
Training in the Dark: Using Target Training for Non-Invasive Application and Validation of Accelerometer Devices for an Endangered Primate (Nycticebus bengalensis)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Data Analysis
3. Results
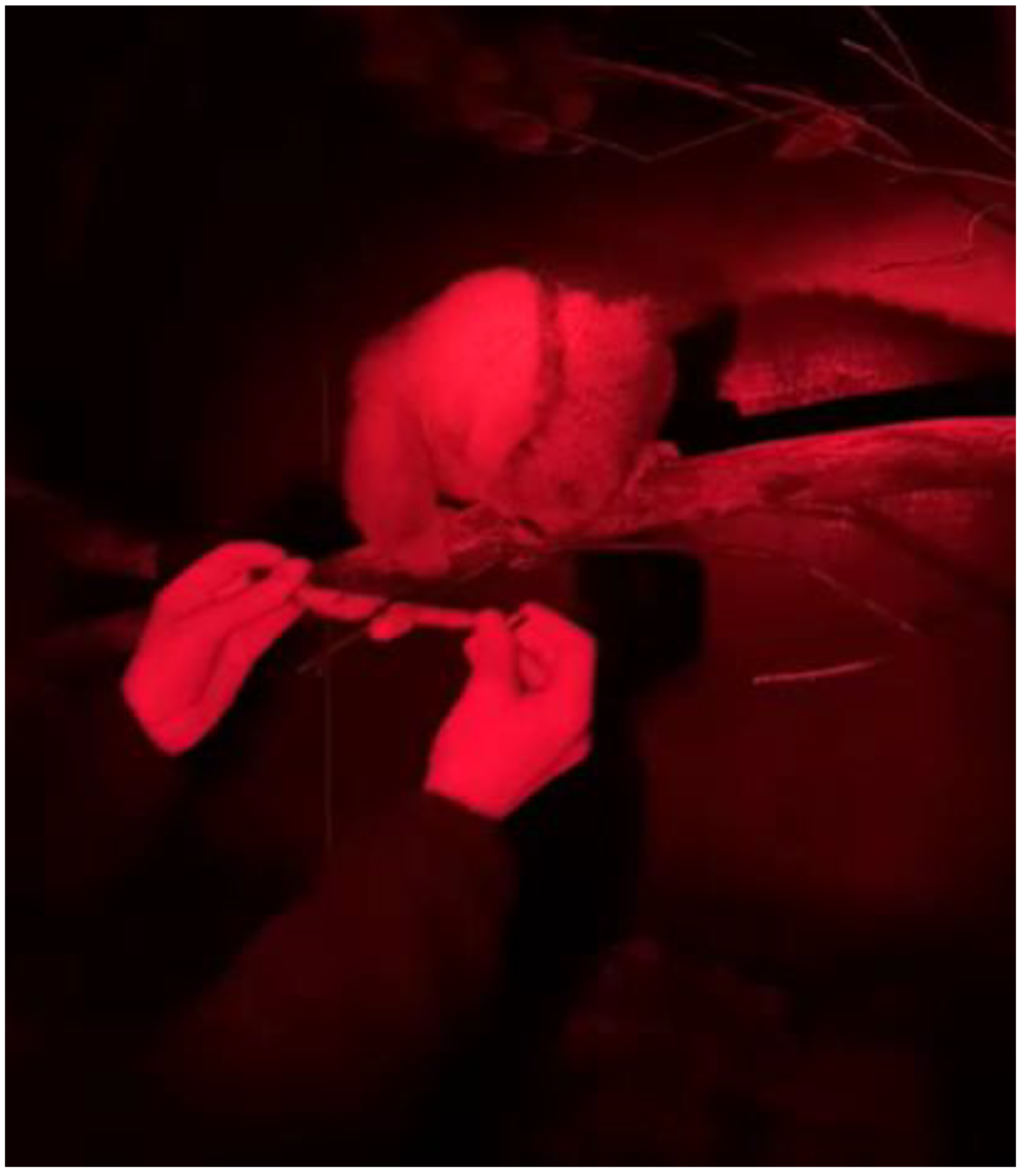
3.1. Training for Application of the Device
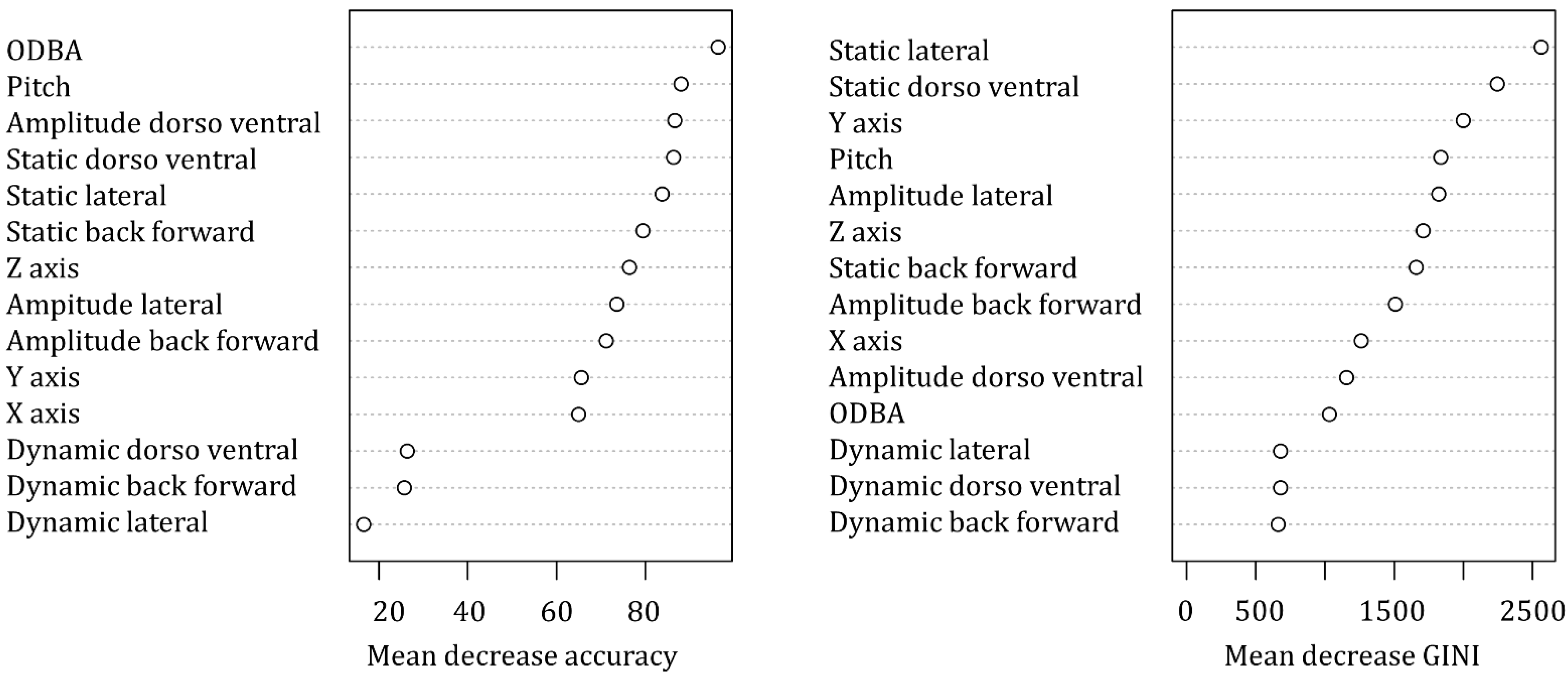
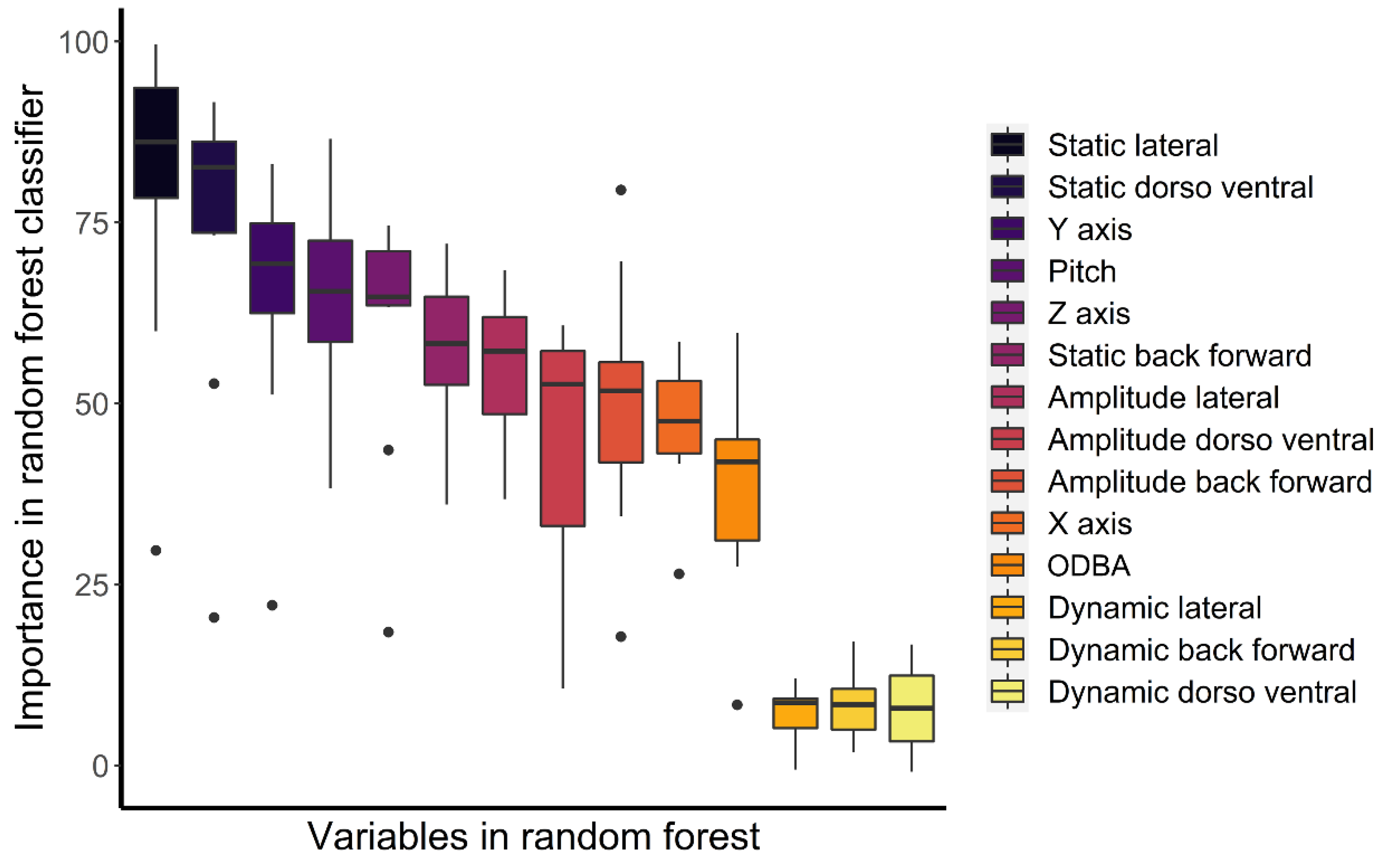
3.2. Accelerometer Validation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, D.D.; Kays, R.; Wikelski, M.; Wilson, R.; Klimley, A. Observing the unwatchable through acceleration logging of animal behavior. Anim. Biotelem. 2013, 1, 20. [Google Scholar] [CrossRef] [Green Version]
- Halsey, L.G.; Shepard, E.L.C.; Wilson, R.P. Assessing the development and application of the accelerometry technique for estimating energy expenditure. Comp. Biochem. Physiol. A 2011, 158, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Furnell, S. A Study of the Locomotor Ecology of the Indriid Primate Propithecus verreauxi in the Dry Deciduous Forest of Kirindy Mitea National Park, Madagascar. Ph.D. Thesis, The University of Manchester, Manchester, UK, 2013. [Google Scholar]
- Campera, M.; Balestri, M.; Chimienti, M.; Nijman, V.; Nekaris, K.A.I.; Donati, G. Temporal niche separation between the two ecologically similar nocturnal primates Avahi meridionalis and Lepilemur fleuretae. Behav. Ecol. Sociobiol. 2019, 73, 55. [Google Scholar] [CrossRef]
- Reinhardt, K.D.; Vyazovskiy, V.V.; Hernandez-Aguilar, R.A.; Imron, M.A.; Nekaris, K.A.I. Environment shapes sleep patterns in a wild nocturnal primate. Sci. Rep. 2019, 9, 9939. [Google Scholar]
- Ladds, M.A.; Salton, M.; Hocking, D.P.; McIntosh, R.R.; Thompson, A.P.; Slip, D.J.; Harcourt, R.G. Using accelerometers to develop time-energy budgets of wild fur seals from captive surrogates. PeerJ 2018, 6, e5814. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, R.E.; Tongen, A.; Gardiner, J.; Miller, C.E.; Schmitt, D. Dynamics of locomotor transitions from arboreal to terrestrial substrates in Verreaux’s sifaka (Propithecus verreauxi). Integr. Comp. Biol. 2014, 54, 1148–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, B.F. Operant behavior. In Operant Behavior: Areas of Research and Application; Honig, W.K., Ed.; Appleton-Century-Crofts: New York, NY, USA, 1966; pp. 12–32. [Google Scholar]
- Prescott, M.J.; Buchanan-Smith, H.M. Training nonhuman primates using positive reinforcement techniques. J. Appl. Anim. Welf. Sci. 2003, 6, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Laule, G.E.; Bloomsmith, M.A.; Schapiro, S.J. The use of positive reinforcement training techniques to enhance the care management, and welfare of primates in the laboratory. J. Appl. Anim. Welf. Sci. 2003, 6, 163–173. [Google Scholar] [CrossRef]
- Melfi, V. Is training zoo animals enriching? Appl. Anim. Behav. Sci. 2013, 147, 299–305. [Google Scholar] [CrossRef]
- Fuller, G.; Lukas, K.E.; Kuhar, C.; Dennis, P.M. A retrospective review of mortality in lorises and pottos in North American zoos, 1980–2010. Endanger. Species Res. 2014, 23, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Rode-Margono, E.J.; Nijman, V.; Wirdateti, W.; Nekaris, K.A.I. Ethology of the Critically Endangered Javan slow loris Nycticebus javanicus E. Geoffroy Saint-Hilaire in West Java. Asian Primates 2014, 4, 27–41. [Google Scholar]
- Poindexter, S.; Nekaris, K.A.I. Vertical clingers and gougers: Rapid acquisition of adult limb proportions facilitates feeding behaviours in young Javan slow lorises (Nycticebus javanicus). Mamm. Biol. 2017, 87, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Hanna, J.B.; Schmitt, D. Locomotor energetics in primates: Gait mechanics and their relationship to the energetics of vertical and horizontal locomotion. Am. J. Phys. Anthropol. 2011, 145, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, N.J. The effect of branch diameter on primate gait sequence pattern. Am. J. Primatol. 2008, 70, 356–362. [Google Scholar] [CrossRef]
- Wiens, F.; Zitzmann, A. Social structure of the solitary slow loris Nycticebus coucang (Lorisidae). J. Zool. 2003, 261, 35–46. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Campera, M.; Nijman, V.; Birot, H.; Rode-Margono, E.J.; Fry, B.G.; Imron, M.A. Slow lorises use venom as a weapon in intraspecific competition. Curr. Biol. 2020, 30, R1252–R1253. [Google Scholar] [CrossRef]
- Chimienti, M.; Cornulier, T.; Owen, E.; Bolton, M.; Davies, I.M.; Travis, J.M.J.; Scott, B.E. The use of an unsupervised learning approach for characterizing latent behaviors in accelerometer data. Ecol. Evol. 2016, 6, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L.; Cutler, A. Package ‘randomForest’. 2018. Available online: https://www.stat.berkeley.edu/~breiman/RandomForests/ (accessed on 3 December 2021).
- Dickinson, E.R.; Twining, J.P.; Wilson, R.; Stephens, P.A.; Westander, J.; Marks, N.; Scantlebury, D.M. Limitations of using surrogates for behaviour classification of accelerometer data: Refining methods using random forest models in Caprids. Mov. Ecol. 2021, 9, 28. [Google Scholar] [CrossRef]
- Fuller, G.; Raghanti, M.A.; Dennis, P.M.; Kuhar, C.W.; Willis, M.A.; Schook, M.W.; Lukas, K.E. A comparison of nocturnal primate behavior in exhibits illuminated with red and blue light. Appl. Anim. Behav. Sci. 2016, 184, 126–134. [Google Scholar] [CrossRef]
- Spiezio, C.; Vaglio, S.; Scala, C.; Regaiolli, B. Does positive reinforcement training affect the behavior and welfare of zoo animals? The case of the ring-tailed lemur (Lemur catta). Appl. Anim. Behav. Sci. 2017, 196, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Schapiro, S.J.; Bloomsmith, M.; Laule, G.E. Positive reinforcement training as a technique to alter nonhuman primate behavior: Quantitative assessments of effectiveness. J. Appl. Anim. Welf. Sci. 2003, 6, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Madeiros, A. Training of North American Porcupine (Erethizon dorsatum) to Allow for Subcutaneous Injections Using Positive Reinforcement; Statislaus State University Honors Program: Turlock, CA, USA, 2018. [Google Scholar]
- Pomerantz, O.; Terkel, J. Effects of positive reinforcement training techniques on the psychological welfare of zoo-housed chimpanzees (Pan troglodytes). Am. J. Primatol. 2009, 71, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Buchanan-Smith, H.M. The benefits of positive reinforcement training and its effects on human non-human animal interactions. In Proceedings of the Fifth Annual Symposium on Zoo Research, Winchester, UK, 7–8 July 2003; pp. 21–26. [Google Scholar]
- Gardiner, M.; Weldon, A.; Poindexter, S.A.; Gibson, N.; Nekaris, K.A.-I. Survey of practitioners handling slow lorises (Primates: Nycticebus): An assessment of the harmful effects of slow loris bites. J. Venom Res. 2018, 9, 1. [Google Scholar] [PubMed]
- Waitt, C.; Buchanan-Smith, H.M.; Morris, K. The effects of caretaker–primate relationship on primates in the laboratory. J. Appl. Anim. Welf. Sci. 2002, 5, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Allan, B.M.; Nimmo, D.G.; Arnould, J.P.; Martin, J.K.; Ritchie, E.G. The secret life of possums: Data loggers reveal the movement ecology of an arboreal mammal. J. Mammal. 2019, 100, 158–168. [Google Scholar] [CrossRef]
- Byrnes, G.; Lim, N.T.L.; Spence, A.J. Take-off and landing kinetics of a free-ranging gliding mammal, the Malayan colugo (Galeopterus variegatus). Proc. Royal Soc. B 2008, 275, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Warren, R.D.; Crompton, R.H. Locomotor ecology of Lepilemur edwardsi and Avahi occidentalis. Am. J. Phys. Anthropol. 1997, 104, 471–486. [Google Scholar] [CrossRef]
- Fehlmann, G.; O’Riain, M.J.; Hopkins, P.W.; O’Sullivan, J.; Holton, M.D.; Shepard, E.L.C.; King, A.J. Identification of behaviours from accelerometer data in a wild social primate. Anim. Biotelemetry 2017, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Nickel, B.; Rutishauser, M.; Bryce, C.M.; Williams, T.M.; Elkaim, G.; Wilmers, C.C. Movement, resting, and attack behaviors of wild pumas are revealed by tri-axial accelerometer measurements. Mov. Ecol. 2015, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Tatler, J.; Cassey, P.; Prowse, T.A.A. High accuracy at low frequency: Detailed behavioural classification from accelerometer data. J. Exp. Biol. 2018, 221, jeb184085. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, G. Casting the net widely for change in animal welfare: The plight of birds in zoos, ex situ conservation, and conservation fieldwork. Animals 2022, 12, 31. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | No. Training Days | Description |
|---|---|---|
| 1 | --- | As part of regular daily management, feed insects from tongs with the bridge word “good”. |
| 2 | 5 | Presenting target stick with insect reward. |
| 3 | 4 | Accepting placement of large bamboo collar over the head from the target stick. |
| 4 | 12 | Learning to push head through quick release cat collar from the target stick. |
| 5 | 2 | Keeper places cat collar over loris’ head with the target stick. |
| 6 | 3 | Introducing the touch command to remove the collar with accelerometer with the tongs. |
| 7 | 12 | Keeper touch with no collar on to become used to being touched by hands. |
| 8 | 2 | Clipping the collar on and off with the hands. |
| Behaviour | Locomotion/Posture | Description |
|---|---|---|
| Alert | Sit/stand | Remain stationary such as in “rest” but active scanning of environment. |
| Explore | Movement associated with looking for food (often includes visual and olfactory searching) or exploring the habitat. | |
| Bridge | Exploring while climbing from one support to the next, (trunk or branches of same or different trees), stretching over a gap of more than 15 cm. | |
| Climb down | Exploring while moving downwards on +/−45° to 90° support. | |
| Climb horizontally | Exploring while moving horizontally through +/−90° or +/−45° support. | |
| Climb up | Exploring while moving upwards on +/−45° to 90° support. | |
| Suspensory walk | Exploring while hanging on +/−0° to 45° support. | |
| Walk | Exploring while walking quadrupedally on +/−0° to 45° support. | |
| Feeding | Actual consumption of a food item. | |
| Non-suspensory | Feeding in a stationary position (e.g., sit or stand). | |
| Suspensory | Feeding in a suspensory position while hanging from a branch. | |
| Resting | Sit/stand | Remain stationary, often with body hunched |
| Behaviour | Prediction Accuracy (%) | Main Confusing Behaviour (% error) | Importance in Random Forest Classifier | ||
|---|---|---|---|---|---|
| 1st Variable | 2nd Variable | 3rd Variable | |||
| Resting | 99.8 | Suspensory walk (0.2) | Amplitude lateral | Pitch | Static back forward |
| Bridge | 85.9 | Suspensory walk (4.3) | Pitch | Static lateral | Static dorso ventral |
| Suspensory feeding | 85.6 | Suspensory walk (5.1) | Static lateral | Static dorso ventral | Y axis |
| Climb down | 82.5 | Walk (8.7) | Static lateral | Static dorso ventral | Y axis |
| Climb up | 80.7 | Walk (9.6) | Static lateral | Static dorso ventral | Y axis |
| Alert | 80.4 | Walk (9.3) | Static lateral | Static dorso ventral | Amplitude back forward |
| Climb horizontally | 79.8 | Suspensory walk (15.5) | Static lateral | Static dorso ventral | Amplitude back forward |
| Walk | 77.2 | Alert (9.6) | Static lateral | Pitch | Static dorso ventral |
| Feeding non-suspensory | 75.0 | Suspensory feeding (9.0) | Static lateral | Static dorso ventral | Pitch |
| Suspensory walk | 60.3 | Feeding suspension (18.8) | Static lateral | Static dorso ventral | Z axis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nekaris, K.A.-I.; Campera, M.; Chimienti, M.; Murray, C.; Balestri, M.; Showell, Z. Training in the Dark: Using Target Training for Non-Invasive Application and Validation of Accelerometer Devices for an Endangered Primate (Nycticebus bengalensis). Animals 2022, 12, 411. https://doi.org/10.3390/ani12040411
Nekaris KA-I, Campera M, Chimienti M, Murray C, Balestri M, Showell Z. Training in the Dark: Using Target Training for Non-Invasive Application and Validation of Accelerometer Devices for an Endangered Primate (Nycticebus bengalensis). Animals. 2022; 12(4):411. https://doi.org/10.3390/ani12040411
Chicago/Turabian StyleNekaris, K. Anne-Isola, Marco Campera, Marianna Chimienti, Carly Murray, Michela Balestri, and Zak Showell. 2022. "Training in the Dark: Using Target Training for Non-Invasive Application and Validation of Accelerometer Devices for an Endangered Primate (Nycticebus bengalensis)" Animals 12, no. 4: 411. https://doi.org/10.3390/ani12040411