
Effect of Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Infestation on the Phenolic Response and Quality of Olive Fruits (Olea europaea L.)


Abstract
:1. Introduction
2. Materials and Methods
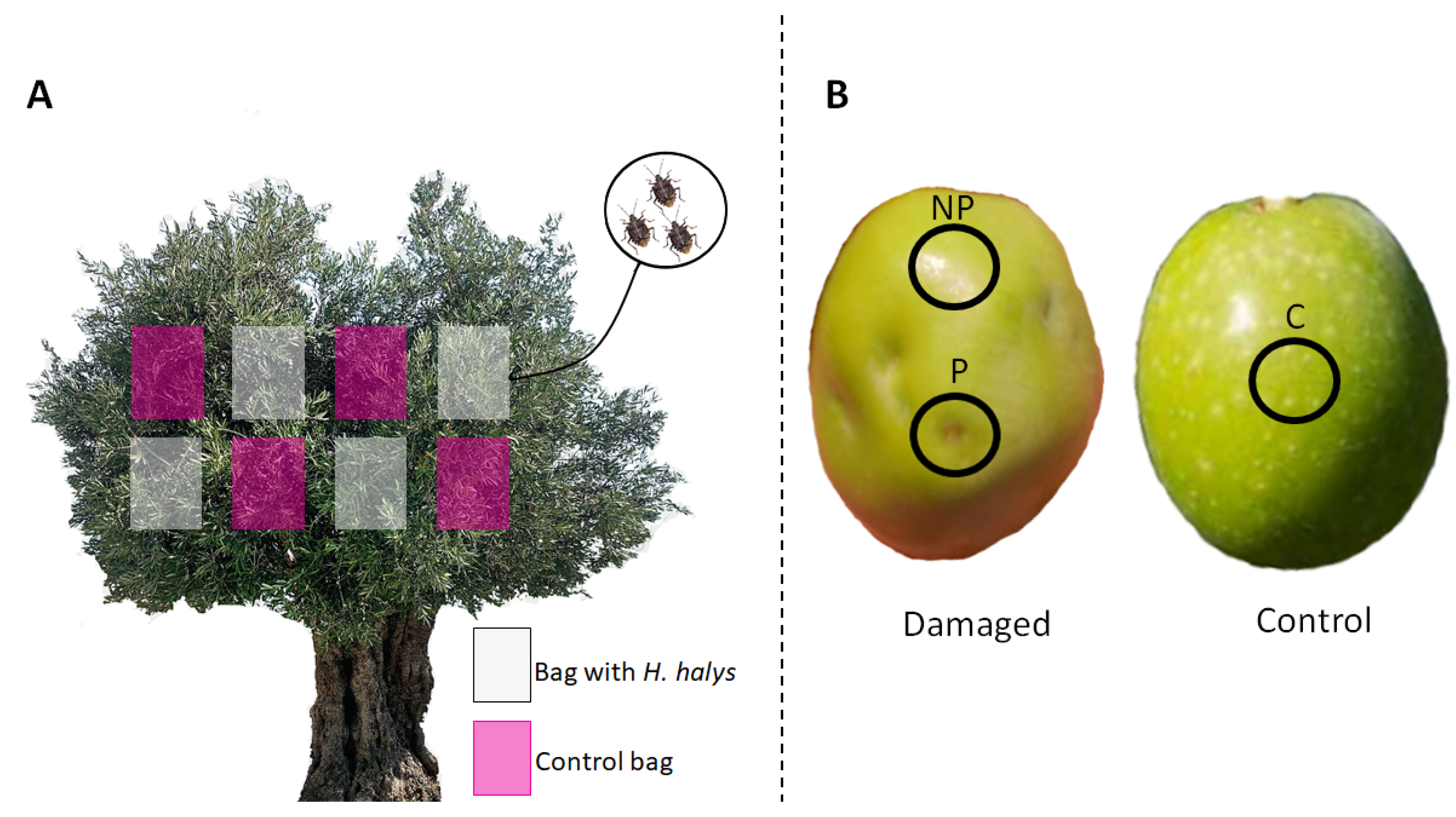
2.1. Plant Materials and Treatments
2.2. Color of the Fruits
2.3. Extraction of TPC, AC, and Individual Phenolic Compounds
2.4. Analysis of Individual Phenolic Compounds
2.5. Analysis of the Total Phenolic Content (TPC)
2.6. Analysis of Antioxidant Capacity (AC)
2.7. Chemicals and Standards
2.8. Statistical Analysis
3. Results
3.1. Visual Appearance, Color, Weight, and Firmness of Olive Fruits
3.2. Total Phenolic Content and Antioxidant Capacity
3.3. Individual Phenolic Compounds
3.3.1. ‘Istrska belica’
3.3.2. ‘Pendolino’
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IOC-International Olive Council. Huiles D’Olive—Olive Oils Consumption. Sensory Analysis of Table Olives. 2021. Available online: https://www.internationaloliveoil.org/wp-content/uploads/2021/12/HO-W901-17-12-2021-C.pdf?fbclid=IwAR3g3QZFnXNUMnHzeJXWUBfvZWk_5e7Oi1iVHeqtvPrNEt739aKhCEDoaPs (accessed on 8 April 2022).
- Spadafora, A.; Mazzuca, S.; Fiorella Chiappetta, F.; Parise, A.; Perri, E.; Innocenti, A.M. Oleuropein Specific Glucosidase Activity Marks the Early Response of Olive Fruits (Olea europaea L.) to Mimed Insect Attack. Agric. Sci. China 2008, 7, 703–712. [Google Scholar] [CrossRef]
- Malheiro, R.; Sousa, A.; Casal, S.; Bento, A.; Pereira, J.A. Cultivar Effect on the Phenolic Composition and Antioxidant Potential of Stoned Table Olives. Food Chem. Toxicol. 2011, 49, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Poyatos, M.D.P.; Llorent-Martínez, E.J.; Ruiz-Medina, A. Effect of Ripening on the Phenolic Composition and Mineral Content of Three Varieties of Olive Fruits. Foods 2021, 10, 380. [Google Scholar] [CrossRef] [PubMed]
- Damos, P.; Soulopoulou, P.; Thomidis, T. First Record and Current Status of the Brown Marmorated Sting Bug Halyomorpha Halys Damaging Peaches and Olives in Northern Greece. J. Plant Prot. Res. 2020, 60, 323–326. [Google Scholar] [CrossRef]
- CABI. Invasive Species Compendium. Halyomorpha halys (Brown Marmorated Stink Bug). 2022. Available online: https://www.cabi.org/isc/datasheet/27377 (accessed on 8 April 2022).
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, Ecology, and Management of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). J. Integr. Pest. Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Rot, M.; Devetak, M.; Carlevaris, B.; Žežlina, J.; Žežlina, I. First Record of Brown Marmorated Stink Bug (Halyomorpha Halys (Stål, 1855)) (Hemiptera: Pentatomidae) in Slovenia. Acta Entomol. Sloven. 2018, 26, 5–12. [Google Scholar]
- Zamljen, T.; Medič, A.; Veberič, R.; Hudina, M.; Štampar, F.; Slatnar, A. Apple Fruit (Malus Domestica Borkh.) Metabolic Response to Infestation by Invasive Brown Marmorated Stink Bug (Halyomorpha Halys Stal.). Horticulturae 2021, 7, 212. [Google Scholar] [CrossRef]
- Kehrli, P.; Rösti, J.; Lorenzini, F.; Deneulin, P.; Linder, C. Influence of Processed Halyomorpha Halys Bugs on the Aroma and Taste of “Chardonnay” and “Merlot” Musts and Wines. Vitis-J. Grapevine Res. 2021, 60, 43–50. [Google Scholar] [CrossRef]
- Sagun, S.; Collins, E.; Martin, C.; Nolan, J.; Horzempa, J. Alarm Odor Compounds of the Brown Marmorated Stink Bug Exhibit Antibacterial Activity. J. Pharmacogn. Nat. Prod. 2016, 2, 119. [Google Scholar] [CrossRef]
- Wiman, N.G.; Parker, J.E.; Rodriguez-Saona, C.; Walton, V.M. Characterizing Damage of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) in Blueberries. J. Econ. Entomol. 2015, 108, 1156–1163. [Google Scholar] [CrossRef]
- Schumm, Z.R.; Alston, D.G.; Spears, L.R.; Manlove, K. Impact of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) Feeding on Tart Cherry (Rosales: Rosaceae) Quality and Yield in Utah. J. Econ. Entomol. 2020, 113, 2328–2334. [Google Scholar] [CrossRef] [PubMed]
- American/Apple Association. Brown Marmorated Stink Bug Causes $37 Million in Losses to Mid-Atlantic Apple Growers. 2011. Available online: http://www.growingproduce.com/news/afg/index.php?storyid=5372 (accessed on 5 March 2022).
- Yactayo-Chang, J.P.; Tang, H.V.; Mendoza, J.; Christensen, S.A.; Block, A.K. Plant Defense Chemicals against Insect Pests. Agronomy 2020, 10, 1156. [Google Scholar] [CrossRef]
- Korth, K.L. Profiling the Response of Plants to Herbivorous Insects. Genome Biol. 2003, 4, 221. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of Phenolics in the Resistance Mechanisms of Plants against Fungal Pathogens and Insects. Phytochem. Adv. Res. 2006, 661, 23–67. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of Plant Defense against Insect Herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Aghaee, M.A.; Lee Rice, S.; Milnes, J.M.; Goding, K.M.; Godfrey, L.D. Is Halyomorpha Halys (Hemiptera: Pentatomidae) a Threat to California Rice? Crop. Prot. 2018, 111, 1–5. [Google Scholar] [CrossRef]
- Patel, D.T.; Stout, M.J.; Fuxa, J.R. Effects of Rice Panicle Age on Quantitative and Qualitative Injury by the Rice Stink Bug (Hemiptera: Pentatomidae). Fla. Entomol. 2006, 89, 321–327. [Google Scholar] [CrossRef]
- Cody Holthouse, M.; Alston, D.G.; Spears, L.R.; Petrizzo, E. Brown Marmorated Stink Bug [Halyomorpha Halys (Stal)]. Utah Pest Fact Sheet 2017, 1, 1–8. [Google Scholar]
- Weber, N.C.; Razinger, J.; Jakopič, J.; Schmitzer, V.; Hudina, M.; Slatnar, A.; Veberič, R.; Štampar, F.; Zamljen, T. Brown Marmorated Stink Bug (Halyomorpha Halys Stål.) Attack Induces a Metabolic Response in Strawberry (Fragaria × Ananassa Duch.) Fruit. Horticulturae 2021, 7, 561. [Google Scholar] [CrossRef]
- Fernando Reyes, L.; Emilio Villarreal, J.; Cisneros-Zevallos, L. The Increase in Antioxidant Capacity after Wounding Depends on the Type of Fruit or Vegetable Tissue. Food Chem. 2007, 101, 1254–1262. [Google Scholar] [CrossRef]
- Arshiya, S. The Antioxidant Effect of Certain Fruits—A Review. J. Pharm. Sci. Res. 2013, 5, 265–268. [Google Scholar]
- Rashid War, A.; Sharma, H.C.; Paulraj, M.G.; War, Y.; Ignacimuthu, S. Plant Signaling & Behavior Herbivore Induced Plant Volatiles: Their Role in Plant Defense for Pest Management. Plant Signal. Behav. 2011, 6, 1973–1978. [Google Scholar] [CrossRef]
- Zhou, Y.; Giusti, M.M.; Parker, J.; Salamanca, J.; Rodriguez-Saona, C. Frugivory by Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) Alters Blueberry Fruit Chemistry and Preference by Conspecifics. Environ. Entomol. 2016, 45, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Zamljen, T.; Veberic, R.; Hudina, M.; Slatnar, A. The Brown Marmorated Stink Bug (Halyomorpha Halys Stål.) Influences Pungent and Non-Pungent Capsicum Cultivars’ Pre-and Post-Harvest Quality. Agronomy 2021, 11, 2252. [Google Scholar] [CrossRef]
- Soriano, I.R.; Asenstorfer, R.E.; Schmidt, O.; Riley, I.T. Inducible Flavone in Oats (Avena Sativa) Is a Novel Defense against Plant-Parasitic Nematodes. Phytopathology 2004, 94, 1207–1214. [Google Scholar] [CrossRef]
- Notario, A.; Sánchez, R.; Luaces, P.; Sanz, C.; Pérez, A.G. The Infestation of Olive Fruits by Bactrocera Oleae (Rossi) Modifies the Expression of Key Genes in the Biosynthesis of Volatile and Phenolic Compounds and Alters the Composition of Virgin Olive Oil. Molecules 2022, 27, 1650. [Google Scholar] [CrossRef]
- Medjkouh, L.; Tamendjari, A.; Alves, R.C.; Araújo, M.; Oliveira, M.B.P.P. Effect of: Bactrocera Oleae on Phenolic Compounds and Antioxidant and Antibacterial Activities of Two Algerian Olive Cultivars. Food Funct. 2016, 7, 4372–4378. [Google Scholar] [CrossRef]
- Valenčič, V.; Butinar, B.; Podgornik, M.; Bučar-Miklavčič, M. The Effect of Olive Fruit Fly Bactrocera Oleae (Rossi) Infestation on Certain Chemical Parameters of Produced Olive Oils. Molecules 2021, 26, 95. [Google Scholar] [CrossRef]
- Medjkouh, L.; Tamendjari, A.; Alves, R.C.; Laribi, R.; Oliveira, M.B.P.P. Phenolic Profiles of Eight Olive Cultivars from Algeria: Effect of: Bactrocera Oleae Attack. Food Funct. 2018, 9, 890–897. [Google Scholar] [CrossRef]
- Cui, M.; Chen, B.; Xu, K.; Rigakou, A.; Diamantakos, P.; Melliou, E.; Logothetis, D.E.; Magiatis, P. Activation of Specific Bitter Taste Receptors by Olive Oil Phenolics and Secoiridoids. Sci. Rep. 2021, 11, 22340. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.J.; Huang, L.X.; Zhang, C.H.; Zhang, Y.L. Phenolic Compositions, and Antioxidant Performance of Olive Leaf and Fruit (Olea Europaea L.) Extracts and Their Structure-Activity Relationships. J. Funct. Foods 2015, 16, 460–471. [Google Scholar] [CrossRef]
- Hamouda, A.; Boussadia, O.; Khaoula, B.; Laarif, A.; Braham, M. Studies on Insecticidal and Deterrent Effects of Olive Leaf Extracts on Myzus Persicae and Phthorimaea Operculella. Artic. J. Entomol. Zool. Stud. 2015, 3, 294–297. [Google Scholar]
- Lo Scalzo, R.; Scarpati, M.L.; Verzegnassi, B.; Vita, G. Olea Europaea Chemicals Repellent to Dacus Oleae Females. J. Chem. Ecol. 1994, 20, 1813–1823. [Google Scholar] [CrossRef]
- Goławska, S.; Kapusta, I.; Łukasik, I. Effect of Phenolics on the Pea Aphid, Acyrthosiphon Pisum (Harris) Population on Pisum Sativum L. (Fabaceae). Pestycydy 2008, 3, 71–77. [Google Scholar]
- Guggenheim, K.G.; Crawford, L.M.; Paradisi, F.; Wang, S.C.; Siegel, J.B. β-Glucosidase Discovery and Design for the Degradation of Oleuropein. ACS Omega 2018, 3, 15754–15762. [Google Scholar] [CrossRef]
- Gucci, R.; Caruso, G.; Canale, A.; Loni, A.; Raspi, A.; Urbani, S.; Taticchi, A.; Esposto, S.; Servili, M. Qualitative Changes of Olive Oils Obtained from Fruits Damaged by Bactrocera Oleae (Rossi). HortScience 2012, 47, 301–306. [Google Scholar] [CrossRef]
- Ramírez, E.; Brenes, M.; García, P.; Medina, E.; Romero, C. Oleuropein Hydrolysis in Natural Green Olives: Importance of the Endogenous Enzymes. Food Chem. 2016, 206, 204–209. [Google Scholar] [CrossRef]
- Tamendjari, A.; Angerosa, F.; Mettouchi, S.; Bellal, M.M. The Effect of Fly Attack (Bactrocera Oleae) on the Quality and Phenolic Content of Chemlal Olive Oil. Grasas Aceites. 2009, 60, 507–513. [Google Scholar] [CrossRef]
- Britton, J.; Davis, R.; O’Connor, K.E. Chemical, Physical and Biotechnological Approaches to the Production of the Potent Antioxidant Hydroxytyrosol. Appl. Microbiol. Biotechnol. 2019, 103, 5957–5974. [Google Scholar] [CrossRef]
- Macoy, D.M.; Kim, W.Y.; Lee, S.Y.; Kim, M.G. Biosynthesis, Physiology, and Functions of Hydroxycinnamic Acid Amides in Plants. Plant Biotechnol. Rep. 2015, 9, 269–278. [Google Scholar] [CrossRef]
- Cardinali, A.; Pati, S.; Minervini, F.; D’Antuono, I.; Linsalata, V.; Lattanzio, V. Verbascoside, Isoverbascoside, and Their Derivatives Recovered from Olive Mill Wastewater as Possible Food Antioxidants. J. Agric. Food Chem. 2012, 60, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, H.M.; Rentzsch, M.; Breuer, M. Anthocyanins Reduce Fungal Growth in Fruits. Nat. Prod. Commun. 2008, 3, 1267–1272. [Google Scholar] [CrossRef]
- Garcıa-Villalba, R.; Larrosa, M.; Possemiers, S.; Tomas-Barberan, F.A.; Espin, J.C. Bioavailability of phenolics from an oleuropein-rich olive (Olea europaea) leaf extract and its acute effect on plasma antioxidant status: Comparison between pre- and postmenopausal women. Eur. J. Nutr. 2014, 53, 1015–1027. [Google Scholar] [CrossRef]
- Fayek, N.M.; Farag, M.A.; Saber, F.R. Metabolome classification via GC/MS and UHPLC/MS of olive fruit varieties grown in Egypt reveal pickling process impact on their composition. Food Chem. 2021, 339, 127861. [Google Scholar] [CrossRef]
- Alañón, M.E.; Ivanović, M.; Gómez-Caravaca, A.M.; Arráez-Román, D.; Segura-Carretero, A. Choline chloride derivative-based deep eutectic liquids as novel green alternative solvents for extraction of phenolic compounds from olive leaf. Arab. J. Chem. 2020, 13, 1685–1701. [Google Scholar] [CrossRef]
- Michel, T.; Khlifc, I.; Kanakis, P.; Termentzi, A.; Allouche, N.; Halabalaki, M.; Skaltsounis, A.L. UHPLC-DAD-FLD and UHPLC-HRMS/MS based metabolic profiling and characterization of different Olea europaea organs of Koroneiki and Chetoui varieties. Phytol. J. 2015, 11, 424–439. [Google Scholar] [CrossRef]
- Li, H.; Yao, W.; Liu, Q.; Xu, J.; Bao, B.; Shan, M.; Cao, Y.; Cheng, F.; Ding, A.; Zhang, L. Application of UHPLC-ESI-Q-TOF-MS to identify multiple constituents in processed products of the herbal medicine ligustri lucidi fructus. Molecules 2017, 22, 689. [Google Scholar] [CrossRef] [PubMed]
- Šuković, D.; Knežević, B.; Gašić, U.; Todić, S.; Mutić, J.; Tešić, Ž. Phenolic profiles of leaves, grapes and wine of grapevine variety Vranac (Vitis vinifera L.) from Montenegro. Foods 2020, 9, 138. [Google Scholar] [CrossRef]
- Ivancic, T.; Jakopic, J.; Veberic, R.; Vesel, V.; Hudina, M. Effect of ripening on the phenolic and sugar contents in the meso- and epicarp of olive fruits (Olea europaea L.) cultivar ‘Leccino’. Agriculture 2022, 12, 1347. [Google Scholar] [CrossRef]


{kind=link}
| Color | Weight (g) | Firmness (N) | |||||
|---|---|---|---|---|---|---|---|
| L | a | b | C | h° | |||
| ‘Istrska belica’ | |||||||
| Control | 51.86 ± 2.80 | −4.86 ± 0.79 | 38.32 ± 2.41 | 38.42 ± 2.52 | 97.30 ± 1.36 | 2.58 ± 0.36 | 6.02 ± 2.06 |
| Damaged | 50.41 ± 2.35 | −5.09 ± 0.76 | 37.14 ± 3.33 | 38.42 ± 2.52 | 97.76 ± 1.09 | 2.01 ± 0.31 | 6.54 ± 3.19 |
| Significance | * | ns | ns | ns | ns | *** | ns |
| ‘Pendolino’ | |||||||
| Control | 23.26 ± 2.17 | 4.68 ± 2.92 | 2.37 ± 2.50 | 5.34 ± 3.74 | 23.66 ± 10.64 | 1.63 ± 0.21 | 3.70 ± 1.53 |
| Damaged | 22.56 ± 2.38 | 3.90 ± 2.20 | 2.96 ± 4.20 | 4.52 ± 2.83 | 28.55 ± 12.15 | 1.47 ± 0.24 | 4.69 ± 1.14 |
| Significance | ns | ns | ns | ns | ns | ** | ** |
| TPC | AC | |
|---|---|---|
| (mg GAE kg−1 FW) | (umol Trolox kg−1 FW) | |
| ‘Istrska belica’ | ||
| C | 2212.28 ± 175.87 a | 27.83 ± 0.54 a |
| NP | 2210.66 ± 49.76 a | 29.81 ± 1.34 b |
| P | 2449.98 ± 71.60 b | 29.53 ± 0.12 b |
| ‘Pendolino’ | ||
| C | 1734.98 ± 128.93 a | 26.36 ± 0.92 a |
| NP | 1928.85 ± 44.98 a | 27.59 ± 1.81 a |
| P | 1873.70 ± 102.32 a | 27.42 ± 0.71 a |
| Phenolic Compound | C | NP | P | |
|---|---|---|---|---|
| Total flavones | 16.57 ± 2.64 a | 17.26 ± 1.32 a | 20.08 ± 1.69 b | |
| Luteoline 7-O-glucoside | 3.91 ± 0.81 a | 4.47 ± 0.97 a | 6.16 ± 0.55 b | |
| Luteoline 7-O-rutinoside | 12.66 ± 3.00 a | 12.79 ± 3.48 a | 13.92 ± 1.50 a | |
| Total flavonols | 14.64 ± 1.48 a | 12.95 ± 2.56 a | 15.93 ± 1.04 b | |
| Kaempferol 7-O-(6″rhamnosyl) hexoside | 1.23 ± 0.25 a | 1.64 ± 0.26 a | 2.23 ± 0.31 b | |
| Quercetin 3-O-rhamnoside | 4.56 ± 0.32 b | 3.00 ± 0.90 a | 3.41 ± 0.31 a | |
| Quercetin 3-O-rutinoside | 8.85 ± 1.35 a | 8.30 ± 1.56 a | 10.29 ± 0.77 b | |
| Total hidroxycinnamic acids | 19.57 ± 2.53 a | 17.36 ± 0.93 a | 18.17 ± 1.88 a | |
| Caffeoil-6′-secologanoside | 6.37 ± 1.04 a | 6.04 ± 0.61 a | 5.47 ± 0.80 a | |
| p-Coumaric acid glucoside | 0.31 ± 0.058 a | 0.31 ± 0.02 a | 0.49 ± 0.08 b | |
| Verbascoside | 8.12 ± 0.98 b | 4.86 ± 0.66 a | 4.15 ± 0.50 a | |
| β-Hydroxy-verbascoside | 4.77 ± 1.42 a | 5.14 ± 2.49 a | 8.06 ± 0.96 b | |
| Total secoiridoids | 1681.49 ± 62.10 a | 1970.80 ± 207.82 a | 2410.56 ± 105.23 b | |
| 2″Methoxyoleuropein isomer | 67.58 ± 8.60 a | 85.60 ± 16.83 a | 71.39 ± 5.32 a | |
| 6-Deoxyhexopyranosyl-oleoside | 54.93 ± 10.16 a | 50.53 ± 10.61 a | 49.61 ± 10.59 a | |
| Acyclodihydroelenolic acid hexoside isomers ˹ | 0.37 ± 0.08 b | 0.27 ± 0.05 a | 0.22 ± 0.05 a | |
| Demethyloleuropein | 12.21 ± 2.64 a | 24.91 ± 13.49 a | 12.07 ± 0.85 a | |
| Elenolic acid | 3.03 ± 0.57 a | 3.095 ± 0.21 a | 4.80 ± 0.33 b | |
| Elenolic acid glucoside isomers ˹ | 2.18 ± 0.25 b | 1.91 ± 0.17 a | 2.18 ± 0.14 b | |
| Hydroxyoleuropein | 2.09 ± 0.15 a | 2.87 ± 0.46 b | 2.08 ± 0.14 a | |
| hydroxytyrosol | 5.45 ± 0.25 c | 1.76 ± 0.24 a | 2.90 ± 0.64 b | |
| Hydroxytyrosol glucoside | 2.66 ± 0.20 a | 3.54 ± 0.69 a | 3.47 ± 0.90 a | |
| Secologanoside isomers ˹ | 10.73 ± 1.11 b | 9.03 ± 0.54 a | 9.77 ± 0.73 ab | |
| Oleuropein | 937.78 ± 74.84 a | 1288.32 ± 219.86 b | 1712.84 ± 123.69 c | |
| Oleuropein aglycone derivatives | 10.18 ± 2.80 c | 8.04 ± 1.37 b | 6.91 ± 0.64 a | |
| Oleuropein aglycone isomers ˹ | 528.10 ± 77.30 b | 449.29 ± 68.10 ab | 492.71 ± 68.73 a | |
| Oleuropein glucoside isomer | 24.78 ± 2.55 a | 27.25 ± 2.28 a | 27.20 ± 1.12 a | |
| Oleuropein isomer | 19.42 ± 0.96 b | 14.35 ± 1.51 a | 14.07 ± 0.87 a | |
| Other | ||||
| Tormentic acid derivatives | 0.61 ± 0.22 a | 0.55 ± 0.17 a | 0.54 ± 0.14 a | |
| Phenolic Compound | C | NP | P |
|---|---|---|---|
| Total flavones | 298.2 ± 32.74 a | 462.61 ± 41.24 b | 574.61 ± 55.32 c |
| Apigenin 7-O-glucoside | 71.32 ± 6.10 a | 103.44 ± 8.28 b | 135.46 ± 38.21 b |
| Luteoline 7-O-glucoside | 171.52 ± 25.39 a | 286.77 ± 33.84 b | 359.74 ± 36.58 c |
| Luteoline 7-O-rutinoside | 55.36 ± 6.85 a | 72.41 ± 3.54 b | 79.40 ± 11.21 b |
| Total flavonols | 131.02 ± 7.43 a | 180.94 ± 13.57 b | 207.33 ± 41.65 b |
| Kaempferol 7-O-(6″rhamnosyl) hexoside | 17.33 ± 2.49 a | 21.86 ± 2.07 b | 21.98 ± 1.39 b |
| Quercetin 3-O-rhamnoside | 77.77 ± 7.11 a | 112.79 ± 13.17 ab | 147.71 ± 41.66 b |
| Quercetin 3-O-rutinoside | 35.91 ± 3.93 a | 46.29 ± 2.55 b | 37.64 ± 2.91 a |
| Total hidroxycinnamic acids | 145.01 ± 7.62 c | 113.02 ± 11.27 a | 139.55 ± 8.62 b |
| Chlorogenic acid | 12.56 ± 0.97 c | 3.57 ± 0.45 a | 6.82 ± 0.95 b |
| Cryptochlorogenic acid | 3.84 ± 0.40 a | 2.30 ± 0.64 a | 5.34 ± 0.33 b |
| p-Coumaric acid glucoside | 0.64 ± 0.08 a | 0.47 ± 0.08 a | 1.38 ± 0.21 b |
| β-Hydroxy-verbascoside | 60.58 ± 2.86 ab | 54.84 ± 4.43 a | 66.73 ± 4.53 b |
| Verbascoside | 67.37 ± 2.33 b | 51.84 ± 5.67 a | 59.28 ± 3.62 a |
| Total secoiridoids | 4498.62 ± 191.67 a | 4668.03 ± 144.17 b | 4650.74 ± 99.84 b |
| 6-Deoxyhexopyranosyl-oleoside | 557.99 ± 45.89 a | 583.77 ± 60.17 a | 586.92 ± 59.60 a |
| Elenolic acid glucoside isomer ˹ | 5.64 ± 0.32 b | 6.11 ± 0.79 b | 4.43 ± 0.48 a |
| Hydroxyoleuropein | 224.23 ± 16.46 b | 197.05 ± 7.98 a | 196.37 ± 25.58 a |
| Hydroxytyrosol | 54.47 ± 4.18 b | 49.09 ± 9.88 ab | 40.50 ± 2.54 a |
| Hydroxytyrosol glucoside | 60.79 ± 6.99 b | 98.42 ± 10.95 c | 43.24 ± 6.75 a |
| Methoxyoleuropein isomer | 29.54 ± 3.85 a | 38.38 ± 4.38 b | 33.32 ± 6.32 b |
| Oleacein | 246.14 ± 32.07 a | 319.79 ± 36.47 b | 277.68 ± 52.64 b |
| Oleoside | 248.10 ± 16.95 a | 232.56 ± 41.41 a | 209.93 ± 35.55 a |
| Oleuropein | 646.96 ± 39.62 a | 784.27 ± 164.36 b | 936.26 ± 122.95 c |
| Oleuropein aglycone derivatives | 63.61 ± 4.42 c | 42.30 ± 6.69 b | 32.88 ± 2.68 a |
| Oleuropein aglycone isomers ˹ | 1709.08 ± 163.74 a | 1628.76 ± 253.70 a | 1645.28 ± 161.71 a |
| Oleuropein glucoside isomer | 346.32 ± 39.02 a | 348.13 ± 34.81 a | 327.55 ± 60.06 a |
| Oleuropein isomer | 272.40 ± 41.16 a | 334.62 ± 18.74 b | 314.19 ± 43.01 b |
| Tyrosol glucoside | 1.51 ± 0.33 a | 4.78 ± 2.13 b | 2.18 ± 0.42 b |
| Total anthocyanins | 330.93 ± 64.03 a | 521.04 ± 91.40 b | 596.38 ± 76.94 c |
| Cyanidin 3-O-glucoside | 29.41 ± 6.53 a | 57.65 ± 10.53 b | 59.62 ± 3.77 b |
| Cyanidin 3-O-rutinoside | 301.45 ± 59.39 a | 463.39 ± 81.46 b | 537.86 ± 77.85 c |
| Other | |||
| Tormentic acid derivatives | 15.00 ± 1.59 a | 16.23 ± 2.79 a | 13.03 ± 1.87 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivancic, T.; Grohar, M.C.; Jakopic, J.; Veberic, R.; Hudina, M. Effect of Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Infestation on the Phenolic Response and Quality of Olive Fruits (Olea europaea L.). Agronomy 2022, 12, 2200. https://doi.org/10.3390/agronomy12092200
Ivancic T, Grohar MC, Jakopic J, Veberic R, Hudina M. Effect of Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Infestation on the Phenolic Response and Quality of Olive Fruits (Olea europaea L.). Agronomy. 2022; 12(9):2200. https://doi.org/10.3390/agronomy12092200
Chicago/Turabian StyleIvancic, Tea, Mariana Cecilia Grohar, Jerneja Jakopic, Robert Veberic, and Metka Hudina. 2022. "Effect of Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Infestation on the Phenolic Response and Quality of Olive Fruits (Olea europaea L.)" Agronomy 12, no. 9: 2200. https://doi.org/10.3390/agronomy12092200