
Deslorelin Slow-Release Implants Delay Ovulation and Increase Plasma AMH Concentration and Small Antral Follicles in Haflinger Mares

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Ultrasonography
2.4. Antral Follicle Count
2.5. Endocrine Analyses
2.6. Statistical Analysis
3. Results
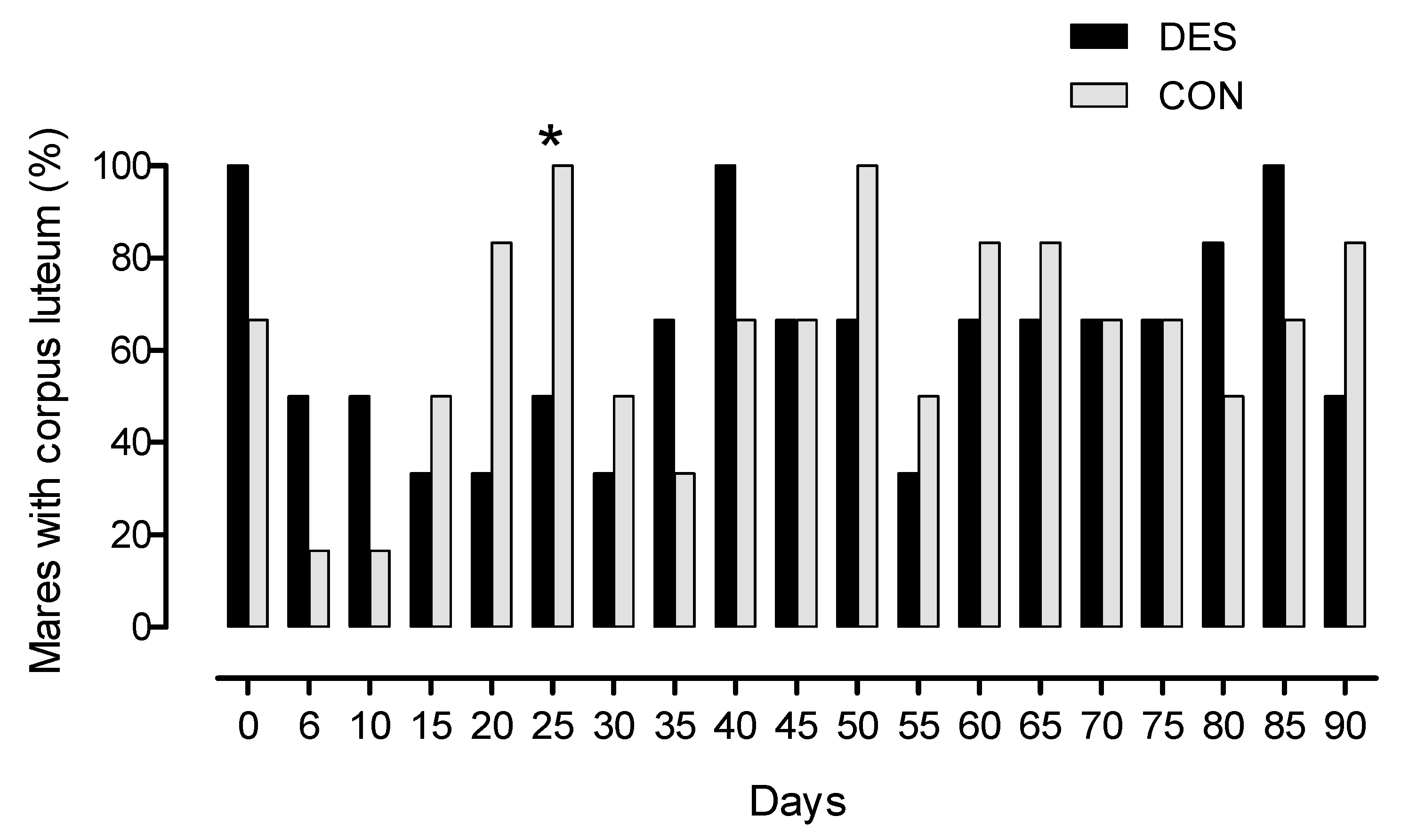
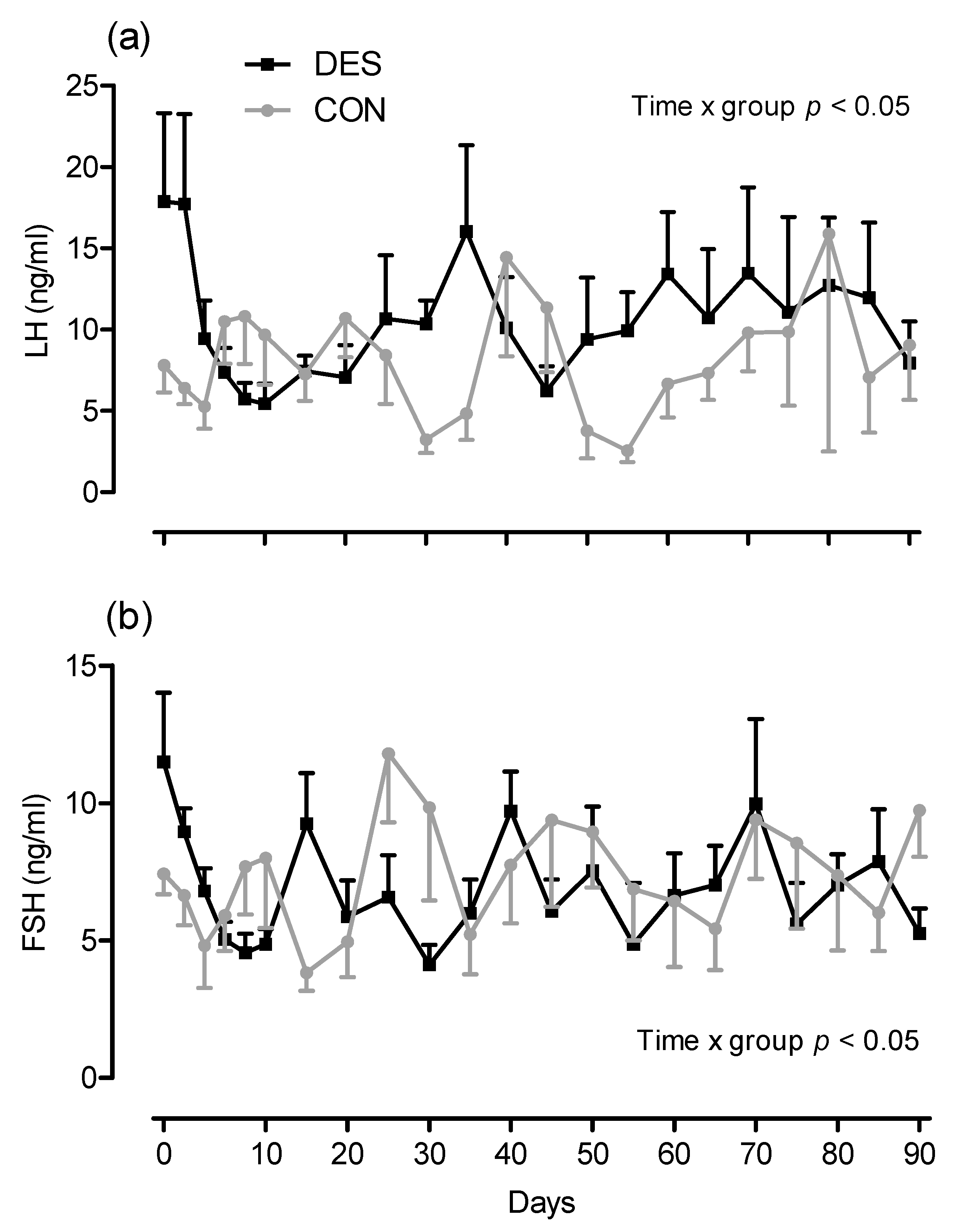
3.1. Influence of Deslorelin on Estrous Cycle Characteristics until Day 90
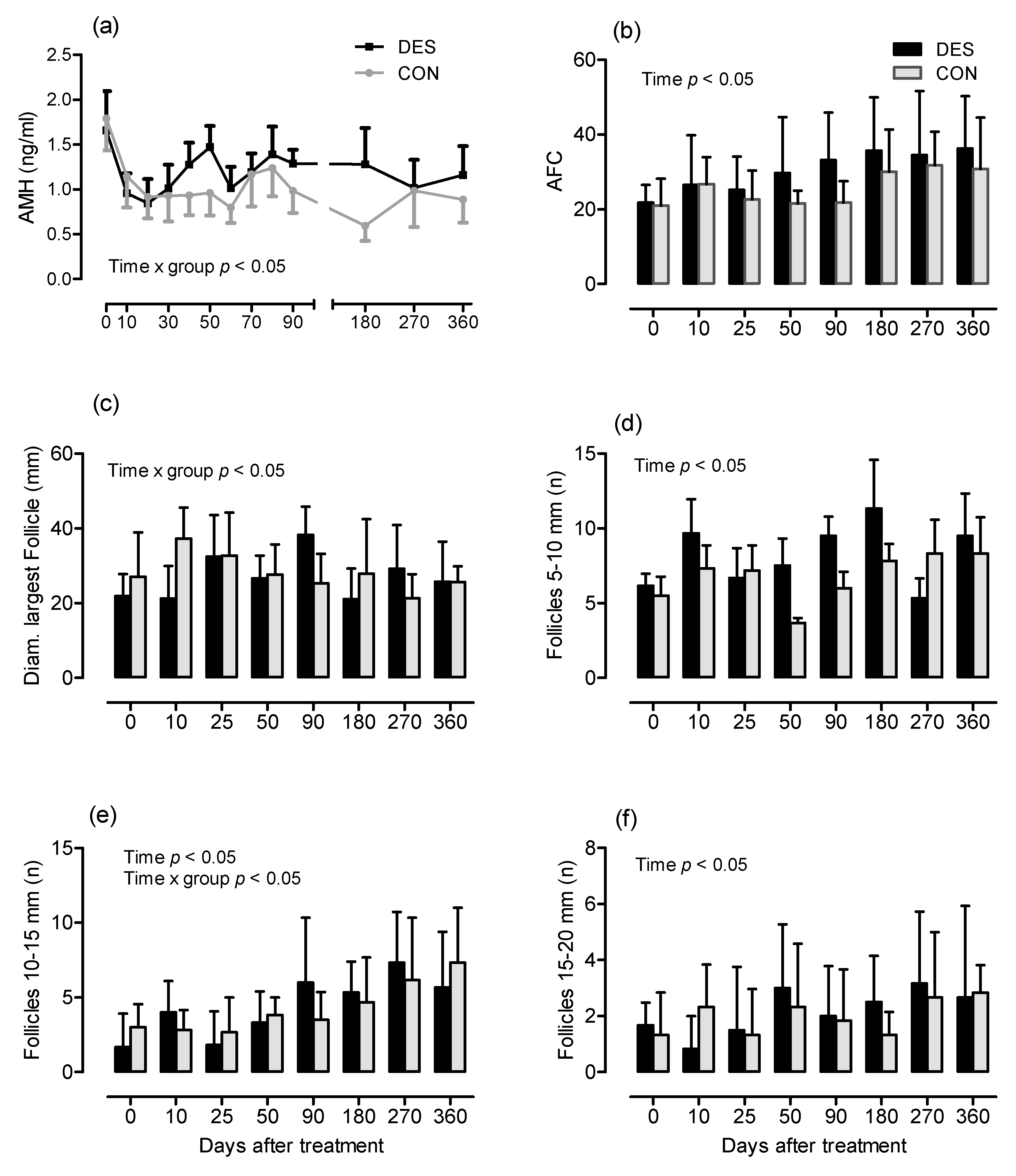
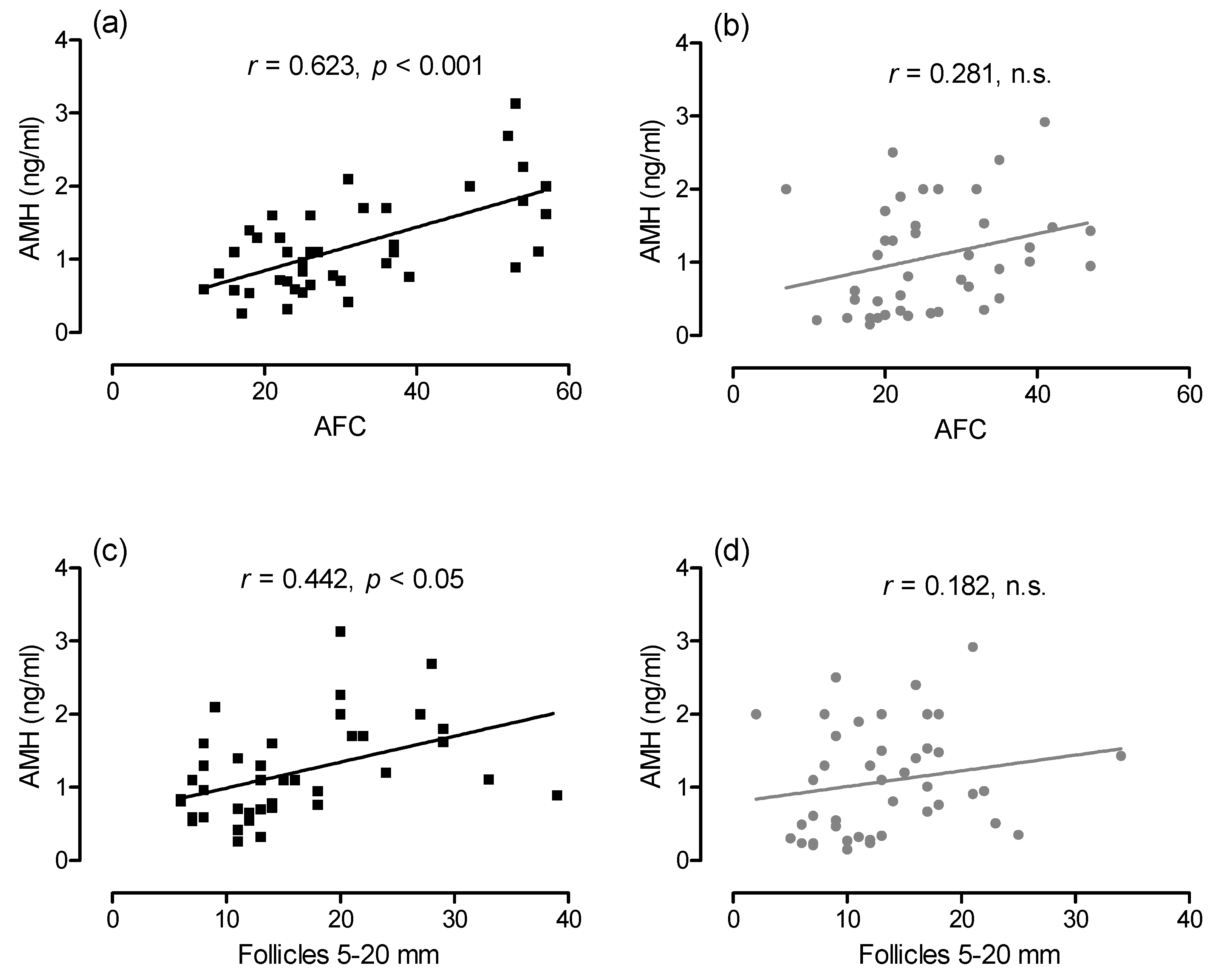
3.2. Influence of Deslorelin Treatment on Antral Follicle Count, Follicle Subpopulations and Plasma AMH Concentration until Day 360
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Junaidi, A.; Williamson, P.E.; Cummins, J.M.; Martin, G.B.; Blackberry, M.A.; Trigg, T.E. Use of a new drug delivery formulation of the gonadotrophin-releasing hormone analogue Deslorelin for reversible long-term contraception in male dogs. Reprod. Fertil. Dev. 2003, 15, 317–322. [Google Scholar] [CrossRef]
- Romagnoli, S.; Stelletta, C.; Milani, C.; Gelli, D.; Falomo, M.E.; Mollo, A. Clinical use of deslorelin for the control of reproduction in the bitch. Reprod. Domest. Anim. 2009, 44, 36–39. [Google Scholar] [CrossRef]
- Munson, L.; Bauman, J.E.; Asa, C.S.; Jöchle, W.; Trigg, T.E. Efficacy of the GnRH analogue deslorelin for suppression of oestrous cycles in cats. J. Reprod. Fertil. Suppl. 2001, 57, 269–273. [Google Scholar]
- Novotny, R.; Cizek, P.; Vitasek, R.; Bartoskova, A.; Prinosilova, P.; Janosovska, M. Reversible suppression of sexual activity in tomcats with deslorelin implant. Theriogenology 2012, 78, 848–857. [Google Scholar] [CrossRef]
- D’Occhio, M.J.; Aspden, W.J.; Whyte, T.R. Controlled, reversible suppression of estrous cycles in beef heifers and cows using agonists of gonadotropin-releasing hormone. J. Anim. Sci. 1996, 74, 218–225. [Google Scholar] [CrossRef]
- Kauffold, J.; Rohrmann, H.; Boehm, J.; Wehrend, A. Effects of long-term treatment with the GnRH agonist deslorelin (Suprelorin) on sexual function in boars. Theriogenology 2010, 74, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Goericke-Pesch, S. Long-term effects of GnRH agonists on fertility and behaviour. Reprod. Domest. Anim. 2017, 52 (Suppl. 2), 336–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montovan, S.M.; Daels, P.P.; Rivier, J.; Hughes, J.P.; Stabenfeldt, G.H.; Lasley, B.L. The effect of a potent GnRH agonist on gonadal and sexual activity in the horse. Theriogenology 1990, 33, 1305–1321. [Google Scholar] [CrossRef]
- Kaps, M.; Okada, C.T.C.; Gautier, C.; Aurich, J.; Scarlet, D.; Kuhl, J.; Aurich, C. Transient suppression of ovulatory ovarian function in pony mares after treatment with slow-release deslorelin implants. Domest. Anim. Endocrinol. 2021, 74, 106505. [Google Scholar] [CrossRef]
- Farquhar, V.J.; McCue, P.M.; Nett, T.M.; Squires, E.L. Effect of deslorelin acetate on gonadotropin secretion and ovarian follicle development in cycling mares. J. Am. Vet. Med. Assoc. 2001, 218, 749–752. [Google Scholar] [CrossRef]
- Johnson, C.A.; Thompson, D.L.; Cartmill, J.A. Pituitary responsiveness to GnRH in mares following deslorelin acetate implantation to hasten ovulation1. J. Anim. Sci. 2002, 80, 2681–2687. [Google Scholar] [CrossRef] [PubMed]
- McCue, P.M.; Farquhar, V.J.; Carnevale, E.M.; Squires, E.L. Removal of deslorelin (Ovuplant™) implant 48 h after administration results in normal interovulatory intervals in mares. Theriogenology 2002, 58, 865–870. [Google Scholar] [CrossRef]
- Farquhar, V.J.; McCue, P.M.; Carnevale, E.M.; Nett, T.M.; Squires, E.L. Deslorelin acetate (Ovuplant) therapy in cycling mares: Effect of implant removal on FSH secretion and ovarian function. Equine Vet. J. 2002, 34, 417–420. [Google Scholar] [CrossRef]
- Hinrichs, K. Assisted reproductive techniques in mares. Reprod. Domest. Anim. 2018, 53, 4–13. [Google Scholar] [CrossRef]
- Alm, H.; Torner, H.; Kanitz, W.; Becker, F.; Hinrichs, K. Comparison of different methods for the recovery of horse oocytes. Equine Vet. J. Suppl. 1997, 47–50. [Google Scholar] [CrossRef]
- Jacobson, C.C.; Choi, Y.-H.; Hayden, S.S.; Hinrichs, K. Recovery of mare oocytes on a fixed biweekly schedule, and resulting blastocyst formation after intracytoplasmic sperm injection. Theriogenology 2010, 73, 1116–1126. [Google Scholar] [CrossRef]
- Galli, C.; Duchi, R.; Colleoni, S.; Lagutina, I.; Lazzari, G. Ovum pick up, intracytoplasmic sperm injection and somatic cell nuclear transfer in cattle, buffalo and horses: From the research laboratory to clinical practice. Theriogenology 2014, 81, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Claes, A.; Ball, B.A.; Scoggin, K.E.; Esteller-Vico, A.; Kalmar, J.J.; Conley, A.J.; Squires, E.L.; Troedsson, M.H.T. The interrelationship between anti-Müllerian hormone, ovarian follicular populations and age in mares. Equine Vet. J. 2015, 47, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Henneke, D.R.; Potter, G.D.; Kreider, J.L.; Yeates, B.F. Relationship between condition score, physical measurements and body fat percentage in mares. Equine Vet. J. 1983, 15, 371–372. [Google Scholar] [CrossRef]
- Kuhl, J.; Nagel, C.; Ille, N.; Aurich, J.E.; Aurich, C. The PGF2α agonists luprostiol and d-cloprostenol reliably induce luteolysis in luteal phase mares without evoking clinical side effects or a stress response. Anim. Reprod. Sci. 2016, 168, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Mateu-Sánchez, S.; Newcombe, J.R.; Garcés-Narro, C.; Cuervo-Arango, J. The period of the follicular phase during which the uterus of mares shows estrus-like echotexture influences the subsequent pregnancy rate. Theriogenology 2016, 86, 1506–1515. [Google Scholar] [CrossRef] [PubMed]
- Scarlet, D.; Wulf, M.; Kuhl, J.; Köhne, M.; Ille, N.; Conley, A.J.; Aurich, C. Anti-Müllerian hormone profiling in prepubertal horses and its relationship with gonadal function. Theriogenology 2018, 117, 72–77. [Google Scholar] [CrossRef]
- Jennings, M.W.; Boime, I.; Daphna-Iken, D.; Jablonka-Shariff, A.; Conley, A.J.; Colgin, M.; Bidstrup, L.A.; Meyers-Brown, G.A.; Famula, T.R.; Roser, J.F. The efficacy of recombinant equine follicle stimulating hormone (reFSH) to promote follicular growth in mares using a follicular suppression model. Anim. Reprod. Sci. 2009, 116, 291–307. [Google Scholar] [CrossRef]
- Matteri, R.L.; Roser, J.F.; Baldwin, D.M.; Lipovetsky, V.; Papkoff, H. Characterization of a monoclonal antibody which detects luteinizing hormone from diverse mammalian species. Domest. Anim. Endocrinol. 1987, 4, 157–165. [Google Scholar] [CrossRef]
- Roser, J.F.; Hughes, J.P. Prolonged pulsatile administration of gonadotrophin-releasing hormone (GnRH) to fertile stallions. J. Reprod. Fertil. Suppl. 1991, 44, 155–168. [Google Scholar]
- Junaidi, A.; Williamson, P.E.; Martin, G.B.; Blackberry, M.A.; Cummins, J.M.; Trigg, T.E. Dose-response studies for pituitary and testicular function in male dogs treated with the GnRH superagonist, deslorelin. Reprod. Domest. Anim. 2009, 44, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Gautier, C.; Schmidt, K.; Aurich, J.; Aurich, C. Effect of implants containing the GnRH agonist deslorelin on testosterone release and semen characteristics in Shetland stallions. Anim. Reprod. Sci. 2018, 195, 230–241. [Google Scholar] [CrossRef]
- Checura, C.M.; Beg, M.A.; Gastal, E.L.; Gastal, M.O.; Wiltbank, M.C.; Parrish, J.J.; Ginther, O.J. Effect of suppression of FSH with a GnRH antagonist (acyline) before and during follicle deviation in the mare. Reprod. Domest. Anim. 2009, 44, 504–511. [Google Scholar] [CrossRef]
- Donadeu, F.X.; Pedersen, H.G. Follicle development in mares. Reprod. Domest. Anim. 2008, 43, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Mc Cue, P.M.; Magee, C.; Gee, E.K. Comparison of compounded deslorlin and hCG for induction of ovulation in mares. J. Equine Vet. Sci. 2007, 27, 58–61. [Google Scholar] [CrossRef]
- Ginther, O.J.; Gastal, E.L.; Gastal, M.O.; Beg, M.A. Effect of prostaglandin F2α on ovarian, adrenal, and pituitary hormones and on luteal blood flow in mares. Domest. Anim. Endocrinol. 2007, 32, 315–328. [Google Scholar] [CrossRef]
- Utt, M.D.; Acosta, T.J.; Wiltbank, M.C.; Ginther, O.J. Acute effects of prostaglandin F(2α) on systemic oxytocin and progesterone concentrations during the mid- or late-luteal phase in mares. Anim. Reprod. Sci. 2007, 97, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Beg, M.A. Dynamics of circulating progesterone concentrations before and during Luteolysis: A comparison between cattle and horses. Biol. Reprod. 2012, 86, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ball, B.A.; Conley, A.J.; MacLaughlin, D.T.; Grundy, S.A.; Sabeur, K.; Liu, I.K.M. Expression of anti-Müllerian hormone (AMH) in equine granulosa-cell tumors and in normal equine ovaries. Theriogenology 2008, 70, 968–977. [Google Scholar] [CrossRef]
- Batista, E.O.S.; Vieira, L.M.; Sá Filho, M.F.; Dias, E.A.R.; Bayeux, B.M.; Accorsi, M.F.; Monteiro, F.M.; Souza, A.H.; Baruselli, P.S.; D’Occhio, M.J. Ovarian follicular growth suppression by long-term treatment with a GnRH agonist and impact on small follicle number, oocyte yield, and in vitro embryo production in Zebu beef cows. Theriogenology 2016, 85, 1680–1687. [Google Scholar] [CrossRef] [PubMed]
- Cuervo-Arango, J.; Claes, A.N.; Stout, T.A.E. Mare and stallion effects on blastocyst production in a commercial equine ovum pick-up-intracytoplasmic sperm injection program. Reprod. Fertil. Dev. 2019, 31, 1894–1903. [Google Scholar] [CrossRef]
- Shabpareh, V.; Squires, E.L.; Seidel, G.E.; Jasko, D.J. Methods for collecting and maturing equine oocytes in vitro. Theriogenology 1993, 40, 1161–1175. [Google Scholar] [CrossRef]
- Roser, J.F.; Meyers-Brown, G. Enhancing fertility in mares: Recombinant equine gonadotropins. J. Equine Vet. Sci. 2019, 76, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.M.; Da Coutinho Silva, M.A.; Cruppe, L.H.; Mussard, M.L.; Bridges, G.A.; Harstine, B.R.; Smith, G.W.; Geary, T.W.; Day, M.L. Role of progesterone concentrations during early follicular development in beef cattle: I. Characteristics of LH secretion and oocyte quality. Anim. Reprod. Sci. 2018, 196, 59–68. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaps, M.; Okada, C.T.C.; Gautier, C.M.; Aurich, J.; Aurich, C. Deslorelin Slow-Release Implants Delay Ovulation and Increase Plasma AMH Concentration and Small Antral Follicles in Haflinger Mares. Animals 2021, 11, 1600. https://doi.org/10.3390/ani11061600
Kaps M, Okada CTC, Gautier CM, Aurich J, Aurich C. Deslorelin Slow-Release Implants Delay Ovulation and Increase Plasma AMH Concentration and Small Antral Follicles in Haflinger Mares. Animals. 2021; 11(6):1600. https://doi.org/10.3390/ani11061600
Chicago/Turabian StyleKaps, Martim, Carolina T. C. Okada, Camille M. Gautier, Jörg Aurich, and Christine Aurich. 2021. "Deslorelin Slow-Release Implants Delay Ovulation and Increase Plasma AMH Concentration and Small Antral Follicles in Haflinger Mares" Animals 11, no. 6: 1600. https://doi.org/10.3390/ani11061600