
Biological Control of Fall Armyworm, Spodoptera frugiperda
by
 , , , ,
, , , ,
Arzlan Abbas
1,† ,
,
Farman Ullah
2,3,† ,
,
Muhammad Hafeez
4,
Xiao Han
1,
Muhammad Zulqar Nain Dara
1,
Hina Gul
5 and
Chen Ri Zhao
1,* 1
College of Plant Protection, Jilin Agricultural University, Changchun 130118, China
2
Department of Plant Biosecurity, College of Plant Protection, China Agricultural University, Beijing 100193, China
3
Key Laboratory of Surveillance and Management for Plant Quarantine Pests, Ministry of Agriculture and Rural Affairs, Beijing 100193, China
4
State Key Laboratory for Managing Biotic and Chemical Threats to the Quality and Safety of Agro-Products, Institute of Plant Protection and Microbiology, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
5
MARA Key Laboratory of Pest Monitoring and Green Management, Department of Entomology, College of Plant Protection, China Agricultural University, Beijing 100193, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2022, 12(11), 2704; https://doi.org/10.3390/agronomy12112704
Submission received: 26 September 2022
/
Revised: 15 October 2022
/
Accepted: 18 October 2022
/
Published: 31 October 2022
(This article belongs to the Special Issue Ecological Management of Pests)
Abstract
:The fall armyworm (FAW), Spodoptera frugiperda, is one of the most important invasive pests worldwide, resulting in considerable losses in host crops. FAW comprises two genetic strains, such as the “rice strain”, which prefers rice and other grass species, and the “maize strain”, which feeds upon maize and sorghum. Potential control measures are generally more applicable to the farmers who lack financial assets to buy chemical insecticides or costly pure seeds. The adverse effects of pesticides on the ecosystem and human’s health and the development of resistance to insect pests have exaggerated efforts to find an alternative strategy that is cost-effective, low-risk and target-specific. Therefore, biological control is widely considered as one of the most important options for insect pest management. This comprehensive review amasses the information on biological control in all phases of their development, including predators, parasitoids, entomopathogenic fungi, viruses, nematodes, bacteria, and biopesticides, with a special focus on their effectiveness against FAW. The findings regarding biological control are briefly discussed in light of improving management programs of the invasive pest S. frugiperda.
1. Introduction
The fall armyworm (FAW), Spodoptera frugiperda, is the most important pest worldwide, resulting in considerable yield losses in maize. FAW was first reported in 1797 as a devouring pest endemic to subtropical and tropical regions of America. It belongs to the family Noctuidae under the order Lepidoptera and was first reported in the African continent [1]. FAW is a devastating pest that damages 186 plant species belonging to 42 families. Poaceae, Fabaceae, Solanaceae, Asteraceae, Rosaceae, Chenopodiaceous Brassicaceae, and Cyperaceae are mostly affected. It results in about 58% yield loss in maize [2,3]. FAW is known to feed voraciously on more than 350 plant species, especially maize, rice, and sorghum, which might cause significant agricultural losses worldwide [3,4,5]. The first confirmed reports of FAW invasion were documented from West Africa in early 2016, and then spread throughout sub-Saharan Africa and Southeast Asia [1,6]. Now, this pest has been spread in more than 109 countries [3]. In India, it was first identified in May 2018, causing major losses to farmers in Karnataka and other southern Indian states [7]. FAW was first discovered in Nepal in May 2019 through morphological and genetic identification approaches [8].
FAW is an important notorious pest which attacks maize and various other crops belonging to the family Gramineae [9]. It is a polyphagous insect pest with over 80 host plants that causes severe damage to cereal crops and vegetables [1,10,11]. The young curl of leaves, ears, and tassels have been preferred, resulting in significant loss to maize crop [12]. FAW travels approximately 500 km before starting oviposition [10]. A single generation of FAW moths can disperse more than 500 km distance from the emergence location, owing to the wind, until they are sexually mature [11,13,14].
FAW is comprised of two genetic strains: the “rice strain”, which prefers rice and other grass species, and the “maize strain”, which feeds primarily on maize and occasionally on sorghum [15]. When FAW arrives in large numbers, especially with an offensive effect, it is determined to pose a long-term and damaging threat to many important crops, as the surrounding circumstances provide a comfortable environment for a variety of host plant species pre-favorable weather conditions for reproduction in various areas [1,4,16].
Biological control strategies are more appropriate to farmers who do not have the financial capability to purchase chemical insecticides and expensive seeds [17]. Microbial formulations are available in the market that are made from pathogens, arthropod natural enemies, and are more profitable in agricultural systems [3,18]. Recently, the microbial formulation production costs have been significantly reduced because these are mainly mass produced in liquid medium [3,19]. The repetitive use of synthetic pesticides in the field may prove detrimental to humans and the environment, have increased input cost, and, furthermore, initiate resistance and resurgence [20,21,22,23,24,25]. The larvae of the FAW caterpillar are deeply embedded in the leaf curls and corn ears, resulting in control failures. However, it comes to feed on plants at night or dawn and twilight [26].
What are the Biological and Biorational Pest Control Options?
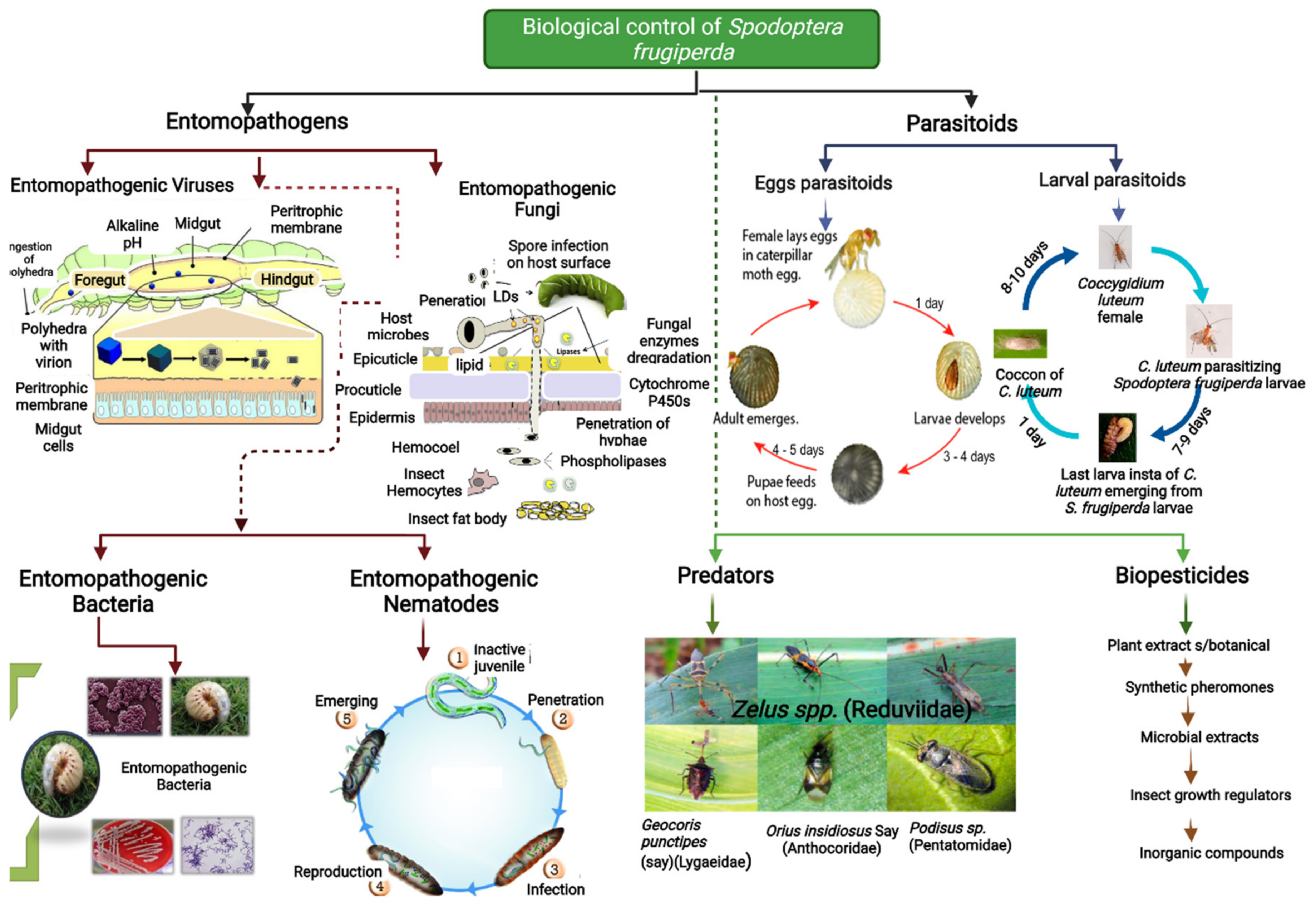
The easiest way to manage the insect pest outbreak is to formulate a self-propelling and self-perpetuating system for the restoration or stimulation of self-sustainable biological control tactics. Biological control agents, such as predators, parasitoids, and pathogens, were supplied to keep close synchronization of the community of other organisms. Trichogramma and Telenomus are the most active biological control agents, which parasitized the FAW eggs and other key pests [27,28]. Parasitoids are closely associated with one of the pest stages and have a higher level of specificity [29]. Predators are rarely linked with a specific insect pest. They feed on the prey with a lower trophic level of specificity than other animals, e.g., the earwigs, ladybird beetles, podisus, and orius prey on different life stages of FAW. Entomopathogens include bacteria, fungi, viruses, protozoans, and nematodes that are the main cause of disease in insects. FAW was infected by several entomopathogens, including Bacillus thuringiensis (Bt), Metarhizium anisopliae, Beauveria bassiana, and Spodoptera frugiperda multiple nucleopolyhedroviruses (SfMNPV) (Figure 1).
In agricultural systems, certain arthropods and microbial pathogens have been used as biocontrol agents [18,30]. Biological pest management is a powerful tool for plant protection [30]. Four hymenopteran and one dipteran species from Kenya, Tanzania, and Ethiopia have been identified, including Chelonus curvimaculatus Cameron (Braconidae), Charops ater Szepligeti (Ichneumonidae), Coccygidium luteum (Brulle) (Braconidae), and one parasitoid of Diptera Palexorista zonata (Curran) (Tachinidae) [31]. The level of parasitism varies in different parasitoids. Cotesia icipe was the dominant larval parasitoid, showing a level of parasitism in Ethiopia ranging from 33.8% to 45.3% [31]. In Kenya and Tanzania, P. zonata parasitizes 12.5%, whereas C. luteum parasitizes 04 to 8.3% [31]. The parasitism levels of two parasitoids, C. luteum and Charops ater, in Kenya and Tanzania, were 04–8.3% and 06–12%, respectively [32]. The most encouraging point under this prospect is the presence of natural enemies despite express invasion and speedy damaging habit of FAW.
Biological Control agent: a contemporary release against FAW
Telenomus or Trichogramma wasps are among the best options for potential development of biological control programs targeting FAW eggs [33], although FAW egg mass scale thickness could lower Trichogramma parasitoid performance on this host [34]. Area-wide releases are made at tactical points (ranging from 20 to 40 per hectare) under 03-day intervals to obtain a constant presence of adult wasps for achieving proactive egg parasitism [35]. This improved system is employed early in the season to limit the season-long spread of FAW in egg tracking and destruction [33]. Synchronization in parasitoid release and the existence of FAW egg masses determine the efficacy of applied biocontrol [36]. However, physically searching for FAW egg masses is a more tiresome activity compared with pheromone traps, regarded as the most effective approach to track the arrival of the FAW moth [37,38,39].
In the Natural Control of FAW, the importance of another beneficial insect
Maize fields (in the Caribbean and America) include a diverse range of beneficial insects [40]. Chelonus insularis is one of the major biological control agents that parasitize the eggs of FAW under natural conditions [41]. However, FAW eggs hatch into the larvae and release the parasitized adult from the larva of FAW [41]. Trichogramma and Telenomus wasps are smaller and less competitive than Chelonus wasps; thus, they have a better chance of surviving [41]. The parasite larvae’s food consumption is steadily reduced, with less than 10% of the biomass eaten by the borehole larvae [42]. The presence of young larvae in the FAW release zone does not necessarily mean that there has been a failure in the biological control of FAW [42]. Deployment of a conservation technique will sustain the natural biological control agents for obtaining a self-perpetuating and self-propelling organized agro-ecosystem. More information is given on the role of Chelonus in IPM [43,44].
In addition to C. insularis, many other parasitoid species are also observed in suppressing populations of FAW larvae [45]. Campoletis flavicincta has been extensively used to suppress the FAW population [46]. Insect predators consume both larvae and eggs that necessarily maintain the FAW population to a sub-economic level [47]. For example, during the maize crop cycle, the predatory earwig Doru luteipes (Scudder) lays its eggs within the maize whorl [47]. Doru luteipes nymphs consume 8–12 larvae per day, while adults consume 10–21 S. frugiperda larvae daily [47]. The comprehensive demonstration of biocontrol is shown in Figure 1. Microorganisms, such as Baculovirus or Bacillus thuringiensis, should be considered in situations where the presence of biocontrol agents is not yet optimal and pesticide applications are needed [45,48].
2. Categories of Biological Control Agents
Predators and Parasitoids
About 150 parasitoid species have been identified from different regions of America and Caribbean. Ashley [49] reported 53 different parasitoid species, including Apanteles marginiventris, Chelonus insularis, Ophion spp. Ternelucha spp. Rogus laphygmae, Campoletis grioti, Ephisoma vitticole, and Meteorus autographae in S. frugiperda eggs and larvae. More than 44% of natural parasitism has been recorded in the non-sprayed fields of America [13]. These species showed at least 45.3% of parasitism level [50]. Seven species of parasitoids and three species of predators were identified from Ghana to control S. frugiperda [51]. These seven parasitoid species are listed as Anatrichus erinaceus (Loew), M. testacea, C. icipe, Bracon sp., C. bifoveolatus, C. luteum, and an uncertain tachinid fly (Diptera: Tachinidae), while the three species of predators include Peprius nodulipes (Signoret), Haematochares obscuripennis (Stal), and Pheidole megacephala F. [51]. The parasitism degree and regional differences in the presence of species have been reported [50,51,52]. This finding is based on crop stage and type changes, agronomic methods, and geographic regions [53,54]. Coccygidium luteum was reported from Kenya and Tanzania, which causes up to 9–19% parasitism in S. frugipera [50].
To control the increasing pest population of S. frugiperda in America, mass breeding and release of predators and parasitoids have been used to manage other pests [2,37,38,39] (Table 1 and Table 2). In Sub-Saharan Africa, the implementation of classical biocontrol due to its high cost is required by the Government to control S. frugiperda [37]. Native parasitoids having a better level of parasitism have been observed from different vicinities of SSA [33,50,51,52]. The best way to control FAW is augmentative biocontrol, releasing predators to overcome the increasing pest population of FAW [37] (Table 1). In America, Trichogramma parasitoids have been used to efficiently control the eggs of S. frugiperda [10,39] (Table 2). Scientists at ICIPE in Kenya and Agboyi et al. [33] observed that Trichogramma and Telenomus parasitoid can efficiently augmentative biocontrol against S. frugiperda. Before FAW neonates emerge, parasitoids (Trichogramma and Telenomus) are introduced into maize fields, search for FAW egg masses, and lay their eggs on them to limit the FAW population at the egg stage [36].
In Africa, Lepidopterous species have been parasitized by C. luteum, the lepidopterous species including Crypsotidia mesosema (Hampson), Spodoptera exigua (Hübner), Prophanti ssp. Spodoptera exempta (Walker), Condica capensis (Guenée), and Cydia ptychora (Meyrick) [55]. Coccygidium luteum has been reported in Africa and many other countries, such as Nigeria, Madagascar, Kenya, Guinea, Congo, Ethiopia, Namibia, Mauritius, Rodrigues Island, Tanzania, Uganda, Somalia, South Africa, and Cameroon [33]. Coccygidium luteum is a solitary koinobiont parasitoid that belongs to the Braconid subfamily Agathidinae, which includes more than 46 species [56,57]. As biocontrol agents, Agathidinae species in this subfamily are not well known regarding their efficacy against insect pests and are rarely studied [58]. In China, C. luteum controls the eggs of many Spodoptera species [59,60]. The comprehensive demonstration of biocontrol is shown in Figure 1. Parasitoids can complete several generations in 90 days, leading to the emergence of early-maturing varieties of maize in West Africa [61]. Populations of natural enemies are affected due to the occurrence of variation in parasitism [62]. In the United States, parasitism levels were lower on average than previously reported levels, i.e., 15.5% [63], 35% [64], 8.1% [65], 28.3% [41], 18.3% [66], and 13.8% [67]. Agboyi et al. [33] reported parasitoids of 10 different species from the various localities of Ghana and Benin. These species are Trichogramma spp. Meteoridea cf, Charops spp, Drino quadrizonula (Thomson), Metopius discolour (Tosquinet), Telenomus remus, Chelonus bifoveolatus (Szpligeti), Coccygidium luteum, Pristomerus pallidus (Kriechbaumer), and Cotesia icipe [33]. Introduction, conservation, and augmentation are the three basic techniques for promotion of a biological control system in an ecosystem. Hymenoptera insects act as egg and larval parasitoids that are collected from FAW-infested areas. Further assistance might be given to stabilize the system either through inoculative or inundative releases. This manipulation chiefly increased the effectiveness of natural enemies.

{kind=link}
Table 1.
Predators against Spodoptera frugiperda.
| Scientific Name | Family | Order | Description | Reference |
|---|---|---|---|---|
| Earwigs: (a) Doru luteipes (Scudder) | Forficulidae | Dermaptera | Very important parasitoid of FAW. According to bioecological studies feeding on FAW larvae, the number of eggs per oviposition is 25–30, with an incubation period of about one week | [68] |
| (b) Euborellia annulipes (Lucas) | Anisolabididae | Dermaptera | A very important parasitoid of Spodoptera frugiperda also feeding on FAW larvae | [69] |
| Flower bug Orius insidiosus (Say) | Anthocoridae | Heteroptera | Extremely abundant species are mostly used in biological control programs. They prey on small arthropods, such as aphids, thrips, whiteflies, mites, and lepidopteran eggs | [70] |
| Ground beetle Calosoma calidum (Fabricius) | Carabidae | Coleoptera | After mating, the females lay their eggs on the soil’s surface or just below it. Before pupation in the ground, the immature stage goes through 3 instars | [71] |
| Spined soldier bug Podisus maculiventris (Say) | Pentatomidae | Heteroptera | Lepidopteran larvae are the primary food source for nymphs and adults. It pierces its prey and injects a toxin that causes it to become paralyzed in a short period. The predator sucks out the prey’s internal fluids, killing it | [72] |
| Assassin bug Pristhesancus plagipennis (Walker) | Reduviidae | Hemiptera | The genus Zelus is the most common killer bug in maize. Females lay their eggs in clusters on plant leaves or even on the ground. The nymphs are wingless and look like adults | [73] |
| Pirate bug Orius sauteri (Poppius) | Anthocoridae | Hemiptera | An important parasitoid. Moth eggs, aphids, and small Lepidoptera larvae are all prey for O. sauteri | [74] |
Table 2.
Parasitoids against Spodoptera frugiperda.
| Scientific Name | Family | Order | Description | Reference |
|---|---|---|---|---|
| Trichogramma pretiosum (Riley) | Trichogrammatidae | Hymenoptera | Egg parasitoids, used in the control of eggs of FAW and Helicoverpa spp. | [75] |
| Trichogrammatoidea armigera (Nagaraja) | Trichogrammatidea | Hymenoptera | Egg parasitoids, used in the control of eggs of Helicoverpa armigera and FAW | [76] |
| Telenomus remus (Nixon) | Scelionidae | Hymenoptera | Egg parasitoid, shows more than 45% parasitism level [31]. Telenomus remus completes its life cycle within 12–13 days at 25 °C, and its female parasitizes more than 250 eggs in [77] | [75] |
| Chelonus insularis (Cresson) | Braconidae | Hymenoptera | Egg–larval parasitoids, The female Chelonus lays eggs inside the eggs of fall armyworm, and the larva, which is parasitized, steadily curtails down its feeding until its death happens | [78] |
| Chelonus bifoveolatus (Szpligeti) | Braconidae | Hymenoptera | Egg–larval koinobiont endoparasitoid, mostly attack the insects belonging to the family of Pyralidae and Noctuidae. The parasitoid attacks by ovipositing into the host eggs [66] | [51] |
| Campoletis sonorensis (Cameron) | Ichneumonidae | Hymenoptera | Larval parasitoids, females lay eggs within the first, second, and third instars of the FAW, and the larva completes its life cycle by feeding on the pest’s internal contents. For the parasitoid, the third-instar larvae are the most favorable stage | [78] |
| Charops sp. | Ichneumonidae | Hymenoptera | Larval parasitoids, used to control FAW population on the larval stage. | [33] |
| Cotesia icipe (Fernández-Triana & Fiaboe) | Braconidae | Hymenoptera | Larval parasitoids, also Lepidopterous larvae and eggs of FAW have been parasitized by C. icipe [79], and the larval instars of 1st and 2nd stage have been parasitized too [80]. | [81] |
| Habrobracon hebetor (Say) | Braconidae | Hymenoptera | Larval parasitoids, under laboratory conditions, a small wasp that has been used to combat the pearl millet head miner, often targets FAW larvae. | [33] |
| Coccygidium luteum (Brullé) | Braconidae | Hymenoptera | Larval parasitoids internally attack larvae belonging to the Noctuidae family [82]. An idiobiont ectoparasitoid Bracon sp. of C. luteum hiddenly attacks the larvae of cereal stored products and cereal stem borers and is also used against S. frugiperda [31,50,83,84]. | [33] |
| Winthemia trinitatis (Thompson) | Tachinidae | Diptera | Larval parasitoids, the female lays her eggs near the head of a fifth- or sixth-instar FAW, making it difficult to detach. The parasitoid’s larvae enter the larva’s body, delaying the pupation and resulting in up to 30% parasitism. | [85] |
| Drino quadrizonula (Thomson) | Tachinidae | Diptera | Larval parasitoids, parasitized family of Noctuidae and other Spodopterous spp. | [86] |
| Archytas marmoratus (Townsend) | Tachinidae | Diptera | Larval–pupal parasitoids, several species of Noctuidae (Lepidoptera), including FAW. Eggs are not laid directly on the host but instead scattered them around. Hatching takes place, and the young larvae emerge. When these larvae come into contact with a host and invade the host’s body, parasitism occurs. | [50] |
| Lespesia archippivora (Riley) | Tachinidae | Diptera | Larval–pupal parasitoids, on the back end of the caterpillar, the female oviposits. Lespesia archippivora has three instars that feed on the host caterpillar, and when they reach maturity, the parasitoid emerges from the larva and pupates in the soil. | [87] |
3. Microorganisms
Entomopathogens infect and cause diseases in insects, and include viruses (Spodoptera frugiperda multiple nucleopolyhedrovirus), bacteria (Bacillus thuringiensis), fungi (Metarhizium anisopliae and Beauveria bassiana), nematodes, protozoans, and others that are well known against FAW management.
3.1. Entomopathogenic Viruses
Virus-based insecticides among microbial control agents within the Baculovirus group have been recognized with higher potential as bioinsecticides for growth and development because of their specific characteristic nature, higher virulence against the host, and increased safety towards vertebrates [88,89]. Two variants of Baculovirus have been studied to control S. frugiperda, named granulovirus “SfGV”, which is Betabaculovirus, and multiple nucleopolyhedroviruses “SfMNPV”, which is Alphabaculovirus. However, the SfMNPV has a good ability to manage FAW [90,91,92]. SfMNPV is specific to FAW larvae. The pest becomes infected by ingesting contaminated (maize leaf) food in natural conditions [93]. After ingestion, polyhedral inclusion bodies (PIB) dissolve in an alkaline midgut, while infectious virions are released. The comprehensive demonstration of biocontrol is shown in Figure 1. These virions cause infection in epithelium cells of the midgut and start multiplying in the nucleus [10]. Further, these viruses are distributed in the body cavity and start infecting other tissues, i.e., tracheal matrix, adipose tissue, epidermal, even including Malpighian tube, salivary glands, and blood cells [94], which leads to its death after ingestion in 6 to 8 days compared to a healthy caterpillar; an infected one with nucleopolyhedroviral only eats 7% of food [94]. In Baculovirus infection, the main symptoms include yellowing of the skin, the appearance of blemishes, and a decrease in feeding [94]. Usually, infected larvae start moving upper parts and are attached to the plants in a hanging-down position upon their death, while some prolegs remain attached to plants [94]. Such larvae are soft with dark color and found with released body fluids rich in polyhedrons, aiding the further spread of the virus [94]. Virus efficacy and speed of killing FAW larvae depends on larva age at the time of infection, ingested virus amount, and its virulence with current climatic conditions, i.e., temperature, humidity, and solar radiation [95,96]. In addition to this, time of spray, equipment type, and formulations are influential in the efficiency of the virus [95,96].
3.2. Entomopathogenic Fungi
Entomopathogenic fungi (EPF) can infect several insect species on different stages at a wide range, causing epizootics under certain natural conditions [97]. Fungal spores of such EPF species infect through integument, and then multiply within the insect’s body. After multiplying, EPF releases certain toxins that destroy tissues and the insect dies. The introduction of epizootics depends upon climatic conditions and the frequency of insect contact [97]. When insects become infected with EPF, they become discolored (cream, green, reddish, or brown), stop eating, and, finally, die as a hard calcareous cadaver, in which the fungus begins to sporulate [98]. Moisture has an important role for fungi as a biocontrol agent. Among the potentially beneficial fungi against insect pests, Beauveria bassiana, Metarhizium anisopliae, and Nomuraea rileyi are the most commonly used for Spodoptera control [98]. FAW larvae are susceptible to B. bassiana compared to other lepidopteran pests [99]. In vitro studies showed 30% and 87% mortality when B. bassiana and M. anisopliae was applied on second instars and eggs, respectively.
3.3. Entomopathogenic Bacteria
Bacillus thuringiensis (Bt) Berliner is one of the most widely used biopesticides for insect control [100]. These bacteria are Gram-positive, soil-dwelling bacteria that aid in producing crystal proteins known as delta-endotoxins, which are naturally insecticidal [101]. Only a few Bt products are effective against FAW available in the market for managing lepidopteran pests [101].
FAW has increased susceptibility to Bt aizawai and Bt thuringiensis [100] compared to Bt kurstaki, which is efficient against numerous lepidopteran pests [102]. There are few limitations in its wide adoption and use, including UV susceptibility of endotoxin, inability to reach pest for inducing toxin consumption, and the high cost of production [102]. Various research groups are trying to determine more effective Bt strains against FAW. In contrast, variation in susceptibility towards different Cyt toxins (also known as Cry toxins) has been observed among populations of FAW [103]. To control FAW, Bt-based biopesticides must be considered throughout the selection process in various locations. Seven Bt strains were highly effective among nineteen in screening against second-instar FAW larvae at ICIPE in Africa, causing 100% mortality within 7 days of post-treatment, having lethal time mortality (LT50) with standard ranges of 2.33 ± 0.33 and 6.50 ± 0.76 days. Bt-based biopesticides have been considered for mass production through fermentation technology using solid-, semi-solid-, or liquid-state fermentation [104]. Not only Cry toxins, but also FAW show susceptibility towards vegetative insecticidal proteins that could be usually seen in Bt culture supernatants [105].
3.4. Entomopathogenic Nematodes EPNs
Entomopathogenic nematodes, i.e., Steinernema feltiae, Steinernema carpocapsae, Heterorhabditis indica, and Heterorhabditis bacteriophora, are used as effective biological control agents [106]. EPNs are eco-friendly and play an essential role in suppressing soil-dwelling insect pests, notably armyworms [106]. The susceptibility rate of FAW to such beneficial nematodes is 23,000 per sq. ft. to target mature and young larvae [10]. Early morning or late night are the most efficient times to apply beneficial nematodes because they are more active at these times and are less exposed to ultraviolet (UV) light, which helps them find armyworm larvae more easily [107].
In a petri dish, 280 infective juveniles of Steinernema sp. can kill 100% of third-instar FAW, while 400 infective juveniles of H. indica kill 75% of FAW [108]. Usage of hydraulic spraying jets, which require 100 filtrating mesh elements, could decrease the concentration of infective juveniles of H. indica and Steinernema species up to 28% and 53%, respectively [10]. The comprehensive demonstration of biocontrol is shown in Figure 1. Molina-Ochoa et al. [109] informed earlier that S. carpocapsae and S. riobravis efficiently manage FAW at the prepupal stage. According to various researchers, EPNs with resistant maize silk could increase FAW mortality at the prepupal stage [110]. These nematode species, S. carpocapsae, S. glaseri, and H. indica, have shown compatibility with various commercial insecticides under laboratory conditions [110]. Heterorhabditis indica shows increased efficacy when mixed with lufenuron against FAW [111]. Moreover, a compatibility evaluation study of insecticides or biopesticides with EPNs is necessary before recommending it to use an IPM for FAW [111].
4. Botanical Pesticides
Local farmers have claimed that botanical extracts from local plants are beneficial [10]. Botanical pesticides are a better alternative to synthetic insecticides, which could be more harmful to the environment, increase consumer cost, and delay recovery [112,113,114,115] (Table 3). Such pesticides are also responsible for increased pest resistance [116,117,118,119]. Few botanical extracts include Chrysanthemum cinerariifollium, Jatropha curcas, Nicotina tabacum, Milletia ferruginea, Phytolacea docendra, and Croton macrostachyus, which could be used as insect pest control [120] (Table 3). Around 50 botanical pesticides were found to be registered for controlling FAW in more than 30 countries (Table 3). Among those, 23 are recommended for field trials and bioassays [2,121]. Solaris 6 SC® was shown to be the most effective insecticide against immature fall armyworms, followed by botanical extracts of garlic and neem, as well as detergent [10].
Botanical pesticides caused 80% mortality under laboratory conditions [122]. According to different reports and studies, these botanicals are effective against FAW [123,124] (Table 3). Neem extracts have shown 70% mortality in FAW [125,126]. Eucalyptus urograndis was found to be more helpful in saving maize from pests [124]. The seed powder of Carica papaya was discovered as an efficient chemical insecticide [127]. Neem oil containing 0.17–0.33% concentration reduces FAW damage in maize [123].
Botanical insecticides are target-specific, non-hazardous for the environment, and safe for natural enemies as compared to chemical pesticides [128]. Thus, their application promotes FAW natural parasitism up to 60% in comparison with pesticide-treated areas [41].
Table 3.
Potential botanical pesticides against FAW.
| Extract | Mode of Action | Species | Family | References |
|---|---|---|---|---|
| 0.25% Neem oil | Larvicidal with up to 80% mortality in the lab | Neem: Azadirachta indica | Meliaceae | [122] |
| Hexane and ethanol extracts of seeds | Larvicidal with up to 100% mortality in the lab | Aglaia cordata Hiern | Meliaceae | [129] |
| Ethanolic extract from seeds | Larval growth inhibition | Annona mucosa Jacquin | Annonaceae | [130] |
| Ethanol extracts from leaves | Ovicidal | Vernonia holosenicea, Lychnophora ramosissima, and Chromolaena chaseae | Asteraceae | [131] |
| Dichloromethane extracts of wood | Insect growth regulating (IGR) and larvicidal with up to 95% mortality | Cedrela salvadorensis and Cedrela dugessi | Meliaceae | [132] |
| Methanol extracts of roots and other aerial parts | Insect growth regulating (IGR), larvicidal, delayed pupation | Myrtillocactus geometrizans | Cactaceae | [133] |
| Essential oil from seeds | Affects spermatogenesis and, hence, egg laying | Long pepper, Piper hispidinervum | Piperaceae | [134] |
| Ethanolic extracts of leaves | Antifeedant to larva; synergistic with pesticide | Melia azedarach | Meliaceae | [135] |
| Ethanolic extracts of leaves | Antifeedant to larva; synergistic with pesticide | Jatropha gossypifolia | Euphorbiaceae | [135] |
| Castor oil and Ricinine (seed extracts) | Growth inhibition and larvicidal | Ricinus communis | Euphorbiaceae | [136] |
(Source: adopted from Fall Armyworm in Africa: A Guide for Integrated Pest Management).
A recent study conducted in Ghana recommends farmers to use intercropping technique during the first nine weeks of crop establishment because foliage of a crop remains too soft for FAW neonates; therefore, moths are attracted by crop to lay eggs [137]. Under this duration, intercropping encouraged natural biological control agents to establish under the prevailing conditions. It resultantly checked the FAW population [13]. Push–pull technology (PPT) is now suggested for FAW management [36]. International Centre of Insect Physiology and Ecology (ICIPE) introduced this method to control stem borers in maize. The push–pull technology comprises intercropping maize with drought-tolerant greenleaf Desmodium intortum (Mill.) Urb., while Brachiaria cv Mulato II is planted as a border crop surrounding the intercrop. Desmodium plays an important role in protecting maize crops by repelling moths away with the emission of semiochemicals [16]. At least 80% of FAW infestations can be eliminated using this technique [16,36]. In Uganda, FAW infestation levels on maize using PPT were 36–38%, compared to 95% when single cropping was used. PPT is better than maize-legume intercropping for controlling FAW infestation [138]. Botanical or biopesticides are recommended as an alternative to hazardous synthetic insecticides, such as pyrethroids and organophosphorus compounds, which can influence and interfere with environmental conditions, and increase expense, resurgence, and insect resistance.
Because of their low cost and accessibility, farmers and growers in developing countries use botanical, eco-friendly, sustainable techniques to manage insect pests of field crops and stored goods. Milletia ferruginea, Azadirachta indica, Croton macrostachyus, Jatropha curcas, Phytolacea docendra, Chrysanthemum cinerariifollium, and Nicotina tabacum extracts have been exploited successfully against insect pests [139]. Azadirachta indica seed extract causes the highest mortality in FAW at the larval stage [126]. Martínez et al. [140] discovered that the Argemone ochroleuca causes FAW mortality by reducing feeding behavior and stunting larval growth. Various botanical plant extracts could increase insecticidal efficiency against FAW [141]. Several products, such as extracts of Azadirachtin from neem and pyrethrins from pyrethrum, have been successfully commercialized, while others, such as those based on garlic, ryanodine, quassia, nicotine, and rotenone, have been registered worldwide [142]. Commercial products are available under different formulations and mode of action, applied diluted with water or sprayed with chemical insecticides and dust formulation. Furthermore, there are difficulties in application mode, such as neem-based solutions having high photosensitivity for Azadirachtin, resulting in a lower residual impact in fields due to sunlight exposure [122]. The comprehensive demonstration of biocontrol is shown in Figure 1. Few botanicals, including neem (Azadirachta indica), pyrethrum (Tanacetum cinerariifolium), fish-poison bean (Tephrosia vogelii), wild sage (Lantana camara), West African pepper (Piper guineense), wild marigold (Tagetes minuta), onion (Allium sativum, Allium cepa), tobacco (Nicotiana sp.), chilies (Capsicum sp.), lemongrass (Cymbopogon citratus), chrysanthemum (Chrysanthemum sp.), wild sunflower (Tithonia diversifolia), acacia (Acacia sp.), jatropha (Jatropha curcas), and Persian lilac (Melia azedarach), have good insecticidal properties in managing stemborers in Africa [143,144,145].
5. Conclusions
Spodoptera frugiperda is a worldwide invasive pest, causing significant losses in host crops. FAW comprises two genetic strains: the “rice strain”, which prefers rice and other grass species, and the “maize strain”, which feeds primarily on maize and occasionally on sorghum. Small and marginal farmers that lack the financial resources to purchase chemical pesticides or expensive seeds are more likely to benefit from potential control methods. Maintaining natural antagonist diversity and intensity is crucial for biological control strategies. It may be accomplished by providing favorable conditions for biological control agents near crops. Shelter crops, alternate food resources, and multiplication favoring environments for such biological species are all important steps in regulating the population of FAWs. Growing Mexican sunflower or Crotalaria on crop edges in maize-growing areas is a good way of boosting beneficial insect biodiversity, even if they are not related to FAW, and achieve landscape management goals. The push–pull technology (PPT) intercropping is also beneficial against pests to repel them out of fields. In addition, economic injury levels (EIL), observation through surveys, and pest management efforts might be used to evaluate biocontrol-based IPM applications against FAW.
Author Contributions
A.A.: Conceptualization, Writing—original draft. F.U.: Conceptualization, Writing—original draft, Writing—review and editing. M.H.: Writing—review and editing. X.H.: Writing—review and editing. M.Z.N.D.: Writing—review and editing. H.G.: Writing—review and editing. C.R.Z.: Conceptualization, review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
We would like to thanks the Government of Jilin Province for funding under the Jilin Government Fund on corn-pest predicting-and-controlling Control (20220203198SF).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I would like to thank the editor and four anonymous reviewers for their attention to the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.M.; Gadratagi, B.G.; Paramesh, V.; Kumar, P.; Madivalar, Y.; Narayanappa, N.; Ullah, F. Sustainable Management of Invasive fall armyworm, Spodoptera frugiperda. Agronomy 2022, 12, 2150. [Google Scholar] [CrossRef]
- Kenis, M.; Benelli, G.; Biondi, A.; Calatayud, P.A.; Day, R.; Desneux, N.; Harrison, R.D.; Kriticos, D.; Rwomushana, I.; Van den Berg, J.; et al. Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomol. Gen. 2022. [Google Scholar] [CrossRef]
- Montezano, D.G.; Sosa-Gómez, D.R.; Roque-Specht, V.F. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 16. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; He, P.C.; Hu, L.; Chi, X.L.; Keller, M.A.; Chu, D. Host selection and adaptation of the invasive pest Spodoptera frugiperda to indica and japonica rice cultivars. Entomol. Gen. 2022, 42, 403–411. [Google Scholar] [CrossRef]
- Nagoshi, R.; Htain, N.; Boughton, D.; Zhang, L.; Xiao, Y.; Nagoshi, B.; Mota-Sanchez, D. Southeastern Asia fall armyworms are closely related to populations in Africa and India, consistent with common origin and recent migration. Sci. Rep. 2020, 10, 1421. [Google Scholar] [CrossRef] [Green Version]
- Sharanabasappa, S.D.; Kalleshwaraswamy, C.M.; Maruthi, M.S.; Pavithra, H.B. Biology of invasive fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) on maize. Ind. J. Entomol. 2018, 80, 540–543. [Google Scholar] [CrossRef]
- Bajracharya, A.; Bhat, B.; Sharma, P.; Shashank, P.; Meshra, N.; Hashmi, T. First Record of fall armyworm. Ind. J. Entomol. 2019, 81, 635–639. [Google Scholar] [CrossRef]
- Andrews, K.L. The whorlworm, Spodoptera frugiperda, in Central America and neighbouring areas. Fla. Entomol. 1980, 63, 456–467. [Google Scholar] [CrossRef]
- Prasanna, B.; Huesing, J.; Eddy, R.; Peschke, V. Fall Armyworm in Africa: A Guide for Integrated Pest Management; USAID; CIMMYT: Mexico City, Mexico, 2018. [Google Scholar]
- Wu, P.; Ren, Q.; Wang, W.; Ma, Z.; Zhang, R. A bet-hedging strategy rather than just a classic fast life-history strategy exhibited by invasive fall armyworm. Entomol. Gen. 2021, 41, 337–344. [Google Scholar] [CrossRef]
- De-Almeida, S.R.; De-Souza, A.R.W.; Vieira, S.M.J.; De-Oliveira, H.G.; Holtz, A.M. Biology review, occurrence and control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in corn in Brazil. J. Biosci. 2002, 18, 41–48. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). Sustainable Management of the Fall Armyworm in Africa; FAO Programme for Action; FAO: Rome, Italy, 2017. [Google Scholar]
- Pogue, M.G. A World Revision of the Genus Spodoptera Guenée: (Lepidoptera: Noctuidae); American Entomological Society Philadelphia, U.S. Department of Agriculture: Washington, DC, USA, 2002. [Google Scholar]
- Sparks, A.N. A review of the biology of fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Midega, C.A.O.; Pittchar, J.O.; Pickett, J.A.; Hailu, G.W.; Khan, Z.R. A climate-adapted push-pull system effectively controls fall armyworm, Spodoptera frugiperda (J.E Smith), in maize in East Africa. Crop Prot. 2018, 105, 10–15. [Google Scholar] [CrossRef]
- Abate, T.; Van-Huis, A.; Ampofo, J.K.O. Pest management strategies in traditional agriculture: An African perspective. Annu. Rev. Entomol. 2000, 45, 631–659. [Google Scholar] [CrossRef]
- Pilkington, L.J.; Messelink, G.; Van-Lenteren, J.C.; Le-Mottee, K. “Protected biological control” Biological pest management in the greenhouse industry. Biol. Control 2010, 52, 216–220. [Google Scholar] [CrossRef]
- Mahmoud, M. Biology and use of entomopathogenic nematodes in insect pest bio-control, a generic view in Moldova. Cercet. Agron. 2016, 49, 85–105. [Google Scholar]
- Weisenburger, D.D. Human health effects of agrichemical use. Hum. Pathol. 1993, 24, 571–576. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Desneux, N.; Gao, X.; Song, D. Imidacloprid-induced hormesis effects on demographic traits of the melon aphid, Aphis gossypii. Entomol. Gen. 2019, 39, 325–337. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Acetamiprid resistance and fitness costs of melon aphid, Aphis gossypii: An age-stage, two-sex life table study. Pestic. Biochem. Physiol. 2020, 171, 104729. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Functional analysis of cytochrome P450 genes linked with acetamiprid resistance in melon aphid, Aphis gossypii. Pestic. Biochem. Physiol. 2020, 175, 104687. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Ullah, F.; Luo, C.; Monticelli, L.S.; Lavoir, A.V.; Gao, X.; Song, D.; Desneux, N. Sublethal effects of beta-cypermethrin modulate interspecific interactions between the specialist and generalist aphid species of soybean crops. Ecotoxicol. Environ. Saf. 2020, 206, 111302. [Google Scholar] [CrossRef] [PubMed]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Gowda, G.B.; Madhusmita, S.; Farman, U.; Naveenkumar, B.P.; Guru Pirasanna, P.; Adak, T.; Pokhare, S.; Mahendiran, A.; Rath, P.C. Insecticide-induced hormesis in a factitious host, Corcyra cephalonica, stimulates the development of its gregarious ecto-parasitoid, Habrobracon hebetor. Biol. Control 2021, 160, 104680. [Google Scholar] [CrossRef]
- Assefa, F.; Ayalew, D. Status and control measures of fall armyworm (Spodoptera frugiperda) infestations in maize fields in Ethiopia: A review. Cogent Food Agric. 2019, 5, 1641902. [Google Scholar] [CrossRef]
- Sisay, B.; Tefera, T.; Wakgari, M.; Ayalew, G.; Mendesil, E. The efficacy of selected synthetic insecticides and botanicals against fall armyworm, Spodoptera frugiperda, in maize. Insects 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Birhanu, S. Evaluation of Different Management Options of Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) and Assessment of Its Parasitoids in Some Parts of Ethiopia; Haramaya University: Dire Dawa, Ethiopia, 2018. [Google Scholar]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R.; et al. Parasitoid Complex of fall armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects 2020, 11, 68. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.Y.; Xu, W.; Desneux, N.; Nkunika, P.O.; Bao, H.P.; Zang, L.S. Spodoptera frugiperda egg mass scale thickness modulates, Trichogramma parasitoid performance. Entomol. Gen. 2022, 42, 589–596. [Google Scholar] [CrossRef]
- Cruz, I.; Lopes, S.R.; Figueiredo, M.D.L.; Viana, P.A.; Mendes, S.M. Controle biológico de pragas do milhodoce. In O Cultivo do Milho-Doce; Pereira Filho, I.A., Teixeira, F.F., Eds.; Embrapa Cap.: Brasília, Brazil, 2016; Volume 11, pp. 205–224. [Google Scholar]
- ICIPE (International Centre of Insect Physiology and Ecology). Combating the Fall Armyworm in Africa—The European Union (EU) Provides New Funding to ICIPE. 2018. Available online: www.icipe.org (accessed on 15 December 2021).
- FAO. Integrated Management of the Fall Armyworm on Maize: A Guide for Farmer Field Schools in Africa; FAO: Rome, Italy, 2018. [Google Scholar]
- Parra, J.R.P.; Zucchi, R.A. Trichogramma in Brazil: Feasibility of use after twenty years of research. Neotrop. Entomol. 2004, 33, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Soares, M.A.; Leão, G.; Leite, D.; Zanuncio, J.C. Quality control of Trichogramma atopovirilia and Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) adults reared under laboratory conditions. Braz. Arch. Biol. Technol. 2012, 55, 305–311. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Carpenter, J.E.; Heinrichs, E.A.; Foster, J.E. Parasitoids and parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean Basin: An inventory. Fla. Entomol. 2003, 86, 254–289. [Google Scholar] [CrossRef]
- Meagher, R.L.; Nuessly, G.S.; Nagoshi, R.N.; Hay-Roe, M.M. Parasitoids attacking fall armyworm (Lepidoptera: Noctuidae) in sweet corn habitats. Biol. Control 2016, 95, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Rezende, M.A.A.; Cruz, I.; Della-Lucia, T.M.C. Consumo foliar de milho e desenvolvimento de lagartas de Spodoptera frugiperda (Smith) parasitadas por Chelonus insularis (Cresson) (Hymenoptera: Braconidae). An. Soc. Entomol. Bras. 1994, 23, 473–478. [Google Scholar] [CrossRef]
- Rezende, M.A.A.; Cruz, I.; Della-Lucia, T.M.C. Aspectos biológicos do parasitoide Chelonus insularis (Cresson) (Hymenoptera, Braconidae) criados em ovos de Spodoptera frugiperda (Smith) (Lepidoptera, Noctuidae). Rev. Bras. Zool. 1995, 12, 779–784. [Google Scholar] [CrossRef]
- Rezende, M.A.A.; Della-Lucia, T.M.C.; Cruz, I. Comportamento de lagartas de Spodoptera frugiperda (Lepidoptera: Noctuidae) parasitadas por Chelonus insularis (Hymenoptera, Braconidae) sobre plantas de milho. Rev. Bras. Entomol. 1995, 39, 675–681. [Google Scholar]
- Figueiredo, M.L.C.; Cruz, I.; Penteado-Dias, A.M.; Silva, R.B. Interaction between Baculovirus Spodoptera and natural enemies on the suppression of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in maize. Rev. Bras. Milho Sorgo 2009, 8, 207–222. [Google Scholar] [CrossRef] [Green Version]
- Matos-Neto, F.C.; Cruz, I.; Zanuncio, J.C.; Silva, C.H.O.; Picanço, M.C. Parasitism by Campoletis flavicincta on Spodoptera frugiperda in corn. Pesqui. Agropecu. Bras. 2004, 39, 1077–1081. [Google Scholar] [CrossRef] [Green Version]
- Reis, L.L.; Oliveira, L.J.; Cruz, I. Biologia e Potencial de Doru luteipes no Controle de Spodoptera frugiperda. Pesqui. Agropecu. Bras. 1988, 23, 333–342. [Google Scholar]
- Cruz, I.; Gonçalves, E.P.; Figueiredo, M.L.C. Effect of a nuclear polyhedrosis virus on Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) larvae, its damage and yield of maize crop. Rev. Bras. Milho Sorgo 2002, 1, 20–27. [Google Scholar] [CrossRef]
- Ashley, T.R. Classification and distribution of fall armyworm parasites. Fla. Entomol. 1979, 62, 114–123. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Tefera, T. First report of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Koffi, D.; Kyerematen, R.; Eziah, V.Y.; Agboka, K.; Adom, M.; Goergen, G.; Meagher, R.L. Natural enemies of the fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in Ghana. Fla. Entomol. 2020, 103, 85. [Google Scholar] [CrossRef]
- Kenis, M.; DuPlessis, H.; VandenBerg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is already Present on the continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef] [Green Version]
- Hay-Roe, M.M.; Meagher, R.L.; Nagoshi, R.N.; Newman, Y. Distributional patterns of fall armyworm parasitoids in a corn field and a pasture field in Florida. Biol. Control 2016, 96, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Ruíz-Nájera, R.E.; Molina-Ochoa, J.; Carpenter, J.E.; Espinosa-Moreno, J.A. Survey for Hymenopteran and Dipteran parasitoids of the fall armyworm (Lepidoptera: Noctuidae) in Chiapas, Mexico. J. Agric. Urban Entomol. 2007, 24, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Van, N.; Wasp, S. Web: Hymenoptera of the Afro-Tropical Region. 2019. Available online: www.waspweb.org (accessed on 15 December 2021).
- Sharkey, M.J. Cladistics and tribal classification of the Agathidinae (Hymenoptera: Braconidae). J. Nat. Hist. 1992, 26, 425–447. [Google Scholar] [CrossRef]
- Sharkey, M.J. Two new genera of Agathidinae (Hymenoptera: Braconidae) with a key to the genera of the New World. Zootaxa 2006, 1185, 37–51. [Google Scholar] [CrossRef]
- Farahani, S.; Talebi, A.; Rakhshani, E.; Achterberg, C.; Sharkey, M. A contribution to the knowledge of Agathidinae (Hymenoptera: Braconidae) from Iran with description of a new species. Biologia 2014, 69, 228–235. [Google Scholar] [CrossRef]
- Chou, L.Y. Note on Telenomus remus (Hymenoptera: Scelionidae). In Bulletin of Society of Entomology; National Chung-Hsing University: Taichung, Taiwan, 1987; Volume 20, pp. 15–20. [Google Scholar]
- Tang, Y.L.; Chen, K.W.; Xu, Z.F. Study on ontogenesis of Telenomus remus Nixon (Hymenoptera: Scelionidae). J. Changjiang Veg. 2010, 18, 1–3. [Google Scholar]
- Oluwaranti, A.; Fakorede, M.A.B.; Badu-Apraku, B. Grain yield of maize varieties of different maturity groups under marginal rainfall conditions. J. Agric. Sci. 2008, 53, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Kogan, M.; Gerling, D.; Maddox, J.V. Enhancement of biological control in annual agricultural environments. In Handbook of Biological Control; Bellows, T., Fisher, T., Eds.; Academic Press: New York, NY, USA, 1999; pp. 789–818. [Google Scholar]
- Wheeler, G.S.; Ashley, T.R.; Andrews, K.L. Larval parasitoids and pathogens of the fall armyworm in Honduran maize. Entomophaga 1989, 34, 331–340. [Google Scholar] [CrossRef]
- Rios-Velasco, C.; Gallegos-Morales, G.; Cambero-Campos, J.; Cerna-Chávez, E.; Rincón-Castro, M.C.D.; Valenzuela-García, R. Natural enemies of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) in Coahuila, México. Fla. Entomol. 2011, 94, 723–726. [Google Scholar] [CrossRef]
- Ordóñez-García, M.; Rios-Velasco, C.; Berlanga-Reyes, D.I.; Acosta-Muñiz, C.H.; Salas-Marina, M.; Cambero-Campos, O.J. Occurrence of natural enemies of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Chihuahua, Mexico. Fla. Entomol. 2015, 98, 843–847. [Google Scholar] [CrossRef]
- Murúa, M.G.; Molina-Ochoa, J.; Fidalgo, P. Natural distribution of parasitoids of larvae of the fall armyworm, Spodoptera frugiperda in Argentina. J. Insect Sci. 2009, 9, 20. [Google Scholar]
- Molina-Ochoa, J.; Carpenter, J.E.; Lezama-Gutiérrez, R.; Foster, J.E.; González-Ramírez, M.; Angel-Sahagún, C.A.; Farías-Larios, J. Natural distribution of hymenopteran parasitoids of Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae in México. Fla. Entomol. 2004, 87, 461–472. [Google Scholar] [CrossRef]
- Frizzas, M.R.; Silveira-Neto, S.; De Oliveira, C.M.; Omoto, C. Genetically modified corn on fall armyworm and earwig populations under field conditions. Ciênc. Rural 2014, 44, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Barros, E.M.; Da Silva-Torres, C.S.A.; Torres, J.B.; Rolim, G.G. Short-term toxicity of insecticides residues to key predators and parasitoids for pest management in cotton. Phytoparasitica 2018, 46, 391–404. [Google Scholar] [CrossRef]
- Ferkovich, S.M.; Venkatesan, T.; Shapiro, J.P.; Carpenter, J.E. Presentation of artificial diet: Effects of composition and size of prey and diet domes on egg production by Orius insidiosus (Heteroptera: Anthocoridae). Fla. Entomol. 2007, 90, 502–508. [Google Scholar] [CrossRef]
- Knutson, A. The Fall Armyworm—Pest of Pasture, Hayfields and Small Grains, AgriLife Extension; Texas A&M Research and Extension Center: College Station, TX, USA, 2008. [Google Scholar]
- Kneeland, K.; Coudron, T.A.; Lindroth, E.; Stanley, D.; Foster, J.E. Genetic variation in field and laboratory populations of the spined soldier bug, Podisus maculiventris. Entomol. Exp. Appl. 2012, 143, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Grundy, P. Utilizing the assassin bug, Pristhesancus plagipennis (Hemiptera: Reduviidae), as a biological control agent within an integrated pest management programme for Helicoverpa spp. (Lepidoptera: Noctuidae) and Creontiades spp. (Hemiptera: Miridae) in cotton. Bull. Entomol. Res. 2007, 97, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaraleño-Teniente, J.; Lomeli-Flores, J.R.; Rodríguez-Leyva, E.; Bujanos-Muñiz, R.; Rodríguez-Rodríguez, S.E. Egg parasitoids survey of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) in maize and sorghum in Central Mexico. Insects 2020, 11, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Freitas Bueno, R.C.O.; Carneiro, T.R.; De Freitas Bueno, A.; Pratissoli, D.; Fernandes, O.A.; Vieira, S.S. Parasitism capacity of Telenomus remus Nixon (Hymenoptera: Scelionidae) on Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) eggs. Braz. Arch. Biol. Technol. 2010, 53, 133–139. [Google Scholar] [CrossRef]
- Karimoune, L.; Ba, M.N.; Baoua, I.B.; Muniappan, R. The parasitoid Trichogrammatoidea armigera Nagaraja (Hymenoptera: Trichogrammatidae) is a potential candidate for biological control of the millet head miner Heliocheilus albipunctella (de Joannis) (Lepidoptera: Noctuidae) in the Sahel. Biol. Control 2018, 127, 9–16. [Google Scholar] [CrossRef]
- De Freitas Bueno, R.C.O.; De Freitas Bueno, A.; Xavier, M.F.D.C.; Carvalho, M.M. Telenomus remus (Hymenoptera: Platygastridae) parasitism on eggs of Anticarsia gemmatalis (Lepidoptera: Eribidae) compared with its natural host Spodoptera frugiperda (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2014, 107, 799–808. [Google Scholar] [CrossRef]
- Jourdie, V.; Virla, E.; Murillo, H.; Bento, J.M.S.; Turlings, T.C.; Alvarez, N. Phylogeography of Chelonus insularis (Hymenoptera: Braconidae) and Campoletis sonorensis (Hymenoptera: Ichneumonidae), two primary neotropical parasitoids of the fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2010, 103, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Ruberson, J.R.; Whitfield, J.B. Facultative egg-larval parasitism of the beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae) by Cotesia marginiventris (Hymenoptera: Braconidae). Fla. Entomol. 1996, 79, 296–302. [Google Scholar] [CrossRef]
- Loke, W.H.; Ashley, T.R.; Sailer, R.I. Influence of fall armyworm, Spodoptera frugiperda, (Lepidoptera: Noctuidae) larvae and corn plant damage on host finding in Apanteles marginiventris (Hymenoptera: Braconidae). Environ. Entomol. 1983, 12, 911–915. [Google Scholar] [CrossRef]
- Agbodzavu, M.; Lagat, Z.; Gikungu, M.; Rwomushana, I.; Ekesi, S.; Fiaboe, K. Performance of the newly identified endoparasitoid Cotesia icipe Fernandez-Triana & Fiaboe on Spodoptera littoralis (Boisduval). J. Appl. Entomol. 2018, 142, 646–653. [Google Scholar]
- Sharkey, M.J.; Yu, D.S.; Noort, S.V.; Seltmann, K.; Penev, L. Revision of the Oriental genera of Agathidinae (Hymenoptera, Braconidae) with an emphasis on Thailand including interactive keys to genera published in three different formats. ZooKeys 2009, 21, 19–54. [Google Scholar] [CrossRef]
- Moolman, H.J.; Vanden-Berg, J.; Conlong, D.; Cugala, D.; Siebert, S.J.; Le-Ru, B.P. Diversity of stem borer parasitoids and their associated wild host plants in South Africa and Mozambique. Phytoparasitica 2013, 41, 89–104. [Google Scholar] [CrossRef]
- Souobou, M.; Nacro, S.; Ouattara, D. Natural enemies associated with rice stem borers in the Kou Valley, Burkina Faso. Int. J. Trop. Insect Sci. 2015, 35, 164–171. [Google Scholar] [CrossRef]
- Bortolotto, O.C.; Menezes, A.D.O.; Hoshino, A.T.; Carvalho, M.G.; Pomari-Fernandes, A.; Salgado-Neto, G. Sugar solution treatment to attract natural enemies and its impact on fall armyworm Spodoptera frugiperda in maize fields. Interciencia 2014, 39, 416. [Google Scholar]
- Crosskey, R.W. The identity of Palexorista quadrizonula (Thomson), a tachinid parasite of lepidopterous pests in Africa. Bull. Entomol. Res. 1970, 59, 579–583. [Google Scholar] [CrossRef]
- Delfín-González, H.; Bojórquez-Acevedo, M.; Manrique-Saide, P. Parasitoids of fall armyworm (Lepidoptera: Noctuidae) from a traditional maize crop in the Mexican State of Yucatan. Fla. Entomol. 2007, 90, 759–761. [Google Scholar] [CrossRef]
- Moscardi, F. Assessment of the application of baculo-viruses for control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef]
- Barrera, G.; Simón, O.; Villamizar, L.; Williams, T.; Caballero, P. Spodoptera frugiperda multiple nucleopolyhedrovirus as a potential biological insecticide: Genetic and phenotypic comparison of field isolates from Colombia. Biol. Control 2011, 58, 113–120. [Google Scholar] [CrossRef]
- Behle, R.W.; Popham, H.J. Laboratory and field evaluations of the efficacy of a fast-killing baculovirus isolate from Spodoptera frugiperda. J. Invertebr. Pathol. 2012, 109, 194–200. [Google Scholar] [CrossRef]
- Gómez, J.; Guevara, J.; Cuartas, P.; Espinel, C.; Villamizar, L. Microencapsulated Spodoptera frugiperda nucleopolyhedrovirus: Insecticidal activity and effect on arthropod populations in maize. Biocontrol Sci. Technol. 2013, 23, 829–846. [Google Scholar] [CrossRef]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus insecticides in Latin America: Historical overview, current status and future perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef] [PubMed]
- CABI. Spodoptera frugiperda (Fall Armyworm). Invasive Species Compendium. CABI. 2020. Available online: https://www.cabi.org/isc/datasheet/29810 (accessed on 3 February 2021).
- Valicente, F.H. Consumo foliar da lagarta do cartucho do milho, Spodoptera frugiperda (J. E. Smith, 1797) infectada com vírus da granulose ou de poliedrose nuclear. An. Soc. Entomológica Bras. 1988, 17, 347–357. [Google Scholar] [CrossRef]
- Hamm, J.J.; Shapiro, M. Infectivity of fall armyworm (Lepidoptera: Noctuidae) nuclear polyhedrosis virus enhanced by a fluorescent brightener. J. Econ. Entomol. 1992, 85, 2149–2152. [Google Scholar] [CrossRef]
- Cisneros, J.; Pérez, J.A.; Penagos, D.I.; Ruiz, J.; Goulson, D.; Caballero, P.; Cave, R.D.; Williams, T. Formulation of a nucleopolyhedrovirus with boric acid for control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize. Biol. Control 2002, 23, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Alves, S.B.; Lopes, R.B.; Vieira, S.A.; Tamai, M.A. Fungos entomopatogênicos usados no controle de pragas na América Latina. In Controle Microbiano de Pragas na América Latina: Avanços e Desafios; Alves, S.B., Ed.; FEALQ: Piracicaba, Brazil, 2008; pp. 69–110. [Google Scholar]
- Sujeetha, J.A.R.P.; Sahayaraj, K. Role of Entomopathogenic Fungus in Pest Management. In Basic and Applied Aspects of Biopesticides; Sahayaraj, K., Ed.; Springer: New Delhi, India, 2014. [Google Scholar]
- Wraight, S.P.; Ramos, M.E.; Avery, P.B.; Jaronski, S.T.; Vandenberg, J.D. Comparative virulence of Beauveria bassiana isolates against Lepidopteran pests of vegetable crops. J. Invertebr. Pathol. 2010, 103, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Polanczyk, R.A.; Silva, R.F.P.D.; Fiuza, L.M. Effectiveness of Bacillus thuringiensis strains against Spodoptera frugiperda (Lepidoptera: Noctuidae). Braz. J. Microbiol. 2000, 31, 164–166. [Google Scholar] [CrossRef] [Green Version]
- De-Bortoli, C.P.; Jurat-Fuentes, J.L. Mechanisms of resistance to commercially relevant entomopathogenic bacteria. Curr. Opin. Insect Sci. 2019, 33, 56–62. [Google Scholar] [CrossRef]
- Silva, S.M.B.; Silva-Werneck, J.O.; Falcão, R.; Gomes, A.C.; Fragoso, R.R.; Quezado, M.T.; Neto, O.B.O.; Aguiar, J.B.; Sá, M.F.G.; Bravo, A.; et al. Characterization of novel Brazilian Bacillus thuringiensis strains active against Spodoptera frugiperda and other insect pests. J. Appl. Entomol. 2004, 128, 102–107. [Google Scholar] [CrossRef]
- Monnerat, R.; Martins, E.; Queiroz, P.; Ordúz, S.; Jaramillo, G.; Benintende, G.; Cozzi, J.; Real, M.D.; Martinez-Ramirez, A.; Rausell, C.; et al. Genetic variability of Spodoptera frugiperda (Lepidoptera: Noctuidae) populations from Latin America is associated with variations in susceptibility to Bacillus thuringiensis Cry toxins. Appl. Environ. Microbiol. 2006, 72, 7029–7035. [Google Scholar] [CrossRef] [Green Version]
- Fontana-Capalbo, D.M.; Valicente, F.H.; Oliveira, M.I.D.; Pelizer, L.H. Solid-state fermentation of Bacillus thuringiensis tolworthi to control fall armyworm in maize. Electron. J. Biotechnol. 2001, 4, 9–10. [Google Scholar] [CrossRef]
- Barreto, M.R.; Loguercio, L.L.; Valicente, F.H.; Paiva, E. Insecticidal activity of culture supernatants from Bacillus thuringiensis Berliner strains against Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) larvae. An. Soc. Entomol. Bras. 1999, 28, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Dillman, A.R.; Chaston, J.M.; Adams, B.J.; Ciche, T.A.; Goodrich-Blair, H.; Stock, S.P.; Sternberg, P.W. An Entomopathogenic Nematode by Any Other Name. PLoS Pathog. 2012, 8, e1002527. [Google Scholar] [CrossRef] [Green Version]
- Shapiro-Ilan, D.I.; Gouge, D.H.; Piggott, S.J.; Fife, J.P. Application technology and environmental considerations for use of entomopathogenic nematodes in biological control. Biol. Control 2006, 38, 124–133. [Google Scholar] [CrossRef]
- Garcia, L.C.; Raetano, C.G.; Leite, L.G. Application technology for the entomopathogenic nematodes Heterorhabditis indica and Steinernema sp. (Rhabditida: Heterorhabditidae and Steinernematidae) to control Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in corn. Neotrop. Entomol. 2008, 37, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Molina-Ochoa, J.; Lezama-Gutierrez, R.; Hamm, J.J.; Wiseman, B.R.; Lopez-Edwards, M. Integrated control of fall armyworm (Lepidoptera: Noctuidae) using resistant plants and entomopathogenic nematodes (Rhabditida: Steinernematidae). Fla. Entomol. 1999, 82, 263–271. [Google Scholar] [CrossRef]
- Negrisoli, A.S.; Garcia, M.S.; Negrisoli, C.R.C.B. Compatibility of entomopathogenic nematodes (Nematoda: Rhabditida) with registered insecticides for Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae) under laboratory conditions. Crop Prot. 2010, 29, 545–549. [Google Scholar] [CrossRef]
- Negrisoli, A.S.; Garcia, M.S.; Negrisoli, C.R.C.B.; Bernardi, D.; Da-Silva, A. Efficacy of entomopathogenic nematodes (Nematoda: Rhabditida) and insecticide mixtures to control Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae) in corn. Crop Prot. 2010, 29, 677–683. [Google Scholar] [CrossRef]
- Shah, F.M.; Razaq, M.; Ali, Q.; Ali, A.; Shad, S.A.; Aslam, M.; Hardy, I.C. Action threshold development in cabbage pest management using synthetic and botanical insecticides. Entomol. Gen. 2020, 40, 157–172. [Google Scholar] [CrossRef]
- Pavela, R.; Morshedloo, M.R.; Mumivand, H.; Khorsand, G.J.; Karami, A.; Maggi, F.; Desneux, N.; Benelli, G. Phenolic monoterpene-rich essential oils from Apiaceae and Lamiaceae species: Insecticidal activity and safety evaluation on non-target earthworms. Entomol. Gen. 2020, 40, 421–435. [Google Scholar] [CrossRef]
- Badshah, K.; Ullah, F.; Ahmad, B.; Ahmad, S.; Alam, S.; Ullah, M.; Jamil, M.; Sardar, S. Management of Lycoriella ingenua (Diptera: Sciaridae) on oyster mushroom (Pleurotus ostreatus) through different botanicals. Int. J. Trop. Insect Sci. 2021, 41, 1435–1440. [Google Scholar] [CrossRef]
- Ullah, M.; Ullah, F.; Khan, M.A.; Ahmad, S.; Jamil, M.; Sardar, S.; Tariq, K.; Ahmed, N. Efficacy of various natural plant extracts and the synthetic insecticide cypermethrin 25EC against Leucinodes orbonalis and their impact on natural enemies in brinjal crop. Int. J. Trop. Insect Sci. 2021, 42, 173–182. [Google Scholar] [CrossRef]
- Gul, H.; Ullah, F.; Biondi, A.; Desneux, N.; Qian, D.; Gao, X.; Song, D. Resistance against clothianidin and associated fitness costs in the chive maggot, Bradysia odoriphaga. Entomol. Gen. 2019, 39, 81–92. [Google Scholar] [CrossRef]
- Wang, X.; Xu, X.; Ullah, F.; Ding, Q.; Gao, X.; Desneux, N.; Song, D. Comparison of full-length transcriptomes of different imidacloprid-resistant strains of Rhopalosiphum padi (L.). Entomol. Gen. 2021, 41, 289–304. [Google Scholar] [CrossRef]
- Shan, J.; Zhu, B.; Gu, S.; Liang, P.; Gao, X. Development of resistance to chlorantraniliprole represses sex pheromone responses in male Plutella xylostella (L.). Entomol. Gen. 2021, 41, 615–625. [Google Scholar] [CrossRef]
- Paula, D.P.; Lozano, R.E.; Menger, J.P.; Andow, D.A.; Koch, R.L. Identification of point mutations related to pyrethroid resistance in voltage-gated sodium channel genes in Aphis glycines. Entomol. Gen. 2021, 41, 243–255. [Google Scholar] [CrossRef]
- Jirnmci, E. Efficacy of botanical extracts against termites, Macrotermes spp., (Lsoptera: Termiticlae) under laboratory conditions. Int. J. Agric. Res. 2014, 9, 60–73. [Google Scholar]
- Bateman, M.L.; Day, R.K.; Luke, B.; Edgington, S.; Kuhlmann, U.; Cock, M.J.W. Assessment of potential biopesticide options for managing fall armyworm (Spodoptera frugiperda) in Africa. J. Appl. Entomol. 2018, 142, 805–819. [Google Scholar] [CrossRef] [Green Version]
- Tavares, W.S.; Costa, M.A.; Cruz, I.; Silveira, R.D.; Serrão, J.E.; Zanuncio, J.C. Selective effects of natural and synthetic insecticides on mortality of Spodoptera frugiperda (Lepidoptera: Noctuidae) and its predator Eriopis connexa (Coleoptera: Coccinellidae). J. Environ. Sci. Health 2010, 45, 557–561. [Google Scholar] [CrossRef]
- Babendreier, D.; Agboyi, L.K.; Beseh, P.; Osae, M.; Nboyine, J.; Ofori, S.E.K.; Frimpong, J.O.; Attuquaye Clottey, V.; Kenis, M. The efficacy of alternative, environmentally friendly plant protection measures for control of fall armyworm, Spodoptera frugiperda, in maize. Insects 2020, 11, 240. [Google Scholar] [CrossRef]
- Hruska, A.J. Fall armyworm (Spodoptera frugiperda) management by smallholders. CAB Rev. 2019, 14, 43. [Google Scholar] [CrossRef] [Green Version]
- Maredia, K.M.; Segura, O.L.; Mihm, J.A. Effects of neem, Azadirachta indica on six species of maize insect pests. Trop. Pest Manag. 1992, 38, 190–195. [Google Scholar] [CrossRef]
- Silva, M.S.; Maria, S.; Broglio, F.; Cristina, R.; Trindade, P.; Ferrreira, E.S.; Gomes, I.B.; Micheletti, L.B. Toxicity and application of neem in fall armyworm. Comun. Sci. 2015, 6, 359–364. [Google Scholar] [CrossRef]
- Figueroa-Brito, R.; Villa-Ayala, P.; López-Olguín, J.F.; Peña, H.; Pacheco-Aguilar, J.R.; Ramos-López, M.A. Nitrogen fertilization sources and insecticidal activity of aqueous seeds extract of Carica papaya against Spodoptera frugiperda in maize. Cienc. Investig. Agrar. 2013, 40, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Mora, J.; Blanco-Metzler, H. Evaluation of botanical insecticides in controlling the population of fall armyworm (Spodoptera frugiperda) present on corn crops (Zea mays) located in Santa Cruz, Guanacate. IOP Conf. Ser. Earth Environ. Sci. 2018, 215, 012013. [Google Scholar] [CrossRef]
- Mikolajczak, K.L.; Zilkowski, B.W.; Bartelt, R.J. Effect of meliaceous seed extracts on growth and survival of Spodoptera frugiperda (J.E Smith). J. Chem. Ecol. 1989, 15, 121–128. [Google Scholar] [CrossRef]
- Ansante, T.F.; Prado Ribeiro, L.; Bicalho, K.U.; Fernandes, J.B.; Vieira, P.C.; Vendramim, J.D. Secondary metabolites from Neotropical Annonaceae: Screening, bioguided fractionation, and toxicity to Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Ind. Crops Prod. 2015, 74, 969–976. [Google Scholar] [CrossRef]
- Tavares, W.S.; Cruz, I.; Petacci, F.; De-Assis, J.S.L.; Sousa-Freitas, S.; Zanuncio, J.C.; Serrão, J.E. Potential use of Asteraceae extracts to control Spodoptera frugiperda (Lepidoptera: Noctuidae) and selectivity to their parasitoids Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) and Telenomus remus (Hymenoptera: Scelionidae). Ind. Crops Prod. 2009, 30, 384–388. [Google Scholar] [CrossRef]
- Céspedes, C.L.; Calderón, J.S.; Lina, L.; Aranda, E. Growth inhibitory effects on fall armyworm Spodoptera frugiperda of some limonoids isolated from Cedrela spp. (Meliaceae). J. Agric. Food Chem. 2000, 48, 1903–1908. [Google Scholar] [CrossRef]
- Céspedes, C.L.; Salazar, J.R.; Martínez, M.; Aranda, E. Insect growth regulatory effects of some extracts and sterols from Myrtillocactus geometrizans (Cactaceae) against Spodoptera frugiperda and Tenebrio molitor. Phytochemistry 2005, 66, 2481. [Google Scholar] [CrossRef]
- Alves, T.J.S.; Cruz, G.S.; Wanderley-Teixeira, V.; Teixeira, A.A.C.; Oliveira, J.V.; Correia, A.A.; Câmara, C.A.G.; Cunha, F.M. Effects of Piper hispidinervum on spermatogenesis and histochemistry of ovarioles of Spodoptera frugiperda. Biotech. Histochem. 2014, 89, 245–255. [Google Scholar] [CrossRef]
- Bullangpoti, V.; Wajnberg, E.; Audant, P.; Feyereisen, R. Antifeedant activity of Jatropha gossypifolia and Melia azedarach senescent leaf extracts on Spodoptera frugiperda (Lepidoptera: Noctuidae) and their potential use as synergists. Pest Manag. Sci. 2012, 68, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Ramos-López, M.A.; Pérez, S.; Rodríguez-Hernández, G.C.; Guevara-Fefer, P.; Zavala-Sanchez, M.A. Activity of Ricinus communis (Euphorbiaceae) against Spodoptera frugiperda (Lepidoptera: Noctuidae). Afr. J. Biotechnol. 2010, 9, 1359–1365. [Google Scholar]
- Nboyine, J.A.; Kusi, F.; Abudulai, M.; Badii, B.K.; Zakaria, M.; Adu, G.B.; Haruna, A.; Seidu, A.; Osei, V.; Alhassan, S.; et al. A new pest, Spodoptera frugiperda (J.E. Smith), in tropical Africa: Its seasonal dynamics and damage in maize fields in northern Ghana. Crop Prot. 2020, 127, 104960. [Google Scholar] [CrossRef]
- Hailu, G.; Niassy, S.; Zeyaur, K.R.; Ochatum, N.; Subramanian, S. Maize-legume intercropping and push–pull for management of fall armyworm, stemborers, and striga in Uganda. Agronomy 2018, 110, 2513–2522. [Google Scholar] [CrossRef] [Green Version]
- Schmutterer, H. Which insect pests can be controlled by application of neem seed kernel extracts under field conditions? Z. Angew. Entomol. 1985, 100, 468–475. [Google Scholar] [CrossRef]
- Martínez, A.M.; Aguado-Pedraza, A.J.; Viñuela, E.; Rodríguez-Enríquez, C.L.; Lobit, P.; Gómez, B.; Pineda, S. Effects of ethanolic extracts of Argemone ochroleuca (Papaveraceae) on the food consumption and development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 2017, 100, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Batista-Pereira, L.G.; Stein, K.; Paula, A.F.; Moreira, J.A.; Cruz, I.; Figueiredo, M.L.C.; Perri, J.; Corrêa, A.G. Isolation, identification, synthesis and field evaluation of the sex pheromone of the Brazilian population of Spodoptera frugiperda. J. Chem. Ecol. 2006, 32, 1085–1099. [Google Scholar] [CrossRef]
- Isman, M.B. Neem and other botanical insecticides: Barriers to commercialization. Phytoparasitica 1997, 25, 339–344. [Google Scholar] [CrossRef]
- Ogendo, J.O.; Deng, A.L.; Omollo, E.O.; Matasyoh, J.C.; Tuey, R.K.; Khan, Z.R. Management of stem borers using selected botanical pesticides in a maize-bean cropping system. Egerton J. Sci. Technol. 2013, 13, 21–38. [Google Scholar]
- Mugisha, K.M.; Deng, A.L.; Ogendo, J.O.; Omolo, E.O.; Mihale, M.J.; Otim, M.; Buyungo, J.P.; Bett, P.K. Indigenous knowledge of field insect pests and their management around Lake Victoria basin in Uganda. Afr. J. Environ. Sci. Technol. 2008, 2, 342–348. [Google Scholar]
- Stevenson, P.C.; Isman, M.B.; Belmain, S.R. Pesticidal plants in Africa: A global vision of new biological control products from local uses. Ind. Crops Prod. 2017, 110, 2–9. [Google Scholar] [CrossRef]
Figure 1.
Biological control of fall armyworm (FAW), Spodoptera frugiperda.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Abbas, A.; Ullah, F.; Hafeez, M.; Han, X.; Dara, M.Z.N.; Gul, H.; Zhao, C.R. Biological Control of Fall Armyworm, Spodoptera frugiperda. Agronomy 2022, 12, 2704. https://doi.org/10.3390/agronomy12112704
AMA Style
Abbas A, Ullah F, Hafeez M, Han X, Dara MZN, Gul H, Zhao CR. Biological Control of Fall Armyworm, Spodoptera frugiperda. Agronomy. 2022; 12(11):2704. https://doi.org/10.3390/agronomy12112704
Chicago/Turabian StyleAbbas, Arzlan, Farman Ullah, Muhammad Hafeez, Xiao Han, Muhammad Zulqar Nain Dara, Hina Gul, and Chen Ri Zhao. 2022. "Biological Control of Fall Armyworm, Spodoptera frugiperda" Agronomy 12, no. 11: 2704. https://doi.org/10.3390/agronomy12112704
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.