
Initial Evidence That Gilthead Seabream (Sparus aurata L.) Is a Host for Lymphocystis Disease Virus Genotype I

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Sampling
2.2. Histopathology
2.3. Electron Microscopy
2.4. Molecular Analysis
3. Results
3.1. Clinical Observations
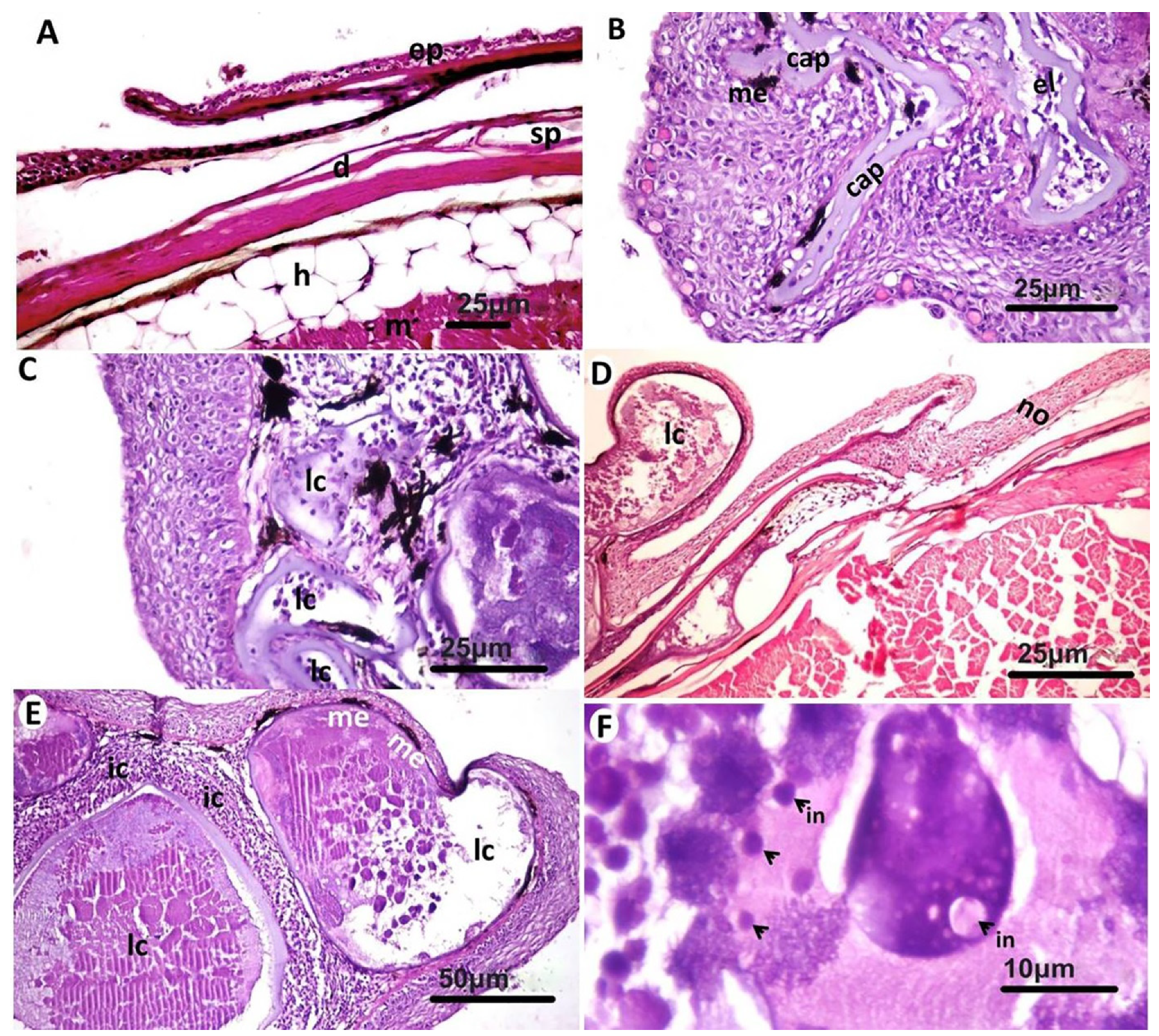
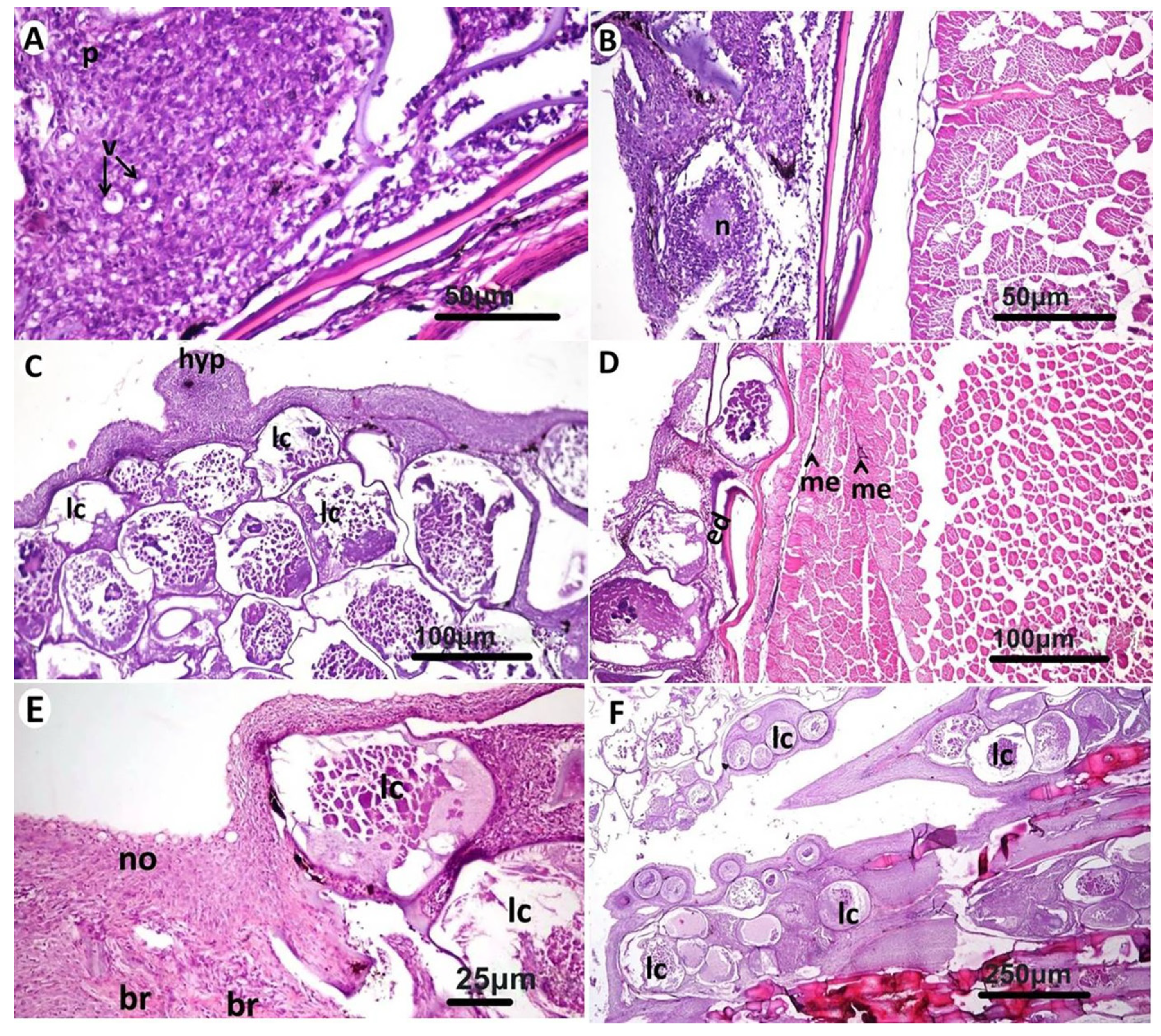
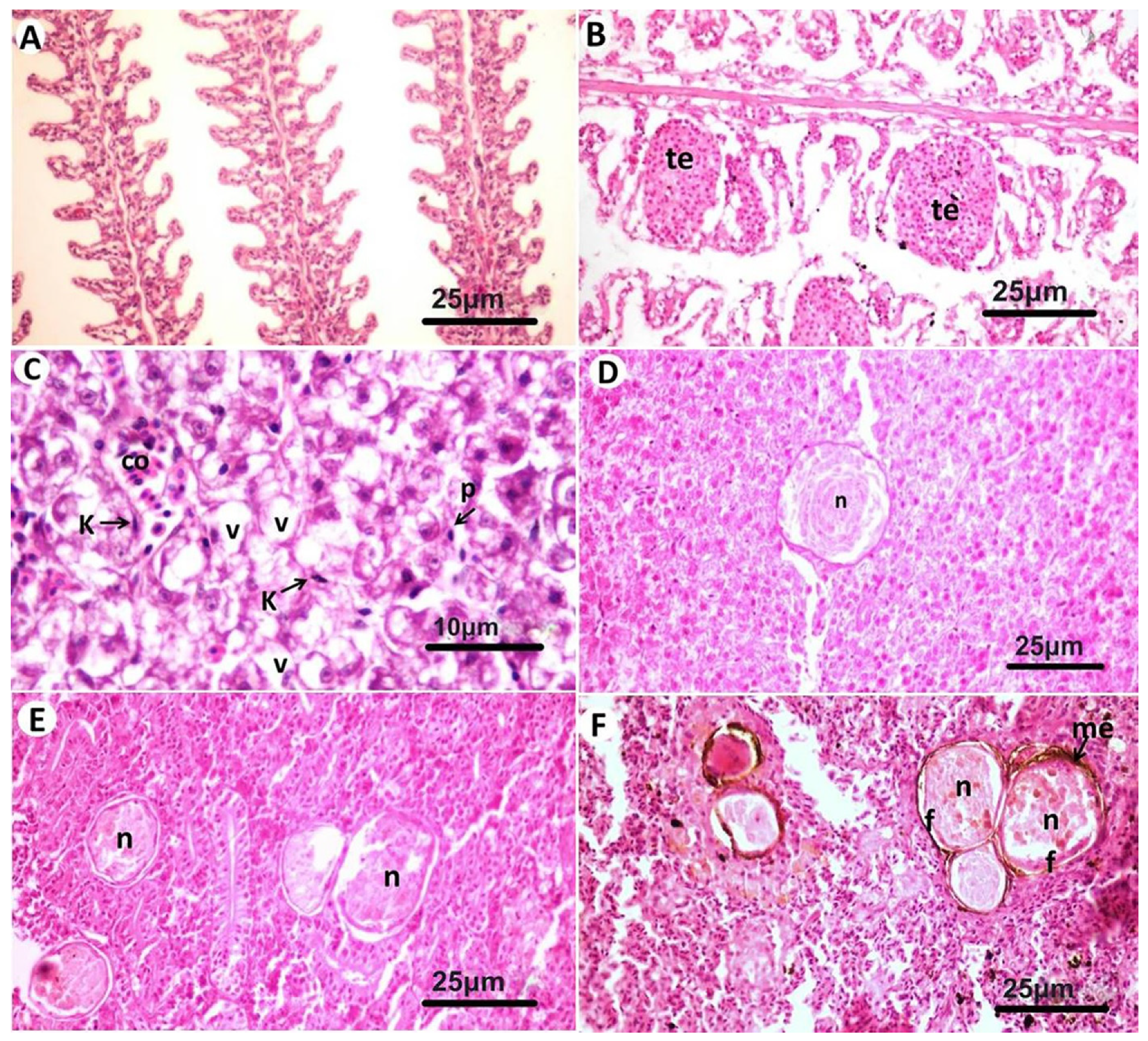
3.2. Histopathological Findings
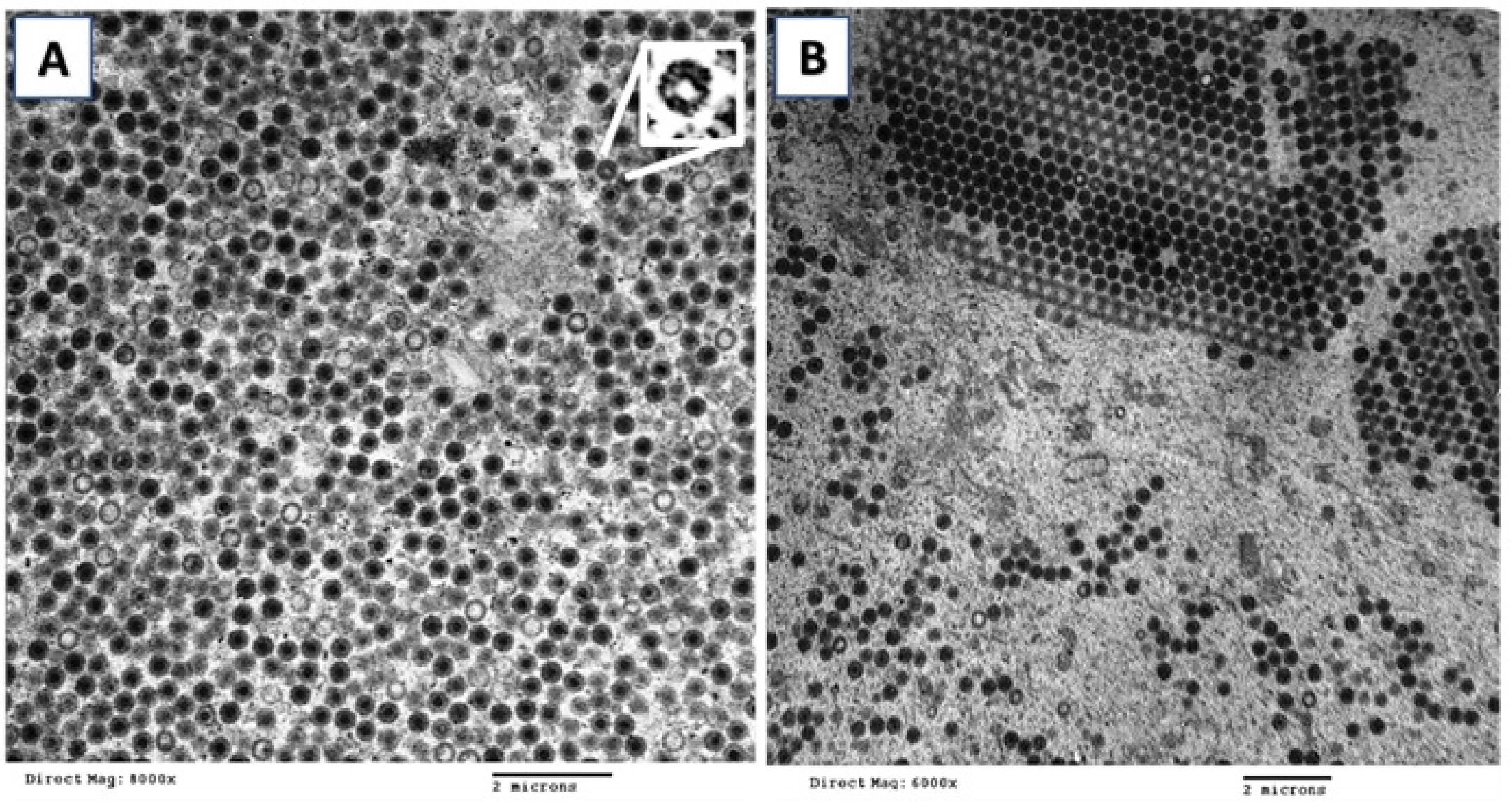
3.3. Electron Microscopy
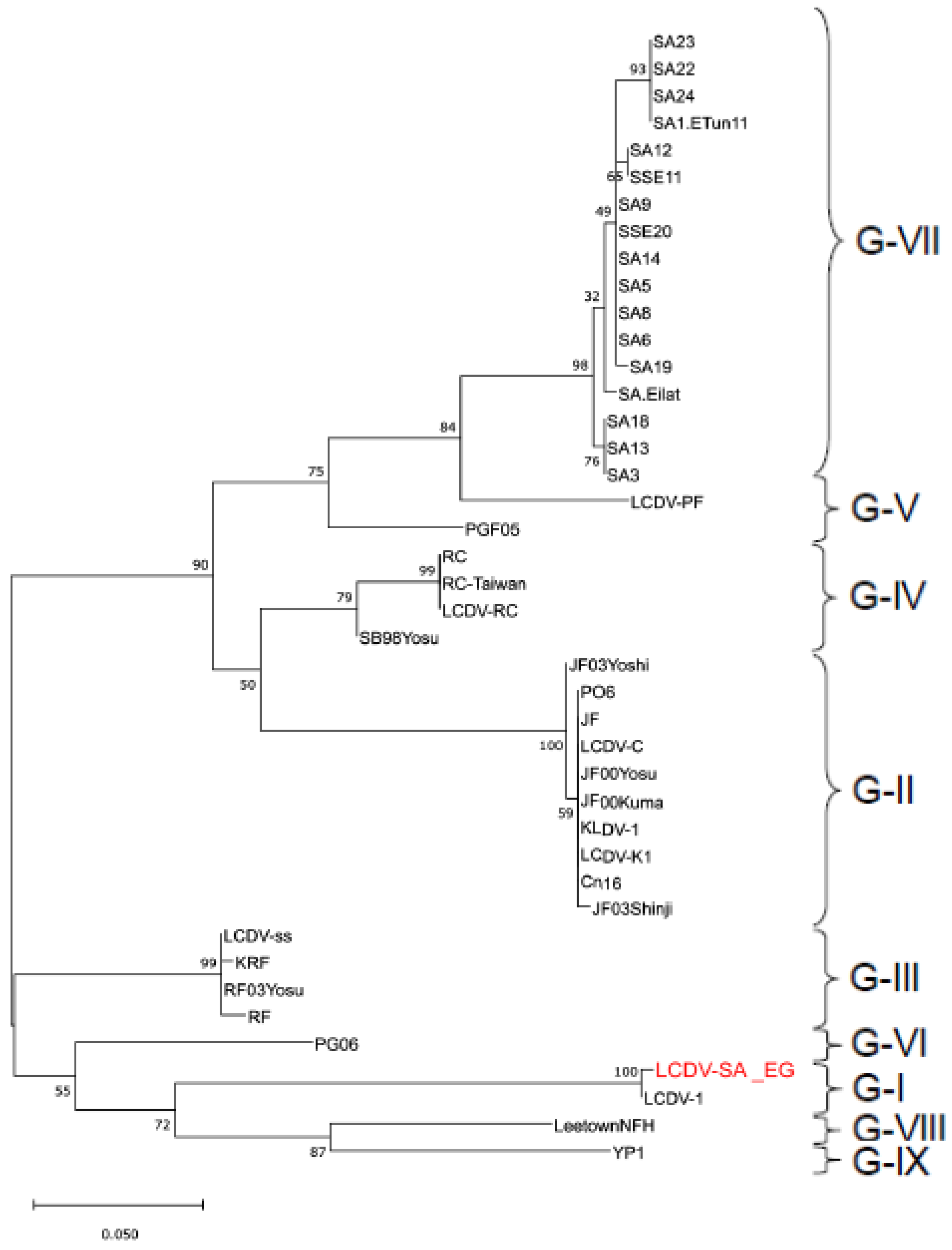
3.4. Gene Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Soliman, N.F.; Yacout, D.M.M. Aquaculture in Egypt: Status, constraints and potentials. Aquac. Int. 2016, 24, 1201–1227. [Google Scholar] [CrossRef]
- Da Silva, J.G. The state of world fisheries and aquaculture 2016. In Contributing to Food Security and Nutrition for All; Food and Agriculture Organization of the United Nations Press: Rome, Italy, 2016; pp. 1–204. [Google Scholar]
- FAO. Rome: Sparus aurata (Linnaeus, 1758). Cultured Aquatic Species Information Programme. FAO Fisheries and Aquaculture Department. Available online: http://www.fao.org/fishery/culturedspecies/Sparus_aurata/en (accessed on 9 November 2019).
- Faisal, M. Health challenges to aquatic animals in the globalization era. In Globalization: Effects on Fisheries Resources; William, W., Taylor, M.G., Schechter, L.G.W., Eds.; Cambridge University Press: New York, NY, USA, 2007; pp. 120–155. [Google Scholar]
- Bovo, G.; Giorgetti, G.; Jørgensen, P.E.V.; Olesen, N.J. Infectious haematopoietic necrosis: First detection in Italy. Bull. Eur. Assoc. Fish Pathol. 1987, 7, 124. [Google Scholar]
- Haenen, O.L.M.; Way, K.; Bergmann, S.M.; Ariel, E. The emergence of koi herpesvirus and its significance to European aquaculture. Bull. Eur. Assoc. Fish Pathol. 2004, 24, 293–307. [Google Scholar]
- Jones, J.B.; Hyatt, A.D.; Hine, P.M.; Whittington, R.J.; Griffin, D.A.; Bax, N.J. Australasian pilchard mortalities. World J. Microbiol. Biotechnol. 1997, 13, 383–392. [Google Scholar] [CrossRef]
- Graham, D.A.; Rowley, H.M.; Walker, I.W.; Weston, J.H.; Branson, E.J.; Todd, D. First isolation of sleeping disease virus from rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2003, 26, 691–694. [Google Scholar] [CrossRef] [PubMed]
- OIE. OIE-Listed Diseases, Infections and Infestations in Force in 2016, World Organization for Animal Health. Available online: http://www.oie.int/animal-health-in-the-world/oie-listed-diseases-2016/ (accessed on 19 January 2016).
- Chinchar, V.G.; Hick, P.; Ince, I.A.; Jancovich, J.K.; Marschang, R.; Qin, Q.; Subramaniam, K.; Waltzek, T.B.; Whittington, R.; Williams, T.; et al. ICTV Virus Taxonomy Profile: Iridoviridae. J. Gen. Virol. 2017, 98, 890–891. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K. Lymphocystis disease. In Fish Viruses and Fish Viral Diseases; Wolf, K., Ed.; Cornell University Press: Ithaca, NY, USA, 1988; pp. 268–291. [Google Scholar]
- Cano, I.; Ferro, P.; Alonso, M.C.; Bergmann, S.M.; Römer-Oberdörfer, A.; Garcia-Rosado, E.; Castro, D.; Borrego, J.J. Development of molecular techniques for detection of lymphocystis disease virus in different marine fish species. J. Appl. Microbiol. 2007, 102, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Cano, I.; Valverde, E.J.; Lopez-Jimena, B.; Alonso, M.C.; Garcia-Rosado, E.; Sarasquete, C.; Borrego, J.J.; Castro, D. A new genotype of Lymphocystivirus isolated from cultured gilthead seabream, Sparus aurata L., and Senegalese sole, Solea senegalensis (Kaup). J. Fish Dis. 2010, 33, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.J.; Hogan, N.S.; Van den Heuvel, M.R. Phylogenetic analysis and molecular methods for the detection of lymphocystis disease virus from yellow perch, Perca flavescens (Mitchell). J. Fish Dis. 2012, 35, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.; Smail, D.A.; Munro, E.S. Laboratory Methodes. In Fish Pathology, 4th ed.; Roberts, R., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 439–481. [Google Scholar]
- Mokhtar, D. Fish Histology From Cells to Organs. In Fish Histology; Mokhtar, D., Ed.; Apple Academic Press: New York, NY, USA, 2017; pp. 1–264. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Michael, L.; Christina, K.; Koichiro, T. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M. Modulation of Cell-Mediated Immune Responses by Lymphocystis Disease in the Gilthead Sea Bream Sparus aurata. In Viruses of Lower Vertebrates; Ahne, W., Kurstak, E., Eds.; Springer: Berlin, Germany, 1989; pp. 487–496. [Google Scholar]
- Faisal, M. Lymphocystis in the Mediterranean golden grouper Epinephelus alexandrinus Valenciennes 1828 (Pisces Serranidae). Bull. Eur. Assoc. Fish Pathol. 1989, 9, 17–18. [Google Scholar]
- Haddad-Boubaker, S.; Bouzgarou, N.; Fakhfakh, E.; Khayech, M.; Mohamed, S.B.; Megdich, A.; Chéhida, N.B. Detection and Genetic Characterization of Lymphocystis Disease Virus (LCDV) Isolated during Disease Outbreaks in Cultured Gilt-head Seabream Sparus aurata in Tunisia. Fish Pathol. 2013, 48, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Menezes, J.; Ramos, M.A.; Pereira, T.G. Lymphocystis disease: An outbreak in Sparus aurata from Ria Formosa, south coast of Portugal. Aquaculture 1987, 67, 222–225. [Google Scholar] [CrossRef]
- Bernardo, B.; Marcotegui, M.A.; Rueda, A.; Tiana, A.; Castellanos, A.; Tarazona, J.; Muñoz, M.J.; Coll, J.M. First report on lymphocystis disease in Sparus aurata (Linnaeus) in Spain. Bull. Eur. Assoc. Fish Pathol. 1990, 10, 71–73. [Google Scholar]
- Le Deuf, R.M.; Renault, T. Lymphocystis outbreaks in farmed seabream Sparus aurata, first report on French Mediterranean coast. Bull. Eur. Assoc. Fish Pathol. 1993, 13, 130–133. [Google Scholar]
- Moate, R.M.; Harris, J.E.; McMahon, S. Lymphocystis infections in cultured gilt-head seabream Sparus aurata in the Aegean Sea. Bull. Eur. Assoc. Fish Pathol. 1992, 12, 134. [Google Scholar]
- Paperna, I.; Sabnai, H.I.; Colorni, A. An outbreak of lymphocystis in Sparus aurata L. in the Gulf of Aqaba, Red Sea. J. Fish Dis. 1982, 5, 433–437. [Google Scholar] [CrossRef]
- Tidona, C.; Schnitzler, P.; Kehm, R.; Darai, G. Is the Major Capsid Protein of Iridoviruses a Suitable Target for the Study of Viral Evolution? Virus Genes 1998, 16, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.I.; Jung, S.J.; Kim, W.S.; Nishizawa, T.; Yoshimizu, M.; Oh, M.J. A new genotype of lymphocystivirus, LCDV-RF, from lymphocystis diseased rockfish. Arch. Virol. 2006, 151, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.C.; Ferro, P.; Garcia-Rosado, E.; Cano, I.; Lang, T.; Bergmann, S.; Borrego, J.J. Comparison of Lymphocystis Disease Virus (LCDV) isolates obtained from different marine fish species and geographical areas. Bull. Eur. Assoc. Fish Pathol. 2007, 27, 157–164. [Google Scholar]
- Valverde, E.J.; Cano, I.; Alejandro, L.; Borrego, J.J.; Dolores, C. Application of a new real-time polymerase chain reaction assay for surveillance studies of lymphocystis disease virus in farmed gilthead seabream. BMC Vet. Res. 2016, 12, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreck, C.B.; Tort, L. The Concept of Stress in Fish. In Fish Physiology; Schreck, C.B., Tort, L., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: London, UK, 2016; Volume 35, pp. 1–34. [Google Scholar]
- Yan, X.Y.; Wu, Z.H.; Chang, J.J.; Lu, Y.S.; Sun, X.Q. Analysis of the genetic diversity of the lymphocystis virus and its evolutionary relationship with its hosts. Virus Genes 2011, 43, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Valverde, E.J.; Labella, A.M.; Borrego, J.J.; Castro, D. Artemia spp., a susceptible host and vector for lymphocystis disease virus. Viruses 2019, 11, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano, I.; Lopez-Jimena, B.; Garcia-Rosado, E.; Ortiz-Delgado, J.; Alonso, M.C.; Borrego, J.; Sarasquete, C.; Castro, D. Detection and persistence of Lymphocystis disease virus (LCDV) in Artemia sp. Aquaculture 2009, 291, 230–236. [Google Scholar]
- Cano, I.; Ferro, P.; Alonso, M.C.; Sarasquete, C.; Garcia-Rosado, E.; Borrego, J.J.; Castro, D. Application of in situ detection techniques to determine the systemic condition of lymphocystis disease virus infection in cultured gilt-head seabream, Sparus aurata L. J. Fish Dis. 2009, 32, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Valverde, E.J.; Borrego, J.J.; Sarasquete, M.C.; Ortiz-Delgado, J.B.; Castro, D. Target organs for lymphocystis disease virus replication in gilthead seabream (Sparus aurata). Vet. Res. 2017, 48, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.; Oh, M.J. Histopathology of marine and freshwater fish lymphocystis disease virus (LCDV). Sains Malays. 2011, 40, 1049–1052. [Google Scholar]
- Cano, I.; Valverde, E.J.; Garcia-Rosado, E.; Alonso, M.C.; Lopez-Jimena, B.; Ortiz-Delgado, J.B.; Borrego, J.J.; Sarasquete, C.; Castro, D. Transmission of lymphocystis disease virus to cultured gilthead seabream, Sparus aurata L. larvae. J. Fish Dis. 2013, 36, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Acc | Abbreviation | Genotype | Fish Species | Year | Origin |
|---|---|---|---|---|---|
| MN128712 | LCDV-SA_EG | G-I | Gilthead seabream (Sparus aurata) | 2017 | (This study) |
| L63545.1 | LCDV-1 | European flounder (Platichthys flesus) Plaice (Pleuronectes platessa L) | 1997 | Europe | |
| GU939626.2 | YP-1 | G-IX | Yellow perch (Perca flavescens) | 2009 | Canada |
| GU290550.1 | Leetown NFH | G-VIII | Largemouth bass (Micropterus salmoides) | 1962 | USA |
| AB299164.1 | PG06 | G-VI | Pearl gourami (Trichopodus leerii) | 2008 | Korea |
| AY849392.1 | KRF | G-III | Korean rock fish (Sebastes schlegeli) | 2004 | Korea |
| AB213004.1 | RF03Yosu | S. schlegeli | 2003 | Korea | |
| KT438164.1 | LCDV-ss | S. schlegeli | 2012 | China | |
| AY823414.1 | RF | S. schlegeli | 2004 | Korea | |
| AB213000.1 | JF03Shinji | G-II | Japanese flounder (P. olivaceus) | 2003 | Korea |
| AF126405.1 | Cn16 | P. olivaceus | 1999 | China | |
| AY303804.1 | LCDV-K1 | P. olivaceus | 2003 | Korea | |
| AY297741.1 | KLDV-1 | P. olivaceus | 2003 | Korea | |
| AB212997.1 | JF00Kuma | P. olivaceus | 2000 | Japan | |
| AB212999.1 | JF00Yosu | P. olivaceus | 2000 | Korea | |
| AY380826.1 | LCDV-C | P. olivaceus | 2003 | China | |
| AY849391.1 | JF | P. olivaceus | 2004 | Korea | |
| KP184512.1 | PO6 | P. olivaceus | 2012 | Korea | |
| AB212998.1 | JF03Yoshi | P. olivaceus | 2003 | Japan | |
| AB247938.1 | SB98Yosu | G-IV | Seabass (Lateolabrax japonicus) | 2006 | Korea |
| EF059992.1 | RC | Cobia (Rachycentron canadum) | 2006 | China | |
| EF103188.1 | LCDV-RC | R. canadum | 2006 | China | |
| EF378607.1 | RC-Taiwan | R. canadum | 2007 | Taiwan | |
| AB299163.1 | PGF05 | G-V | Painted glass fish (Parambassis ranga) | 2007 | Japan |
| KJ408271.1 | LCDV-PF | Paradise fish (Macropodus opercularis) | 2014 | China | |
| GU320724.1 | SA3 | G-VII | S. aurata | 1997 | Spain |
| GU320731.1 | SA13 | S. aurata | 2005 | Tunisia | |
| GU320734.1 | SA18 | S. aurata | 2008 | Portugal | |
| EF184306.1 | SA-Eilat | S. aurata | 2006 | Israel | |
| GU320735.1 | SA19 | S. aurata | 2008 | Spain | |
| GU320729.1 | SSE11 | Senegalese sole (Solea senegalensis) | 2001 | Spain | |
| GU320730.1 | SA12 | S. aurata | 2003 | Spain | |
| GU320726.1 | SA6 | S. aurata | 1998 | Spain | |
| GU320727.1 | SA8 | S. aurata | 2000 | Spain | |
| GU320725.1 | SA5 | S. aurata | 1998 | Spain | |
| KX643370.1 | SA9 | S. aurata | 2001 | Spain | |
| GU320736.1 | SSE20 | S. senegalensis | 2008 | Spain | |
| GU320732.1 | SA14 | S. aurata | 2008 | Spain | |
| HE650105.1 | SA1-ETun11 | S. aurata | 2011 | Tunisia | |
| GU320739.1 | SA24 | S. aurata | 2009 | France | |
| GU320737.1 | SA22 | S. aurata | 2009 | France | |
| GU320738.1 | SA23 | S. aurata | 2009 | France |
| Percent Identity | |||||||||||
| LCDV-SA_EG | (LCDV-1) G-I | (YP1) G-IX | (Leetown NFH) G-VIII | (PG06) G-VI | (RF) G-III | (JF03Yoshi) G-II | (LCDV-RC) G-IV | (SA-Eilat) G-VII | (PGF05) G-V | ||
| Divergence | LCDV-SA_EG | 100 | 89.6 | 90.6 | 89.6 | 88.5 | 86.5 | 89.6 | 88.5 | 88.5 | |
| (LCDV-1) G-I | 0.0 | 89.6 | 90.6 | 89.6 | 88.5 | 86.5 | 89.6 | 88.5 | 88.5 | ||
| (YP1) G-IX | 11.2 | 11.2 | 91.7 | 91.7 | 91.7 | 89.6 | 91.7 | 90.6 | 91.7 | ||
| (Leetown NFH) G-VIII | 10.0 | 10.0 | 8.9 | 93.8 | 93.8 | 93.8 | 93.8 | 92.7 | 92.7 | ||
| (PG06) G-VI | 11.2 | 11.2 | 8.9 | 6.5 | 93.8 | 92.7 | 91.7 | 91.7 | 90.6 | ||
| (RF) G-III | 12.5 | 12.5 | 8.9 | 6.5 | 6.5 | 94.8 | 95.8 | 95.8 | 94.8 | ||
| (JF03Yoshi) G-II | 15.0 | 15.0 | 11.2 | 6.5 | 7.7 | 5.4 | 95.8 | 95.8 | 94.8 | ||
| (LCDV-RC) G-IV | 11.2 | 11.2 | 8.9 | 6.5 | 8.9 | 4.3 | 4.3 | 97.9 | 97.9 | ||
| (SA-Eilat) G-VII | 12.5 | 12.5 | 10.0 | 7.7 | 8.9 | 4.3 | 4.3 | 2.1 | 99.0 | ||
| (PGF05) G-V | 12.5 | 12.5 | 8.9 | 7.7 | 10.0 | 5.4 | 5.4 | 2.1 | 1.0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shawky, M.; Taha, E.; Ahmed, B.; Mahmoud, M.A.; Abdelaziz, M.; Faisal, M.; Yousif, A. Initial Evidence That Gilthead Seabream (Sparus aurata L.) Is a Host for Lymphocystis Disease Virus Genotype I. Animals 2021, 11, 3032. https://doi.org/10.3390/ani11113032
Shawky M, Taha E, Ahmed B, Mahmoud MA, Abdelaziz M, Faisal M, Yousif A. Initial Evidence That Gilthead Seabream (Sparus aurata L.) Is a Host for Lymphocystis Disease Virus Genotype I. Animals. 2021; 11(11):3032. https://doi.org/10.3390/ani11113032
Chicago/Turabian StyleShawky, Mohamed, Engy Taha, Basem Ahmed, Mahmoud Aly Mahmoud, Mohamed Abdelaziz, Mohamed Faisal, and Ausama Yousif. 2021. "Initial Evidence That Gilthead Seabream (Sparus aurata L.) Is a Host for Lymphocystis Disease Virus Genotype I" Animals 11, no. 11: 3032. https://doi.org/10.3390/ani11113032