
Transcriptomic Characterizations of the Molecular Mechanisms Regulating Lipid Production and Composition in Mortierella alpina in Response to Cold Stress

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
2.2. Lipid Analysis
2.3. RNA Extraction, Library Construction, and Illumina Sequencing
2.4. RNA-Seq Analysis, De Novo Assembly, and Functional Annotation
2.5. Analysis of Differentially Expressed Unigenes
2.6. Quantitative Real-Time PCR Analysis
3. Results
3.1. Effects of Temperature on Fatty Acid Compositions in Mortierella alpina
3.2. RNA-Seq and Transcriptome Analyses of Lipid Synthesis in Mortierella alpina under Cold Stress
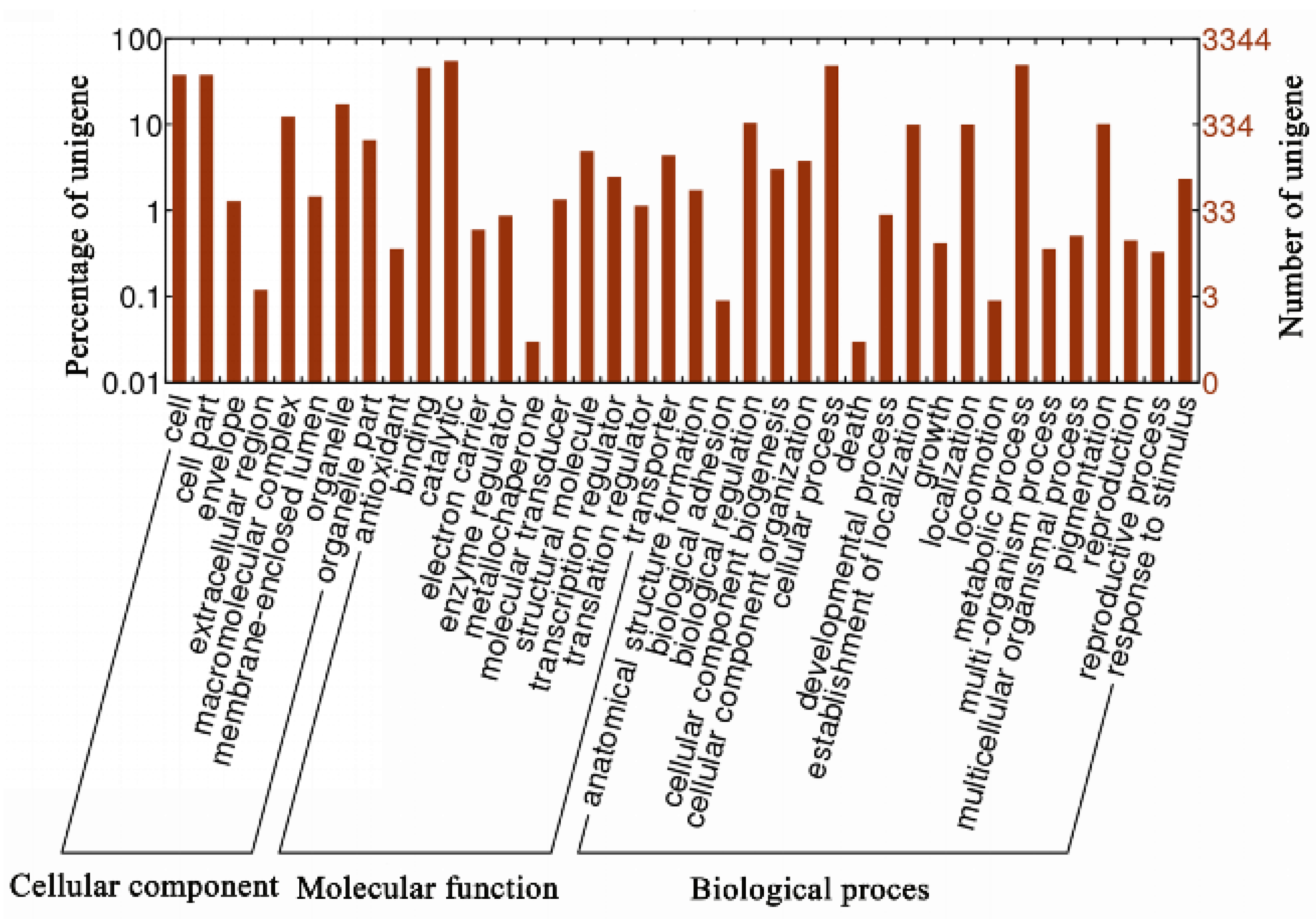
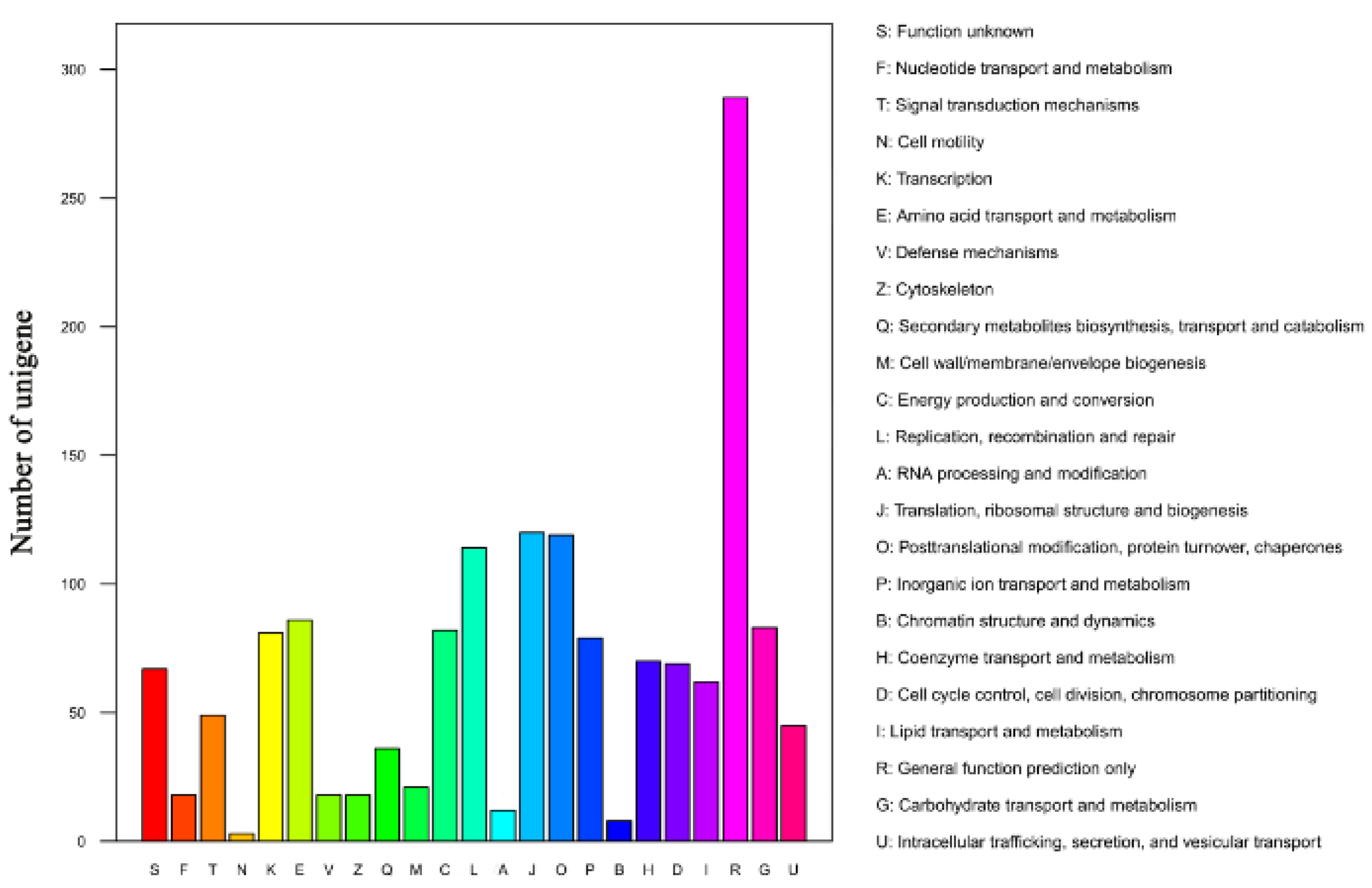
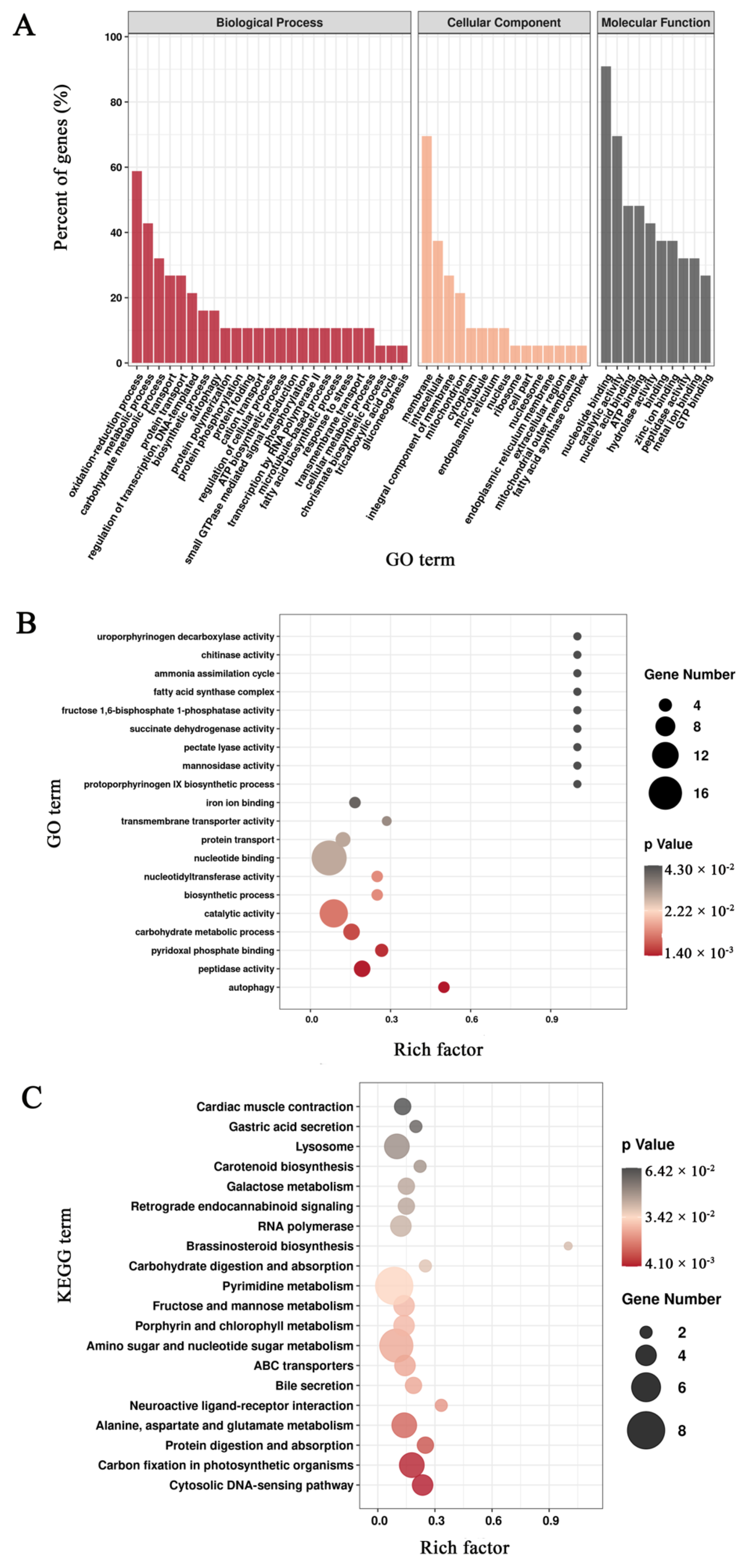
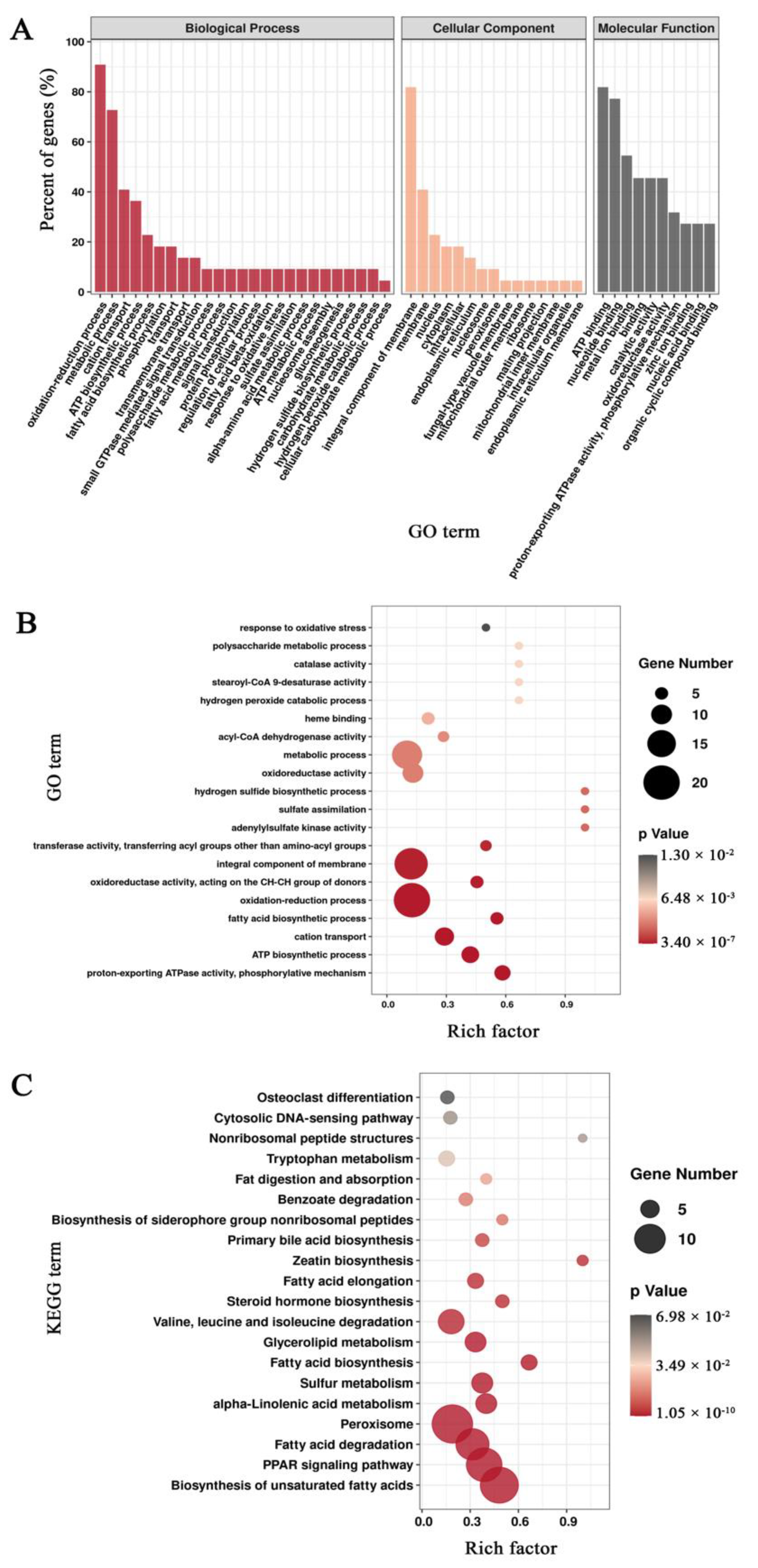
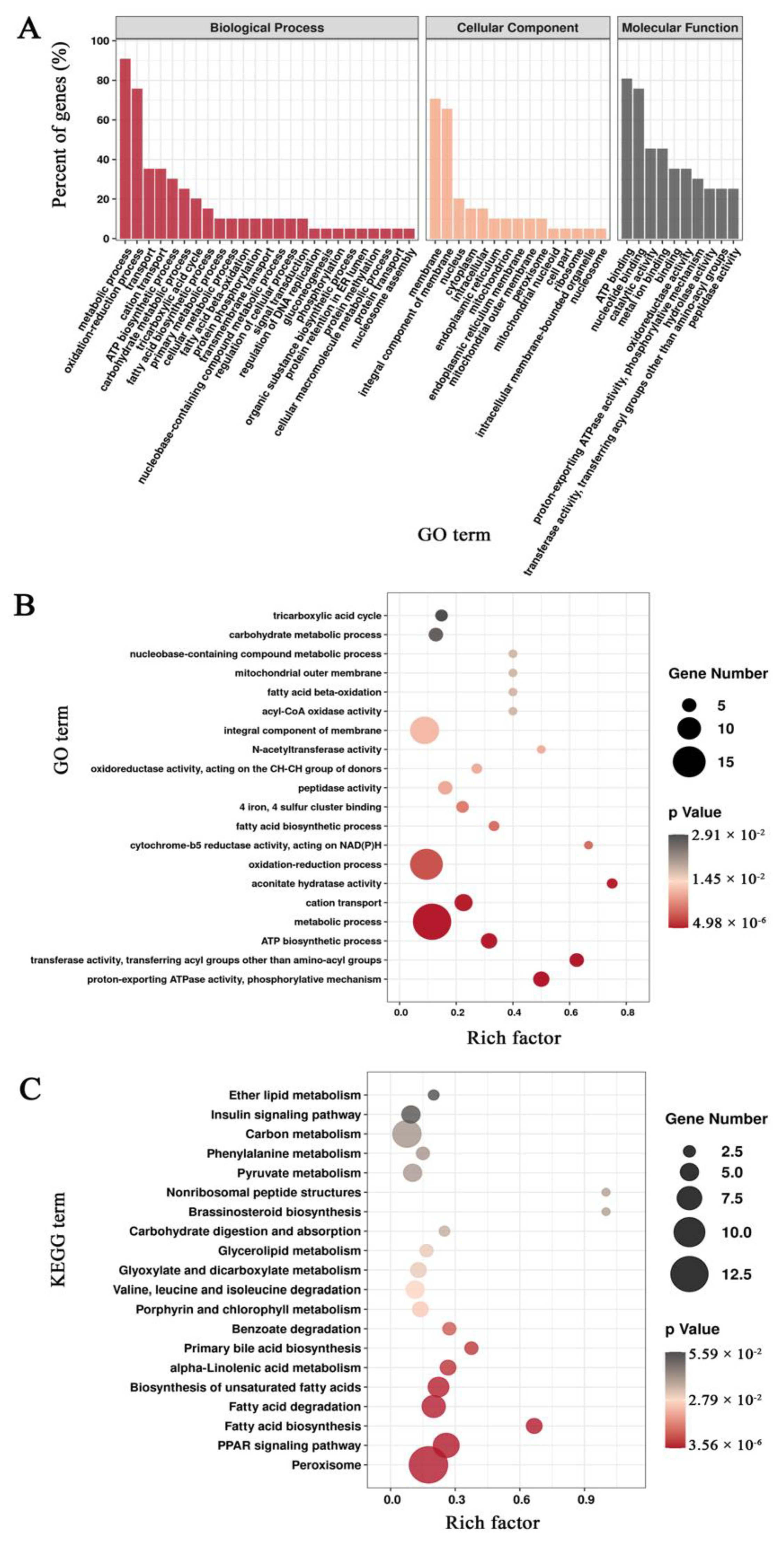
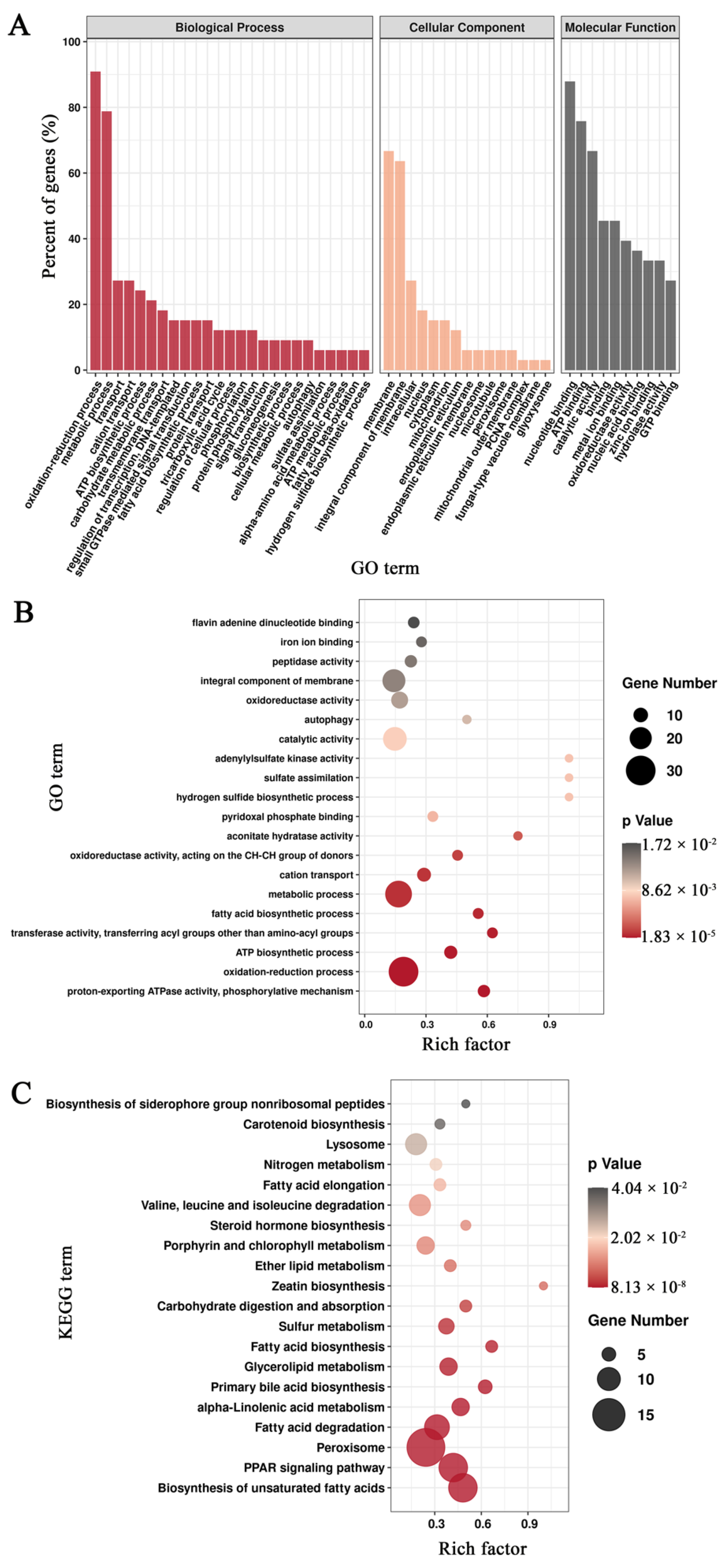
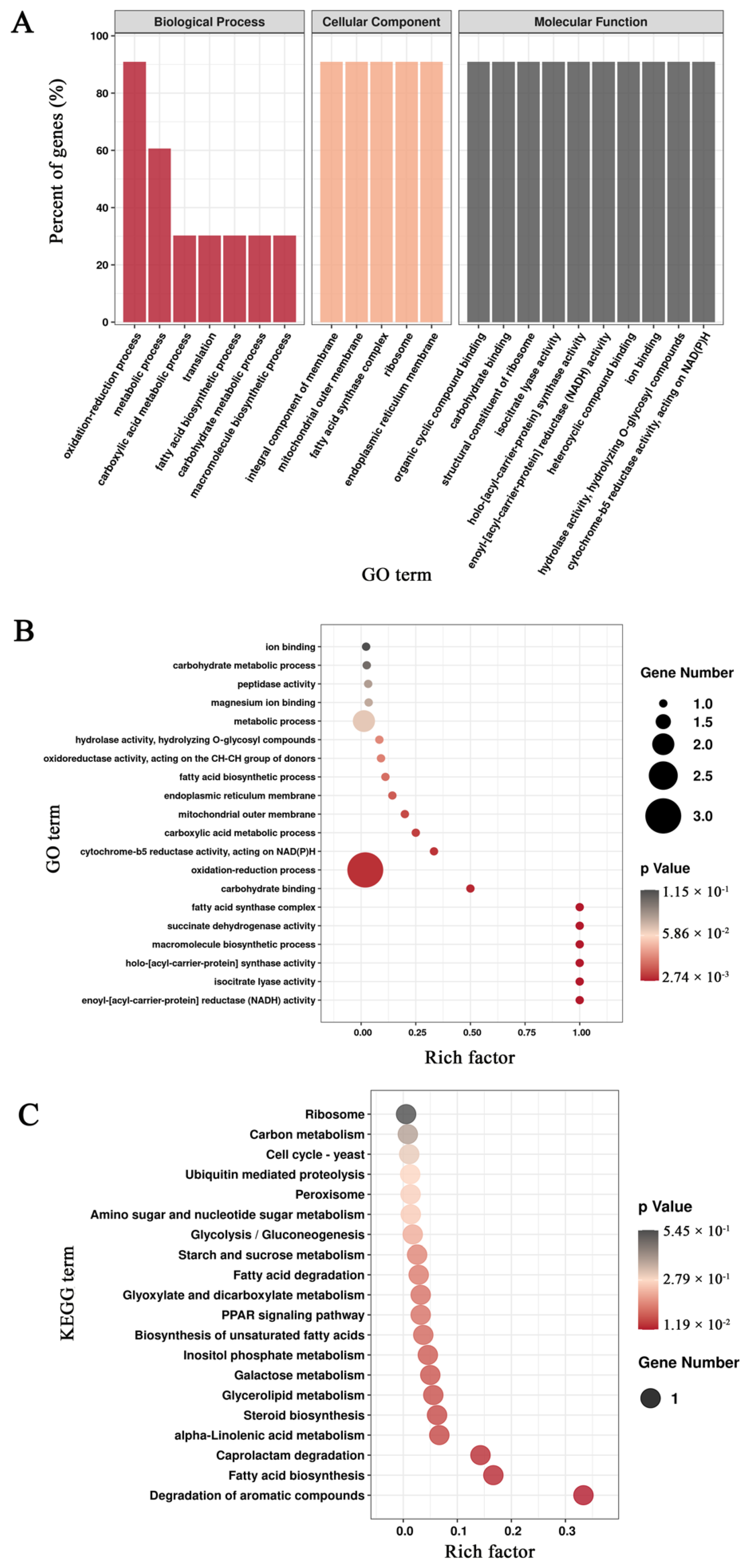
3.3. Functional Annotation of Unigenes in Mortierella alpina
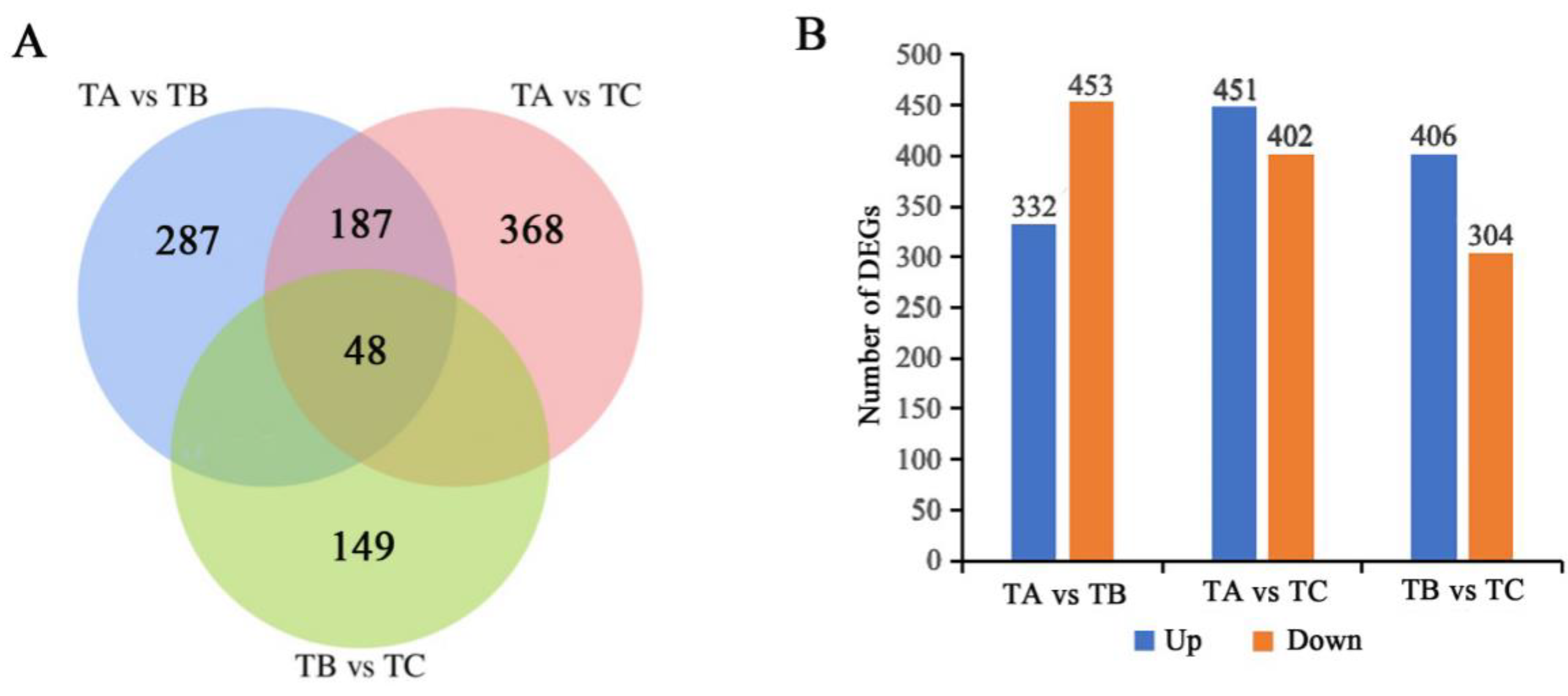
3.4. Identification of Differentially Expressed Unigenes in Mortierella alpina
3.5. Enrichment Analysis of Differentially Expressed Genes Based on the Pairwise Comparisons of Mortierella alpina
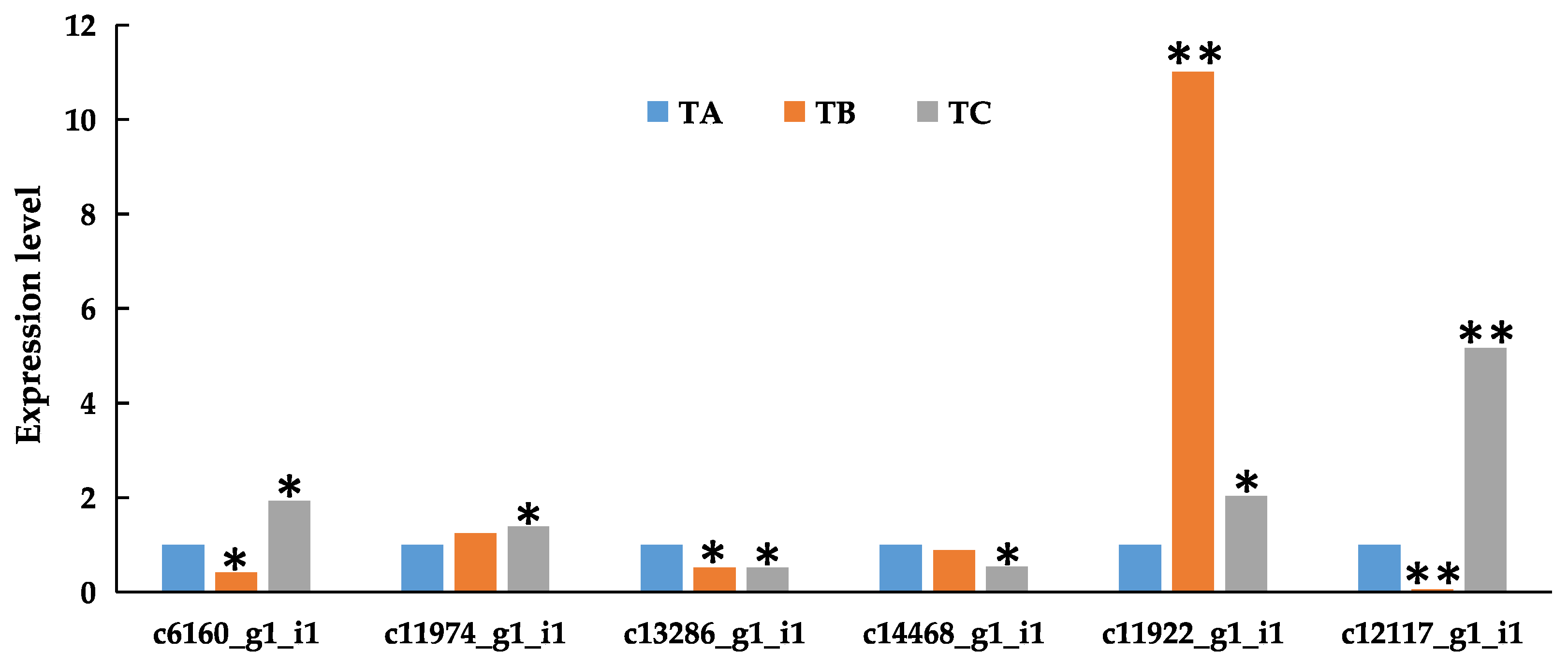
3.6. Verification of the Expression of Differentially Expressed Genes by qRT-PCR Analysis
3.7. Expression of Genes Involved in the Synthesis of Polyunsaturated Fatty Acids
4. Discussion
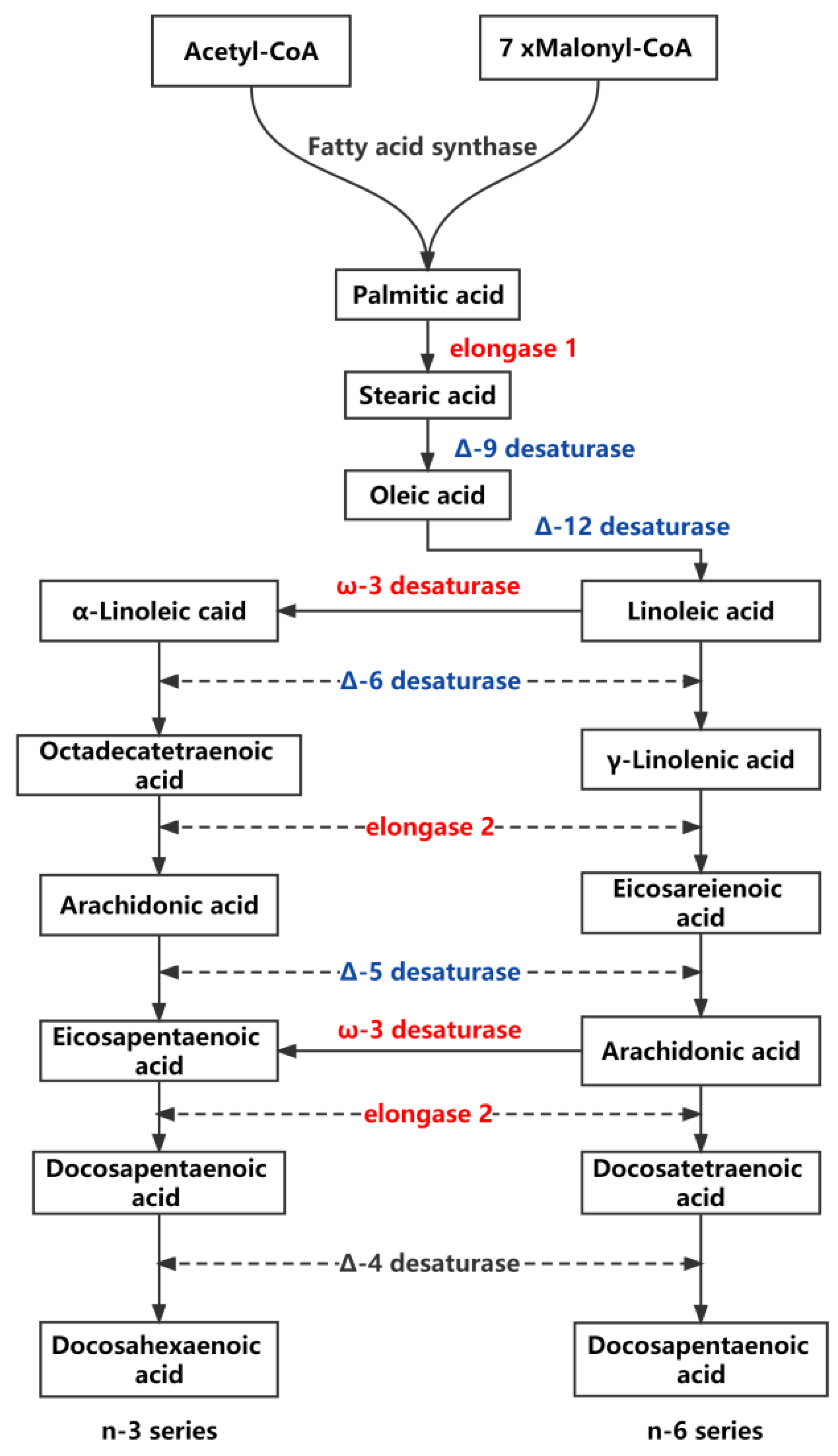
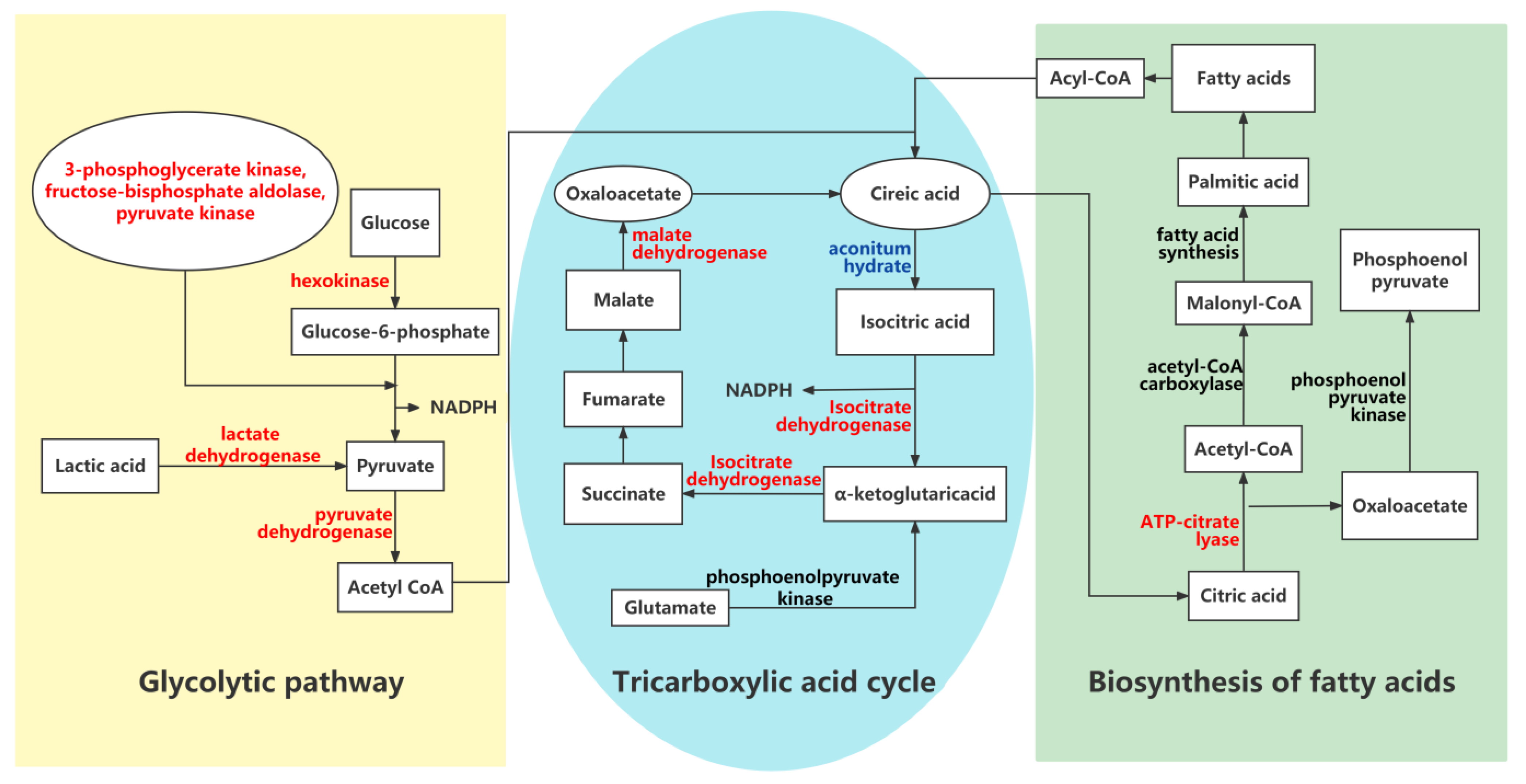
4.1. Fatty Acid Metabolic Pathway
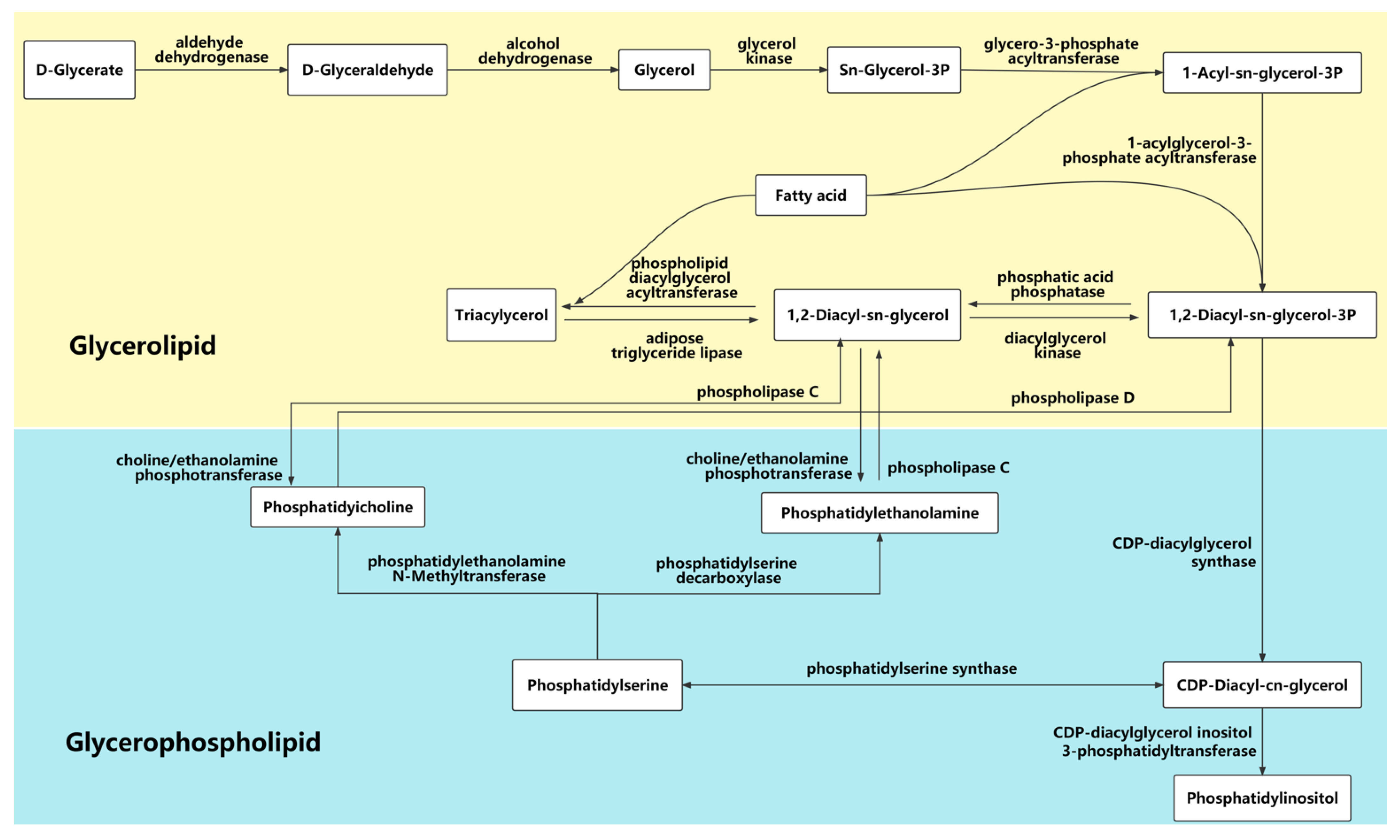
4.2. Glycerolipid and Glycerophospholipid Metabolic Pathways
4.3. Acetyl-CoA and NADPH Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ji, X.J.; Ren, L.J.; Nie, Z.K.; Huang, H.; Ouyang, P.K. Fungal arachidonic acid-rich oil: Research, development and industrialization. Crit. Rev. Biotechnol. 2014, 34, 197–214. [Google Scholar] [CrossRef]
- Samadlouie, H.; Hamidi-Esfahani, Z.; Soltani-Najafabadi, S.M.A.M.; Sahari, M.A.; Abbasi, S. Statistical approach to optimization of fermentative production of oil and arachidonic acid from Mortierella alpina CBS 754.68. Afr. J. Microbiol. Res. 2012, 6, 1559–1567. [Google Scholar]
- Birch, E.E. Visual Acuity and the Essentiality of Docosahexaenoic Acid and Arachidonic Acid in the Diet of Term Infants. Pediatr. Res. 1998, 44, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Nisha, A.; Venkateswaran, G. Effect of Culture Variables on Mycelial Arachidonic acid Production by Mortierella alpina. Food Bioproc. Technol. 2011, 4, 232–240. [Google Scholar]
- Wu, W.J.; Zhang, A.H.; Peng, C.; Ren, L.J.; Song, P.; Yu, Y.D.; Huang, H.; Ji, X.J. An efficient multi-stage fermentation strategy for the production of microbial oil rich in arachidonic acid in Mortierella alpina. Bioresour. Bioprocess. 2017, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Sakuradani, E.; Shimizu, S. Single cell oil production by Mortierella alpina. J. Biotechnol. 2009, 144, 31–36. [Google Scholar]
- Wang, L.; Chen, W.; Feng, Y.; Ren, Y.; Gu, Z.; Chen, H.; Wang, H.; Thomas, M.J.; Zhang, B.; Berquin, I.M.; et al. Genome characterization of the oleaginous fungus Mortierella alpina. PLoS ONE 2011, 6, e28319. [Google Scholar] [CrossRef] [Green Version]
- Etienne, K.A.; Chibucos, M.C.; Su, Q.; Orvis, J.; Daugherty, S.; Ott, S.; Sengamalay, N.A.; Fraser, C.M.; Lockhart, S.R.; Bruno, V.M. Draft genome sequence of Mortierella alpina isolate CDC-B6842. Genome Announc. 2014, 2, 2007–2008. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Hao, G.; Wang, L.; Wang, H.; Gu, Z.; Liu, L.; Zhang, H.; Chen, W.; Chen, Y.Q. Identification of a critical determinant that enables efficient fatty acid synthesis in oleaginous fungi. Sci. Rep. 2015, 5, 11247. [Google Scholar] [CrossRef]
- Yu, Y.; Li, T.; Wu, N.; Ren, L.; Jiang, L.; Ji, X.; Huang, H. Mechanism of Arachidonic Acid Accumulation during Aging in Mortierella alpina: A Large-Scale Label-Free Comparative Proteomics Study. J. Agric. Food Chem. 2016, 64, 9124–9134. [Google Scholar]
- Lu, H.; Chen, H.; Tang, X.; Yang, Q.; Zhang, H.; Chen, Y.Q.; Chen, W. Time-resolved multi-omics analysis reveals the role of nutrient stress-induced resource reallocation for TAG accumulation in oleaginous fungus Mortierella alpina. Biotechnol. Biofuels 2020, 13, 116. [Google Scholar] [CrossRef]
- Okuda, T.; Ando, A.; Negoro, H.; Kikukawa, H.; Sakamoto, T.; Sakuradani, E.; Shimizu, S.; Ogawa, J. Omega-3 eicosatetraenoic acid production by molecular breeding of the mutant strain S14 derived from Mortierella alpina 1S-4. J. Biosci. Bioeng. 2015, 120, 299–304. [Google Scholar] [CrossRef]
- Vadivelan, G.; Venkateswaran, G. Production and enhancement of Omega-3 fatty acid from Mortierella alpina CFR-GV15: Its food and therapeutic application. Biomed Res. Int. 2014, 9, 657414. [Google Scholar]
- Okuda, T.; Ando, A.; Negoro, H.; Muratsubaki, T.; Kikukawa, H.; Sakamoto, T.; Sakuradani, E.; Shimizu, S.; Ogawa, J. Eicosapentaenoic acid (EPA) production by an oleaginous fungus Mortierella alpina expressing heterologous the Δ-17-desaturase gene under ordinary temperature. Eur. J. Lipid Sci. Technol. 2015, 117, 1919–1927. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Bruneel, C.; Muylaert, K.; Foubert, I. Microalgae as an alternative source of omega-3 long chain polyunsaturated fatty acids. Lipid Technol. 2012, 24, 128–130. [Google Scholar] [CrossRef]
- Winwood, R.J. Recent developments in the commercial production of DHA and EPA rich oils from micro-algae. OCL Oilseeds Fats Crops Lipids 2013, 20, D604. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhao, X.; Strait, M.; Wen, Z. Use of dry-milling derived thin stillage for producing eicosapentaenoic acid (EPA) by the fungus Pythium irregulare. Bioresour. Technol. 2012, 111, 404–409. [Google Scholar] [CrossRef]
- Dedyukhina, E.G.; Chistyakova, T.I.; Mironov, A.A.; Kamzolova, S.V.; Minkevich, I.G.; Vainshtein, M.B. The effect of pH, aeration, and temperature on arachidonic acid synthesis by Mortierella alpina. Appl. Biochem. Microbiol. 2015, 51, 242–248. [Google Scholar] [CrossRef]
- Li, X.; Lin, Y.; Chang, M.; Jin, Q.; Wang, X. Efficient production of arachidonic acid by Mortierella alpina through integrating fed-batch culture with a two-stage pH control strategy. Bioresour. Technol. 2015, 181, 275–282. [Google Scholar] [CrossRef]
- Qin, X.Z.; Yang, X.P.; Chen, J.; Feng, L. Optimization of incubated mycelium of Mortierella alpina based on response surface methodology. Xinjiang Agric. Sci. 2014, 51, 89–97. [Google Scholar]
- Huang, M.; Chen, H.; Tang, X.; Lu, H.; Chen, W. Two-stage ph control combined with oxygen-enriched air strategies for the highly efficient production of EPA by Mortierella alpina ccfm698 with fed-batch fermentation. Bioprocess Biosyst. Eng. 2020, 43, 1725–1733. [Google Scholar] [CrossRef]
- Nie, Z.K.; Ji, X.J.; Shang, J.S.; Zhang, A.H.; Ren, L.J.; Huang, H. Arachidonic acid-rich oil production by Mortierella alpina with different gas distributors. Bioprocess Biosyst. Eng. 2014, 37, 1127–1132. [Google Scholar] [CrossRef]
- Ling, X.; Zeng, S.; Chen, C.; Liu, X.; Lu, Y. Enhanced arachidonic acid production using a bioreactor culture of Mortierella alpina with a combined organic nitrogen source. Bioresour. Bioprocess. 2016, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Liao, X.; Gao, Z.; Li, Y.; Lei, Y.; Chen, W.; Chen, L.; Fan, X.; Zhang, K.; Chen, S.; et al. Effects of methanol on carotenoids as well as biomass and fatty acid biosynthesis in Schizochytrium limacinum B4D1. Appl. Environ. Microbiol. 2019, 85, 19. [Google Scholar] [CrossRef] [Green Version]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [Green Version]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Dewey, C.N.; Li, B. Rsem: Accurate transcript quantification from RNA-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar]
- Robinson, M.D.; Smyth, G.K. Moderated statistical tests for assessing differences in tag abundance. Bioinformatics 2007, 23, 2881–2887. [Google Scholar] [CrossRef] [Green Version]
- Bellou, S.; Triantaphyllidou, I.E.; Aggeli, D.; Elazzazy, A.M.; Baeshen, M.N.; Aggelis, G. Microbial oils as food additives: Recent approaches for improving microbial oil production and its polyunsaturated fatty acid content. Curr. Opin. Biotechnol. 2016, 37, 24–35. [Google Scholar] [CrossRef]
- Kothri, M.; Mavrommati, M.; Elazzazy, A.M.; Baeshen, M.N.; Moussa, T.A.A.; Aggelis, G. Microbial sources of polyunsaturated fatty acids (PUFAs) and the prospect of organic residues and wastes as growth media for PUFA-producing microorganisms. FEMS Microbiol. Lett. 2020, 367, fnaa028. [Google Scholar] [CrossRef]
- Wongwathanarat, P.; Michaelson, L.V.; Carter, A.T.; Lazarus, C.M.; Griffiths, G.; Stobart, A.K.; Archer, D.B.; MacKenzie, D.A. Two fatty acid Delta 9-desaturase genes, ole1 and ole2, from Mortierella alpina complement the yeast ole1 mutation. Microbiology 1999, 145, 2939–2946. [Google Scholar] [CrossRef] [Green Version]
- Hao, G.; Chen, H.; Yang, B.; Du, K.; Wang, H.; Gu, Z.; Zhang, H.; Chen, W.; Chen, Y.Q. Substrate specificity of Mortierella alpina Δ-9-III fatty acid desaturase and its value for the production of omega-9 MUFA. Eur. J. Lipid Sci. Technol. 2016, 118, 753–760. [Google Scholar] [CrossRef]
- Huang, Y.S.; Chaudhary, S.; Thurmond, J.M.; Bobik, E.G.; Yuan, L.; Chan, G.M.; Kirchner, S.J.; Mukerji, P.; Knutzon, D.S. Cloning of Delta 12-and Delta 6- desaturases from Mortierella alpina and recombinant production of gamma-linolenic acid in Saccharomyces cerevisiae. Lipids 1999, 34, 649–659. [Google Scholar] [CrossRef]
- Wang, C.; Qing, X.; Yu, M.; Sun, Q.; Liu, F.; Qi, B.; Li, X. Production of eicosapentaenoic acid (EPA, 20:5n-3) in transgenic peanut (Arachis hypogaea L.) through the alternative Delta 8-desaturase pathway. Mol. Biol. Rep. 2019, 46, 333–342. [Google Scholar] [CrossRef]
- Sakuradani, E.; Abe, T.; Iguchi, K.; Shimizu, S. A novel fungal omega 3-desaturase with wide substrate specificity from arachidonic acid-producing Mortierella alpina 1S-4. Appl. Microbiol. Biotechnol. 2005, 66, 648–654. [Google Scholar] [CrossRef]
- Tv, A.; Ct, A.; Tp, B.; Ts, C.; Pj, D.; Tw, D. Systematic genome analysis of a novel arachidonic acid-producing strain uncovered unique metabolic traits in the production of acetyl-CoA-derived products in mortierellale fungi. Gene 2020, 741, 144550. [Google Scholar]
- Chang, L.; Chen, H.; Tang, X.; Zhao, J.; Zhang, H.; Chen, Y.Q.; Chen, W. Advances in improving the biotechnological application of oleaginous fungus Mortierella alpina. Appl. Microbiol. Biotechnol. 2021, 105, 6275–6289. [Google Scholar] [CrossRef]
- Houten, S.M.; Wanders, R.J.A. A general introduction to the biochemistry of mitochondrial fatty acid β-oxidation. J. Inherit. Metab. Dis. 2010, 33, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Adams, I.P.; Ratledge, C. Malic enzyme: The controlling activity for lipid production? Overexpression of malic enzyme in Mucor circinelloides leads to a 2.5-fold increase in lipid accumulation. Microbiology 2007, 153, 2013–2025. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
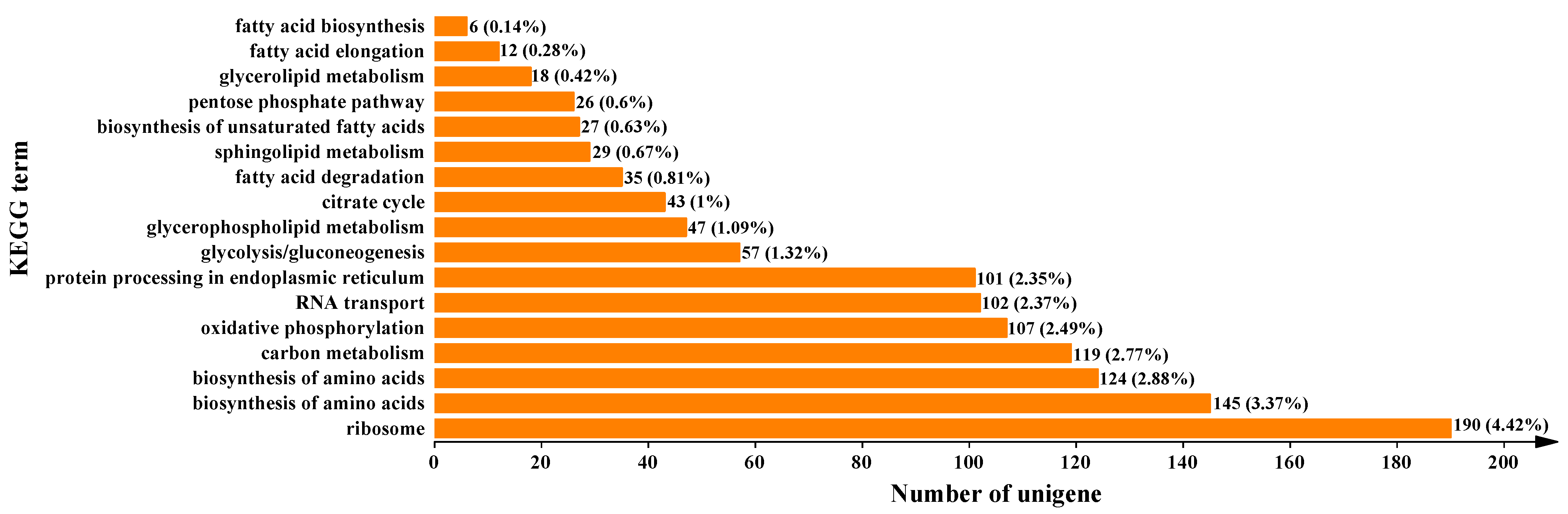{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Lipid Content (%) | Biomass (g/L) |
|---|---|---|
| TA (25 °C) | 17.59 ± 0.27 | 38.04 ± 0.24 |
| TB (15 °C) | 9.89 ± 0.41 ** | 32.54 ± 0.52 ** |
| TC (25 °C–15 °C) | 15.58 ± 0.13 ** | 36.03 ± 0.34 ** |
| Fatty Acid | TA (25 °C) | TB (15 °C) | TC (25 °C–15 °C) |
|---|---|---|---|
| Tetradecanoic acid (C14:0) | 0.29 ± 0.035 | 0.678 ± 0.061 ** | 0.35 ± 0.027 * |
| Palmitic acid (C16:0) | 7.84 ± 0.145 | 9.60 ± 0.225 ** | 8.33 ± 0.232 ** |
| Stearic acid (C18:0) | 11.20 ± 0.917 | 15.30 ± 0.874 ** | 11.29 ± 0.657 |
| Oleic acid (C18:1n9t) | 0.06 ± 0.013 | 0.07 ± 0.011 * | 0.05 ± 0.013 * |
| Oleic acid (C18:1n9c) | 7.58 ± 0.750 | 5.36 ± 0.53 ** | 7.36 ± 0.032 ** |
| Linoleic acid (C18:2n6c) | 5.82 ± 0.232 | 4.11 ± 0.211 ** | 5.44 ± 0.307 ** |
| r-linolenic acid (C18:3n6) | 4.34 ± 0.371 | 3.61 ± 0.341 ** | 4.12 ± 0.429 ** |
| Arachidic acid (C20:0) | 0.58 ± 0.043 | 0.55 ± 0.026 * | 0.56 ± 0.033 ** |
| Eicosenoic acid (C20:1) | 0.30 ± 0.029 | 0.17 ± 0.014 ** | 0.32 ± 0.028 * |
| Eicosadienoic acid (C20:2) | 0.48 ± 0.054 | 0.41 ± 0.043 ** | 0.52 ± 0.056 ** |
| Eicosatrienoic acid (C20:3) | 4.40 ± 0.238 | 7.45 ± 0.657 ** | 4.66 ± 0.382 ** |
| Arachidonic acid (C20:4) | 56.40 ± 1.835 | 51.77 ± 1.677 ** | 56.59 ± 1.971 * |
| Eicosapentaenoic acid (C20:5) | 0.01 ± 0.001 | 0.22 ± 0.014 ** | 0.22 ± 0.021 ** |
| Sample | Length (nt) | Reads | Nucleotide | Q20 (%) | Q30 (%) | GC (%) | N (ppm) |
|---|---|---|---|---|---|---|---|
| TA | 101 (100.27) | 55,701,908 (46,514,296) | 5,625,892,708 (4,663,901,130) | 95.57 (99.69) | 90.44 (96.78) | 53.72 (53.51) | 933.86 (0.46) |
| TB | 101 (100.24) | 48,534,748 (40,287,982) | 4,902,009,548 (4,038,356,115) | 95.45 (99.67) | 90.06 (96.62) | 53.88 (53.61) | 927.96 (0.46) |
| TC | 101 (100.24) | 65,639,098 (55,154,048) | 6,629,548,898 (5,528,511,690) | 95.89 (99.69) | 90.77 (96.73) | 54.25 (54.00) | 938.59 (0.46) |
| Assembled Unit | Total | <200 nt | (200–500) nt | (500–1000) nt | (1000–1500) nt | (1500–2000) nt | ≥2000 nt |
|---|---|---|---|---|---|---|---|
| Contig | 1,417,688 (100) | 1,397,239 (98.56) | 9006 (0.64) | 3761 (0.27) | 2017 (0.14) | 1633 (0.12) | 4031 (0.28) |
| Trinity assembly | 23,727 (100) | 0 | 8159 (34.39) | 4254 (17.93) | 2668 (11.24) | 2238 (9.43) | 6407 (27) |
| Unigene (TGICL) | 22,059 (100) | 0 | 7936 (35.98) | 4048 (18.35) | 2495 (11.31) | 2039 (9.24) | 5540 (25.11) |
| Sample | Total Read | Total Read Mapped (%) | Unique Read Mapped (%) | Multi-Region Mapped (%) |
|---|---|---|---|---|
| TA | 46,514,296 | 43,589,424 (93.71) | 39,026,470 (83.90) | 4,562,954 (9.81) |
| TB | 40,287,982 | 37,644,954 (93.44) | 33,595,404 (83.39) | 4,049,550 (10.05) |
| TC | 55,154,048 | 51,442,932 (93.27) | 45,945,988 (83.30) | 5,496,944 (9.97) |
| Gene | Gene Annotation | TA | TB | TC |
|---|---|---|---|---|
| c6160_g1_i1 | Sterol desaturase | 1.38 | 0.57 | 2.66 |
| c11974_g1_i1 | Delta-6 fatty acid desaturase | 28.3 | 35.2 | 39.27 |
| c13286_g1_i1 | Delta-5 fatty acid desaturase | 552.74 | 287.16 | 287.03 |
| c14468_g1_i1 | Delta-6 fatty acid elongase | 885.54 | 790.17 | 473.17 |
| c11922_g1_i1 | OPT oligopeptide transporter | 30.2 | 332.6 | 61.63 |
| c12117_g1_i1 | Isocitrate lyase | 74.24 | 4.76 | 383.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, J.; Wang, H.; Sun, F.; Meng, C.; Zhu, X.; Wei, Z.; Gao, Z.; Li, D. Transcriptomic Characterizations of the Molecular Mechanisms Regulating Lipid Production and Composition in Mortierella alpina in Response to Cold Stress. Agronomy 2022, 12, 599. https://doi.org/10.3390/agronomy12030599
Ren J, Wang H, Sun F, Meng C, Zhu X, Wei Z, Gao Z, Li D. Transcriptomic Characterizations of the Molecular Mechanisms Regulating Lipid Production and Composition in Mortierella alpina in Response to Cold Stress. Agronomy. 2022; 12(3):599. https://doi.org/10.3390/agronomy12030599
Chicago/Turabian StyleRen, Jiali, Haijun Wang, Fengjie Sun, Chunxiao Meng, Xiangyu Zhu, Zuoxi Wei, Zhengquan Gao, and Demao Li. 2022. "Transcriptomic Characterizations of the Molecular Mechanisms Regulating Lipid Production and Composition in Mortierella alpina in Response to Cold Stress" Agronomy 12, no. 3: 599. https://doi.org/10.3390/agronomy12030599