

Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland


1
Gaolan Ecological and Agricultural Research Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
2
Naiman Desertification Research Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(9), 2131; https://doi.org/10.3390/agronomy12092131
Submission received: 10 August 2022
/
Revised: 31 August 2022
/
Accepted: 5 September 2022
/
Published: 8 September 2022
(This article belongs to the Special Issue Impact of Grazing on Sustainability of Rangelands under Changing Climate)
Abstract
:Seedlings are a crucial part of the plant community and the basis of vegetation restoration and regeneration. Grazing and climate change are two main factors contributing to vegetation changes in semiarid grasslands. However, the effects of grazing on seedling emergence with rainfall variation are not clearly understood. This study aimed to address how seedling abundance and the emergence dynamics of annuals respond to grazing with rainfall changes in different years. We conducted a seedling investigation experiment in a grassland along a grazing intensity gradient (light grazing—LG; moderate grazing—MG; heavy grazing—HG) in contrasting wet and dry years. The results showed that MG increased the seedling richness of grasses and forbs in the wet year, but increasing grazing resulted in a decline in forb richness in the dry year. Seedling abundance of grasses was enhanced under MG, while seedling abundance of forbs was higher under HG. The monthly emergence dynamics were generally in a single peak pattern, but the peak time of seedling emergence for forbs advanced under HG. Seedling abundance of whole annuals increased with grazing in the wet year but was much higher under MG in the dry year. Our results suggest that more judicious grazing management based on rainfall variation and functional composition of the plant community should be undertaken to facilitate grassland restoration.
1. Introduction
Grazing has been identified as one of the main drivers of vegetation changes in grasslands, resulting in variations in species composition, fluctuations in plant abundance, and production [1,2,3,4]. Vegetation degradation of grasslands caused by grazing combined with climate changes, including changing precipitation, may lead to cascading effects on ecosystem function and services [5,6]. Overgrazing and precipitation uncertainty have become critical challenges to biodiversity and the ecological equilibrium in grasslands, particularly for those in arid and semi-arid regions [7,8]. Assessing the impacts of grazing on vegetation changes scientifically and adopting suitable management practices for promoting natural vegetation restoration under post-grazing grassland is the basis and effective measure to prevent and control grassland ecological degradation.
Seedlings are an important part of the plant community and the basis of vegetation restoration and regeneration [9,10]. Seedlings in the seedling bank usually affect the distribution and richness of adult plants in the community by participating in the natural regeneration of vegetation, as well as the composition, dynamic changes and species diversity maintenance of the aboveground vegetation [11,12]. The abundance and species composition of seedlings and the time of seedling emergence all restrict the community formation and vegetation dynamics, and they also affect the anti-interference ability and restoration ability of the ecosystem [13]. Therefore, the study of seedling emergence and the abundance of the seedling bank under different environmental circumstances can inform restoration planning and vegetation management [14,15].
Grazing disturbance and rainfall changes have been considered as two important factors impacting seedling emergence and growth in semi-arid grasslands due to seedling emergence being dependent both on seed availability in the soil and on water availability for seedling emergence [16,17]. Grazing strongly influenced seedling emergence through changing seed bank size and seed distribution from livestock feeding and trampling [18,19]. Although the effect of grazing on seedlings has been frequently studied, no consistent conclusion has been found. Some studies found a higher seedling density and higher species richness under moderate grazing intensities [13]. Some researchers reported that higher grazing intensity increased the richness and the density of seedlings [19]. Others suggested that seedling number and diversity reduced with intensive grazing [20]. Furthermore, seedling emergence and growth are sensitive to water availability in arid environments and are closely related to rainfall [17,21]. Higher precipitation and better water conditions might be beneficial for seedling emergence and establishment. On the contrary, seedling emergence was negatively affected by drought and fewer seedlings would emerge and settle in aridity conditions [12,22]. Moreover, the impacts of grazing and precipitation variability on seedlings always interact with each other, which results in the complexity and inconsistency of results [2,23]. For instance, studies found that higher-intensity grazing could buffer the negative impacts of drought on seedling richness and density [24]. Thus, understanding how grazing influences the seedling patterns with rainfall variation is essential and of crucial importance for guiding restoration practices in grasslands against the background of climate change.
Although previous studies have shown the important role of grazing and precipitation variation on shaping seedling communities [6,12,20], most studies have only focused on seedlings during early spring or during a certain stage in the growing season (e.g., in June) about the whole community [13,25]. However, the total emergence and dynamics of seedlings over the whole growing season and the interaction effects of grazing and rainfall variation on seedling patterns are less investigated, and whether the effects of grazing on seedlings will alter with functional types has rarely been addressed. These aspects limit the accuracy and integrity of evaluation of grazing impacts on seedlings. Annuals (including annual grasses and annual forbs) are the primary component of vegetation in semi-arid regions, playing important ecological functions in restoring grassland ecosystems [26]. To understand how grazing affects the seedling patterns of annual plants, we conducted a field investigation experiment and examined the differences in seedling composition, total abundance, and monthly dynamics of seedlings along the grazing gradient and compared the seedling patterns of different functional types in contrasting wet and dry years. The aims of this study were to clarify: (1) how grazing affects seedling composition, abundance, and emergence dynamics; (2) whether the effects of grazing on seedlings were consistent in wet and dry years; and (3) whether the impacts of grazing on seedling emergence differed with functional groups.
2. Materials and Methods
2.1. Study Area
The study was conducted on a sandy grassland near the Naiman Desertification Research Station, Chinese Academy of Sciences (42°54′ N, 120°42′ E, 360 m a.s.l.) in the southwestern Horqin sandy grassland, Inner Mongolia, China (Figure 1). The study area has a typical temperate semi-arid continental and monsoonal climate. The mean annual precipitation is 353.2 mm, nearly 87% of which falls in the growing season from May to September [27]. The mean annual potential evaporation is 1935 mm. The mean annual temperature is approximately 6.9 °C, and the frost-free period averages about 150 days per year [3]. The landform is flat and open; soils are classified as sandy chestnut soils [28]. Plant species are mainly annual grasses and forbs, such as Setaria viridis, Corispermum macrocarpum, Salsola collina, Artemisia scoparia, and Bassia dasyphylla [3,29].
The precipitation and temperature during the experimental period in 2019 and 2020 are shown in Table 1. The precipitation from April to August in 2019 was 341.0 mm, 18.3% higher than the long-term average precipitation in the same period in the study area [30]. The precipitation from April to August in 2020 was 229.2 mm, 20.0% lower than the average precipitation. The average temperatures during the growing seasons of the two years were similar. In terms of precipitation in the growing season, 2019 was a relatively wet year but 2020 was a relatively dry year for plant growth.
2.2. Experimental Design and Data Collection
To explore the seedling pattern of annuals along a grazing gradient, the experiment was established at selected sites in each grazing intensity grassland, managed with different grazing intensities during the last 25 years. The light grazing (LG), moderate grazing (MG), and heavy grazing (HG) grasslands had been managed as a long-term free and public grazing pasture, and livestock was allowed to enter during the autumn and winter of each year according to the local seasonal grazing prohibition policy, with approximately 2 sheep units ha−1, 4 sheep units ha−1, and 6 sheep units ha−1 in light, moderate, and heavy grazing areas, respectively, which were classified according to the standard and method of Zhao et al. [3]. No grazers were present in the study field during experiment. Three groups of fifteen 0.5 m × 0.5 m fixed quadrats in total were randomly set up in each grazing intensity grassland in early April 2019. The seedling emergence of annuals was monitored using the seedling removal method [31] during the growing seasons in 2019 and 2020.
In each year, we investigated the number of species and emerged seedlings of each species every fifteen days during the period of the main growing season from mid-April to mid-August. In each quadrat, the emerging seedlings were identified, recorded, and removed at each investigation. Then, species richness (species number) and the total abundance (number of total seedling emergence) of seedling emerged were obtained by summing the data from each investigation. During the investigation, we marked and kept the seedlings, which were difficult to identify until they grew up to be recognizable. We identified these seedlings species by referring to the online Flora of China (www.cn-flora.ac.cn, (accessed on 15 July 2020)) website. All of the emerged annual seedlings were classified into two plant functional groups as an annual grass species group and an annual forb species group in this study. The functional composition percentage of seedlings was the seedling number of grasses or forbs divided by the total number of seedling emergence, and the percentage of monthly seedling emergence was the number of emergence in each month divided by the total number of seedling emergence in that year. The soil water content (0–20 cm soil depth) in each quadrat was measured simultaneously during the experiment and was measured in triplicate using a soil moisture meter (TDR300). The mean soil water content from each month and each year were calculated under each grazing intensity.
2.3. Data Analyses
Two-way ANOVA was used to test the effects of grazing intensity impact on the abundance in the functional groups of the seedling bank in the two contrasting years. The difference in the functional composition percentage of the seedling bank and monthly seedling emergence among the grazing intensities were tested by one-way ANOVA. Results were considered to be significantly different at the level of p < 0.05 with the Tukey test. Dependent data of emergence were square root-transformed to meet the assumption of normality and homogeneous variances. The difference in average annual soil water content between the two experimental years under different grazing intensities were tested by paired t-test, and a Pearson correlation analysis was used to examine the relationship between seedling emergence and the average monthly soil water content under different grazing intensities. Data analysis and graphing were carried out with the IBM SPSS 22.0 and Origin 22.0 (OriginLab Corporation, Northampton, MA, USA).
3. Results
3.1. Species Richness of the Seedling Bank
In 2019 (the wet year), there were 9, 15, and 7 total annual species recorded in the LG, MG, and HG grasslands, respectively, and 13, 13, and 11 annual species in 2020 (the dry year), with forb species dominating in the seedling bank under all grazing intensities in both years (Figure 2; Table S1). Species richness of grasses, forbs, and all annuals showed the trends of, firstly, an increase, then a decrease along the grazing gradient in 2019. The species richness of grasses showed the same trend in 2020, while the species richness of forbs and all annuals decreased with grazing intensity in 2020 (Figure 2).
3.2. Abundance and Functional Composition of the Seedling Bank
The abundance of the seedling bank of grasses, forbs, and all annuals were significantly different among the different grazing intensities and between the two years in the semi-arid sandy grassland. There were significant interactions between the grazing intensity and year on the abundance of the seedling bank of forbs and all annuals, but no significant interaction with the abundance of the seedling bank of grasses (Table 2).
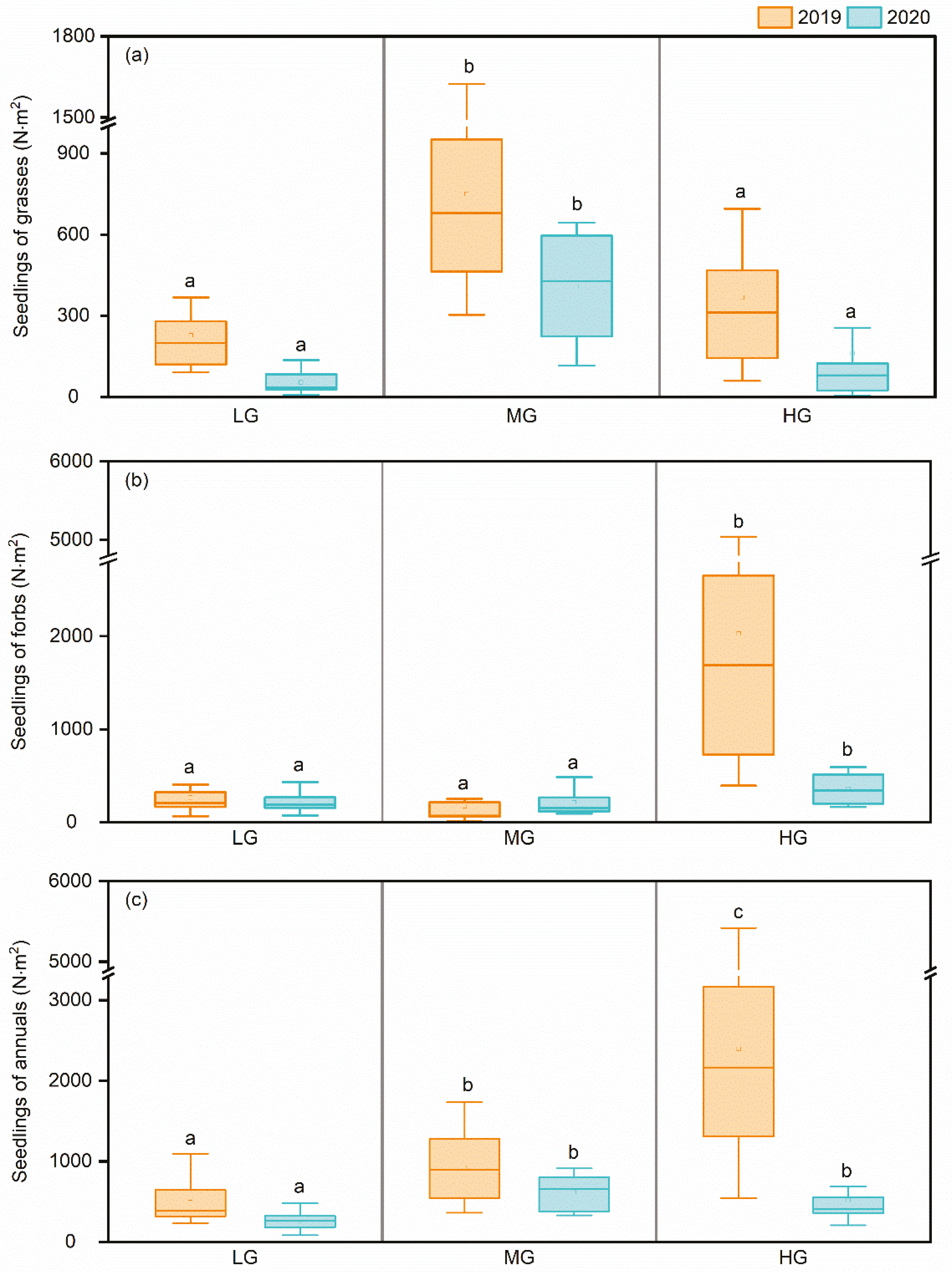
The total seedling emergence of grasses significantly differed (p < 0.05) among grazing intensities with the highest value in MG in both years, and it was always higher in the wet year than in the dry year under all grazing intensities (Figure 3a). The total seedling emergence of forbs was significantly higher (p < 0.05) in HG than in LG and MG in both years, and there were more forbs seedlings in the wet year than in the dry year under HG, but the seedling emergence in terms of the number of forbs changed little from the wet year to the dry year under LG and MG (Figure 3b). The total seedling emergence of all annuals increased with grazing and was significantly higher (p < 0.05) under HG in the wet year but was higher under MG in dry year (Figure 3c).
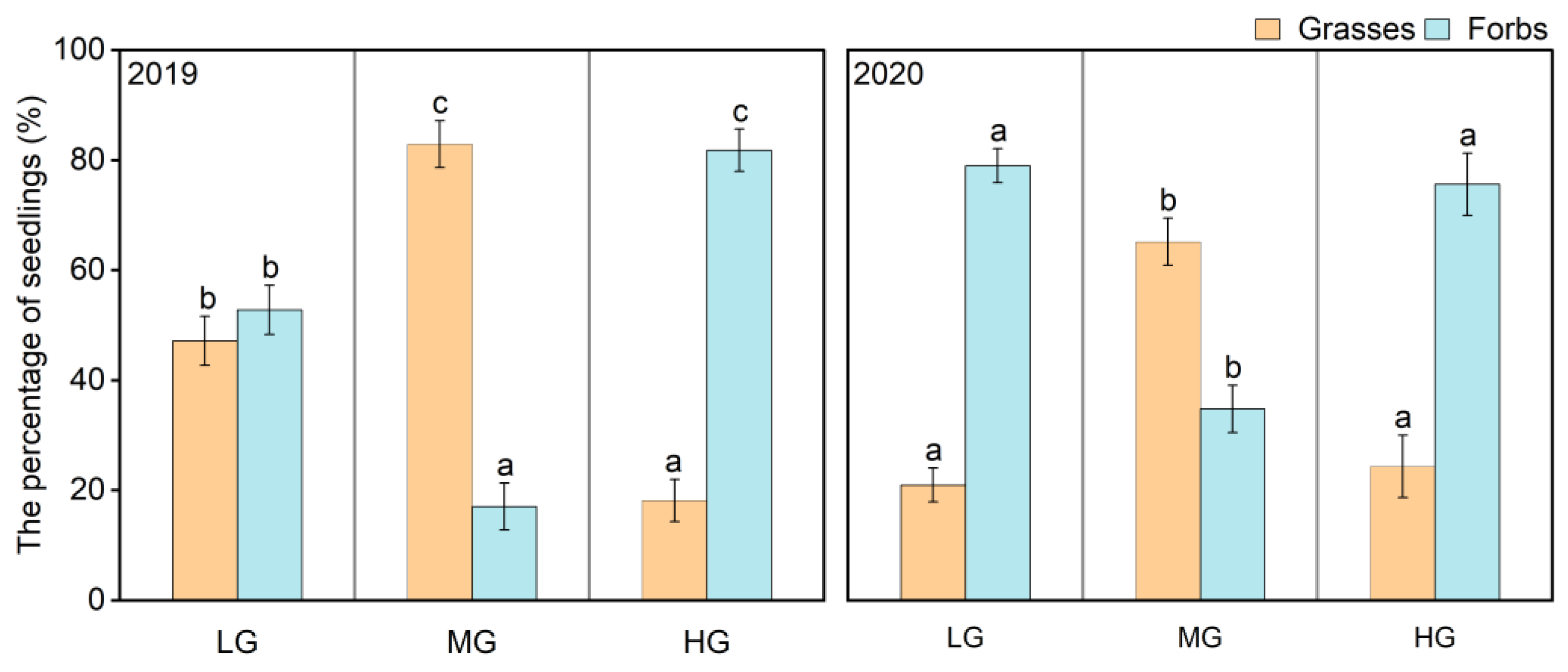
There was a higher proportion of forb seedlings but a lower proportion of grass seedlings under LG and HG; however, there was a higher proportion of grass seedlings but a lower proportion of forb seedlings under MG in both years. In contrast to the wet year, the percentage of grass seedlings decreased but the percentage of forb seedlings increased in the dry year under LG and MG; however, the functional composition percentage of seedlings was similar in the wet year and the dry year under HG (Figure 4).
3.3. Seedling Emergence Dynamics
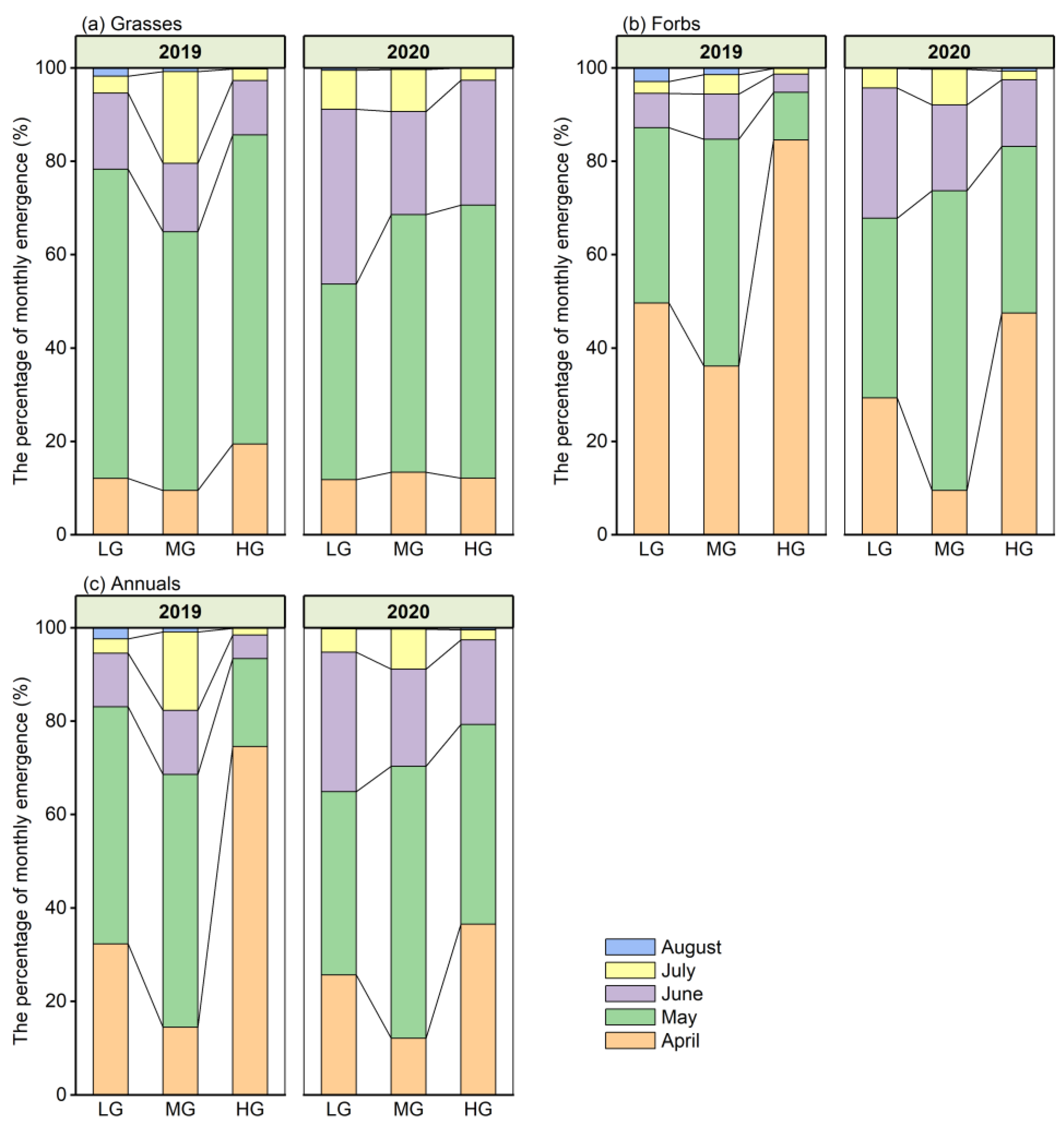
The monthly seedling emergence of grasses was always highest under MG, followed by HG, and was lowest under LG in both years. The seedling emergence dynamics of grasses under LG and HG showed a similar trend, with one peak in May in both years; however, there were two emergence peaks (in May and July) under MG in 2019 but one peak in May in 2020 (Figure 5a,b). The monthly seedling emergence of forbs was always in the order of HG > LG > MG in the wet year, and the emergence dynamics showed a single peak pattern, peaking in April under HG in both years and under LG in the wet year; however, the peak occurred in May under MG in both years and under LG in the dry year (Figure 5c,d). For all annuals, the difference in monthly seedling emergence among grazing intensities was similar to that of grasses, in the order of MG > HG > LG, except in April. The emergence dynamics showed a single peak pattern in most cases, with the exception of two peaks under MG in the wet year (Figure 5e,f).
Although there was higher proportion of grass seedlings emerging in May in both years, the percentage of seedling emergence in May decreased in 2020 under LG and HG and was similar under MG between the two years (Figure 6a). However, the emergence percentage in June increased in 2020 under all grazing intensities, compared with those in 2019 (Figure 6a). A higher proportion of forb seedlings emerged in April and May; however, in contrast to 2019, the emergence percentage in April decreased in 2020, while the emergence percentage in May and June increased under all grazing intensities (Figure 6b). The emergence percentage of all annuals in the first two months (April–May) under LG and HG decreased in 2020, compared to those in 2019 (Figure 6c). However, the percentage of emergence for all annuals in June–July obviously increased in 2020, compared with those in 2019 under LG and HG (Figure 6c).
3.4. Correlation between Seedling Emergence and Soil Water Content
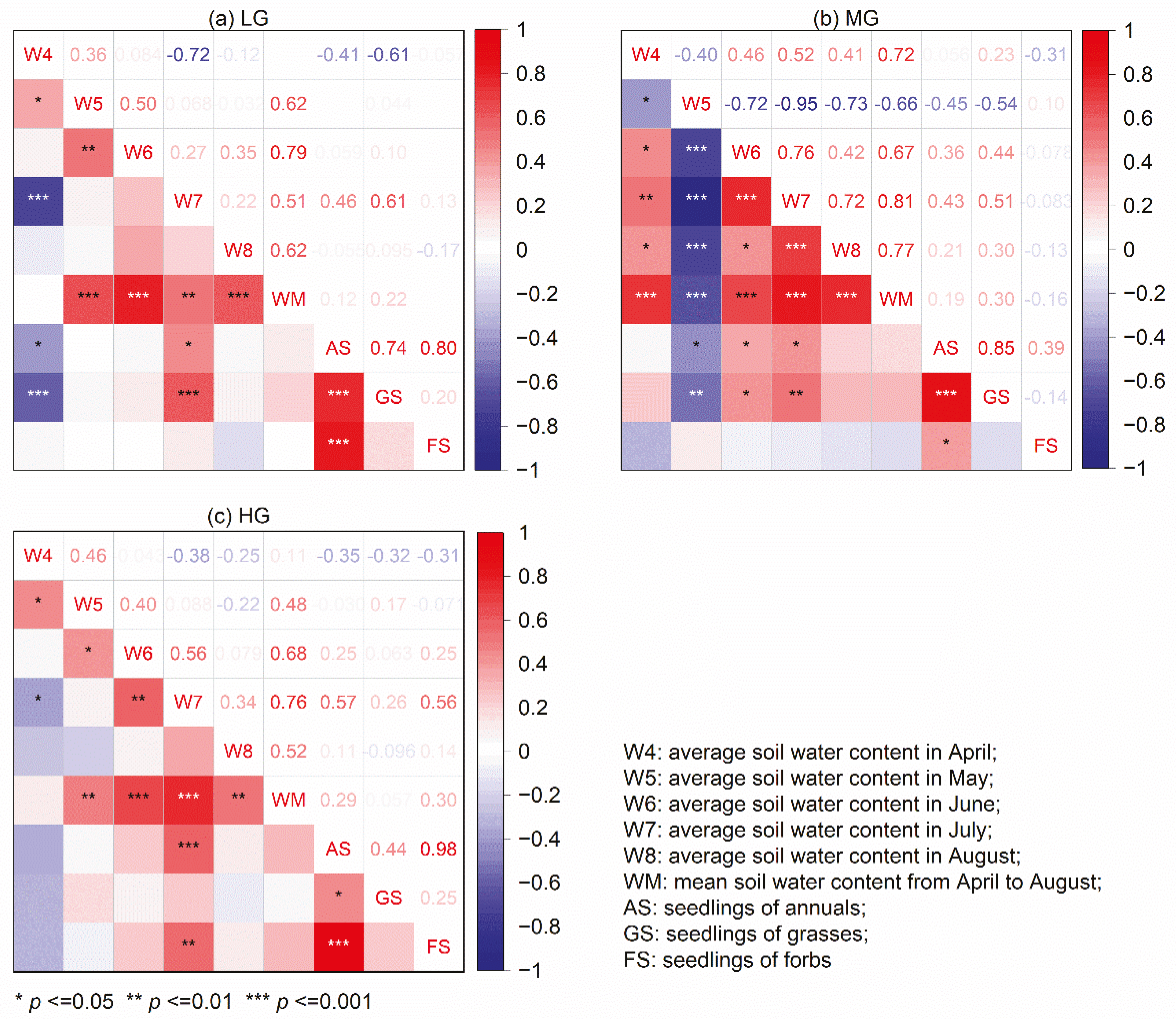
The results showed that the soil water contents in 2019 were obviously higher than those in 2020 under all grazing intensities (Figure 7). There were significant positive responses in the total seedling emergence of annuals and grasses to soil water content in July but negative responses to soil water content in April under LG (Figure 8a). However, under MG, the total seedling emergence of annuals and grasses positively correlated with the soil water content in June and July but negatively correlated with the soil water content in May (Figure 8b). Significant positive correlations were only found between the total seedling emergence of annuals and forbs and the soil water content in July under HG (Figure 8c). The total seedling emergence of annuals significantly correlated with the seedling emergence of both grasses and forbs under LG, while there was a higher correlation with seedling emergence of grasses under MG and a higher correlation with seedling emergence of forbs under HG.
4. Discussion
4.1. Effects of Grazing on Species Richness of Seedlings in Wet and Dry Years
Our data showed that species richness of the total emerged seedlings along the grazing gradient differed between the wet and dry years. It suggested that rainfall variation in the growing season influenced the impacts of grazing on seedling species richness. In the wet year, seedlings of more species of both grasses and forbs presented with MG, which was in agreement with the results of other studies, showing that higher species richness of seedlings was found under moderate grazing than the other grazing intensities [13]. This suggests that moderate grazing enhanced the seedling emergence of more species in wet conditions. Previous studies have suggested that moderate grazing could promote the enhancement of the plant richness and diversity in semiarid grasslands [32]; there is a higher similarity of species composition between the seedlings emerged from the seed bank and the aboveground vegetation [33,34,35,36], and grazing influenced the richness of seedlings by affected the richness of aboveground species [12]; thus, more species being present under MG conditions may be explained by the higher species diversity of vegetation in the moderate grazing habitat [3,36], and adequate rainfall in the wet year might enable seeds from more species to germinate and emerge. However, low species richness in the aboveground vegetation under LG and HG limited the species diversity of emerged seedlings, especially in HG [13,36,37,38], even though the water conditions were better. Furthermore, the species richness of seedlings (especially for forbs) decreased with grazing in the dry year. It is possible that intensive livestock trampling with heavily grazing might lead to more seeds on the soil surface being buried into deeper soil [39], and it is difficult for those deep buried seeds to germinate and emerge under the scant rainfall conditions in the dry year. Additionally, the seeds of forbs with flat or non-spherical diaspores are more likely to be buried and they have high requirements for water conditions for seedling emergence [34,40,41].
4.2. Effects of Grazing on Seedling Abundance of the Seedling Bank in Wet and Dry Years
The results from the present study indicated that grazing and rainfall variation had a remarkable individual and interactive effect on the abundance of the seedling bank. More seedlings of grasses emerged in the wet year and under MG, which suggested that moderate grazing and adequate rainfall were beneficial for the seedling emergence of grasses, while the seedling emergence of grasses was greatly limited under light and heavy grazing, particularly in a drought year. This may be due to the possibility of moderate grazing facilitating seed storage in the soil seed bank [42] and more grasses species emerging under MG. The results were in line with the conclusions from other studies, in which a higher seedling emergence was also found under a moderate grazing intensity [11]. Moreover, although many environmental factors, such as temperature, influenced seedling emergence, studies have shown that rainfall and water availability were the most important environmental factors that restrict seed germination and seedling emergence in arid and semi-arid regions [41,43,44]. Furthermore, water availability in the soil is mostly regulated by rainfall variation in this region [26,45], which resulted in a higher seedling emergence in the wet year than in the dry year. In addition, the temperatures of the two experimental years was similar (Table 1), but the soil water contents were obviously higher in the wet year than in the dry year, then the seedling emergence was higher in the wet year than in the dry year, indicating the effect of rainfall variation.
In contrast, a higher seedling abundance of forbs was found under HG in the wet year in this study, which suggested that heavy grazing and higher soil water availability could stimulate more seedlings of forbs to emerge. This was in line with previous studies, which have demonstrated that decreasing the grazing intensity reduced the species number and number of emerged seedlings; however, intensive grazing could be effective for enhancing seedling growth [46,47]. The results could be attributed to the seedling emergence of plants being affected by both the depth of seed burial and the amount of precipitation [20], as livestock trampling with intensive grazing could strongly promote seedling emergence by pressing surface-lying seeds into the uppermost soil layer, and create suitable establishment gaps for emergence [19,48]. Therefore, a certain depth of seed burial might be needed and be helpful for the seedling emergence of forbs in the study area. This was also evident in our results regarding a lower seedling abundance of forbs under LG and MG than under HG, regardless of whether it was a wet or dry year. Furthermore, our results showed that the seedling abundance of all annuals in 2019, with a relatively higher rainfall, was highest under HG; however, it was higher under MG in 2020, with a relative lower rainfall. This is likely to be related to the higher proportion of forb seedlings under HG, but the higher proportion of grass seedlings under MG. Overall, considering the whole annual community in the study region, the results suggested that increasing the grazing intensity, especially with heavy grazing, could strongly promote an increase in the seedling abundance with higher rainfall, while moderate grazing has the greatest promoting effect on seedling emergence during drought.
4.3. Grazing Effects on Emergence Dynamics in Wet and Dry Years
Studies have shown that the pattern of seed germination and seedling emergence over time is similar to a single peak, which is the manifestation of the biological characteristics of seeds under appropriate conditions and is also the adaptive responses of plants to environmental conditions [49,50,51]. In our study, seedling emergence also showed a single peak pattern in most cases while the time of the peak differed with functional types, grazing intensities and between the wet and dry year, and the peak of seedling emergence was advanced in April under heavy grazing, compared with that in May under other conditions. This suggested that the emergence dynamics of plants in semi-arid grasslands are not only related to plant characteristics but also influenced by grazing disturbance and rainfall variation. Previous studies have reported that annual plants generally exhibit two different germination and emergence strategies, including an opportunistic strategy and a cautious strategy, which can effectively improve the competitive advantage of plants [43,52]. Here, most species under LG and MG might exhibit a cautious strategy; they generally emerge in batches, avoiding competition for resources and deterring the possible risks of mass loss through heterogeneity in the timing of seed germination and seedling emergence [43,53]. However, most species (in particular for forbs) under HG were likely to exhibit an opportunistic strategy: to adapt to uncertainty environments in the following summer through rapid emergence and early settlement. In addition, a large number of seedlings emerged in early spring, providing enough individuals for environmental screening so that a certain number of seedlings could survive and eventually establish, which provided a material basis for population reproduction and regeneration [54,55].
A notable exception was that the emergence dynamics of grasses under MG showed two peak patterns with the second peak occurring in July 2019. A possible explanation is that sufficient rainfall in July in that year stimulated the germination and emergence of dormant seeds in the deeper soil under MG, due to the higher species richness under MG (Figure 2). The correlation between seedling abundance and the soil water content also confirmed that seedling emergence was strongly influenced by soil water content in July. Studies have shown that delaying germination and emergence is another effective risk-dispersion strategy for plants in unpredictable environments [56,57]. In the present study, the emergence percentage of grasses and forbs in April and May decreased, but the emergence percentage in June and July increased in 2020 with low rainfall, compared with that in 2019 with more rainfall, especially under HG. This could be partly due to the lower rainfall during spring, especially in April 2020, not meeting the water conditions for seed germination, which limited the seedling emergence of most plants [29,31], while more rainfall in June–July during the rainy season stimulated the germination and emergence of more plant seeds. Moreover, previous studies have suggested that the seeds of some plants would remain dormant in severe environments with lower water conditions but overcome this dormancy when the environment improves with soil water availability increasing [31,36,58]. Hence, some annual plants in the studied region may adopt the strategy of delaying emergence to adapt to drought and severe environments in years with low rainfall. However, due to the complexity of the interactive effects of grazing and rainfall on the plants, continuous observation and widely ranging experiments are still required to explore the mechanisms of the grazing impacts on vegetation changes under the background of climate change.
5. Conclusions
Grazing strongly affected seedling richness, seedling abundance, and emergence dynamics, while the effects altered with functional types and rainfall variation. Moderate grazing could enhance forb richness in a wet year and enhance grass richness in both wet and dry years, while forb richness decreased with the increase in grazing intensity in a dry year. There were more seedlings emerging in a wet year than in a dry year, and the emergence peak varied with rainfall variation and grazing intensity. Moderate grazing enhanced the seedling abundance of grasses, while heavy grazing promoted the seedling abundance of forbs and advanced the peak time of seedling emergence. This study showed that increasing grazing was beneficial for seedling emergence in the year with more rainfall during the growing season, but moderate grazing was more helpful for enhancing seedling emergence in the year with low rainfall during the growing season. It is necessary to adopt more judicious grazing management measures based on rainfall variation and the functional composition of the plant community, thus promoting the vegetation restoration of semi-arid grasslands through affecting the seedling emergence patterns.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12092131/s1, Table S1: species composition under the different grazing intensities during the two experimental years.
Author Contributions
Conceptualization, Y.H.; methodology, Y.H. and X.L.; validation, Y.H.; formal analysis, Y.H.; investigation, X.L.; resources, X.L.; data curation, Y.H.; writing—original draft preparation, Y.H.; writing—review and editing, Y.H. and X.L.; visualization, Y.H.; supervision, X.L.; project administration, Y.H. and X.L.; funding acquisition, Y.H. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science and Technology Planning Project of Inner Mongolia Autonomous Region (2022YFHH0063); the National Natural Science Foundation of China (41801076).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are thankful to Mingming Wang, Shanshan Sun, and Li Chen for their collaboration in the experimental work, and to the Naiman Desertification Research Station, Chinese Academy of Sciences, for the support in field work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheng, H.; Jin, B.C.; Luo, K.; Pei, J.Y.; Zhang, X.L.; Zhang, Y.H.; Tang, J.Q.; Yang, Q.; Sun, G.J. Vegetation response to goats grazing intensity in semiarid hilly grassland of the Loess Plateau. Sustainability 2021, 13, 3569. [Google Scholar] [CrossRef]
- Pfeiffer, M.; Langan, L.; Linstadter, A.; Martens, C.; Gaillard, C.; Ruppert, J.C.; Higgins, S.I.; Mudongo, E.I.; Scheiter, S. Grazing and aridity reduce perennial grass abundance in semi-arid rangelands-Insights from a trait-based dynamic vegetation model. Ecol. Model. 2019, 395, 11–22. [Google Scholar] [CrossRef]
- Zhao, H.L.; Zhang, T.H.; Zhao, X.Y.; Zhou, R.L. Effects of sheep grazing and precipitation patterns on sandy grassland vegetation in Inner Mongolia, China. Environ. Sci. Develop. 2011, 4, 68–75. [Google Scholar]
- Zainelabdeen, Y.M.; Yan, R.; Xin, X.; Yan, Y.; Ahmed, A.I.; Hou, L.; Zhang, Y. The Impact of Grazing on the Grass Composition in Temperate Grassland. Agronomy 2020, 10, 1230. [Google Scholar] [CrossRef]
- Bat-Oyun, T.; Shinoda, M.; Cheng, Y.X.; Purevdorj, Y. Effects of grazing and precipitation variability on vegetation dynamics in a Mongolian dry steppe. J. Plant Ecol. 2016, 9, 508–519. [Google Scholar] [CrossRef]
- Wan, H.; Bai, Y.; Hooper, D.U.; Schönbach, P.; Gierus, M.; Schiborra, A.; Taube, F. Selective grazing and seasonal precipitation play key roles in shaping plant community structure of semi-arid grasslands. Landsc. Ecol. 2015, 30, 1767–1782. [Google Scholar] [CrossRef]
- Huang, X.T.; Luo, G.P.; Chen, C.B.; Peng, J.; Zhang, C.J.; Zhou, H.K.; Yao, B.Q.; Ma, Z.; Xi, X.Y. How precipitation and grazing influence the ecological functions of drought-prone grasslands on the northern slopes of the Tianshan Mountains, China? J. Arid Land 2021, 13, 88–97. [Google Scholar] [CrossRef]
- Ren, H.; Schönbach, P.; Wan, H.; Gierus, M.; Taubeet, F. Effects of grazing intensity and environmental factors on species composition and diversity in typical steppe of Inner Mongolia, China. PLoS ONE 2012, 7, e52180. [Google Scholar] [CrossRef] [PubMed]
- Esaete, J.; Muwanika, V.B.; Musiba, R.; Mawa, C.; Tabuti, J.R.S. Understory seedling bank in forest areas with a differing period of recovery in Mabira Central Forest Reserve, South Central Uganda. Environ. Manag. 2022, 1–11. [Google Scholar] [CrossRef]
- Gonzalez, M.; Deconchat, M.; Balent, G.; Cabanettes, A. Diversity of woody plant seedling banks under closed canopy in fragmented coppice forests. Ann. Forest Sci. 2008, 65, 511. [Google Scholar] [CrossRef]
- Albrecht, M.A.; McCarthy, B.C. Seedling establishment shapes the distribution of shade-adapted forest herbs across a topographical moisture gradient. J. Ecol. 2009, 97, 1037–1049. [Google Scholar] [CrossRef]
- Hu, A.; Zhang, J.; Chen, X.J.; Millner, J.P.; Chang, S.H.; Bowatte, S.; Hou, F.J. The composition, richness, and evenness of seedlings from the soil seed bank of a semi-arid steppe in northern China are affected by long-term stocking rates of sheep and rainfall variation. Range. J. 2019, 41, 23–32. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chu, L.; Daryanto, S.; Wang, L.X.; Lin, J.X.; Ala, M. The impact of grazing on seedling patterns in degraded sparse-elm grassland. Land Degrad. Dev. 2018, 29, 2330–2337. [Google Scholar] [CrossRef]
- Janisova, M.; Skodova, I.; Hegedusova, K.; Kochjarova, J. Seed bank and seedling recruitment of endangered Tephroseris longifolia subsp moravica (Asteraceae). Folia Geobot. 2016, 51, 343–360. [Google Scholar] [CrossRef]
- Grewel, B.J.; Gillard, M.B.; Futrell, C.J.; Castillo, J.M. Seedling Emergence from Seed Banks in Ludwigia hexapetala-Invaded Wetlands: Implications for Restoration. Plants 2019, 8, 451. [Google Scholar] [CrossRef] [PubMed]
- Oesterheld, M.; Sala, O.E. Effects of Grazing on Seedling Establishment-the Role of Seed and Safe-Site Availability. J. Veg. Sci. 1990, 1, 353–358. [Google Scholar] [CrossRef]
- Fay, P.A.; Schultz, M.J. Germination, survival, and growth of grass and forb seedlings: Effects of soil moisture variability. Acta Oecol. 2009, 35, 679–684. [Google Scholar] [CrossRef]
- Osem, Y.; Konsens, I.; Perevolotsky, A.; Kigel, J. Soil seed bank and seedling emergence of Sarcopoterium spinosum as affected by grazing in a patchy semiarid shrubland. Israel J. Plant Sci. 2007, 55, 35–43. [Google Scholar] [CrossRef]
- Eichberg, C.; Donath, T.W. Sheep trampling on surface-lying seeds improves seedling recruitment in open sand ecosystems. Restor. Ecol. 2018, 26, 211–219. [Google Scholar] [CrossRef]
- Gallacher, D.J.; Hill, J.P. Effects of camel grazing on density and species diversity of seedling emergence in the Dubai (UAE) inland desert. J. Arid Environ. 2008, 72, 853–860. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Yang, X.J.; Baskin, C.C.; Baskin, J.M.; Dong, M.; Huang, Z.Y. Effects of amount and frequency of precipitation and sand burial on seed germination, seedling emergence and survival of the dune grass Leymus secalinus in semiarid China. Plant Soil 2014, 374, 399–409. [Google Scholar] [CrossRef]
- Classen, A.T.; Norby, R.J.; Campany, C.E.; Sides, K.E.; Weltzin, J.F. Climate change alters seedling emergence and establishment in an old-field ecosystem. PLoS ONE 2010, 5, e13476. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Davison, E.A. Emergence and survival of herbaceous seedlings in temperate grassy woodlands: Recruitment limitations and regeneration niche. Austral Ecol. 2004, 29, 320–331. [Google Scholar] [CrossRef]
- Wang, D.B.; Wang, X.Y.; Wu, Y.; Lin, H.L. Grazing buffers the effect of climate change on the species diversity of seedlings in an alpine meadow on the Tibetan Plateau. Ecol. Evol. 2019, 9, 1119–1126. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Li, F.R. Characteristics of the soil seed bank and the seedling bank in fenced sandy meadow. Acta Bot. Boreali-Occident. Sin. 2003, 23, 1725–1730. [Google Scholar]
- Yue, X.F.; Zhang, T.H.; Zhao, X.Y.; Liu, X.P.; Ma, Y.H. Effects of rainfall patterns on annual plants in Horqin Sandy Land, Inner Mongolia of China. J. Arid Land. 2016, 8, 389–398. [Google Scholar] [CrossRef]
- Liu, X.P.; He, Y.H.; Sun, S.S.; Zhang, T.H.; Luo, Y.Q.; Zhang, L.M.; Wang, M.M.; Cheng, L.; Hu, H.J.; Xu, Y.Z. Restoration of sand-stabilizing vegetation reduces deep percolation of precipitation in semi-arid sandy lands, northern China. Catena 2022, 208, 105728. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhao, H.L.; Zhao, X.Y.; Zhang, T.H.; Li, Y.L.; Cui, J.Y. Effects of grazing and livestock exclusion on soil physical and chemical properties in desertified sandy grassland, Inner Mongolia, northern China. Environ. Earth Sci. 2011, 63, 771–783. [Google Scholar] [CrossRef]
- Ma, Y.H.; Zhang, T.H.; Liu, X.P. Effect of intensity of small rainfall simulation in spring on annuals in Horqin Sandy Land, China. Environ. Earth Sci. 2015, 74, 727–735. [Google Scholar] [CrossRef]
- Yao, S.; Zhang, T.; Zhao, C. Multiple-time-scale Analysis on Precipitation in Naiman Banner of Horqin Sandy Land in 1970–2010. J. Desert Res. 2014, 34, 42–549. [Google Scholar] [CrossRef]
- He, Y.H.; Zhao, H.L.; Liu, X.P.; Zhang, T.H. Seed germination and population dynamic of two annual plants in sand land restoration process. J. Desert Res. 2010, 30, 1331–1335. [Google Scholar]
- Zhao, H.L.; Okuro, T.; Zhou, R.L.; Li, Y.L.; Zuo, X.A.; Huang, G. Effects of grazing and climate change on species diversity in sandy grassland, Inner Mongolia, China. Sci. Cold Arid Reg. 2009, 1, 30–38. [Google Scholar]
- Osem, Y.; Perevolotsky, A.; Kigel, J. Similarity between seed bank and vegetation in a semi-arid annual plant community: The role of productivity and grazing. J. Veg. Sci. 2006, 17, 29–36. [Google Scholar] [CrossRef]
- Li, X.H.; Li, X.L.; Jiang, D.; Liu, Z.; Yu, Q.H. Annual plants in arid and semi-arid desert regions. Front. Biol. 2008, 3, 259–264. [Google Scholar] [CrossRef]
- Harrison, S.; Inouye, B.D.; Safford, H.D. Ecological heterogeneity in the effects of grazing and fire on grassland diversity. Conserv. Biol. 2003, 17, 837–845. [Google Scholar] [CrossRef]
- Hopfensperger, K.N. A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 2007, 116, 1438–1448. [Google Scholar] [CrossRef]
- Tessema, Z.K.; de Boer, W.F.; Baars, R.M.T.; Prins, H.H.T. Influence of grazing on soil seed banks determines the restoration potential of aboveground vegetation in a semi-arid Savanna of Ethiopia. Biotropica 2012, 44, 211–219. [Google Scholar] [CrossRef]
- Zhao, W.Z.; Liu, Z.M.; Chang, X.L. Influence of grazing intensity on seed bank of a sandy grassland in Horqin Steppe of China. Ann. Arid Zone. 2001, 40, 397–404. [Google Scholar]
- Jahantab, E.; Yazdanshenas, H.; Saray, A.A.; Matinkhah, S.H.; Khazaei, M. Seed burial depth, seedling emergence, and height as affected by animal trampling in marl soils. Plant Ecol. 2022, 223, 493–506. [Google Scholar] [CrossRef]
- Liu, Z.M.; Hua, L.X.; Ping, L.R.; Luo, Y.M.; Wang, H.M.; Jiang, D.M. A comparative study on diaspore shape of 70 species found in the sandy land of Horqin. Acta Prataculturae Sci. 2003, 12, 55–61. [Google Scholar]
- Ramawat, K.G. Desert Plants: Biology and Biotechnology; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Willms, W.D.; Quinton, D.A. Grazing effects on germinable seeds on the fescue prairie. J. Range Manag. 1995, 48, 423–430. [Google Scholar] [CrossRef]
- Gutterman, Y. Seed Germination in Desert Plants; Springer: Berlin/Heidelberg, Germany, 1993; p. 253. [Google Scholar]
- Tobe, K.; Zhang, L.; Omasa, K. Seed germination and seedling emergence of three annuals growing on desert sand dunes in China. Ann. Bot. 2005, 95, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.P.; He, Y.H.; Zhao, X.Y.; Zhang, T.H.; Li, Y.L.; Yun, J.Y. The response of soil water and deep percolation under Caragana microphylla to rainfall in the Horqin Sand Land, northern China. Catena 2016, 139, 82–91. [Google Scholar] [CrossRef]
- Tadey, M.; Souto, C.P. Unexpectedly, intense livestock grazing in arid rangelands strengthens the seedling vigor of consumed plants. Agron. Sustain. Dev. 2016, 36, 63. [Google Scholar] [CrossRef]
- Schuster, M.Z.; Pelissari, A.; de Vioraes, A.; Harrison, S.K.; Sulc, R.M.; Lustosa, S.B.C.; Anghinoni, I.; Carvalho, P.C.F. Grazing intensities affect weed seedling emergence and the seed bank in an integrated crop-livestock system. Agric. Ecosyst. Environ. 2016, 232, 232–239. [Google Scholar] [CrossRef]
- Kiss, R.; Deak, B.; Tothmeresz, B.; Miglecz, T. Establishment gaps in species-poor grasslands: Artificial biodiversity hotspots to support the colonization of target species. Restor. Ecol. 2021, 29, e13135. [Google Scholar] [CrossRef] [Green Version]
- Garwood, N.C. Seed germination in a seasonal tropical forest in Panama: A community study. Ecol. Monogr. 1983, 53, 159–181. [Google Scholar] [CrossRef]
- Miller, T.E. Effects of emergence time on survival and growth in an early old-field plant community. Oecologia 1987, 72, 272–278. [Google Scholar] [CrossRef]
- Noe, G.B.; Zedler, J.B. Spatio-temporal variation of salt marsh seedling establishment in relation to the abiotic and biotic environment. J. Veg. Sci. 2001, 12, 61–74. [Google Scholar] [CrossRef]
- Li, X.H. Biology of Annual Plants in Arid and Semi-Arid Desert Regions of China; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Nunes, F.P.; Dayrell, R.L.; Silveira, F.A.; Negreiros, D.; Santana, D.G.d.; Carvalho, F.J.; Garcia, Q.S.; Fernandes, G.W. Seed Germination Ecology in Rupestrian Grasslands. In Ecology and Conservation of Mountaintop Grasslands in Brazil; Fernandes, G., Ed.; Springer: Cham, Switzerland, 2016; pp. 207–225. [Google Scholar]
- Houle, G. Environmental filters and seedling recruitment on a coastal dune in subarctic Quebec (Canada). Can. J. Bot. 1996, 74, 1507–1513. [Google Scholar] [CrossRef]
- Castillo-Díaz, D.; Chen, H.Y.; Harrison, R.D.; Wen, B.; Goodale, U.M. Seedling emergence and environmental filters determine Ficus recruitment in a subtropical landscape. Forest Ecol. Manag. 2021, 497, 119536. [Google Scholar] [CrossRef]
- Venable, D.L.; Lawlor, L. Delayed germination and dispersal in desert annuals: Escape in space and time. Oecologia 1980, 46, 272–282. [Google Scholar] [CrossRef] [PubMed]
- López, R.P.; Hayes, M.; Maidana-Tuco, Y.; Naoki, K. Summer annual plants exhibit germination responses to an aridity gradient consistent with both delayed germination and phenotypic plasticity. Plant Ecol. 2020, 221, 1233–1242. [Google Scholar] [CrossRef]
- Li, X.; Li, X.L.; Jiang, D.M.; Liu, Z.M. Germination strategies and patterns of annual species in the temperate semiarid region of China. Arid Land Res. Manag. 2006, 20, 195–207. [Google Scholar] [CrossRef]
Figure 1.
Location of the study area.

Figure 2.
Species richness of the seedling bank in different grazing intensities during the two years.
Figure 2.
Species richness of the seedling bank in different grazing intensities during the two years.

Figure 3.
Total seedling emergence of annual grasses (a), annual forbs (b), and all annuals (c) under the different grazing intensities during contrasting wet and dry years. Different letters indicate significant differences (p < 0.05) among the different grazing intensities in the same year.
Figure 3.
Total seedling emergence of annual grasses (a), annual forbs (b), and all annuals (c) under the different grazing intensities during contrasting wet and dry years. Different letters indicate significant differences (p < 0.05) among the different grazing intensities in the same year.

Figure 4.
Functional composition percentage of the seedling bank under the different grazing intensities during the two years. Mean ± SE. Different letters indicate significant differences (p < 0.05) among the different grazing intensities in the same functional groups.
Figure 4.
Functional composition percentage of the seedling bank under the different grazing intensities during the two years. Mean ± SE. Different letters indicate significant differences (p < 0.05) among the different grazing intensities in the same functional groups.

Figure 5.
The monthly seedling emergence of different functional groups under the different grazing intensities during the two contrasting years. The asterisk indicates significant difference among grazing intensities each month. (a,b) the seedling emergence of grasses; (c,d) the seedling emergence of forbs; (e,f) the seedling emergence of all annuals. Mean ± SE. * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 5.
The monthly seedling emergence of different functional groups under the different grazing intensities during the two contrasting years. The asterisk indicates significant difference among grazing intensities each month. (a,b) the seedling emergence of grasses; (c,d) the seedling emergence of forbs; (e,f) the seedling emergence of all annuals. Mean ± SE. * p < 0.05, ** p < 0.01, *** p < 0.001.

Figure 6.
The percentage of monthly seedling emergence under the different grazing intensities during the two years. (a) annual grasses; (b) annual forbs; (c) all annuals.
Figure 6.
The percentage of monthly seedling emergence under the different grazing intensities during the two years. (a) annual grasses; (b) annual forbs; (c) all annuals.

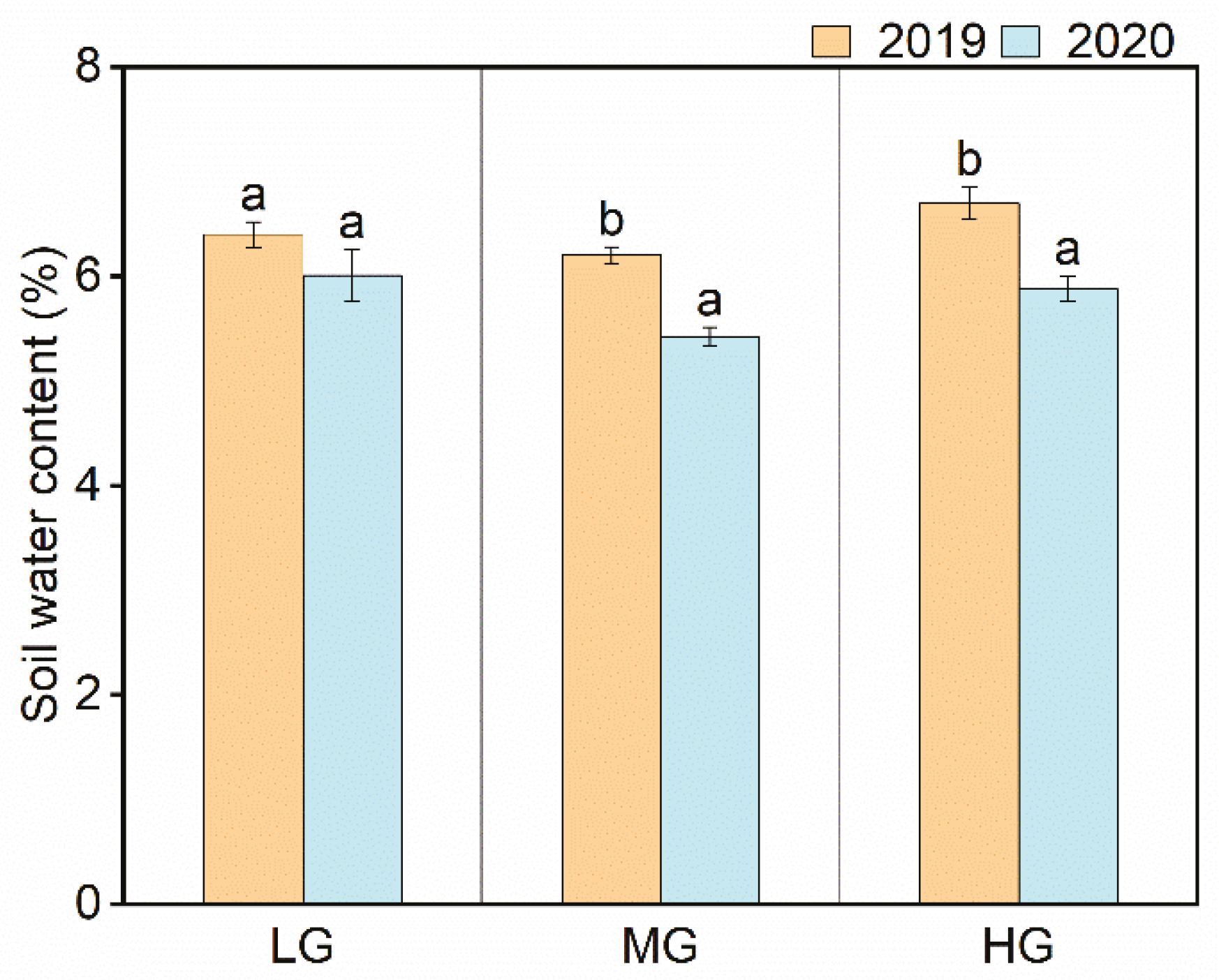
Figure 7.
The soil water contents under the different grazing intensities in the two contrasting years. Mean ± SE. Different letters indicate significant differences (p < 0.05) between the two years under the same grazing intensity.
Figure 7.
The soil water contents under the different grazing intensities in the two contrasting years. Mean ± SE. Different letters indicate significant differences (p < 0.05) between the two years under the same grazing intensity.

Figure 8.
Pearson correlations between the seedling emergence and the monthly soil water content under the different grazing intensities. (a) Correlations in LG; (b) Correlations in MG; (c) Correlations in HG.
Figure 8.
Pearson correlations between the seedling emergence and the monthly soil water content under the different grazing intensities. (a) Correlations in LG; (b) Correlations in MG; (c) Correlations in HG.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Precipitation and temperature during the growing season from April to August in two experimental years.
Table 1.
Precipitation and temperature during the growing season from April to August in two experimental years.
| Time | Precipitation (mm) | Temperature (°C) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| April | May | June | July | August | Total | April | May | June | July | August | Average | |
| 2019 | 6.2 | 72.0 | 59.8 | 61.6 | 141.4 | 341.0 | 9.8 | 18.3 | 21.4 | 24.5 | 21.2 | 19.0 |
| 2020 | 3.0 | 92.6 | 43.6 | 30.6 | 59.4 | 229.2 | 8.7 | 16.5 | 22.3 | 24.6 | 22.4 | 18.9 |
Table 2.
F values of two-way ANOVAs of the seedling abundance with grazing and year.
| Factors | Seedlings of Grasses | Seedlings of Forbs | Seedlings of All Annuals | |||
|---|---|---|---|---|---|---|
| F | Sig. | F | Sig. | F | Sig. | |
| Grazing | 24.283 | 0.000 *** | 17.353 | 0.000 *** | 21.666 | 0.000 *** |
| Year | 20.744 | 0.000 *** | 28.160 | 0.000 *** | 16.129 | 0.000 *** |
| Grazing × Year | 0.950 | 0.391 | 12.611 | 0.000 *** | 15.680 | 0.000 *** |
Note: *** p < 0.001.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
He, Y.; Liu, X. Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland. Agronomy 2022, 12, 2131. https://doi.org/10.3390/agronomy12092131
AMA Style
He Y, Liu X. Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland. Agronomy. 2022; 12(9):2131. https://doi.org/10.3390/agronomy12092131
Chicago/Turabian StyleHe, Yuhui, and Xinping Liu. 2022. "Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland" Agronomy 12, no. 9: 2131. https://doi.org/10.3390/agronomy12092131
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.