
The Influence of Dehulling on the Nutritional Quality of Lupine Seeds (Lupinus albus L.) and the Effect of Their Use in the Feed of Laying Quails on the Live Performance and Quality of Eggs

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Lupine Seeds, Chemical Composition and Nutritional Quality Analysis
2.2. Quails, Experimental Design and Diets

{kind=link}
| Specification | 1 C | Experimental Diets | |||
|---|---|---|---|---|---|
| 2 WLS20 | 3 WLS25 | 4 DLS20 | 5 DLS25 | ||
| Composition of feed (%) | |||||
| Maize (8.0% Cp) | 46.03 | 41.85 | 41.10 | 49.03 | 49.75 |
| Triticale (11.4% Cp) | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Soybean meal (46% Cp) | 33.00 | 16.50 | 12.30 | 12.00 | 6.80 |
| Lupine whole seeds | - | 20.00 | 25.00 | - | - |
| Lupine dehulled seeds | - | - | - | 20.00 | 25.00 |
| Sunflower oil | 3.20 | 3.85 | 3.80 | 1.10 | 0.50 |
| DL-Methionine | 0.02 | 0.05 | 0.05 | 0.05 | 0.05 |
| L-lysine HCl | - | - | - | 0.07 | 0.15 |
| Limestone | 5.25 | 5.25 | 5.25 | 5.25 | 5.25 |
| Vitamin–Mineral premix 6 | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| TOTAL | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Nutritional characteristics (calculated values) | |||||
| Metabolizable energy (kcal/kg) | 2901 | 2906 | 2904 | 2903 | 2903 |
| Crude protein (%) | 20.02 | 20.04 | 20.03 | 20.02 | 20.04 |
| Ether extract (%) | 5.83 | 7.90 | 8.23 | 5.55 | 5.42 |
| Crude fiber (%) | 2.88 | 4.55 | 4.98 | 2.66 | 2.61 |
| Lysine (%) | 1.02 | 1.03 | 1.01 | 1.00 | 1.00 |
| Methionine (%) | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 |
| Methionine + cysteine (%) | 0.80 | 0.81 | 0.81 | 0.83 | 0.83 |
| Calcium (%) | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| Available phosphorus (%) | 0.43 | 0.44 | 0.44 | 0.43 | 0.43 |
2.3. Performance Responses
2.4. Egg Quality Measurements
2.5. Chemical Analysis and Nutritional Quality of the Eggs
2.6. Carotenoids Content of Egg Yolk
2.7. Statistical Analysis
3. Results
3.1. The Effect of Dehulling on the Chemical Composition of White Lupine Seeds
3.2. Influence of Utilizing Dehulled White Lupine Seeds in the Diets of Laying Japanese quails
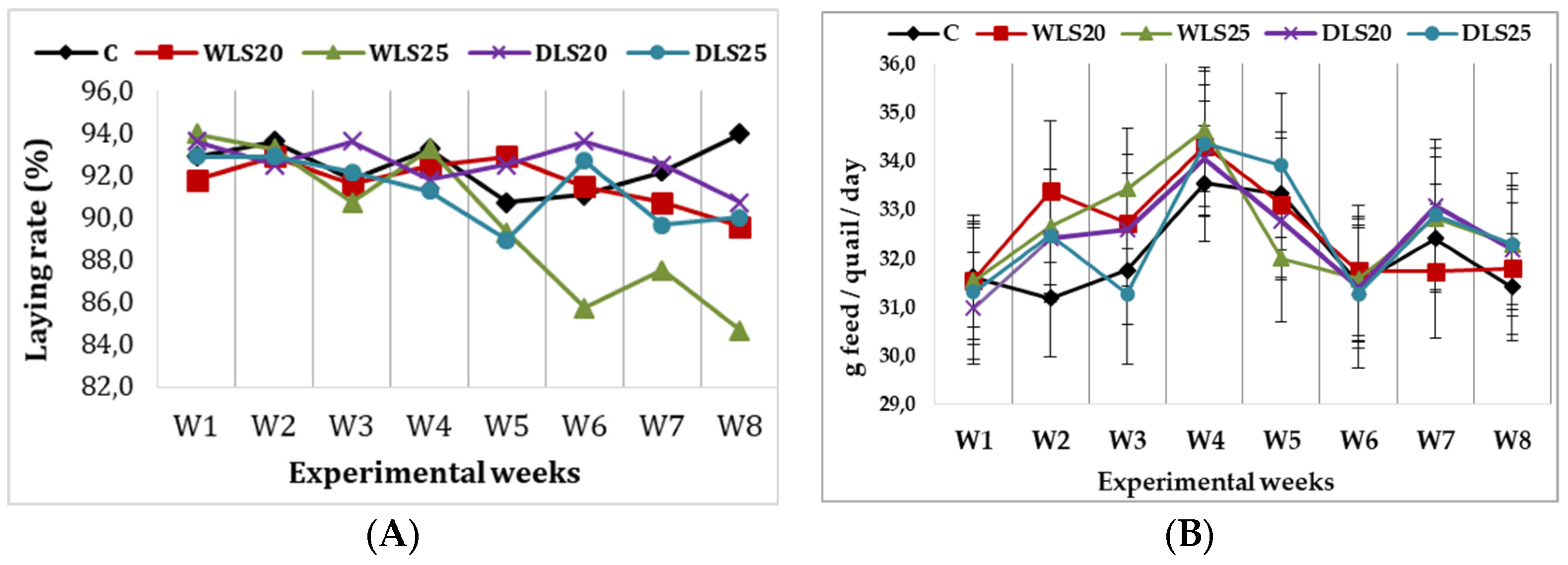
3.2.1. Performance Response
3.2.2. Egg Quality Traits
3.2.3. Egg Chemical and Nutritional Composition
3.2.4. Carotenoids Content of Egg Yolk
4. Discussion
4.1. Dehulled White Lupine Seeds
4.2. Performance Responses of Quails
4.3. Egg Quality Parameters
4.4. Chemical Composition and Nutritional Quality of Eggs
4.5. Carotenoids Content in Egg Yolk
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Visser, C.L.M.; Schreuder, R.; Stoddard, F. The EU’s dependency on soya bean import for the animal feed industry and potential for EU produced alternatives. Oilseeds Fats Crop. Lipids 2014, 21, D407. [Google Scholar] [CrossRef] [Green Version]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frías, J.; Martínez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Peter, M.Z.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [Google Scholar] [CrossRef]
- Pieper, R.; Taciak, M.; Pieper, L.; Święch, E.; Tuśnio, A.; Barszcz, M.; Vahjen, W.; Skomial, J.; Zentek, J. Comparison of the nutritional value of diets containing differentially processed blue sweet lupin seeds or soybean meal for growing pigs. Anim. Feed Sci. Technol. 2015, 221, 79–86. [Google Scholar] [CrossRef]
- Gresta, F.; Wink, M.; Prins, U.; Abberton, M.; Capraro, J.; Scarafoni, A.; Hill, G. Lupins in European cropping systems. In Legumes in Cropping Systems; Murphy-Bokern, D., Stoddard, F.L., Watson, C.A., Eds.; CAB International: Wallingford, UK, 2017; pp. 88–108. [Google Scholar]
- Kim, J.C.; Mullan, B.P.; Heo, J.M.; Hernandez, A.; Pluske, J.R. Variation in digestible energy content of Australian sweet lupins (Lupinus angustifolius L.) and the development of prediction equations for its estimation. Anim. Sci. J. 2009, 87, 2565–2573. [Google Scholar] [CrossRef] [Green Version]
- Chiofalo, B.; Lo Presti, V.; Chiofalo, V.; Gresta, F. The productive traits, fatty acid profile and nutritional indices of three lupin (Lupinus spp.) species cultivated in a Mediterranean environment for the livestock. Anim. Feed Sci. Technol. 2012, 171, 230–239. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Manunza, P.; Arnoldi, A.; Boschin, G. Quality of Lupinus albus L. (White Lupin) seed: Extent of genotypic and environmental effects. J. Agric. Food Chem. 2014, 62, 6539–6545. [Google Scholar] [CrossRef]
- Księża, J.; Staniak, M.; Bojarszczuk, J. Nutrient contents in yellow lupine (Lupinus luteus L.) and blue lupine (Lupinus angustifolius L.) cultivars depending on habitat conditions. Pol. J. Environ. Stud. 2018, 27, 1145–1153. [Google Scholar] [CrossRef]
- Písaříková, B.; Zralý, Z. Nutritional value of lupine in the diets for pigs (a review). Acta Vet. Brno 2009, 78, 399–409. [Google Scholar] [CrossRef]
- Musco, N.; Cutrignelli, M.I.; Calabrò, S.; Tudisco, R.; Infascelli, F.; Grazioli, R.; Lo Presti, V.; Gresta, F.; Chiofalo, B. Comparison of nutritional and antinutritional traits among different species (Lupinus albus L., Lupinus luteus L., Lupinus angustifolius L.) and varieties of lupin seeds. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1227–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sujak, A.; Kotlarz, A.; Strobel, W. Compositional and nutritional evaluation of several lupin seeds. Food Chem. 2006, 98, 711–719. [Google Scholar] [CrossRef]
- Boschin, G.; D’Agostina, A.; Annicchiarico, P.; Arnoldi, A. Effect of genotype and environment on fatty acid composition of Lupinus albus L. seed. Food Chem. 2008, 108, 600–606. [Google Scholar] [CrossRef]
- Mierliţă, D.; Simeanu, D.; Pop, I.M.; Criste, F.; Pop, C.; Simeanu, C.; Lup, F. Chemical composition and nutritional evaluation of the lupine seeds (Lupinus albus L.) from low-alkaloid varieties. Rev. Chim. 2018, 69, 453–458. [Google Scholar] [CrossRef]
- Knudsen, K.E.B. Fiber and nonstarch polysaccharide content and variation in common crops used in broiler diets. Poult. Sci. 2014, 93, 2380–2393. [Google Scholar] [CrossRef] [PubMed]
- Zdunczyk, Z.; Mikulski, D.; Jankowski, J.; Przybylska-Gornowicz, B.; Juskiewicz, J. Gastrointestinal response of laying hens to graded dietary inclusion levels of yellow lupine seeds. Anim. Feed Sci. Technol. 2019, 255, 114214. [Google Scholar] [CrossRef]
- Jeroch, H.; Kozłowski, K.; Schöne, F.; Zduńczyk, Z. Lupines (Lupinus spp.) as a protein feedstuff for poultry. Varieties, composition and nutritional values for poultry. Eur. Poult. Sci. 2016, 80. [Google Scholar] [CrossRef]
- Wils, P.E.L.; Dilger, R.N. Combined dietary effects of supplemental threonine and purified fiber on growth performance and intestinal health of young chicks. Poult. Sci. 2013, 92, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Smulikowska, S.; Konieczka, P.; Czerwinski, J.; Mieczkowska, A.; Jankowiak, J. Feeding broiler chickens with practical diets containing lupin seeds (L. angustifolius or L. luteus): Effects of incorporation level and mannanase supplementation on growth performance, digesta viscosity, microbial fermentation and gut morphology. J. Anim. Feed Sci. 2014, 23, 64–72. [Google Scholar] [CrossRef]
- Drażbo, A.; Mikulski, D.; Zduńczyk, Z.; Szmatowicz, B.; Rutkowski, A.; Jankowski, J. Fatty acid composition, physicochemical and sensory properties of eggs from laying hens fed diets containing blue lupine seeds. Eur. Poult. Sci. 2014, 78. [Google Scholar] [CrossRef]
- Lee, M.R.; Parkinson, S.; Fleming, H.R.; Theobald, V.J.; Leemans, D.K.; Burgess, T. The potential of blue lupins as a protein source, in the diets of laying hens. Vet. Anim. Sci. 2016, 1-2, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubiś, M.; Kaczmarek, S.A.; Nowaczewski, S.; Adamski, M.; Hejdysz, M.; Rutkowski, A. Influence of graded inclusion of white lupin (Lupinus albus) meal on performance, nutrient digestibility and ileal viscosity of laying hens. Br. Poult. Sci. 2018, 59, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, S.A.; Kasprowicz, P.M.; Hejdysz, M.; Mikuła, R.; Rutkowski, A. The nutritional value of narrow-leafed lupin (Lupinus angustifolius) for broilers. J. Anim. Feed Sci. 2014, 23, 160–166. [Google Scholar] [CrossRef]
- Mierliţă, D. The effect of lupine seed in broiler diet on animal performance and fatty acids profile of their meat. Bull. UASVM Anim. Sci. Biotech. 2015, 72, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Mikulski, D.; Zduńczyk, Z.; Juśkiewicz, J.; Rogiewicz, A.; Jankowski, J. The effect of different blue lupine (L. angustifolius) inclusion levels on gastrointestinal function, growth performance and meat quality in growing-finishing turkeys. Anim. Feed Sci. Technol. 2014, 198, 347–352. [Google Scholar] [CrossRef]
- Embaby, H.E.-S. Effect of soaking, dehulling, and cooking methods on certain antinutrients and in vitro protein digestibility of bitter and sweet lupin seeds. Food Sci. Biotechnol. 2010, 19, 1055–1062. [Google Scholar] [CrossRef]
- Batterham, E.S.; Andersen, L.M.; Lowe, R.F.; Darnell, R.E. Nutritional value of lupin (Lupinus albus)—seed meal for growing pigs: Availability of lysine, effect of autoclaving and net energy content. Br. J. Nutr. 1986, 56, 645. [Google Scholar] [CrossRef] [Green Version]
- Omer, M.A.M.; Mohamed, E.A.; Ahmed, I.A.M.; Yagoub, A.A.; Babiker, E.E. Effect of different processing methods on anti-nutrients content and protein quality of improved lupin (Lupinus albus L.) cultivar seeds. Turk. J. Agric. Food Sci. Technol. 2016, 4, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Rutkowski, A.; Kaczmarek, S.A.; Hejdysz, M.; Jamroz, D. Effect of extrusion on nutrients digestibility, metabolizable energy and nutritional value of yellow lupine seeds for broiler chickens. Ann. Anim. Sci. 2016, 16, 1059. [Google Scholar] [CrossRef] [Green Version]
- Hejdysz, M.; Kaczmarek, S.A.; Kubiś, M.; Jamroz, D.; Kasprowicz-Potocka, M.; Zaworska, A.; Rutkowski, A. Effect of increasing levels of raw and extruded narrow-leafed lupin seeds in broiler diet on performance parameters, nutrient digestibility and AMEN value of diet. J. Anim. Feed Sci. 2018, 27, 55–64. [Google Scholar] [CrossRef]
- Chilomer, K.; Kasprowicz-Potocka, M.; Gulewicz, P.; Frankiewicz, A. The influence of lupin seed germination on the chemical composition and standardized ileal digestibility of protein and amino acids in pigs. J. Anim. Physiol. Anim. Nutr. 2012, 97, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Mieczkowska, A.; Jansman, A.J.M.; Kwakkel, R.P.; Smulikowska, S. Effect of dehulling and α-galactosidase supplement on the ileal digestibility of yellow lupin based diets in broiler chickens and adult roosters. J. Anim. Feed Sci. 2005, 14, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Písaříková, B.; Zraly, Z.; Bunka, F.; Trckova, M. Nutritional value of white lupine cultivar Butan in diets for fattening pigs. Vet. Med. 2008, 53, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Saez, P.; Borquez, A.; Dantagnan, P.; Hernández, A. Effects of dehulling, steam-cooking and microwave-irradiation on digestive value of white lupin (Lupinus albus) seed meal for rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Arch. Anim. Nutr. 2015, 69, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Brenes, A.; Slominski, B.; Marquardt, R.; Guenter, W.; Viveros, A. Effect of enzyme addition on the digestibilities of cell wall polysaccharides and oligosaccharides from whole, dehulled, and ethanol-extracted white lupins in chickens. Poult. Sci. 2003, 82, 1716–1725. [Google Scholar] [CrossRef]
- Nalle, C.L.; Ravindran, G.; Ravindran, V. Influence of dehulling on the apparent metabolisable energy and ileal amino acid digestibility of grain legumes for broilers. J. Sci. Food Agric. 2010, 90, 1227–1231. [Google Scholar] [CrossRef]
- Laudadio, V.; Tufarelli, V. Dehulled-micronised lupin (Lupinus albus L. cv. Multitalia) as the main protein source for broilers: Influence on growth performance, carcass traits and meat fatty acid composition. J. Sci. Food Agric. 2011, 91, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Laudadio, V.; Tufarelli, V. Influence of substituting dietary soybean meal for dehulled-micronized lupin (Lupinus albus cv. Multitalia) on early phase laying hens production and egg quality. Livest. Sci. 2011, 140, 184–188. [Google Scholar] [CrossRef]
- Arslan, C.; Seker, E. Effects of processed white lupin seed (Lupinus albus L.) on growth performance of japanese quail. Revue Méd. Vét. 2002, 153, 643–646. [Google Scholar]
- Minvielle, F.; Mills, A.D.; Faure, J.M.; Monvoisin, J.L.; Gourichon, D. Fearfulness and performance related traits in selected lines of Japanese quail (Coturnix japonica). Poult. Sci. 2002, 81, 321–326. [Google Scholar] [CrossRef]
- Minvielle, F. The future of Japanese quail for research and production. Worlds Poult. Sci. J. 2004, 60, 500–507. [Google Scholar] [CrossRef]
- Bragagnolo, N.; Rodriguez, A.D.B. Comparison of the cholesterol content of Brazilian chicken and quail eggs. J. Food Compos. Anal. 2003, 16, 147–153. [Google Scholar] [CrossRef]
- Genchev, A. Quality and composition of Japanese quail eggs (Coturnix japonica). Trakia J. Sci. 2012, 10, 91–101. [Google Scholar]
- Akram, M.; Hussain, J.; Ahmad, S.; Rehman, A.; Lohani, F.; Munir, A.; Amjad, R.; Noshahi, H. Comparative study on production performance, egg geometry, quality and hatching traits in four close-bred stocks of Japanese quail. Adv. Agric. 2014, 3, 13–18. [Google Scholar] [CrossRef]
- Fernandez, I.B.; Calixto, L.F.L.; Torres-Cordido, K.A.A.; Lemos, M.J.; Togashi, C.K.; Souza, D.S.; Pizzolante, C.C. Feeding time under performance and eggs quality of quails in production. Rev. Bras. Saúde Produção Anim. 2018, 19, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Official Method of Analysis, 18th ed.; The Association of Official Analytical Chemists: Washington, DC, USA, 2005.
- Sibbald, I.R.; Morse, P.M. Effects of the nitrogen correction and of feed intake on true metabolizable energy values. Poult. Sci. 1983, 62, 138–142. [Google Scholar] [CrossRef]
- National Research Council (NRC). Subcommittee on Poultry Nutrition 1994. Nutrient requirements of ring-necked pheasants, Japanese quail, and bobwhite quail. In Nutrient Requirements of Poultry, 9th ed.; National Academy of Sciences: Washington, DC, USA, 1994; pp. 44–45. [Google Scholar]
- Williams, K.C. Some factors affecting albumen quality with particular reference to Haugh units score. Worlds Poult. Sci. J. 1992, 48, 5–16. [Google Scholar] [CrossRef]
- Official Method of Analysis of the Association of Official Analytical Chemists, 16th ed.; The Association of Official Analytical Chemists, Inc.: Arlington, VA, USA, 1996.
- Horwitz, W. AOAC Guidelines for Single Laboratory Validation of Chemical Methods for Dietary Supplements and Botanicals; AOAC International: Gaithersburg, MD, USA, 2002; pp. 1–38. [Google Scholar]
- Panaite, T.; Criste, R.D.; Ropota, M.; Cornescu, G.M.; Alexandrescu, D.C.; Criste, V.; Vasile, G.; Olteanu, M.; Untea, A. Effect of layer diets enriched in omega-3 fatty acids supplemented with Cu on the nutritive value of the eggs. Rom. Biotechnol. Lett. 2016, 21, 11652–11660. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W. Preparation of methyl ester and other derivatives. In Gas-Chromatography and Lipids. A Practical Guide, 1st ed.; Christie, W.W., Ed.; Oily Press: Glasgow, UK, 1989; pp. 36–47. [Google Scholar]
- Timmons, J.S.; Weiss, W.P.; Palmquist, D.L.; Harper, W.J. Relationships among dietary roasted soybeans, milk components, and spontaneous oxidized flavor of milk. J. Dairy Sci. 2001, 84, 2440–2449. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Fernandez, M.; Ordóñez, J.A.; Cambero, I.; Santos, C.; Pin, C.; de la Hoz, L. Fatty acid compositions of selected varieties of Spanish dry ham related to their nutritional implications. Food Chem. 2007, 101, 107–112. [Google Scholar] [CrossRef]
- Chen, S.; Bobe, G.; Zimmerman, S.; Hammond, E.G.; Luhman, C.M.; Boylston, T.D.; Freeman, A.E.; Beitz, D.C. Physical and sensory properties of dairy products from cows with various milk fatty acid compositions. J. Agric. Food Chem. 2004, 52, 3422–3428. [Google Scholar] [CrossRef] [PubMed]
- Schlatterer, J.; Breithaupt, D.E. Xanthophylls in commercial egg yolks: Quantification and identification by HPLC and LC-(APCI) MS using a C30 phase. J. Agric. Food Chem. 2006, 54, 2267–2273. [Google Scholar] [CrossRef]
- Mierliță, D. Studies on cultivation suitability and nutritional characterization of lupine alkaloid-free varieties. An. Univ. Din Oradea Fasc. Ecotoxicologie Zooteh. Și Tehnol. De Ind. Aliment. 2012, 11, 501–507. [Google Scholar]
- Krawczy, M.; Przywitowski, M.; Mikulski, D. Effect of yellow lupine (L. luteus) on the egg yolk fatty acid profile, the physicochemical and sensory properties of eggs, and laying hen performance. Poult. Sci. 2015, 94, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Criste, F.L.; Mierlita, D.; Simeanu, D.; Boisteanu, P.; Pop, I.M.; Georgescu, B.; Nacu, G. Study of fatty acids profile and oxidative stability of egg yolk from hens fed a diet containing white lupine seeds meal. Rev. Chim. 2018, 69, 2454–2460. [Google Scholar] [CrossRef]
- Rybiński, W.; Święcicki, W.; Bocianowski, J.; Börner, A.; Starzycka, K.E.; Starzycki, M. Variability of fat content and fatty acids profiles in seeds of a Polish white lupin (Lupinus albus L.) collection. Genet. Resour. Crop Evol. 2018, 65, 417–431. [Google Scholar] [CrossRef] [Green Version]
- Suchý, P.; Straková, E.; Kroupa, L.; Vecerek, V. The fatty acid content of oil from seeds of some lupin varieties. In Lupins for Health and Wealth, Proceedings of the 12th International Lupin Conference, Fremantle, Western Australia, 14–18 September 2008; Palta, J.A., Berger, J.D., Eds.; International Lupin Association: Canterbury, New Zealand, 2008; pp. 188–191. [Google Scholar]
- Volek, Z.; Bureš, D.; Uhlířová, L. Effect of dietary dehulled white lupine seed supplementation on the growth, carcass traits and chemical, physical and sensory meat quality parameters of growing-fattening rabbits. Meat Sci. 2018, 141, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Grela, E.R.; Kiczorowska, B.; Samolińska, W.; Matras, J.; Kiczorowski, P.; Rybiński, W.; Hanczakowska, E. Chemical composition of leguminous seeds: Part I—content of basic nutrients, amino acids, phytochemical compounds, and antioxidant activity. Eur. Food Res. Technol. 2017, 243, 1385–1395. [Google Scholar] [CrossRef]
- Mera-Zúñiga, F.; Pro-Martínez, A.; Zamora, N.J.F.; Sosa, M.E.; Guerrero, R.J.D.; Mendoza, P.S.I.; Cuca, G.J.M.; López, R.R.M.; Chan, D.D.; Becerril, P.C.M.; et al. Soybean meal substitution by dehulled lupine (Lupinus angustifolius) with enzymes in broiler diets. Asian-Australas. J. Anim. Sci. 2018, 32, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, X.H.; Nguyen, H.T.; Morgan, N.K. Dietary soluble non-starch polysaccharide level and xylanase supplementation influence performance, egg quality and nutrient utilization in laying hens fed wheat-based diets. Anim. Nutr. 2021, 7, 512–520. [Google Scholar] [CrossRef]
- Rutkowski, A.; Hejdysz, M.; Kaczmarek, S.; Adamski, M.; Nowaczewski, S.; Jamroz, D. The effect of addition of yellow lupin seeds (Lupinus luteus L.) to laying hen diets on performance and egg quality parameters. J. Anim. Feed Sci. 2017, 26, 247–256. [Google Scholar] [CrossRef]
- Canogullari, S.; Baylan, M.; Erdogan, Z.; Duzguner, V.; Kucukgul, A. The effects of dietary garlic powder on performance, egg yolk and serum cholesterol concentrations in laying quails. Czech J. Anim. Sci. 2010, 55, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, S.S.; Yesilbag, D.; Meral, Y.; Cetin, I.; Biricik, H. Juniper oil improves oxidative stability and eggshell and albumin quality of quail eggs. Br. Poult. Sci. 2015, 56, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Fathi, M.M.; Al-Homidan, I.; Ebeid, T.A.; Galal, A.; Abou-Emera, O.K. Assessment of residual feed intake and its relevant measurements in two varieties of Japanese quails (Coturnix coturnix japonica) under high environmental temperature. Animals 2019, 9, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Moneim, A.E.; Sabic, E.M. Beneficial effect of feeding olive pulp and Aspergillus awamori on productive performance, egg quality, serum/yolk cholesterol and oxidative status in laying Japanese quails. J. Anim. Feed Sci. 2019, 28, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Lee, S.I.; Kim, I.H. Effects of lupin seed supplementation on egg production performance, and qualitative egg traits in laying hens. Vet. Med. 2016, 61, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Zdunczyk, Z.; Jankowski, J.; Rutkowski, A.; Sosnowska, E.; Drazbo, A.; Zdunczyk, P.; Juskiewicz, J. The composition and enzymatic activity of gut microbiota in laying hens fed diets supplemented with blue lupine seeds. Anim. Feed Sci. Technol. 2014, 191, 57–66. [Google Scholar] [CrossRef]
- Al-Daraji, H.J.; Razuki, W.M.; Al-Hayani, W.K.; Al-Hassani, A.S. Effect of dietary linseed on egg quality of laying quail. Int. J. Poult. Sci. 2010, 9, 584–590. [Google Scholar] [CrossRef] [Green Version]
- Christaki, E.V.; Bonos, E.M.; Florou-Paneri, P.C. Comparative evaluation of dietary oregano, anise and olive leaves in laying Japanese quails. Braz. J. Poult. Sci. 2011, 13, 97–101. [Google Scholar] [CrossRef]
- Çabuk, M.; Eratak, S.; Basmacioğlu, M.H. Effects of dietary inclusion of lentil byproduct on performance and oxidative stability of eggs in laying quail. Sci. World J. 2014, 1–5. [Google Scholar] [CrossRef]
- Elangovan, A.V.; Verma, S.V.S.; Sastry, V.R.B.; Singh, S.D. Laying performance of Japanese quail fed graded levels of neem (Azadirachta indica) kernel meal incorporated diets. Anim. Feed Sci. Technol. 2000, 88, 113–120. [Google Scholar] [CrossRef]
- Konca, Y.; Yalcin, H.; Karabacak, M.; Kaliber, M.; Durmuscelebi, F.Z. Effect of hempseed (Cannabis sativa L.) on performance, egg traits and blood biochemical parameters and antioxidant activity in laying Japanese Quail (Coturnix coturnix japonica). Br. Poult. Sci. 2014, 55, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Nemati, Z.; Ahmadian, H.; Besharati, M.; Lesson, S.; Alirezalu, K.; Domínguez, R.; Lorenzo, J.M. Assessment of dietary selenium and vitamin E on laying performance and quality parameters of fresh and stored eggs in Japanese Quails. Foods 2020, 9, 1324. [Google Scholar] [CrossRef]
- Silva, W.J.; Gouveia, A.B.V.; Sousa, F.E.; Santos, F.R.; Minafra-Rezende, C.S.; Silva, J.M.S.; Minafra, C.S. Turmeric and sorghum for egg-laying quails. Ital. J. Anim. Sci. 2017, 17, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Abou-Elkhair, R.; Abdo Basha, H.; Slouma Hamouda Abd El Naby, W.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A.; Naiel, M.A.E. Effect of a diet supplemented with the Moringa oleifera seed powder on the performance, egg quality, and gene expression in Japanese Laying quail under heat-stress. Animals 2020, 10, 809. [Google Scholar] [CrossRef]
- Parizadian, B.; Ahangari, Y.J.; Shargh, M.S.; Sardarzadeh, A. Effects of different levels of L-carnitine supplementation on egg quality and blood parameters of laying Japanese quail. Int. J. Poult. Sci. 2011, 10, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Dalle Zotte, A.; Singh, Y.; Michiels, J.; Cullere, M. Black soldier fly (Hermetia Illucens) as dietary source for laying quails: Live performance, and egg physico-chemical quality, sensory profile and storage stability. Animals 2019, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammershøj, M.; Steenfeldt, S. Effects of blue lupin (Lupinus angustifolius) in organic layer diets and supplementation with foraging material on egg production and some egg quality parameters. Poult. Sci. 2005, 84, 723–733. [Google Scholar] [CrossRef]
- Timová, I.; Straková, E.; Všetičková, L.; Suchý, P. Impact of feeding mixture containing lupin meal on improvement of polyunsaturated fatty acids in egg yolk. Czech J. Anim. Sci. 2020, 65, 311–321. [Google Scholar] [CrossRef]
- Ozogul, Y.; Ozogul, F. Fatty acid profiles of commercially important fish species from the Mediterranean, Aegen and Black Seas. Food Chem. 2007, 100, 1634–1638. [Google Scholar] [CrossRef]
- Simopoulos, A. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Akdemir, F.; Orhan, C.; Sahin, N.; Sahin, K.; Hayirli, A. Tomato powder in laying hen diets: Effects on concentrations of yolk carotenoids and lipid peroxidation. Br. Poult. Sci. 2012, 53, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Errington, S.; Yap, H.H. Studies on carotenoids from lupin seeds. In Lupins for Health and Wealth, Proceedings of the 12th International Lupin Conference, Fremantle, Western Australia, 14–18 September 2008; Palta, J.A., Berger, J.D., Eds.; International Lupin Association: Canterbury, New Zealand, 2008; pp. 14–18. [Google Scholar]
- Karamać, M.; Orak, H.H.; Amarowicz, R.; Orak, A.; Piekoszewski, W. Phenolic contents and antioxidant capacities of wild and cultivated white lupin (Lupinus albus L.) seeds. Food chem. 2018, 258, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Karadas, F.; Grammenidis, E.; Surai, P.F.; Acamovic, T.; Sparks, N.H.C. Effects of carotenoids from lucerne, marigold and tomato on egg yolk pigmentation and carotenoid composition. Br. Poult. Sci. 2006, 47, 561–566. [Google Scholar] [CrossRef] [PubMed]

| Specification (n = 5) | Whole Lupine Seeds | Dehulled Lupine Seeds | Tukey Test |
|---|---|---|---|
| Mean ± sd | Mean ± sd | p-Value | |
| Organic Matter | 96.03 ± 0.03 | 96.10 ± 0.09 | 0.201 |
| Crude ash | 3.97 ± 0.03 | 3.90 ± 0.09 | 0.202 |
| Crude protein | 42.98 ± 0.49 | 51.83 ± 0.27 | 0.001 |
| Ether extract | 10.69 ± 0.31 | 11.90 ± 0.53 | 0.002 |
| Crude fiber | 14.01 ± 0.81 | 4.40 ± 0.19 | 0.001 |
| N-FE extract | 28.36 ± 1.03 | 27.97 ± 0.27 | 0.463 |
| AMEN (kcal/kg DM) | 3127.82 ± 71.87 | 4049.65 ± 40.76 | 0.001 |
| Fatty Acids (n = 5) | Whole Lupine Seeds | Dehulled Lupine Seeds | Tukey Test |
|---|---|---|---|
| Mean ± sd | Mean ± sd | p-Value | |
| Miristic Acid (C14:0) | 0.27 ± 0.02 | 0.21 ± 0.01 | 0.001 |
| Pentadecanoic Acid (C15:0) | 0.13 ± 0.01 | 0.09 ± 0.01 | 0.001 |
| Palmitic Acid (C16:0) | 10.46 ± 0.18 | 10.3 ± 0.14 | 0.148 |
| Palmitoleic Acid (C16:1) | 0.67 ± 0.04 | 0.65 ± 0.01 | 0.391 |
| Stearic Acid (C18:0) | 2.61 ± 0.09 | 2.58 ± 0.01 | 0.497 |
| Oleic-cis Acid (C18:1 n-9) | 50.72 ± 0.09 | 51.04 ± 0.1 | 0.001 |
| Linoleic Acid (C18:2 n-6) | 14.51 ± 0.09 | 13.79 ± 0.18 | 0.001 |
| γ-linolenic Acid (C18:3 n-6) | 0.07 ± 0.09 | 0.09 ± 0.05 | 0.811 |
| α-linolenic Acid (C18:3 n-3) | 13.09 ± 0.17 | 13.26 ± 0.17 | 0.161 |
| Arachidic Acid (C20:0) | 0.95 ± 0.07 | 0.97 ± 0.02 | 0.541 |
| Eicosenoic Acid (C20:1 n-9) | 3.01 ± 0.03 | 3.39 ± 0.25 | 0.008 |
| Eicosadienoic Acid (C20:2 n-6) | 0.19 ± 0.01 | 0.18 ± 0.01 | 0.056 |
| Behenic Acid (C22:0) | 2.07 ± 0.02 | 2.21 ± 0.05 | 0.001 |
| Erucic Acid (C22:1 n-9) | 0.73 ± 0.02 | 0.7 ± 0.05 | 0.289 |
| Eicosapentaenoic Acid (C20:5 n-3) | 0.28 ± 0.02 | 0.18 ± 0.07 | 0.015 |
| Other fatty acids | 0.28 ± 0.05 | 0.38 ± 0.38 | 0.004 |
| Σ SFA | 16.46 ± 0.28 | 16.36 ± 0.09 | 0.466 |
| Σ MUFA | 55.12 ± 0.78 | 55.78 ± 0.14 | 0.001 |
| Σ PUFA | 28.14 ± 0.17 | 27.48 ± 0.19 | 0.001 |
| Σ UFA | 83.26 ± 0.29 | 83.28 ± 0.10 | 0.881 |
| Amino Acids (n = 6) | Whole Lupine Seeds | Dehulled Lupine Seeds | Tukey Test |
|---|---|---|---|
| Mean ± sd | Mean ± sd | p-Value | |
| Essential amino acids | |||
| Lysine | 4.99 ± 0.02 | 5.73 ± 0.14 | 0.001 |
| Methionine | 0.48 ± 0.01 | 0.55 ± 0.01 | 0.001 |
| Tryptophan | 0.47 ± 0.01 | 0.51 ± 0.01 | 0.001 |
| Histidine | 4.84 ± 0.18 | 3.61 ± 0.05 | 0.001 |
| Valine | 3.24 ± 0.02 | 3.38 ± 0.02 | 0.035 |
| Phenylalanine | 2.14 ± 0.03 | 1.06 ± 0.02 | 0.001 |
| Isoleucine | 2.33 ± 0.06 | 1.64 ± 0.03 | 0.001 |
| Leucine | 5.76 ± 0.04 | 4.01 ± 0.06 | 0.001 |
| Arginine | 9.42 ± 0.04 | 10.06 ± 0.03 | 0.001 |
| Non-essential amino acids | |||
| Serine + aspartic acid | 7.55 ± 0.10 | 9.03 ± 0.11 | 0.001 |
| Glutamine | 31.66 ± 0.55 | 36.64 ± 0.25 | 0.001 |
| Proline | 6.29 ± 0.09 | 7.99 ± 0.06 | 0.001 |
| Alanine | 2.08 ± 0.04 | 5.54 ± 0.02 | 0.001 |
| Specification | C | Experimental Treatments (Mean ± sd) | ANOVA Two-Way | ||||
|---|---|---|---|---|---|---|---|
| WLS20 | WLS25 | DLS20 | DLS25 | F-Value | p-Value | ||
| Initial weight of quails (g/quail) | 288.26 ± 17.90 a | 287.47 ± 19.67 a | 290.77 ± 15.03 a | 287.22 ± 17.79 a | 286.86 ± 14.52 a | 0.312 | 0.869 |
| Final weight of quails (g/quail) | 313.76 ± 26.73 a | 297.03 ± 24.72 b | 297.87 ± 23.52 b | 301.16 ± 24.45 ab | 297.77 ± 27.81 b | 2.981 | 0.021 |
| Daily feed intake (g of feed/quail) | 32.09 ± 1.12 a | 32.54 ± 1.33 a | 32.62 ± 1.21 a | 32.43 ± 1.18 a | 32.47 ± 1.43 a | 0.795 | 0.530 |
| Laying rate (%) | 92.41 ± 8.62 a | 91.65 ± 9.76 ab | 89.78 ± 9.69 b | 92.59 ± 9.03 a | 91.29 ± 10.07 ab | 4.132 | 0.002 |
| Feed conversion ratio: kg feed/kg egg mass | 2.68 ± 0.14 a | 2.78 ± 0.20 ab | 2.82 ± 0.16 b | 2.70 ± 0.16 a | 2.77 ± 0.21 ab | 5.983 | 0.000 |
| kg feed/dozen eggs | 0.417 ± 0.02 a | 0.426 ± 0.02 ab | 0.436 ± 0.03 b | 0.420 ± 0.03 ab | 0.427 ± 0.02 ab | 1.962 | 0.018 |
| Specification | C | Experimental Treatments (Mean ± sd) | ANOVA Two-Way | ||||
|---|---|---|---|---|---|---|---|
| WLS20 | WLS25 | DLS20 | DLS25 | F-Value | p-Value | ||
| Egg weight (g) | 13.02 ± 0.07 a | 12.89 ± 0.14 b | 12.87 ± 0.13 b | 13.04 ± 0.20 a | 12.90 ± 0.15 ab | 1.759 | 0.002 |
| Albumen (%) | 58.00 ± 1.80 a | 58.33 ± 1.94 a | 58.35 ± 1.66 a | 58.26 ± 1.85 a | 57.76 ± 1.65 a | 3.573 | 0.064 |
| Yolk (%) | 29.81 ± 1.44 a | 29.88 ± 1.66 a | 29.60 ± 1.42 a | 29.66 ± 1.80 a | 30.08 ± 1.44 a | 1.877 | 0.113 |
| Shell (%) | 12.18 ± 0.85 a | 11.81 ± 0.76 b | 12.05 ± 0.85 b | 12.10 ± 0.93 ab | 12.16 ± 0.89 a | 7.725 | 0.000 |
| Shell thickness (mm) | 0.212 ± 0.01 a | 0.197 ± 0.01 c | 0.194 ± 0.01 c | 0.203 ± 0.01 b | 0.198 ± 0.01 bc | 4.081 | 0.000 |
| Fatty Acids | C | Experimental Treatments (Mean ± sd) | ANOVA Single-Way | ||
|---|---|---|---|---|---|
| WLS20 | DLS20 | F-Value | p-Value | ||
| Miristic Acid (C14:0) | 0.30 ± 0.03 a | 0.30 ± 0.01 a | 0.29 ± 0.02 a | 0.40 | 0.680 |
| Miristoleic Acid (C14:1) | 0.04 ± 0.01 a | 0.03 ± 0.01 a | 0.04 ± 0.01 a | 2.90 | 0.094 |
| Pentadecanoic Acid (C15:0) | 0.03 ± 0.01 a | 0.03 ± 0.01 a | 0.04 ± 0.01 a | 17.49 | 0.089 |
| Pentadecenoic Acid (C15:1) | 0.13 ± 0.03 a | 0.20 ± 0.13 a | 0.14 ± 0.06 a | 1.34 | 0.298 |
| Palmitic Acid (C16:0) | 25.84 ± 0.41 a | 23.42 ± 2.41 b | 21.38 ± 0.12 b | 4.33 | 0.038 |
| Palmitoleic Acid (C16:1) | 2.92 ± 0.25 a | 2.90 ± 0.33 a | 3.50 ± 0.08 b | 9.84 | 0.003 |
| Heptadecanoic Acid (C17:0) | 0.11 ± 0.02 a | 0.10 ± 0.01 a | 0.15 ± 0.01 b | 26.82 | 0.000 |
| Heptadecenoic Acid (C17:1) | 0.18 ± 0.050 a | 0.21 ± 0.03 a | 0.18 ± 0.07 a | 0.78 | 0.482 |
| Stearic Acid (C18:0) | 15.91 ± 0.50 a | 14.22 ± 0.99 a | 13.95 ± 0.12 a | 0.35 | 0.714 |
| Oleic Acid (C18:1 n-9) | 32.26 ± 1.12 a | 33.92 ± 1.04 ab | 34.66 ± 0.22 b | 30.61 | 0.000 |
| Linoleic Acid (C18:2 n-6) | 12.49 ± 1.55 a | 15.31 ± 1.63 b | 16.31 ± 0.22 b | 25.15 | 0.000 |
| Γ-linolenic Acid (C18:3 n-6) | 0.18 ± 0.03 a | 0.27 ± 0.01 b | 0.27 ± 0.02 b | 31.84 | 0.000 |
| α-linolenic Acid (C18:3 n-3) | 0.27 ± 0.03 a | 0.12 ± 0.03 b | 0.27 ± 0.01 a | 55.79 | 0.000 |
| Eicosadienoic Acid (C20:2 n-6) | 0.17 ± 0.05 a | 0.13 ± 0.03 a | 0.17 ± 0.03 a | 2.61 | 0.115 |
| Eicosatrienoic Acid (C20:3 n-6) | 0.23 ± 0.01 a | 0.20 ± 0.05 ab | 0.15 ± 0.02 b | 9.27 | 0.004 |
| Eicosatrienoic Acid (C20:3 n-3) | 0.22 ± 0.02 a | 0.23 ± 0.01 a | 0.20 ± 0.04 a | 1.68 | 0.227 |
| Erucic Acid (C22:1 n-9) | 0.04 ± 0.01 a | 0.04 ± 0.01 a | 0.05 ± 0.01 a | 3.52 | 0.063 |
| Arachidonic Acid (C20:4 n-6) | 5.02 ± 0.12 a | 5.36 ± 0.49 b | 5.16 ± 0.04 b | 14.75 | 0.001 |
| Nervonic Acid (C24:1 n-9) | 0.32 ± 0.02 a | 0.25 ± 0.03 b | 0.16 ± 0.03 c | 39.02 | 0.000 |
| Docosatetraenoic Acid (C22:4 n-6) | 0.55 ± 0.15 a | 1.41 ± 0.17 b | 0.84 ± 0.04 a | 53.27 | 0.000 |
| Docosapentaenoic Acid (C22:5 n-3) | 0.27 ± 0.02 a | 0.14 ± 0.04 b | 0.25 ± 0.04 a | 18.89 | 0.000 |
| Docosahexaenoic Acid (C22:6 n-3) | 2.06 ± 0.05 a | 0.86 ± 0.18 b | 1.44 ± 0.07 c | 131.26 | 0.000 |
| Other fatty acids | 0.47 ± 0.28 a | 0.55 ± 0.18 a | 0.40 ± 0.17 a | 0.10 | 0.908 |
| Σ SFA | 42.19 ± 0.74 a | 38.06 ± 2.02 b | 35.80 ± 2.02 c | 5.16 | 0.024 |
| Σ UFA | 57.35 ± 0.68 s | 61.39 ± 1.97 b | 63.80 ± 1.97 c | 5.77 | 0.018 |
| Σ MUFA | 35.88 ± 1.33 a | 37.56 ± 1.35 ab | 38.74 ± 1.35 b | 24.33 | 0.000 |
| Σ PUFA | 21.47 ± 1.78 a | 23.83 ± 2.45 b | 25.06 ± 2.45 b | 14.06 | 0.001 |
| n-3 FA | 2.82 ± 0.08 a | 1.34 ± 0.22 b | 2.16 ± 0.22 c | 105.57 | 0.000 |
| n-6 FA | 18.65 ± 1.75 a | 22.48 ± 2.26 b | 22.89 ± 2.26 b | 23.17 | 0.000 |
| HFA | 26.14 ± 0.38 a | 23.71 ± 2.42 b | 21.66 ± 2.42 b | 4.37 | 0.038 |
| hFA | 53.72 ± 0.74 a | 52.74 ± 2.02 a | 59.72 ± 2.02 b | 7.26 | 0.009 |
| Indices | C | Experimental Treatments (Mean ± sd) | ANOVA Single Way | ||
|---|---|---|---|---|---|
| WLS20 | DLS20 | F-value | p-Value | ||
| PUFA/SFA | 0.51 ± 0.06 a | 0.63 ± 0.12 ab | 0.70 ± 0.12 b | 8.10 | 0.006 |
| MUFA/SFA | 0.85 ± 0.02 a | 0.99 ± 0.07 b | 1.08 ± 0.07 c | 13.29 | 0.001 |
| UFA/SFA | 1.36 ± 0.05 a | 1.61 ± 0.17 b | 1.79 ± 0.17 c | 4.88 | 0.028 |
| n-6/n-3 FA | 6.63 ± 1.32 a | 16.77 ± 0.67 b | 10.63 ± 0.67 c | 222.52 | 0.000 |
| h/H | 2.06 ± 0.07 a | 2.44 ± 0.59 ab | 2.80 ± 0.59 b | 3.68 | 0.057 |
| PI | 13.02 ± 1.60 a | 15.26 ± 1.69 ab | 16.85 ± 1.69 b | 22.60 | 0.000 |
| AI | 0.47 ± 0.02 a | 0.40 ± 0.05 b | 0.35 ± 0.05 b | 21.59 | 0.000 |
| TI | 1.17 ± 0.03 a | 1.11 ± 0.08 a | 0.95 ± 0.08 b | 20.87 | 0.000 |
| HPI | 2.12 ± 0.05 a | 2.50 ± 0.55 ab | 2.87 ± 0.55 b | 10.68 | 0.002 |
| Yolk Carotenoids | C | Experimental Treatments (Mean ± sd) | ANOVA Single-Way | ||
|---|---|---|---|---|---|
| WLS20 | DLS20 | F-Value | p-Value | ||
| Lutein | 4.28 ± 1.36 a | 7.01 ± 0.50 b | 6.01 ± 0.76 b | 10.746 | 0.002 |
| Zeaxanthin | 6.02 ± 1.52 a | 9.31 ± 1.04 b | 8.38 ± 0.18 b | 12.623 | 0.001 |
| Canthaxanthin | 3.08 ± 0.66 a | 5.55 ± 2.13 b | 3.73 ± 0.84 ab | 4.325 | 0.039 |
| ß-cryptoxanine | 0.53 ± 0.11 a | 0.93 ± 0.35 b | 0.57 ± 0.07 ab | 5.227 | 0.023 |
| Total carotenoids | 13.91 ± 3.14 a | 22.8 ± 2.13 b | 18.63 ± 1.20 b | 18.804 | 0.000 |
| Yolk color intensity | 8.86 ± 1.14 a | 12.77 ± 3.14 b | 12.14 ± 3.14 b | 120.00 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Struţi, D.I.; Bunea, A.; Pop, I.M.; Păpuc, T.A.; Mierliţă, D.P. The Influence of Dehulling on the Nutritional Quality of Lupine Seeds (Lupinus albus L.) and the Effect of Their Use in the Feed of Laying Quails on the Live Performance and Quality of Eggs. Animals 2021, 11, 2898. https://doi.org/10.3390/ani11102898
Struţi DI, Bunea A, Pop IM, Păpuc TA, Mierliţă DP. The Influence of Dehulling on the Nutritional Quality of Lupine Seeds (Lupinus albus L.) and the Effect of Their Use in the Feed of Laying Quails on the Live Performance and Quality of Eggs. Animals. 2021; 11(10):2898. https://doi.org/10.3390/ani11102898
Chicago/Turabian StyleStruţi, Dănuţ Ioan, Andrea Bunea, Ioan Mircea Pop, Tudor Andrei Păpuc, and Daniel Pantea Mierliţă. 2021. "The Influence of Dehulling on the Nutritional Quality of Lupine Seeds (Lupinus albus L.) and the Effect of Their Use in the Feed of Laying Quails on the Live Performance and Quality of Eggs" Animals 11, no. 10: 2898. https://doi.org/10.3390/ani11102898