
Effects of Weaning Age on Plasma Biomarkers and Growth Performance in Simmental Calves

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Calves Management and Experimental Design
2.2. Body Measurements, Blood Samples, and Analysis
2.3. Statistical Analysis
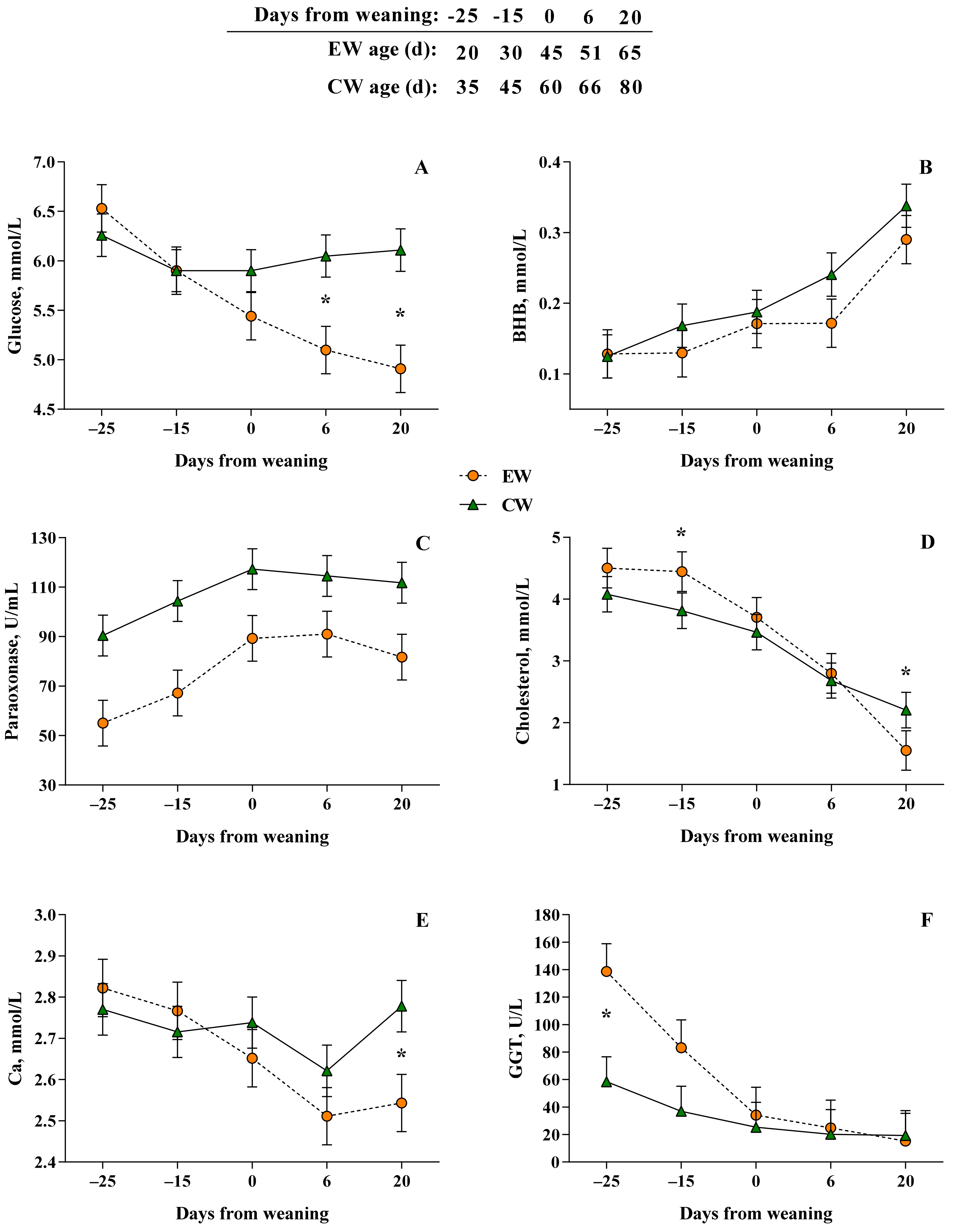
3. Results
3.1. Growth Performance
3.2. Plasma Biomarkers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wickramasinghe, H.K.J.P.; Kaya, C.A.; Baumgard, L.H.; Appuhamy, J.A.D.R.N. Early step-down weaning of dairy calves from a high milk volume with glutamine supplementation. J. Dairy Sci. 2022, 105, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, W.; Liu, T.; Zhang, Q.; Wang, G.; Li, F.; Li, F.; Yue, X.; Li, T. Effect of early weaning on the intestinal microbiota and expression of genes related to barrier function in lambs. Front. Microbiol. 2018, 9, 1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopreiato, V.; Minuti, A.; Cappelli, F.P.; Vailati-Riboni, M.; Britti, D.; Trevisi, E.; Morittu, V.M. Daily rumination pattern recorded by an automatic rumination-monitoring system in pre-weaned calves fed whole bulk milk and ad libitum calf starter. Livest. Sci. 2018, 212, 127–130. [Google Scholar] [CrossRef]
- Mikuš, T.; Marzel, R.; Mikuš, O. Early weaning: New insights on an ever-persistent problem in the dairy industry. J. Dairy Res. 2020, 87, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Morittu, V.M.; Minuti, A.; Spina, A.; Riboni, M.V.; Piccioli-Cappelli, F.; Trevisi, E.; Britti, D.; Lopreiato, V. Age-related metabolic changes of pre-weaned Simmental calves fed whole bulk milk and ad libitum calf starter. Res. Vet. Sci. 2019, 135, 237–243. [Google Scholar] [CrossRef]
- Eckert, E.; Brown, H.E.; Leslie, K.E.; DeVries, T.J.; Steele, M.A. Weaning age affects growth, feed intake, gastrointestinal development, and behavior in Holstein calves fed an elevated plane of nutrition during the preweaning stage. J. Dairy Sci. 2015, 98, 6315–6326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazemi-Bonchenari, M.; Khanaki, H.; Jafari, A.; Eghbali, M.; Poorhamdollah, M.; Ghaffari, M.H. Milk feeding level and starter protein content: Effects on growth performance, blood metabolites, and urinary purine derivatives of Holstein dairy calves. J. Dairy Sci. 2022, 105, 1115–1130. [Google Scholar] [CrossRef] [PubMed]
- Meale, S.J.; Li, S.C.; Azevedo, P.; Derakhshani, H.; DeVries, T.J.; Plaizier, J.C.; Steele, M.A.; Khafipour, E. Weaning age influences the severity of gastrointestinal microbiome shifts in dairy calves. Sci. Rep. 2017, 7, 2699. [Google Scholar] [CrossRef]
- Khan, M.A.; Lee, H.J.; Lee, W.S.; Kim, H.S.; Ki, K.S.; Hur, T.Y.; Suh, G.H.; Kang, S.J.; Choi, Y.J. Structural growth, rumen development, and metabolic and immune responses of holstein male calves fed Milk through step-down and conventional methods. J. Dairy Sci. 2007, 90, 3376–3387. [Google Scholar] [CrossRef] [Green Version]
- Benetton, J.B.; Neave, H.W.; Costa, J.H.C.; von Keyserlingk, M.A.G.; Weary, D.M. Automatic weaning based on individual solid feed intake: Effects on behavior and performance of dairy calves. J. Dairy Sci. 2019, 102, 5475–5491. [Google Scholar] [CrossRef]
- Kertz, A.F.; Hill, T.M.; Quigley, J.D.; Heinrichs, A.J.; Linn, J.G.; Drackley, J.K. A 100-Year Review: Calf nutrition and management. J. Dairy Sci. 2017, 100, 10151–10172. [Google Scholar] [CrossRef]
- Lopreiato, V.; Vailati-Riboni, M.; Morittu, V.M.; Britti, D.; Piccioli-Cappelli, F.; Trevisi, E.; Minuti, A. Post-weaning rumen fermentation of Simmental calves in response to weaning age and relationship with rumination time measured by the Hr-Tag rumination-monitoring system. Livest. Sci. 2020, 232, 103918. [Google Scholar] [CrossRef]
- Palombo, V.; Milanesi, M.; Sgorlon, S.; Capomaccio, S.; Mele, M.; Nicolazzi, E.; Ajmone-Marsan, P.; Pilla, F.; Stefanon, B.; D’Andrea, M. Genome-wide association study of milk fatty acid composition in Italian Simmental and Italian Holstein cows using single nucleotide polymorphism arrays. J. Dairy Sci. 2018, 101, 11004–11019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoni, G.; Trevisi, E.; Han, X.; Bionaz, M. Effects of Inflammatory Conditions on Liver Activity in Puerperium Period and Consequences for Performance in Dairy Cows. J. Dairy Sci. 2008, 91, 3300–3310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferré, N.; Camps, J.; Prats, E.; Vilella, E.; Paul, A.; Figuera, L.; Joven, J. Serum paraoxonase activity: A new additional test for the improved evaluation of chronic liver damage. Clin. Chem. 2002, 48, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Trevisi, E.; Calamari, L.; Librandi, F.; Ferrari, A.; Bertoni, G. Plasma paraoxonase, health, inflammatory conditions, and liver function in transition dairy cows. J. Dairy Sci. 2007, 90, 1740–1750. [Google Scholar] [CrossRef] [Green Version]
- Lopreiato, V.; Minuti, A.; Trimboli, F.; Britti, D.; Morittu, V.M.; Cappelli, F.P.; Loor, J.J.; Trevisi, E. Immunometabolic status and productive performance differences between periparturient Simmental and Holstein dairy cows in response to pegbovigrastim. J. Dairy Sci. 2019, 102, 9312–9327. [Google Scholar] [CrossRef]
- Mirzaei, M.; Khanaki, H.; Kazemi-Bonchenari, M.; Khan, M.A.; Khaltabadi-Farahani, A.H.; Hossein-Yazdi, M.; Ghaffari, M.H. Effects of step-down weaning implementation time on growth performance and blood metabolites of dairy calves. J. Dairy Sci. 2020, 103, 10099–10107. [Google Scholar] [CrossRef]
- Quigley, J.D.; Caldwell, L.A.; Sinks, G.D.; Heitmann, R.N. Changes in Blood Glucose, Nonesterified Fatty Acids, and Ketones in Response to Weaning and Feed Intake in Young Calves. J. Dairy Sci. 1991, 74, 250–257. [Google Scholar] [CrossRef]
- Kertz, A.F.; Prewitt, L.R.; Everett, J.P. An Early Weaning Calf Program: Summarization and Review. J. Dairy Sci. 1979, 62, 1835–1843. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirement of Dairy Cattle, 7th ed.; National Academies Press: Washington, DC, USA, 2001; ISBN 9780309069977. [Google Scholar]
- De Passillé, A.M.; Borderas, T.F.; Rushen, J. Weaning age of calves fed a high milk allowance by automated feeders: Effects on feed, water, and energy intake, behavioral signs of hunger, and weight gains. J. Dairy Sci. 2011, 94, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Bach, A.; Weary, D.M.; von Keyserlingk, M.A.G. Invited review: Transitioning from milk to solid feed in dairy heifers. J. Dairy Sci. 2016, 99, 885–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosadiuk, J.P.; Bruinjé, T.C.; Moslemipur, F.; Fischer-Tlustos, A.J.; Renaud, D.L.; Ambrose, D.J.; Steele, M.A. Differing planes of pre- and postweaning phase nutrition in Holstein heifers: I. Effects on feed intake, growth efficiency, and metabolic and development indicators. J. Dairy Sci. 2021, 104, 1136–1152. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Mena, F.X.; Hu, W.; Dennis, T.S.; Hill, T.M.; Schlotterbeck, R.L. β-Hydroxybutyrate (BHB) and glucose concentrations in the blood of dairy calves as influenced by age, vaccination stress, weaning, and starter intake including evaluation of BHB and glucose markers of starter intake. J. Dairy Sci. 2017, 100, 2614–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, R.L., VI; McLeod, K.R.; Klotz, J.L.; Heitmann, R.N. Rumen development, intestinal growth and hepatic metabolism in the pre- and postweaning ruminant. J. Dairy Sci. 2004, 87, E55–E65. [Google Scholar] [CrossRef] [Green Version]
- Haga, S.; Fujimoto, S.; Yonezawa, T.; Yoshioka, K.; Shingu, H.; Kobayashi, Y.; Takahashi, T.; Otani, Y.; Katoh, K.; Obara, Y. Changes in hepatic key enzymes of dairy calves in early weaning production systems. J. Dairy Sci. 2008, 91, 3156–3164. [Google Scholar] [CrossRef] [Green Version]
- Donkin, S.S.; Armentano, L.E. Insulin and glucagon regulation of gluconeogenesis in preruminating and ruminating bovine. J. Anim. Sci. 1995, 73, 546–551. [Google Scholar] [CrossRef]
- Deelen, S.M.; Leslie, K.E.; Steele, M.A.; Eckert, E.; Brown, H.E.; DeVries, T.J. Validation of a calf-side β-hydroxybutyrate test and its utility for estimation of starter intake in dairy calves around weaning. J. Dairy Sci. 2016, 99, 7624–7633. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Weary, D.M.; von Keyserlingk, M.A.G. Hay intake improves performance and rumen development of calves fed higher quantities of milk. J. Dairy Sci. 2011, 94, 3547–3553. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.; Enjalbert, F.; Combes, S.; Cauquil, L.; Bouchez, O.; Monteils, V. Establishment of ruminal bacterial community in dairy calves from birth to weaning is sequential. J. Appl. Microbiol. 2014, 116, 245–257. [Google Scholar] [CrossRef]
- Puppione, D.L. Implications of Unique Features of Blood Transport in the Lactating Cow. J. Dairy Sci. 1978, 61, 651–659. [Google Scholar] [CrossRef]
- Moody, D.E.; Hohenboken, W.D.; Beal, W.E.; Thye, F.W. Concentration of plasma cholesterol in beef cows and calves, milk production and calf gain. J. Anim. Sci. 1992, 70, 1464–1470. [Google Scholar] [CrossRef] [Green Version]
- Bertoni, G.; Ferrari, A.; Gubbiotti, A.; Trevisi, E. Blood indices calves: Relationship with mother values and changes in the first days of life. Ital. J. Anim. Sci. 2016, 8, 595–597. [Google Scholar] [CrossRef]
- Pérez-Santos, M.; Castillo, C.; Hernández, J.; Abuelo, Á. Biochemical variables from Holstein-Friesian calves older than one week are comparable to those obtained from adult animals of stable metabolic status on the same farm. Vet. Clin. Pathol. 2015, 44, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazana, P.; Siachos, N.; Panousis, N.; Kalaitzakis, E.; Arsenos, G.; Valergakis, G.E. Effect of weaning on serum biochemistry and establishment of reference intervals for peri-weaning period of Holstein calves. Livest. Sci. 2021, 250, 104550. [Google Scholar] [CrossRef]
- Fleck, A. Clinical and nutritional aspects of changes in acute-phase proteins during inflammation. Proc. Nutr. Soc. 1989, 48, 347–354. [Google Scholar] [CrossRef]
- Giordano, A.; Veronesi, M.C.; Rossi, G.; Pezzia, F.; Probo, M.; Giori, L.; Paltrinieri, S. Serum paraoxonase-1 activity in neonatal calves: Age related variations and comparison between healthy and sick animals. Vet. J. 2013, 197, 499–501. [Google Scholar] [CrossRef]
- Turk, R.; Juretic, D.; Geres, D.; Turk, N.; Rekic, B.; Simeon-Rudolf, V.; Svetina, A. Serum paraoxonase activity and lipid parameters in the early postpartum period of dairy cows. Res. Vet. Sci. 2004, 76, 57–61. [Google Scholar] [CrossRef]
- Oetzel, G.R. Management of dry cows for the prevention of milk fever and other mineral disorders. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 369–386. [Google Scholar] [CrossRef]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef] [Green Version]
- Celi, P.; Robinson, A. Effects of Yerba Mate (Ilex paraguariensis) supplementation on the performance of dairy calves. Anim. Prod. Sci. 2010, 50, 376–381. [Google Scholar] [CrossRef]
- Ranade, R.; Talukder, S.; Muscatello, G.; Celi, P. Assessment of oxidative stress biomarkers in exhaled breath condensate and blood of dairy heifer calves from birth to weaning. Vet. J. 2014, 202, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Celi, P. Oxidative stress in ruminants. In Studies on Veterinary Medicine. Oxidative Stress in Applied Basic Research and Clinical Practice; Mandelker, L., Vajdovich, P., Eds.; Humana Press: Totowa, NJ, USA, 2011; pp. 191–231. ISBN 978-1-61779-070-6. [Google Scholar]
- Premi, M.; Mezzetti, M.; Ferronato, G.; Barbato, M.; Cappelli, F.P.; Minuti, A.; Trevisi, E. Changes of plasma analytes reflecting metabolic adaptation to the different stages of the lactation cycle in healthy multiparous holstein dairy cows raised in high-welfare conditions. Animals 2021, 11, 1714. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Item | Bulk Whole Milk 1 | Calf Starter 2 | Grass Hay |
|---|---|---|---|
| N. of samples | 13 | 1 | 1 |
| % on as fed | |||
| Fat | 3.35 | - | - |
| Protein | 3.63 | - | - |
| Lactose | 4.67 | - | - |
| Casein | 2.79 | - | - |
| True protein | 3.43 | - | - |
| Urea, mg/dL | 30.38 | - | - |
| Dry matter (DM), % | 11.65 | 87.87 | 11.13 |
| % on DM | |||
| Starch | - | 24.38 | 3.42 |
| Crude Protein | - | 18.59 | 7.64 |
| Ether Extract | - | 3.05 | 1.38 |
| Neutral Detergent Fiber | - | 33.22 | 65.51 |
| Acid Detergent Fiber | - | 23.18 | 37.66 |
| Acid Detergent Lignin | - | 6.85 | 5.5 |
| Ash | - | 7.34 | 9.78 |
| Item 1 | Pre-Weaning | Weaning | Post-Weaning | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| EW | CW | EW | CW | EW | CW | Wa 3 | Time 4 | Wa × Time 5 | ||
| Glucose, mmol/L | 6.22 | 6.08 | 5.44 | 5.90 | 5.00 | 6.08 | 0.32 | 0.06 | <0.05 | <0.05 |
| BHB, mmol/L | 0.13 | 0.15 | 0.17 | 0.19 | 0.23 | 0.29 | 0.06 | 0.09 | <0.05 | 0.92 |
| NEFA, mmol/L | 0.18 | 0.20 | 0.18 | 0.26 | 0.11 | 0.07 | 0.11 | 0.89 | <0.05 | 0.88 |
| Urea, mmol/L | 3.24 | 3.62 | 3.71 | 4.07 | 4.04 | 4.20 | 0.58 | 0.50 | 0.20 | 0.85 |
| Creatinine, mmol/L | 113.06 | 116.41 | 121.20 | 115.05 | 110.97 | 108.44 | 12.22 | 0.94 | 0.15 | 0.65 |
| Total protein, g/L | 63.40 | 60.78 | 62.30 | 62.16 | 59.40 | 62.07 | 29.25 | 0.99 | 0.85 | 0.71 |
| Item 1 | Pre-Weaning | Weaning | Post-Weaning | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| EW | CW | EW | CW | EW | CW | Wa 3 | Time 4 | Wa × Time 5 | ||
| ROM, mg of H2O2/0.1 L | 15.51 | 14.76 | 13.52 | 11.87 | 11.45 | 11.99 | 2.13 | 0.76 | <0.05 | 0.54 |
| MPO, U/L | 249.73 | 262.69 | 173.16 | 282.48 | 297.68 | 296.05 | 92.61 | 0.59 | 0.63 | 0.83 |
| AOPP, μmol/L | 84.20 | 69.55 | 78.40 | 45.41 | 54.38 | 49.87 | 27.94 | <0.05 | 0.12 | 0.58 |
| FRAP, μmol/L | 134.11 | 124.18 | 141.46 | 146.97 | 155.27 | 134.38 | 14.31 | 0.24 | 0.31 | 0.73 |
| Item 1 | Pre-Weaning | Weaning | Post-Weaning | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| EW | CW | EW | CW | EW | CW | Wa 3 | Time 4 | Wa × Time 5 | ||
| Liver metabolism | ||||||||||
| Albumin, g/L | 33.13 | 33.86 | 34.85 | 34.95 | 33.63 | 34.89 | 1.22 | 0.43 | <0.05 | 0.43 |
| Globulin, g/L | 30.27 | 26.92 | 27.45 | 27.21 | 25.77 | 27.18 | 1.99 | 0.59 | 0.39 | 0.33 |
| Cholesterol, mmol/L | 4.47 | 3.95 | 3.71 | 3.47 | 2.17 | 2.44 | 0.43 | 0.70 | <0.05 | <0.05 |
| Triglycerides, mg/dL | 0.40 | 0.31 | 0.35 | 0.19 | 0.22 | 0.25 | 0.07 | 0.13 | <0.05 | 0.34 |
| PON, U/mL | 61.11 | 97.40 | 89.31 | 117.32 | 86.35 | 113.18 | 12.40 | <0.05 | <0.05 | 0.43 |
| Inflammatory response | ||||||||||
| Haptoglobin, g/L | 0.25 | 0.25 | 0.22 | 0.19 | 0.31 | 0.25 | 0.05 | 0.18 | 0.37 | 0.38 |
| Ceruloplasmin, μmol/L | 1.96 | 1.89 | 1.67 | 1.69 | 1.58 | 1.66 | 0.21 | 0.97 | <0.05 | 0.74 |
| Minerals | ||||||||||
| Ca, mmol/L | 2.79 | 2.74 | 2.65 | 2.74 | 2.53 | 2.70 | 0.11 | 0.32 | <0.05 | <0.05 |
| Mg, mmol/L | 0.86 | 0.88 | 0.96 | 0.96 | 0.90 | 0.98 | 0.05 | 0.23 | <0.05 | 0.51 |
| Zn, μmol/L | 17.87 | 16.63 | 15.64 | 15.16 | 13.47 | 13.16 | 3.30 | 0.16 | 0.54 | 0.94 |
| P, mmol/L | 3.33 | 3.35 | 3.23 | 3.44 | 3.05 | 3.27 | 0.16 | 0.24 | 0.06 | 0.37 |
| Liver Enzymes | ||||||||||
| AST, U/L | 54.18 | 71.47 | 64.82 | 97.27 | 83.37 | 126.42 | 31.90 | 0.11 | 0.41 | 0.90 |
| GGT, U/L | 110.92 | 47.69 | 34.07 | 25.34 | 20.05 | 19.72 | 27.14 | 0.29 | <0.05 | <0.05 |
| ALP, U/L | 421.02 | 535.87 | 370.21 | 413.17 | 310.45 | 417.49 | 104.65 | 0.32 | <0.05 | 0.18 |
| LH, U/L | 1575.71 | 2020.83 | 1805.98 | 2294.98 | 1890.65 | 2626.99 | 451.05 | 0.16 | <0.05 | 0.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferronato, G.; Cattaneo, L.; Trevisi, E.; Liotta, L.; Minuti, A.; Arfuso, F.; Lopreiato, V. Effects of Weaning Age on Plasma Biomarkers and Growth Performance in Simmental Calves. Animals 2022, 12, 1168. https://doi.org/10.3390/ani12091168
Ferronato G, Cattaneo L, Trevisi E, Liotta L, Minuti A, Arfuso F, Lopreiato V. Effects of Weaning Age on Plasma Biomarkers and Growth Performance in Simmental Calves. Animals. 2022; 12(9):1168. https://doi.org/10.3390/ani12091168
Chicago/Turabian StyleFerronato, Giulia, Luca Cattaneo, Erminio Trevisi, Luigi Liotta, Andrea Minuti, Francesca Arfuso, and Vincenzo Lopreiato. 2022. "Effects of Weaning Age on Plasma Biomarkers and Growth Performance in Simmental Calves" Animals 12, no. 9: 1168. https://doi.org/10.3390/ani12091168