Response of Bt and Non-Bt Cottons to High Infestations of Bollworm (Helicoverpa zea Boddie) and Tobacco Budworm (Heliothis virescens (F.)) under Sprayed and Unsprayed Conditions

 and
and
Abstract
:1. Introduction
2. Materials and Methods
Data Analysis
3. Results
3.1. Insect Densities
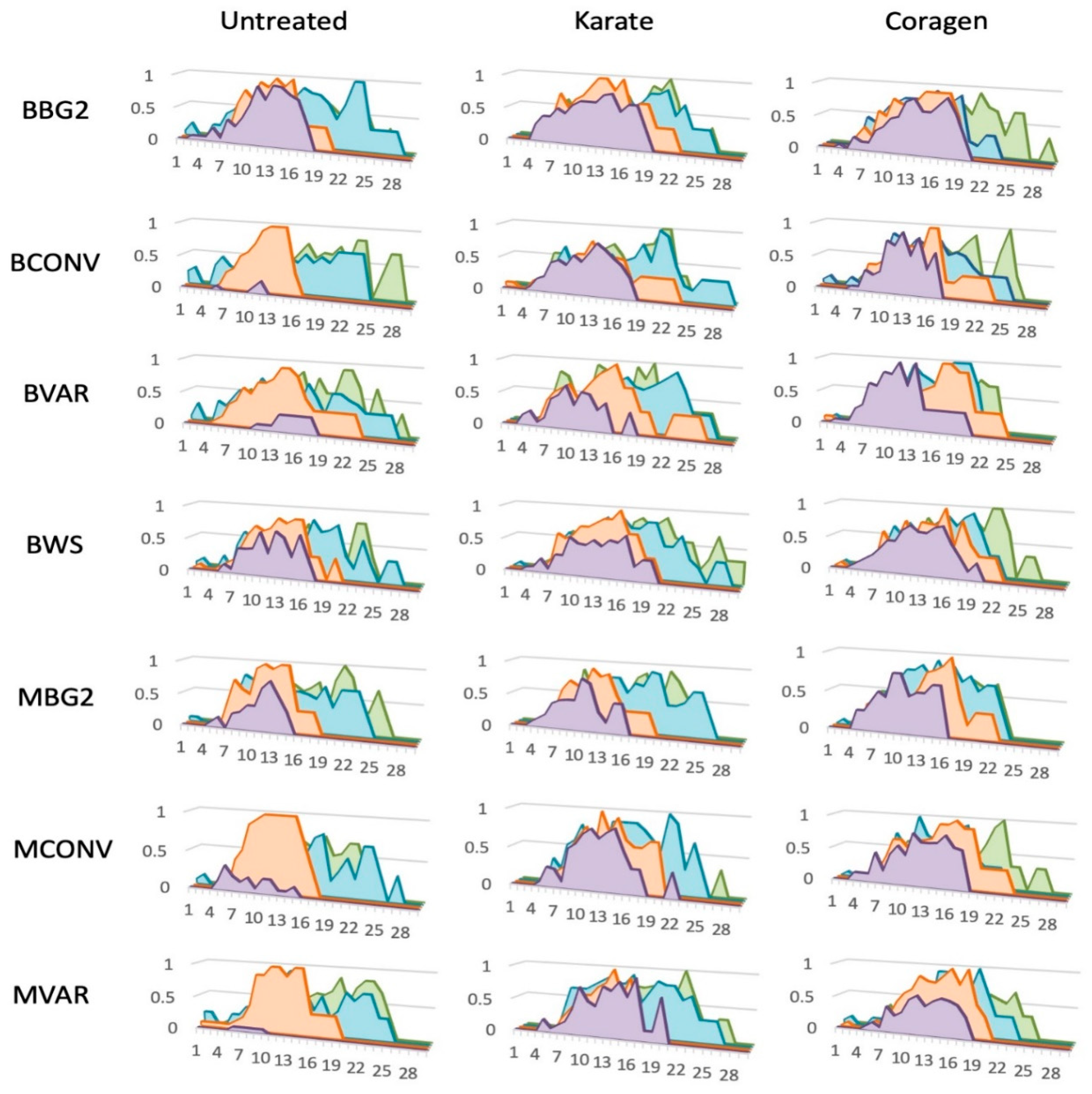
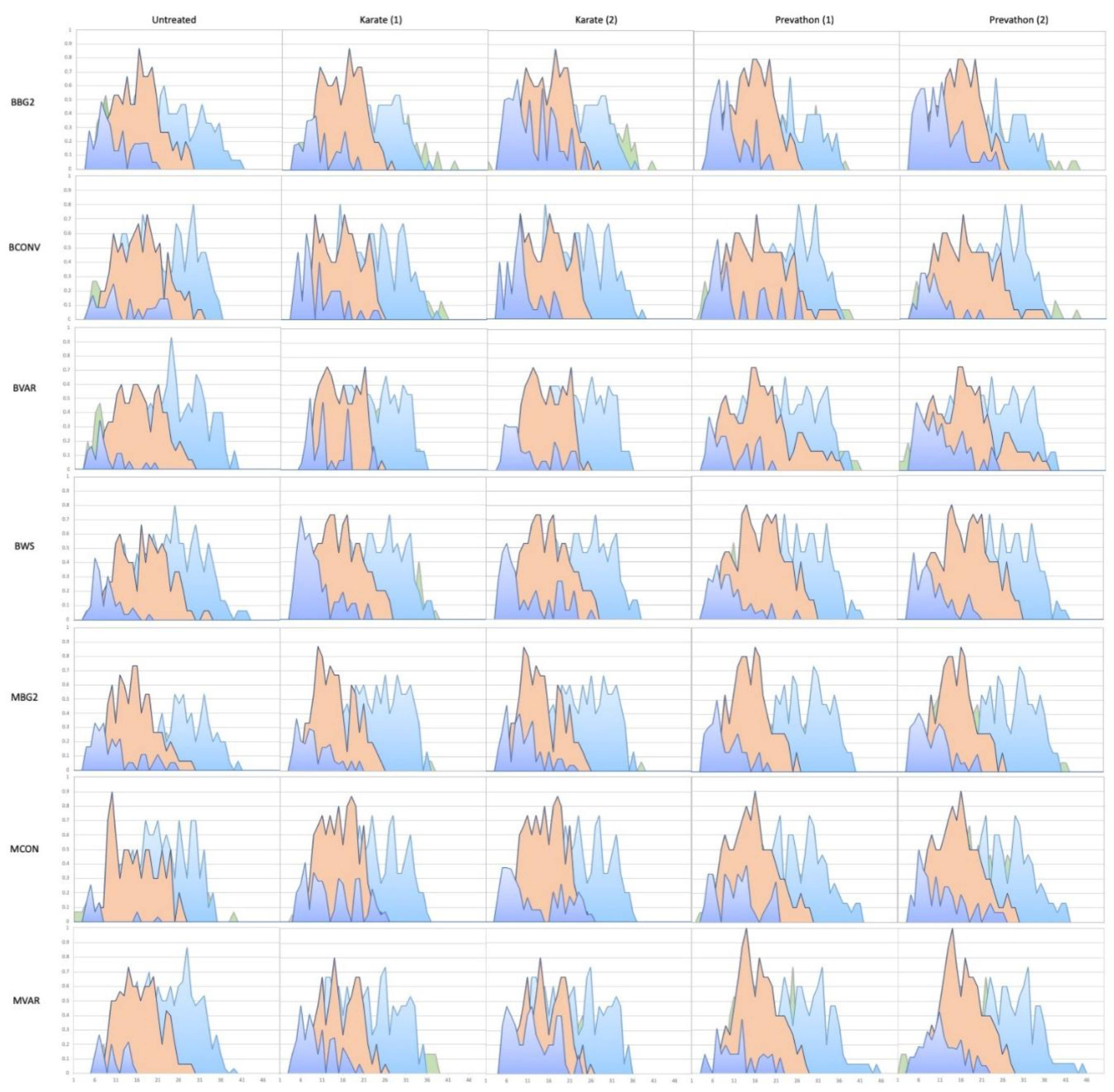
3.2. Within-Season Retention of 1st Postion Fruit
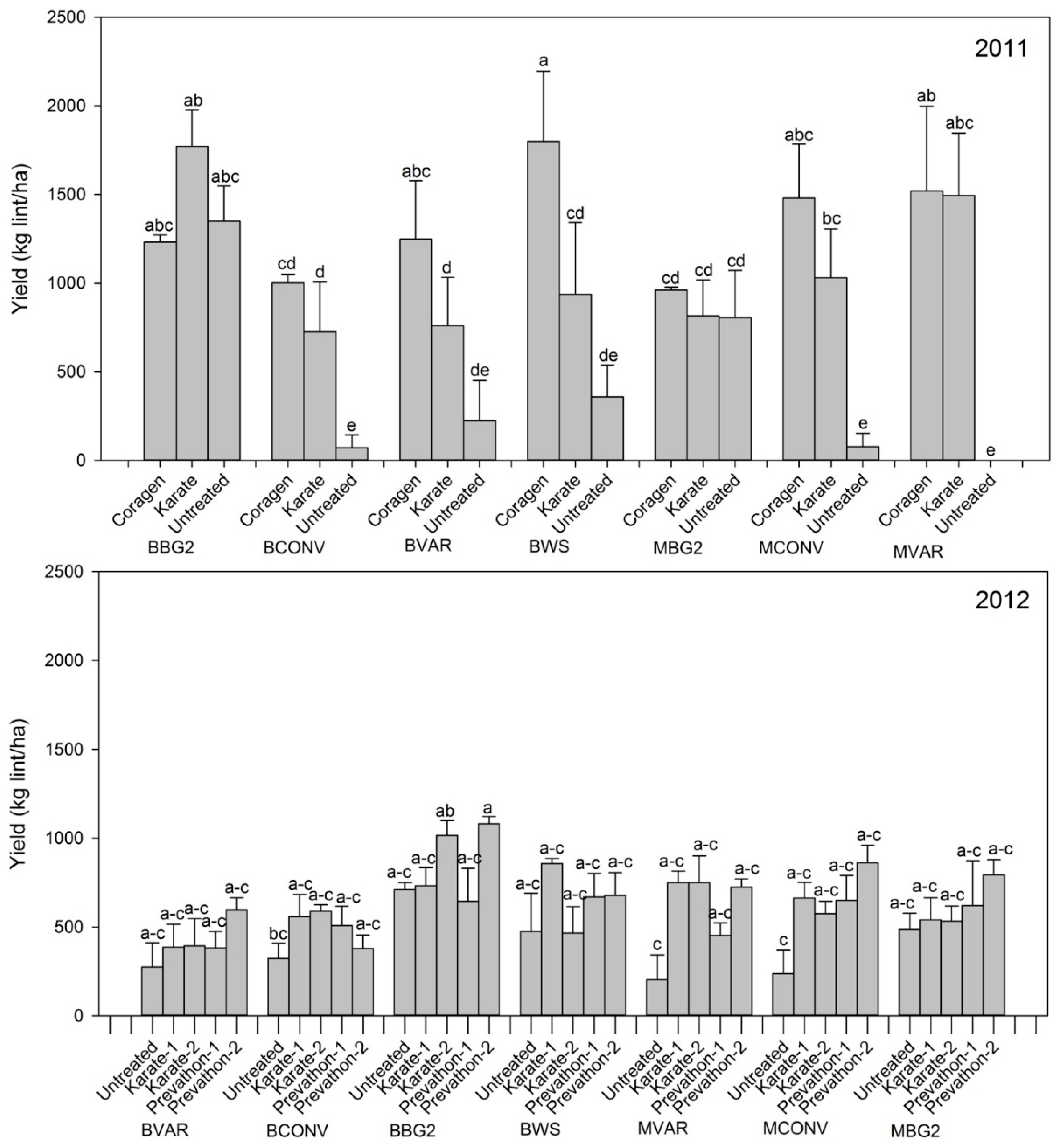
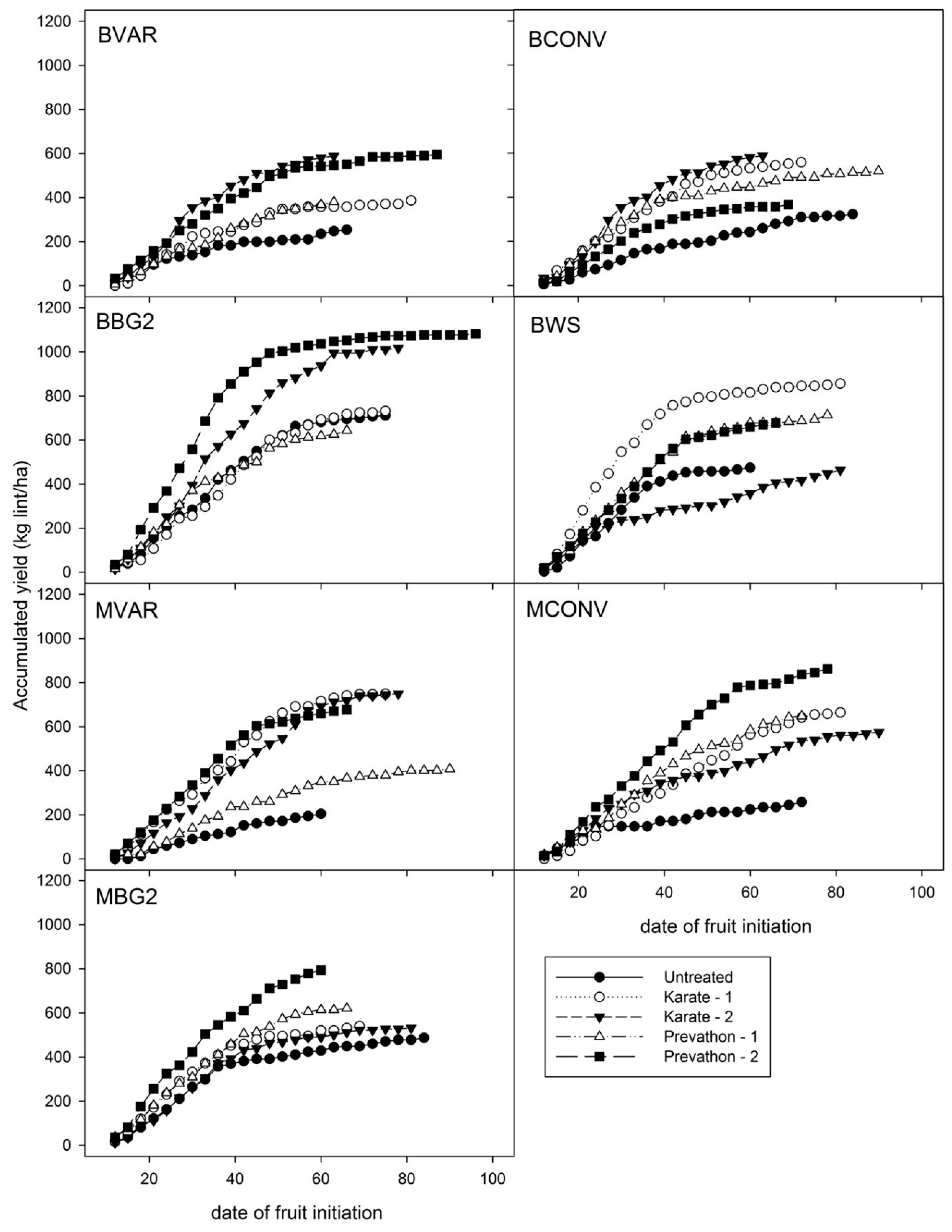
3.3. Overall Yield Response
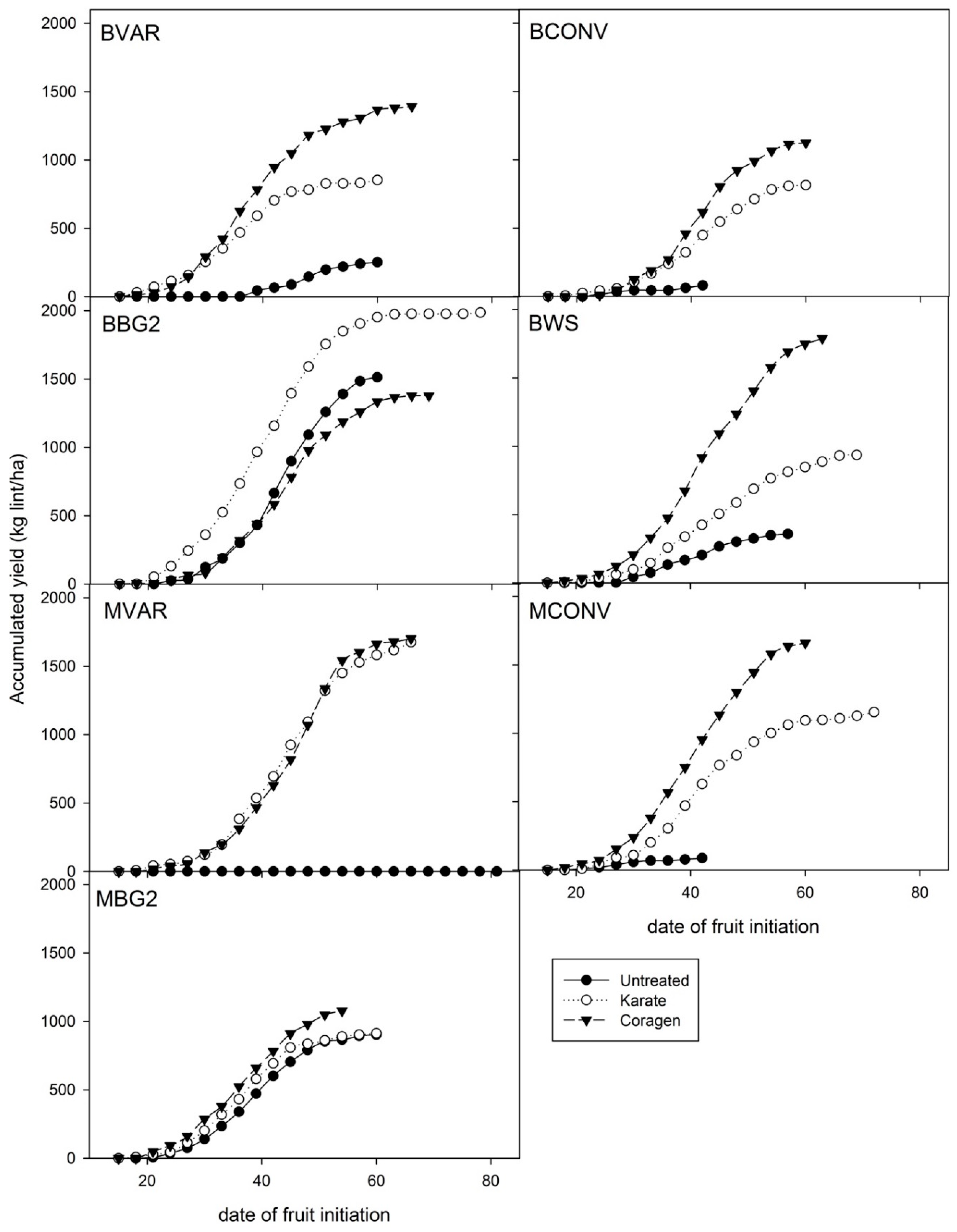
3.4. End of Seaon Box Maps of Harvested Fruit
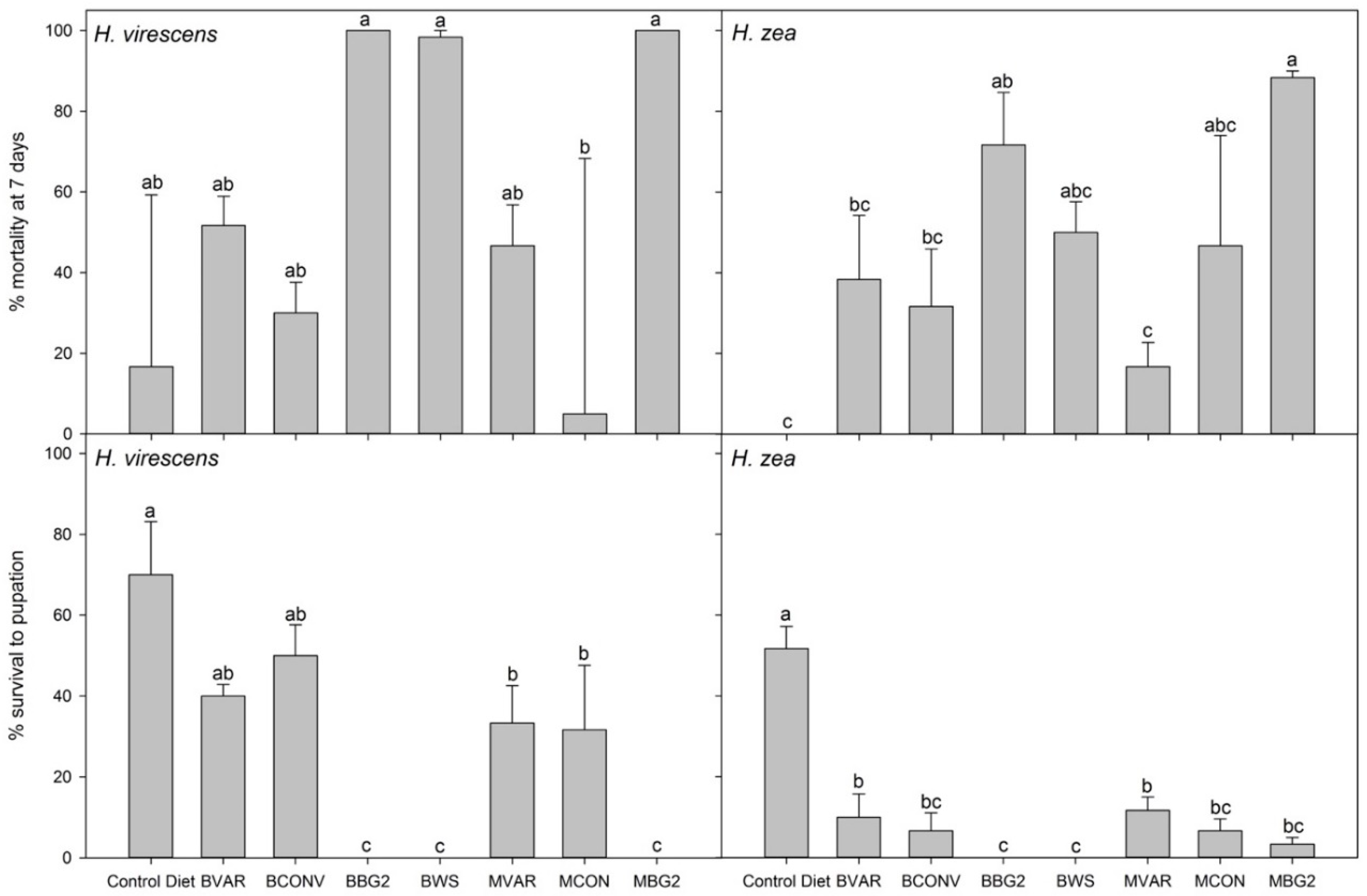
3.5. Bioassays of Cotton Plant Tissue
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luttrell, R.G.; Teague, T.G.; Brewer, M.J. Cotton insect pest management. In Cotton, 2nd ed.; Fang, D.D., Percey, R.G., Eds.; Agronomy Monograph 57; American Society of Agronomy: Madison, WI, USA; Crop Science Society: Fitchburg, WI, USA; Soil Science Society of America: Madison, WI, USA, 2015; pp. 509–546. [Google Scholar]
- Frisvold, G.B.; Reeves, J.M. The costs and benefits of refuge requirements: The case of Bt cotton. Ecol. Econ. 2008, 65, 87–97. [Google Scholar] [CrossRef]
- Luttrell, R.G.; Wan, L.; Knighten, K. Variation in susceptibility of noctuid (lepidoptera) larvae attacking cotton and soybean to purified endotoxin proteins and commercial formulations of Bacillus thuringiensis. J. Econ. Entomol. 1999, 92, 21–32. [Google Scholar] [CrossRef]
- MacIntosh, S.C.; Stone, T.B.; Sims, S.R.; Hunst, P.L.; Greenplate, J.T.; Marrone, P.G.; Perlak, F.J.; Fischoff, D.A.; Fuchs, R.L. Specificity and efficacy of purified Bacillus thuringiensis proteins against economically important insects. J. Invertebr. Pathol. 1990, 56, 258–266. [Google Scholar] [CrossRef]
- Stewart, S.D.; Adamczyk, J.J., Jr.; Knighten, K.S.; Davis, F.M. Impact of Bt cottons expressing one or two insecticidal proteins of Bacillus thuringiensis Berliner on growth and survival of noctuid (lepidoptera) larvae. J. Econ. Entomol. 2001, 94, 752–760. [Google Scholar] [CrossRef]
- Luttrell, R.G.; Jackson, R.E. Helicoverpa zea and Bt Cotton in the United States. GM Crops Food 2012, 3, 213–227. [Google Scholar] [CrossRef]
- Edge, J.M.; Benedict, J.H.; Carroll, J.P.; Reding, H.K. Bollgard cotton: An assessment of global economic, environmental, and social benefits. J. Cotton Sci. 2001, 5, 121–136. [Google Scholar]
- Jackson, R.E.; Bradley, J.R., Jr.; Van Duyn, J.W.; Gould, F. Comparative production of Helicoverpa zea (lepidoptera: Noctuidae) from transgenic cotton expressing either one or two Bacillus thuringiensis proteins with and without insecticide oversprays. J. Econ. Entomol. 2004, 97, 1719–1725. [Google Scholar] [CrossRef]
- Little, N.S.; Catchot, A.L.; Allen, K.C.; Gore, J.; Musser, F.R.; Cook, D.R.; Luttrell, R.G. Supplemental control with diamides for heliothines in Bt Cotton. Southwest. Entomol. 2017, 42, 15–26. [Google Scholar] [CrossRef]
- Naranjo, S.E. Impact of Bt transgenic cotton on integrated pest management. J. Agric. Food Chem. 2011, 59, 5842–5851. [Google Scholar] [CrossRef]
- Mascarenhas, V.J.; Luttrell, R.G. Combined effect of sublethal exposure to cotton expressing the endotoxin protein of Bacillus thuringiensis and natural enemies on survival of bollworm (lepidoptera: Noctuidae) larvae. Environ. Entomol. 1997, 26, 939–945. [Google Scholar] [CrossRef]
- Naranjo, S.E.; Ellsworth, P.C. Arthropod communities and transgenic cotton in the western United States: Implications for biological control. In Proceedings of the 1st International Symposium on Biological Control of Arthropods, Honolulu, HI, USA, 14–18 January 2002; pp. 284–291. [Google Scholar]
- Head, G.; Moar, W.; Eubanks, M.; Freeman, B.; Ruberson, J.; Hagerty, A.; Turnipseed, S. A multiyear, large-scale comparison of arthropod populations on commercially managed Bt and non-Bt cotton fields. Environ. Entomol. 2005, 34, 1257–1266. [Google Scholar] [CrossRef]
- Torres, J.B.; Ruberson, J.R. Spatial and temporal dynamics of oviposition behavior of bollworm and three of its predators in Bt and non-Bt cotton fields. Entomol. Exp. Appl. 2006, 120, 11–13. [Google Scholar] [CrossRef]
- Naranjo, S.E.; Luttrell, R.G. Cotton arthropod IPM. In Integrated Pest Management: Concepts, Tactics, Strategies and Case Studies; Ratcliffe, E.B., Hutchinson, W.D., Cancelado, R.E., Eds.; Cambridge University Press: New York, NY, USA, 2009; p. 529. [Google Scholar]
- Luttrell, R.G. Cotton pest management: Part 2. A US perspective. Annu. Rev. Entomol. 1994, 39, 527–542. [Google Scholar] [CrossRef]
- Lambert, A.L.; Bradley, J.R., Jr.; Van Duyn, J.W. Effects of natural enemy conservation and planting date on the susceptibility of Bt cotton to Helicoverpa zea in North Carolina. In Proceedings of the Beltwide Cotton Conferences, Nashville, TN, USA, 9–12 January 1996; pp. 931–935. [Google Scholar]
- Mahaffey, J.S.; Bradley, J.R., Jr.; Van Duyn, J.W. Bt cotton: Field performance in North Carolina under conditions of unusually high bollworm populations. In Proceedings of the Beltwide Cotton Conferences, San Antonio, TX, USA, 4–7 January 1995; pp. 795–798. [Google Scholar]
- Reisig, D.D.; Huseth, A.S.; Batcheler, J.S.; Aghaee, M.A.; Braswell, L.; Burrack, H.J.; Flanders, K.; Greene, J.K.; Herbert, D.A.; Jacobson, A.; et al. Long-term empirical and observational evidence of practical Helicoverpa zea resistance to cotton with pyramided Bt toxins. J. Econ. Entomol. 2018, 111, 1824–1833. [Google Scholar] [CrossRef]
- Williams, M.R. Cotton Insect Losses-2015. In Proceedings of the Beltwide Cotton Conferences, New Orleans, LA, USA, 5–7 January 2016; pp. 507–526. [Google Scholar]
- Little, N.S.; Luttrell, R.G.; Allen, K.C.; Perera, O.P.; Parys, K.A. Effectiveness of microbial and chemical insecticides for supplemental control of bollworm on Bt and non-Bt cottons. J. Econ. Entomol. 2017, 110, 1039–1051. [Google Scholar] [CrossRef]
- Bourland, F.M.; Jones, D.C. Registration of ‘UA48’ cotton cultivar. J. Plant Regist. 2011, 6, 15–18. [Google Scholar] [CrossRef]
- Meredith, W.R., Jr.; Nokes, W.S. Registration of MD 9ne and MD 25 high-fiber-quality germplasm lines of cotton. J. Plant Regist. 2011, 5, 202–206. [Google Scholar] [CrossRef]
- Blanco, C.A.; Portilla, M.; Abel, C.A.; Winters, H.; Ford, R.; Streett, D. Soybean flour and wheat germ proportions in artificial diet and their effect on the growth rates of tobacco budworm, Heliothis virescens. J. Insect Sci. 2009, 9, 1–9. [Google Scholar] [CrossRef]
- Allen, K.C.; Luttrell, R.G.; Parker, C.D., Jr. Influence of within-season densities of heliothines and tarnished plant bugs on variability in end-of-season cotton yield mapping. J. Cotton Sci. 2009, 133, 11–22. [Google Scholar]
- Jenkins, J.N.; McCarty, J.C., Jr. Useful Tools in Managing Cotton Production: End of Season Plant Maps; Bulletin 1024; Mississippi State University: Starkville, MS, USA, 1995. [Google Scholar]
- Allen, K.C.; Andrews, G.; Burdine, B.; Catchot, A.; Cook, D.; Dodds, D.; Gore, J.; Howell, M.; Jackson, R.; Larson, E.; et al. Insect Control Guide for Agronomic Crops; Mississippi State University Extension Service: Starkville, MS, USA, 2011; p. 104. [Google Scholar]
- Parker, C.D., Jr.; Luttrell, R.G. Interplant movement of Heliothis virescens (lepidoptera: Noctuidae) larvae in pure and mixed plantings of cotton with and without the expression of the CryIA(c) delta-endotoxin protein of Bacillus thuringiensis. J. Econ. Entomol. 1999, 92, 837–845. [Google Scholar] [CrossRef]
- Storer, N.P.; Peck, S.L.; Gould, F.; Van Duyn, J.W.; Kennedy, G.C. Spatial processes in the evolution of resistance in Helicoverpa zea (lepidoptera: Noctuidae) to Bt transgenic corn and cotton in a mixed agroecosystem: A biology-rich stochastic simulation model. J. Econ. Entomol. 2003, 96, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Danforth, D.M.; O’Leary, P.F. (Eds.) COTMAN Expert System 5.0. User’s Manual; Argricultural Experiment Station Bulletin; University of Arkansas: Fayetteville, AR, USA, 1998; p. 198. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Abbreviation | Trait | Maturity |
|---|---|---|---|
| ARK48 | BVAR | Non-Bt | Early |
| DP121 | BCONV | Non-Bt | Early |
| DP0912 | BBG2 | Bollgard II | Early |
| PHY375 | BWS | Widestrike | Early |
| MD25 | MVAR | Non-Bt | Full |
| DP174 | MCONV | Non-Bt | Full |
| DP1048 | MBG2 | Bollgard II | Full |
| Variety | Insecticide Commercial Name | Eggs/Plant 20 July 2011 | Eggs/Plant 25 July 2011 | Larvae/Plant 15 Aug 2011 |
|---|---|---|---|---|
| BBG2 | Coragen | 8.47 ± 1.142 | 0.67 ± 0.27 | 0.1 ± 0.072 cd |
| BBG2 | Karate | 10.27 ± 1.59 | 0.87 ± 0.256 | 0 ± 0 d |
| BBG2 | Untreated | 10.6 ± 1.45 | 0.8 ± 0.312 | 0 ± 0 d |
| BCONV | Coragen | 12.93 ± 0.954 | 1.07 ± 0.358 | 0.3 ± 0.107 b |
| BCONV | Karate | 13.27 ± 1.406 | 0.33 ± 0.159 | 0 ± 0 d |
| BCONV | Untreated | 11.33 ± 1.728 | 0.93 ± 0.345 | 0.3 ± 0.107 b |
| BVAR | Coragen | 12.4 ±1.279 | 1.33 ± 0.681 | 0.5 ± 0.129 a |
| BVAR | Karate | 13 ± 1.483 | 1.53 ± 0.533 | 0 ± 0 d |
| BVAR | Untreated | 11.13 ± 0.576 | 0.8 ± 0.296 | 0 ± 0 d |
| BWS | Coragen | 12.27 ± 1.66 | 0.73 ± 0.33 | 0.3 ± 0.107 b |
| BWS | Karate | 9.93 ± 1.248 | 0.93 ± 0.267 | 0 ± 0 d |
| BWS | Untreated | 8.57 ± 1.137 | 0.21 ± 0.114 | 0 ± 0 d |
| MBG2 | Coragen | 10.87 ± 0.827 | 0.73 ± 0.248 | 0.4 ± 0.111 ab |
| MBG2 | Karate | 11.4 ± 2.537 | 0.73 ± 0.228 | 0 ± 0 d |
| MBG2 | Untreated | 11.44 ± 1.228 | 0.31 ± 0.151 | 0.06 ± 0.043 d |
| MCOBV | Coragen | 12.47 ± 1.664 | 0.33 ± 0.159 | 0 ± 0 d |
| MCONV | Karate | 11.07 ± 1.631 | 0.67 ± 0.252 | 0 ± 0 d |
| MCONV | Untreated | 11.33 ± 0.95 | 0.6 ± 0.163 | 0.27 ± 0.108 bc |
| MVAR | Coragen | 11.53 ± 1.129 | 0.33 ± 0.211 | 0 ± 0 d |
| MVAR | Karate | 11.13 ± 1.897 | 0.4 ± 0.131 | 0 ± 0 d |
| MVAR | Untreated | 12 ± 1.47 | 0.67 ± 0.232 | 0.07 ± 0.067 d |
| Variety | Insecticide Commercial Name | Eggs/Plant 23 July 2012 | Eggs/Plant 6 August 2012 | Eggs/Plant 13 August 2012 |
|---|---|---|---|---|
| BBG2 BBG2 | Karate-1 | 0.13 ± 0.063 | 0 | |
| BBG2 | Karate-2 | 0 | ||
| BBG2 | Prevathon-1 | 0.13 ± 0.063 | 0 | |
| BBG2 | Prevathon-2 | 0 | ||
| BBG2 | Untreated | 0.83 ± 0.017 ab | 0.03 ± 0.033 | 0 |
| BCONV | Karate-1 | 0.2 ± 0.074 | 0 | |
| BCONV | Karate-2 | 0 | ||
| BCONV | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| BCONV | Prevathon-2 | 0 | ||
| BCONV | Untreated | 0.52 ± 0.1 b | 0.1 ± 0.056 | 0 |
| BVAR | Karate-1 | 0.1 ± 0.056 | 0 | |
| BVAR | Karate-2 | 0 | ||
| BVAR | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| BVAR | Prevathon-2 | 0 | ||
| BVAR | Untreated | 0.88 ± 0.1 ab | 0.1 ± 0.056 | 0 |
| BWS | Karate-1 | 0.1 ± 0.056 | 0 | |
| BWS | Karate-2 | 0 | ||
| BWS | Prevathon-1 | 0.2 ± 0.074 | 0 | |
| BWS | Prevathon-2 | 0.05 ± 0.05 | ||
| BWS | Untreated | 0.72 ± 0.017 ab | 0.13 ± 0.063 | 0 |
| MBG2 | Karate-1 | 0.17 ± 0.069 | 0 | |
| MBG2 | Karate-2 | 0 | ||
| MBG2 | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| MBG2 | Prevathon-2 | 0 | ||
| MBG2 | Untreated | 0.92 ± 0.033 a | 0.13 ± 0.063 | 0 |
| MCONV | Karate-1 | 0.1 ± 0.056 | 0 | |
| MCONV | Karate-2 | 0 | ||
| MCONV | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| MCONV | Prevathon-2 | 0 | ||
| MCONV | Untreated | 0.85 ± 0.083 ab | 0.07 ± 0.046 | 0 |
| MVAR | Karate-1 | 0.07 ± 0.046 | 0 | |
| MVAR | Karate-2 | 0 | ||
| MVAR | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| MVAR | Prevathon-2 | 0 | ||
| MVAR | Untreated | 0.7 ± 0.1 ab | 0.13 ± 0.063 | 0 |
| Variety | Insecticide | Larvae/Plant 23 July 2012 | Larvae/Plant 6 August 2012 | Larvae/Plant 13 August 2012 |
|---|---|---|---|---|
| BBG2 | Karate-1 | 0.2 ± 0.074 | 0 | |
| BBG2 | Karate-2 | 0 | ||
| BBG2 | Prevathon-1 | 0.2 ± 0.074 | 0 | |
| BBG2 | Prevathon-2 | 0 | ||
| BBG2 | Untreated | 0.02 ± 0.017 | 0.37 ± 0.089 | 0 |
| BCONV | Karate-1 | 0.13 ± 0.063 | 0 | |
| BCONV | Karate-2 | 0 | ||
| BCONV | Prevathon-1 | 0.23 ± 0.079 | 0 | |
| BCONV | Prevathon-2 | 0 | ||
| BCONV | Untreated | 0.1 ± 0.039 | 0.27 ± 0.082 | 0 |
| BVAR | Karate-1 | 0.27 ± 0.082 | 0 | |
| BVAR | Karate-2 | 0 | ||
| BVAR | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| BVAR | Prevathon-2 | 0 | ||
| BVAR | Untreated | 0.1 ± 0.039 | 0.23 ± 0.079 | 0 |
| BWS | Karate-1 | 0.27 ± 0.082 | 0 | |
| BWS | Karate-2 | 0 | ||
| BWS | Prevathon-1 | 0.23 ± 0.079 | 0 | |
| BWS | Prevathon-2 | 0 | ||
| BWS | Untreated | 0.02 ± 0.017 | 0.27 ± 0.082 | 0 |
| MBG2 | Karate-1 | 0.13 ± 0.063 | 0 | |
| MBG2 | Karate-2 | 0 | ||
| MBG2 | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| MBG2 | Prevathon-2 | 0 | ||
| MBG2 | Untreated | 0.1 ± 0.023 | 0.23 ± 0.079 | 0 |
| MCONV | Karate-1 | 0.27 ± 0.082 | 0 | |
| MCONV | Karate-2 | 0 | ||
| MCONV | Prevathon-1 | 0.2 ± 0.074 | 0 | |
| MCONV | Prevathon-2 | 0 | ||
| MCONV | Untreated | 0.1 ± 0.036 | 0.3 ± 0.085 | 0.05 ± 0.05 |
| MVAR | Karate-1 | 0.2 ± 0.074 | 0 | |
| MVAR | Karate-2 | 0 | ||
| MVAR | Prevathon-1 | 0.17 ± 0.069 | 0 | |
| MVAR | Prevathon-2 | 0 | ||
| MVAR | Untreated | 0.1 ± 0.017 | 0.2 ± 0.074 | 0 |
| Variety | Insecticide Commercial Name | Uppermost Node 13 July 2011 | Uppermost Node 25 July 2011 | Uppermost Node 1 August 2011 | Nodes with Open Bolls at Harvest 6 October 2011 |
|---|---|---|---|---|---|
| BBG2 | Coragen | 16.5 ± 0.532 | 18.9 ± 0.446 | 21.2 ± 0.901 | 16.3 ± 0.622 a |
| BBG2 | Karate | 15.6 ± 0.716 | 21.3 ± 0.583 | 18.9 ± 0.768 | 15.6 ± 0.505 a |
| BBG2 | Untreated | 15.3 ± 0.485 | 20.9 ± 0.808 | 19.9 ± 0.892 | 14.4 ± 1.025 a |
| BCONV | Coragen | 16.1 ± 0.736 | 20 ± 0.64 | 20.9 ± 0.867 | 14.4 ± 0.631 a |
| BCONV | Karate | 15.6 ± 0.844 | 21.4 ± 0.838 | 20 ± 0.552 | 14.2 ± 0.813 a |
| BCONV | Untreated | 14.1 ± 0.33 | 20.1 ± 0.511 | 22.5 ± 0.761 | 12.9 ± 1.429 a |
| BVAR | Coragen | 18.3 ± 0.79 | 19 ± 0.447 | 20 ± 0.586 | 12.9 ± 0.808 a |
| BVAR | Karate | 16 ± 0.845 | 21.9 ± 0.621 | 20.2 ± 0.649 | 8.3 ± 1.899 b |
| BVAR | Untreated | 15.1 ± 0.876 | 20.5 ± 0.91 | 20.8 ± 0.932 | 9.4 ± 3.894 ab |
| BWS | Coragen | 15.8 ± 0.829 | 18.9 ± 0.597 | 19.7 ± 0.929 | 15.2 ± 0.96 a |
| BWS | Karate | 15.8 ± 0.649 | 21.7 ± 0.74 | 21.9 ± 1.016 | 14.6 ± 1.675 a |
| BWS | Untreated | 15 ± 0.61 | 20.5 ± 0.85 | 19.6 ± 0.918 | 13.1 ± 0.589 a |
| MBG2 | Coragen | 16.2 ± 0.571 | 18.6 ± 0.638 | 19.2 ± 0.712 | 12.5 ± 0.682 a |
| MBG2 | Karate | 14.3 ± 0.591 | 20.1 ± 0.904 | 20.8 ± 0.656 | 11.8 ± 1.244 a |
| MBG2 | Untreated | 13.9 ± 0.403 | 20.4 ± 0.632 | 20.1 ± 0.844 | 9.8 ± 1.235 ab |
| MCONV | Coragen | 17.4 ± 0.809 | 18.9 ± 0.425 | 20.6 ± 0.755 | 15.6 ± 0.521 a |
| MCONV | Karate | 16.3 ± 0.708 | 20.9 ± 0.628 | 20.7 ± 0.831 | 14.2 ± 1.028 a |
| MCONV | Untreated | 13.7 ± 0.565 | 20.7 ± 0.785 | 20.3 ± 0.41 | 6 ± 2.471 b |
| MVAR | Coragen | 16.1 ± 0.593 | 19.5 ± 0.376 | 19.7 ± 0.773 | 15.4 ± 0.618 a |
| MVAR | Karate | 14.4 ± 0.423 | 20.5 ± 0.965 | 20.2 ± 0.751 | 15.8 ± 0.757 a |
| MVAR | Untreated | 14.2 ± 0.418 | 21 ± 0.64 | 20.9 ± 0.733 | 2.5 ± 2.5 b |
| Variety | Insecticide Commercial Name | Fruit/Plant 13 July 2011 | Fruit/Plant 25 July 2011 | Fruit/Plant1 August 2011 | Fruit/Plant Harvest 6 October 2011 |
|---|---|---|---|---|---|
| BBG2 | Coragen | 7.8 ± 0.656 | 9.5 ± 0.456 | 9.8 ± 0.712 a | 5.7 ± 0.597 |
| BBG2 | Karate | 7.7 ± 0.621 | 10.3 ± 0.475 | 7.5 ± 0.878 ab | 7.5 ± 0.904 |
| BBG2 | Untreated | 6.8 ± 0.368 | 10.7 ± 0.422 | 7.7 ± 0.621 ab | 5.2 ± 0.654 |
| BCONV | Coragen | 5.7 ± 0.433 | 8.9 ± 0.749 | 8.5 ± 0.925 ab | 4.5 ± 0.376 |
| BCONV | Karate | 6.2 ± 0.699 | 8.7 ± 0.539 | 8.7 ± 0.771 ab | 5.1 ± 0.691 |
| BCONV | Untreated | 5.8 ± 0.327 | 9.5 ± 0.446 | 10.5 ± 0.661 a | 0.3 ± 0.141 |
| BVAR | Coragen | 7.4 ± 0.838 | 9.7 ± 0.54 | 8.7 ± 0.747 ab | 6 ± 0.75 |
| BVAR | Karate | 7.2 ± 0.705 | 10.3 ± 0.396 | 9.5 ± 0.533 ab | 3.4 ± 0.872 |
| BVAR | Untreated | 7.2 ± 0.547 | 9.5 ± 0.568 | 8.7 ± 0.959 ab | 2.2 ± 1.158 |
| BWS | Coragen | 6.3 ± 0.566 | 9.1 ± 0.452 | 8.3 ± 0.939 ab | 6 ± 0.78 |
| BWS | Karate | 6.9 ± 0.53 | 10.5 ± 0.624 | 11.1 ± 0.646 a | 6.5 ± 0.976 |
| BWS | Untreated | 7.4 ± 0.58 | 10.1 ± 0.467 | 6.6 ± 0.894 b | 3.6 ± 0.456 |
| MBG2 | Coragen | 6.1 ± 0.51 | 9.1 ± 0.636 | 7.3 ± 0.773 ab | 4.9 ± 0.401 |
| MBG2 | Karate | 6 ± 0.425 | 9.8 ± 0.536 | 9.1 ± 0.456 ab | 4.8 ± 0.716 |
| MBG2 | Untreated | 6.3 ± 0.546 | 9.5 ± 0.465 | 9.3 ± 0.8 ab | 1.8 ± 0.393 |
| MCONV | Coragen | 7.9 ± 0.827 | 9.4 ± 0.308 | 8.7 ± 0.803 ab | 7.1 ± 1.19 |
| MCONV | Karate | 7.1 ± 0.643 | 10.4 ± 0.515 | 5.8 ± 0.788 b | 5.1 ± 0.619 |
| MCONV | Untreated | 6.7 ± 0.419 | 9.5 ± 0.467 | 8.9 ± 0.473 ab | 3.3 ± 1.521 |
| MVAR | Coragen | 6.8 ± 0.67 | 8.9 ± 0.496 | 7.3 ± 0.813 ab | 6.2 ± 0.757 |
| MVAR | Karate | 5.1 ± 0.452 | 10.5 ± 0.716 | 8.1 ± 0.978 ab | 6.1 ± 1.106 |
| MVAR | Untreated | 7.0 ± 0.535 | 10.2 ± 0.36 | 9.4 ± 0.702 ab | 1.3 ± 1.25 |
| Variety | Insecticide Commercial Name | Uppermost Node 17 July 2012 | Uppermost Node 3 August 2012 | Uppermost Node 15 August 2012 | Nodes with Open Bolls at Harvest 20 October 2012 |
|---|---|---|---|---|---|
| BBG2 | Karate-1 | 16.7 ± 0.667 | 23.3 ± 9.733 | 21 ± 6.533 | 8.3 ± 2.675 |
| BBG2 | Karate-2 | 23.3 ± 8.4 | 13.3 ± 6.017 | ||
| BBG2 | Prevathon-1 | 16.7 ± 1.202 | 25 ± 11.9 | 17.3 ± 5.733 | 9.3 ± 4.122 |
| BBG2 | Prevathon-2 | 21.7 ± 8 | 14.3 ± 6.178 | ||
| BBG2 | Untreated | 16.3 ± 1.453 | 23 ± 11.333 | 21 ± 6.8 | 10 ± 3.381 |
| BCONV | Karate-1 | 16.3 ± 1.333 | 24 ± 11.467 | 17.7 ± 4.8 | 8.7 ± 3.333 |
| BCONV | Karate-2 | 12.3 ± 3.2 | 8.3 ± 3.344 | ||
| BCONV | Prevathon-1 | 20.7 ± 3.712 | 24 ± 11.133 | 17.3 ± 6.133 | 6 ± 3.156 |
| BCONV | Prevathon-2 | 19 ± 7.067 | 6.7 ± 2 | ||
| BCONV | Untreated | 18 ± 1.528 | 24 ± 11.733 | 4.3 ± 1.2 | 8 ± 1.869 |
| BVAR | Karate-1 | 15 ± 1 | 23.7 ± 11.267 | 18.7 ± 6.367 | 6 ± 2.293 |
| BVAR | Karate-2 | 16.7 ± 4.6 | 8 ± 2.556 | ||
| BVAR | Prevathon-1 | 19.7 ± 3.18 | 26.7 ± 11.533 | 18.7 ± 6.511 | 7 ± 2.378 |
| BVAR | Prevathon-2 | 18.3 ± 5.467 | 10.7 ± 4.289 | ||
| BVAR | Untreated | 17 ± 1.155 | 26.7 ± 12.333 | 12.3 ± 4.133 | 5 ± 1.327 |
| BWS | Karate-1 | 17.7 ± 0.333 | 23 ± 10.733 | 20.7 ± 6.8 | 11 ± 4.644 |
| BWS | Karate-2 | 18.7 ± 5.867 | 9.7 ± 3.733 | ||
| BWS | Prevathon-1 | 19.7 ± 2.028 | 25.3 ± 11.067 | 20 ± 6.933 | 9.7 ± 2.808 |
| BWS | Prevathon-2 | 15.7 ± 5 | 9.7 ± 3.128 | ||
| BWS | Untreated | 19 ± 1.528 | 26 ± 11.933 | 14 ± 4.333 | 8 ± 2 |
| MBG2 | Karate-1 | 16 ± 1 | 22.3 ± 11 | 15 ± 4.267 | 7.7 ± 2.333 |
| MBG2 | Karate-2 | 18.3 ± 4.8 | 9 ± 3.075 | ||
| MBG2 | Prevathon-1 | 15.7 ± 0.667 | 26.7 ± 12.733 | 18.3 ± 6.533 | 8.3 ± 2.944 |
| MBG2 | Prevathon-2 | 19.3 ± 8.067 | 10 ± 3.333 | ||
| MBG2 | Untreated | 16.7 ± 2.906 | 26.5 ± 12.8 | 17.7 ± 5.4 | 9.3 ± 2.667 |
| MCONV | Karate-1 | 15.7 ± 0.333 | 23.3 ± 11 | 18.5 ± 3.7 | 10 ± 3.905 |
| MCONV | Karate-2 | 14 ± 4.667 | 12 ± 3.698 | ||
| MCONV | Prevathon-1 | 18.7 ± 2.028 | 25.3 ± 12.6 | 19 ± 5.8 | 8 ± 3.5 |
| MCONV | Prevathon-2 | 22 ± 9 | 11 ± 3.644 | ||
| MCONV | Untreated | 17 ± 1 | 22.5 ± 11.3 | 12.3 ± 3.067 | 3.3 ± 0.689 |
| MVAR | Karate-1 | 14.3 ± 0.667 | 24 ± 11.667 | 15 ± 4.867 | 8.7 ± 3.133 |
| MVAR | Karate-2 | 16.7 ± 5.667 | 12 ± 4.8 | ||
| MVAR | Prevathon-1 | 18 ± 2.082 | 27.7 ± 13.4 | 16 ± 6.667 | 6 ± 2.33 |
| MVAR | Prevathon-2 | 20 ± 7.733 | 12.3 ± 3.3 | ||
| MVAR | Untreated | 17.3 ± 1.764 | 27 ± 13.2 | 9.3 ± 2.267 | 4 ± 1.25 |
| Variety | Insecticide Commercial Name | Fruit/Plant 7 July 2012 | Fruit/Plant 3 August 2012 | Fruit/Plant 15 August 2012 | Fruit/Plantat Harvest 20 October 2012 |
|---|---|---|---|---|---|
| BBG2 | Karate-1 | 9.7 ± 0.24 | 9.7 ± 0.822 | 6.5 ± 0.577 | 2.7 ± 2.028 |
| BBG2 | Karate-2 | 8.4 ± 0.333 | 6 ± 1.856 | ||
| BBG2 | Prevathon-1 | 9.5 ± 0.291 | 11.9 ± 1 | 5.7 ± 3.18 | 4.1 ± 1.856 |
| BBG2 | Prevathon-2 | 8 ± 1.202 | 6.2 ± 0.882 | ||
| BBG2 | Untreated | 8.9 ± 0.742 | 11.3 ± 3.055 | 6.8 ± 3.606 | 3.4 ± 0.577 |
| BCONV | Karate-1 | 8.5 ± 0.467 | 11.5 ± 1.732 | 4.8 ± 1.453 | 3.3 ± 0.882 |
| BCONV | Karate-2 | 3.2 ± 1.333 | 3.3 ± 0.822 | ||
| BCONV | Prevathon-1 | 9.7 ± 0.882 | 11.1 ± 1.155 | 6.1 ± 3.18 | 3.2 ± 1 |
| BCONV | Prevathon-2 | 7.1 ± 1.528 | 2 ± 1.667 | ||
| BCONV | Untreated | 9.1 ± 0.467 | 11.7 ± 1.155 | 1.2 ± 0.667 | 1.9 ± 1 |
| BVAR | Karate-1 | 8.2 + 0.2 | 11.3 ± 0.882 | 6.4 ± 3.844 | 2.3 ± 1.528 |
| BVAR | Karate-2 | 4.6 ± 3.712 | 2.6 ± 1 | ||
| BVAR | Prevathon-1 | 10 ± 0.4 | 11.5 ± 0.882 | 5.6 ± 2.848 | 2.4 ± 1.732 |
| BVAR | Prevathon-2 | 5.5 ± 1.667 | 4.3 ± 2.848 | ||
| BVAR | Untreated | 8.2 ± 0.6 | 12.3 ± 0.667 | 4.1 ± 2.028 | 1.3 ± 0.577 |
| BWS | Karate-1 | 9.3 ± 0.521 | 10.7 ± 0.577 | 6.8 ± 1.202 | 4.6 ± 1 |
| BWS | Karate-2 | 5.9 ± 1.202 | 3.7 ± 2.186 | ||
| BWS | Prevathon-1 | 11 ± 0.917 | 11.1 ± 4.177 | 6.9 ± 1.528 | 2.8 ± 0.882 |
| BWS | Prevathon-2 | 5 ± 1.202 | 3.1 ± 1.453 | ||
| BWS | Untreated | 8.5 ± 0.667 | 11.9 ± 1.155 | 4.3 ± 2.517 | 2 ± 2.887 |
| MBG2 | Karate-1 | 8.6 ± 0.4 | 11 ± 1.667 | 4.3 ± 2.646 | 2.3 ± 2.333 |
| MBG2 | Karate-2 | 4.8 ± 1.453 | 3.1 ± 1.528 | ||
| MBG2 | Prevathon-1 | 8.8 ± 1 | 12.7 ± 2.404 | 6.5 ± 3.283 | 2.9 ± 3.18 |
| MBG2 | Prevathon-2 | 8.1 ± 3.18 | 3.3 ± 0.577 | ||
| MBG2 | Untreated | 7.9 ± 0.811 | 12.8 ± 2.5 | 5.4 ± 2.333 | 2.7 ± 1.764 |
| MCONV | Karate-1 | 9.6 ± 0.757 | 11 ± 1.202 | 3.7 ± 3.5 | 3.9 ± 1.155 |
| MCONV | Karate-2 | 4.7 ± 4.583 | 3.7 ± 0.577 | ||
| MCONV | Prevathon-1 | 10.3 ± 0.267 | 12.6 ± 2.404 | 5.8 ± 3.215 | 3.5 ± 2.082 |
| MCONV | Prevathon-2 | 9 ± 2.082 | 3.6 ± 2.082 | ||
| MCONV | Untreated | 8.1 ± 0.1 | 11.3 ± 1.5 | 3.1 ± 5.044 | 0.7 ± 0.333 |
| MVAR | Karate-1 | 7.5 ± 0.406 | 11.7 ± 0.577 | 4.9 ± 3.464 | 3.1 ± 2.728 |
| MVAR | Karate-2 | 5.7 ± 3.93 | 4.8 ± 1 | ||
| MVAR | Prevathon-1 | 10.1 ± 0.636 | 13.4 ± 1.453 | 6.7 ± 3.055 | 2.3 ± 1.732 |
| MVAR | Prevathon-2 | 7.7 ± 1.155 | 3.3 ± 1.856 | ||
| MVAR | Untreated | 8.7 ± 0.353 | 13.2 ± 1.732 | 2.3 ± 2.2.963 | 1.3 ± 1 |
| Variety | Insecticide Commercial Name | Average Days after Planting | Average Lint Weight (g) |
|---|---|---|---|
| BBG2 | Coragen | 46.3 ± 0.883 a | 45.8 ± 0.892 ab |
| BBG2 | Karate | 41.7 ± 1.003 a–c | 41.2 ± 1.026 a–e |
| BBG2 | Untreated | 45 ± 1.06 a | 44.5 ± 1.265 a–c |
| BCONV | Coragen | 38.5 ± 2.434 bc | 38.4 ± 2.662 c–e |
| BCONV | Karate | 41.9 ± 1.971 ab | 41.3 ± 1.921 a–e |
| BCONV | Untreated | 32.4 ± 0cd | 32.7 ± 0 ef |
| BVAR | Coragen | 38.5 ± 2.434 bc | 38.4 ± 2.662 c–e |
| BVAR | Karate | 41.9 ± 1.971 ab | 41.3 ± 1.921 a–e |
| BVAR | Untreated | --- | --- |
| BWS | Coragen | 46.6 ± 1.318 a | 46.2 ± 1.565 ab |
| BWS | Karate | 42.4 ± 4.27 ab | 42.4 ± 3.522 a–d |
| BWS | Untreated | 41.5 ± 1.583 a–c | 41.4 ± 0.942 a–e |
| MBG2 | Coragen | 37.8 ± 3.564 bc | 37.6 ± 3.755 de |
| MBG2 | Karate | 37.7 ± s.461 bc | 38 ± 2.133 c–e |
| MBG2 | Untreated | 40.7 ± 0.452 a–c | 40.2 ± 0.373 b–e |
| MCONV | Coragen | 42.2 ± 1.249 ab | 41.6 ± 1.254 a–e |
| MCONV | Karate | 42.9 ± 3.53 ab | 41.9 ± 3.504 |
| MCONV | Untreated | 27.2 ± 3.167 d | 27.2 ± 3.219 |
| MVAR | Coragen | 47.2 ± 2.318 | 46.9 ± 2.648 |
| MVAR | Karate | 45.3 ± 0.455 a | 45.2 ± 0.293 ab |
| MVAR | Untreated | --- | --- |
| Variety | Insecticide Commercial Name | Average Days after Planting | Average Lint Weight (g) |
|---|---|---|---|
| BBG2 | Karate-1 | 38.7 ± 3.425 | 37.8 ± 3.671 |
| BBG2 | Karate-2 | 37 ± 1 | 37.4 ± 0.898 |
| BBG2 | Prevathon-1 | 34.1 ± 3.124 | 34.5 ± 3.43 |
| BBG2 | Prevathon-2 | 32.8 ± 2.986 | 32.5 ± 2.952 |
| BBG2 | Untreated | 36.9 ± 1.795 | 35.4 ± 0.849 |
| BCONV | Karate-1 | 33.7 ± 3.433 | 33.5 ± 3.52 |
| BCONV | Karate-2 | 31.8 ± 0.556 | 32 ± 0.655 |
| BCONV | Prevathon-1 | 31.8 ± 6.407 | 31.5 ± 6.078 |
| BCONV | Prevathon-2 | 29.5 ± 5.378 | 29.1 ± 5.115 |
| BCONV | Untreated | 43.1 ± 10.176 | 42.6 ± 10.584 |
| BVAR | Karate-1 | 39.3 ± 5.677 | 38.6 ± 5.732 |
| BVAR | Karate-2 | 35.9 ± 4.619 | 35.2 ± 5.774 |
| BVAR | Prevathon-1 | 35.6 ± 4.786 | 35.1 ± 4.332 |
| BVAR | Prevathon-2 | 33 ± 5.241 | 32.5 ± 5.841 |
| BVAR | Untreated | 32.4 ± 6.496 | 33.1 ± 7.582 |
| BWS | Karate-1 | 32.3 ± 4.149 | 31.9 ± 4.842 |
| BWS | Karate-2 | 39.1 ± 3.703 | 39.8 ± 4.285 |
| BWS | Prevathon-1 | 30.2 ± 4.294 | 29.9 ± 4.365 |
| BWS | Prevathon-2 | 30.8 ± 4.264 | 30.9 ± 4.2 |
| BWS | Untreated | 28.9 ± 2.24 | 28.7 ± 2.099 |
| MBG2 | Karate-1 | 31.4 ± 1.613 | 31.3 ± 1.784 |
| MBG2 | Karate-2 | 33.8 ± 2.671 | 33.9 ± 2.213 |
| MBG2 | Prevathon-1 | 31.4 ± 2.658 | 31 ± 1.909 |
| MBG2 | Prevathon-2 | 32.1 ± 2.258 | 31.9 ± 2.289 |
| MBG2 | Untreated | 36.8 ± 5.751 | 36.1 ± 5.937 |
| MCONV | Karate-1 | 45.2 ± 1.907 | 44.3 ± 2.163 |
| MCONV | Karate-2 | 42.1 ± 5.995 | 43.1 ± 7.114 |
| MCONV | Prevathon-1 | 39.6 ± 5.701 | 39.3 ± 5.772 |
| MCONV | Prevathon-2 | 37.7 ± 5.803 | 37 ± 5.167 |
| MCONV | Untreated | 26.1 ± 5.798 | 26.4 ± 5.557 |
| MVAR | Karate-1 | 36 ± 1.971 | 35.7 ± 2.025 |
| MVAR | Karate-2 | 40.2 ± 2.063 | 39.3 ± 1.964 |
| MVAR | Prevathon-1 | 45.5 ± 7.128 | 45.7 ± 7.302 |
| MVAR | Prevathon-2 | 37.2 ± 1.503 | 36.4 ± 1.672 |
| MVAR | Untreated | 37.4 ± 4.475 | 38.3 ± 5.058 |
| 2011 | ||||
| 13 July 2011 | 25 July 2011 | 1 August 2011 | 6 October 2011 (Harvest) | |
| Untreated Bt | 6.8(0.318) | 10.1(0.346) | 7.9(0.784) | 3.5(0.982) ab |
| Insecticide Bt | 6.8(0.327) | 9.7(0.243) | 8.9(0.593) | 5.9(0.416) a |
| Untreated non Bt | 6.7(0.309) | 9.7(0.175) | 9.4(0.403) | 1.8(0.639) b |
| Insecticide non Bt | 6.7(0.332) | 9.6(0.26) | 8.2(0.403) | 5.4(0.409) a |
| 2012 | ||||
| 17 July 2012 | 3 August 2012 | 15 August 2012 | 20 October 2012 (Harvest) | |
| Untreated Bt | 8.4(0.291) | 12(0.436) | 5.5(0.723) ab | 2.7(0.404) ab |
| Insecticide Bt | 9.5(0.348) | 11.2(0.42) | 6.4(0.385) a | 3.7(0.367) a |
| Untreated non Bt | 8.5(0.232) | 12.1(0.413) | 2.7(0.614) b | 1.3(0.245) b |
| Insecticide non Bt | 9.2(0.364) | 11.8(0.292) | 5.8(0.372) a | 3.2(0.192) a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allen, K.C.; Luttrell, R.G.; Little, N.S.; Parys, K.A.; Perera, O.P. Response of Bt and Non-Bt Cottons to High Infestations of Bollworm (Helicoverpa zea Boddie) and Tobacco Budworm (Heliothis virescens (F.)) under Sprayed and Unsprayed Conditions. Agronomy 2019, 9, 759. https://doi.org/10.3390/agronomy9110759
Allen KC, Luttrell RG, Little NS, Parys KA, Perera OP. Response of Bt and Non-Bt Cottons to High Infestations of Bollworm (Helicoverpa zea Boddie) and Tobacco Budworm (Heliothis virescens (F.)) under Sprayed and Unsprayed Conditions. Agronomy. 2019; 9(11):759. https://doi.org/10.3390/agronomy9110759
Chicago/Turabian StyleAllen, Kerry C., Randall G. Luttrell, Nathan S. Little, Katherine A. Parys, and Omaththage P. Perera. 2019. "Response of Bt and Non-Bt Cottons to High Infestations of Bollworm (Helicoverpa zea Boddie) and Tobacco Budworm (Heliothis virescens (F.)) under Sprayed and Unsprayed Conditions" Agronomy 9, no. 11: 759. https://doi.org/10.3390/agronomy9110759