
Growth of Vegetables in an Agroecological Garden-Orchard System: The Role of Spatiotemporal Variations of Microclimatic Conditions and Soil Properties


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
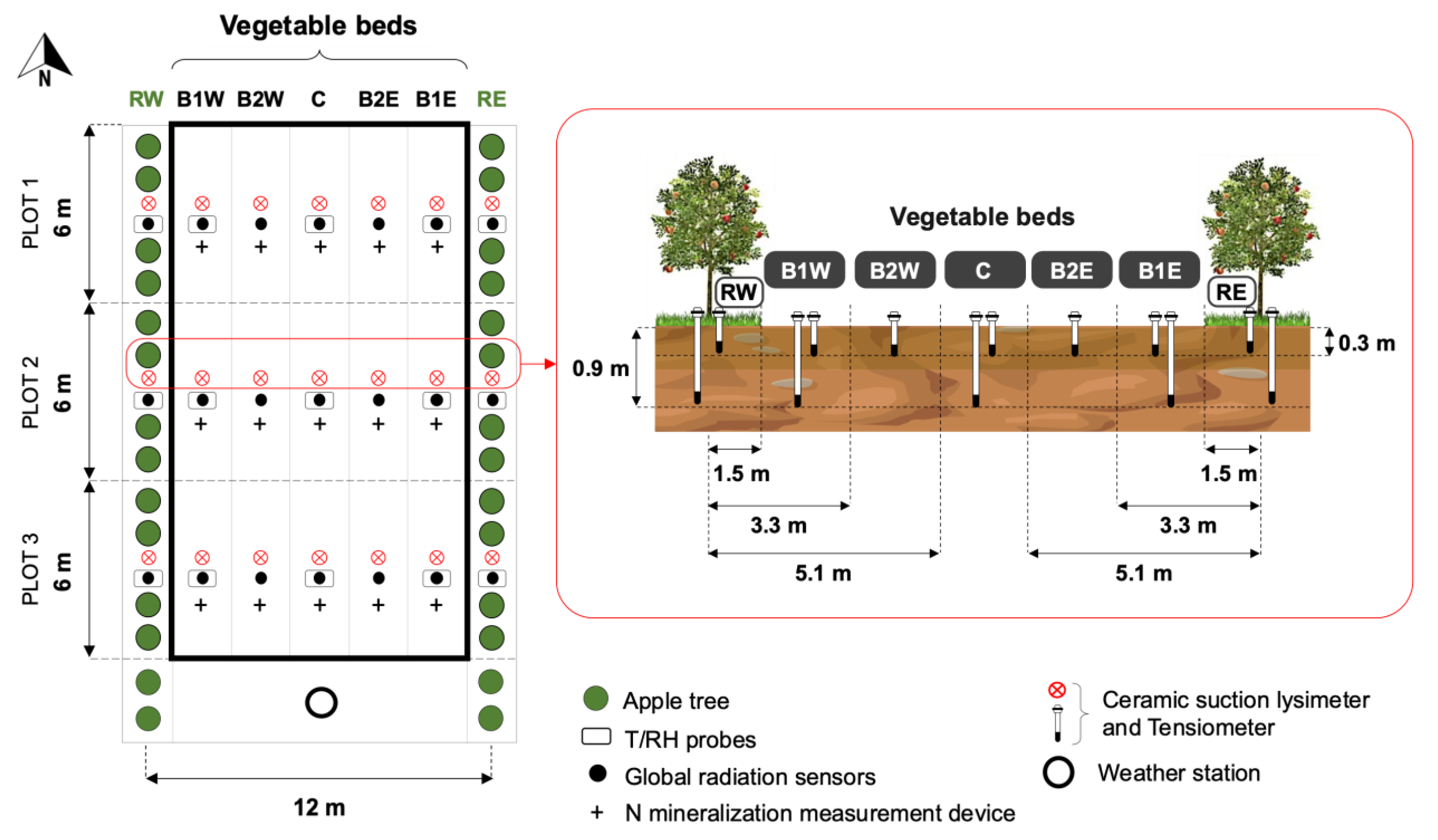
2.2. Experimental Design
2.3. Microclimate Characterization
2.4. Soil Properties Measurements
2.4.1. Soil Water Content
2.4.2. Soil Bulk Density
2.4.3. Soil Organic Matter
2.4.4. N Mineralization
2.4.5. Mineral Nitrogen Concentration in Soil and Soil Solution
2.5. Vegetable Growth and Yield
2.6. Data Analysis
3. Results
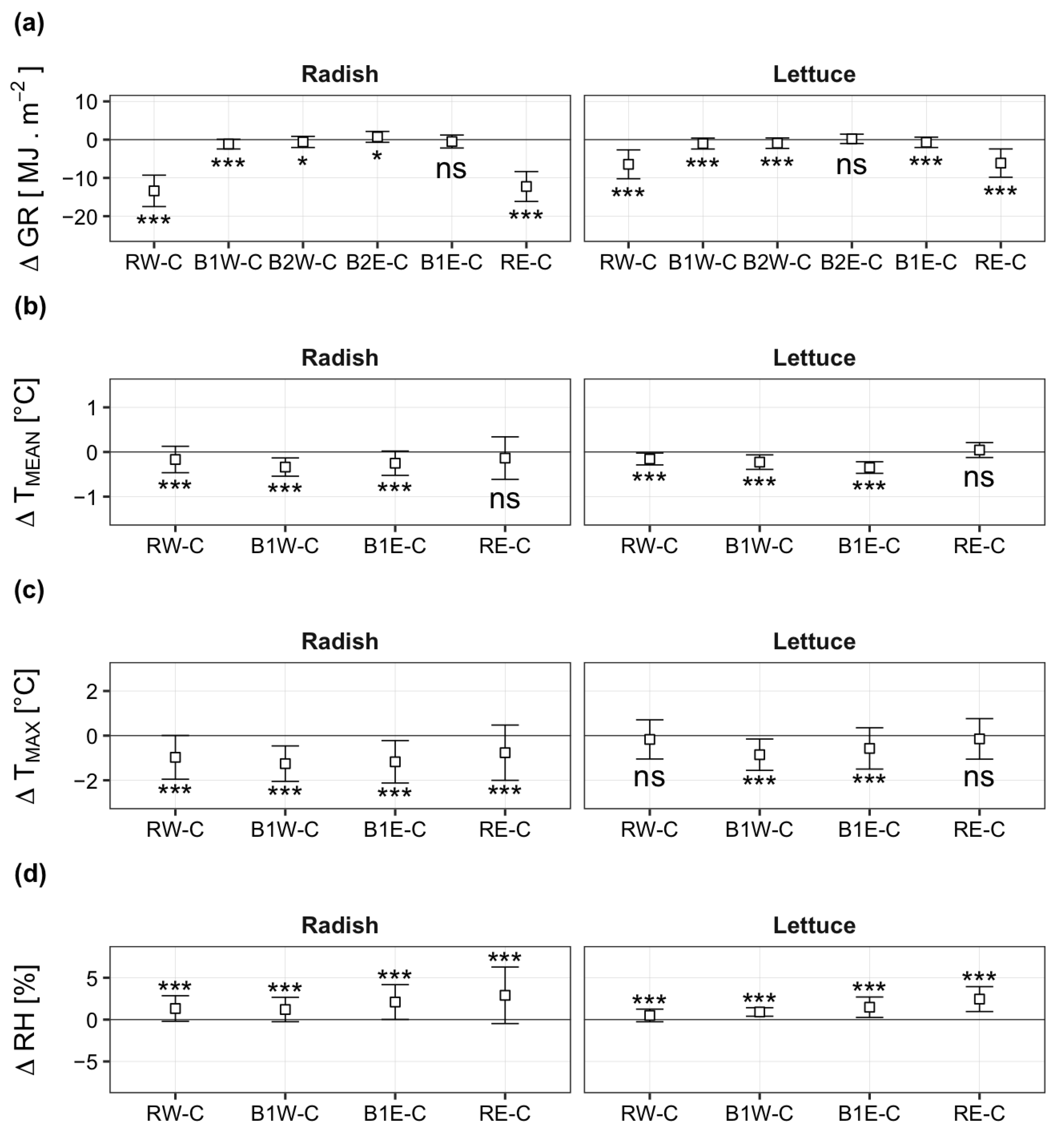
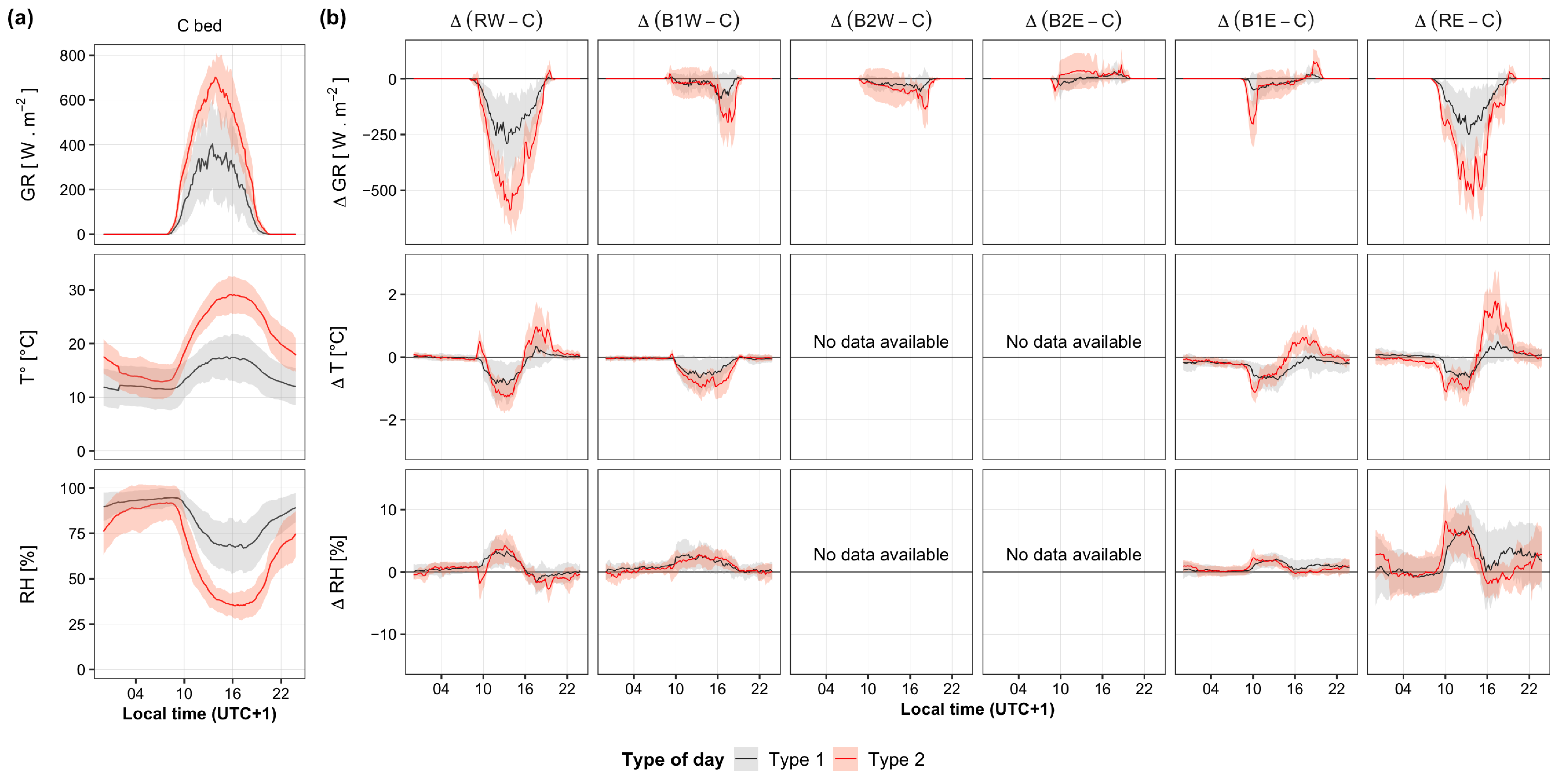
3.1. Spatiotemporal Variation of Microclimatic Conditions
- Day type 1: cloudy sky, with cold and humid conditions
- Day type 2: clear sky, with warm and dry conditions
3.2. Spatiotemporal Variation of Soil Properties
3.2.1. Soil Water Content
3.2.2. Soil Bulk Density and Soil Organic Matter
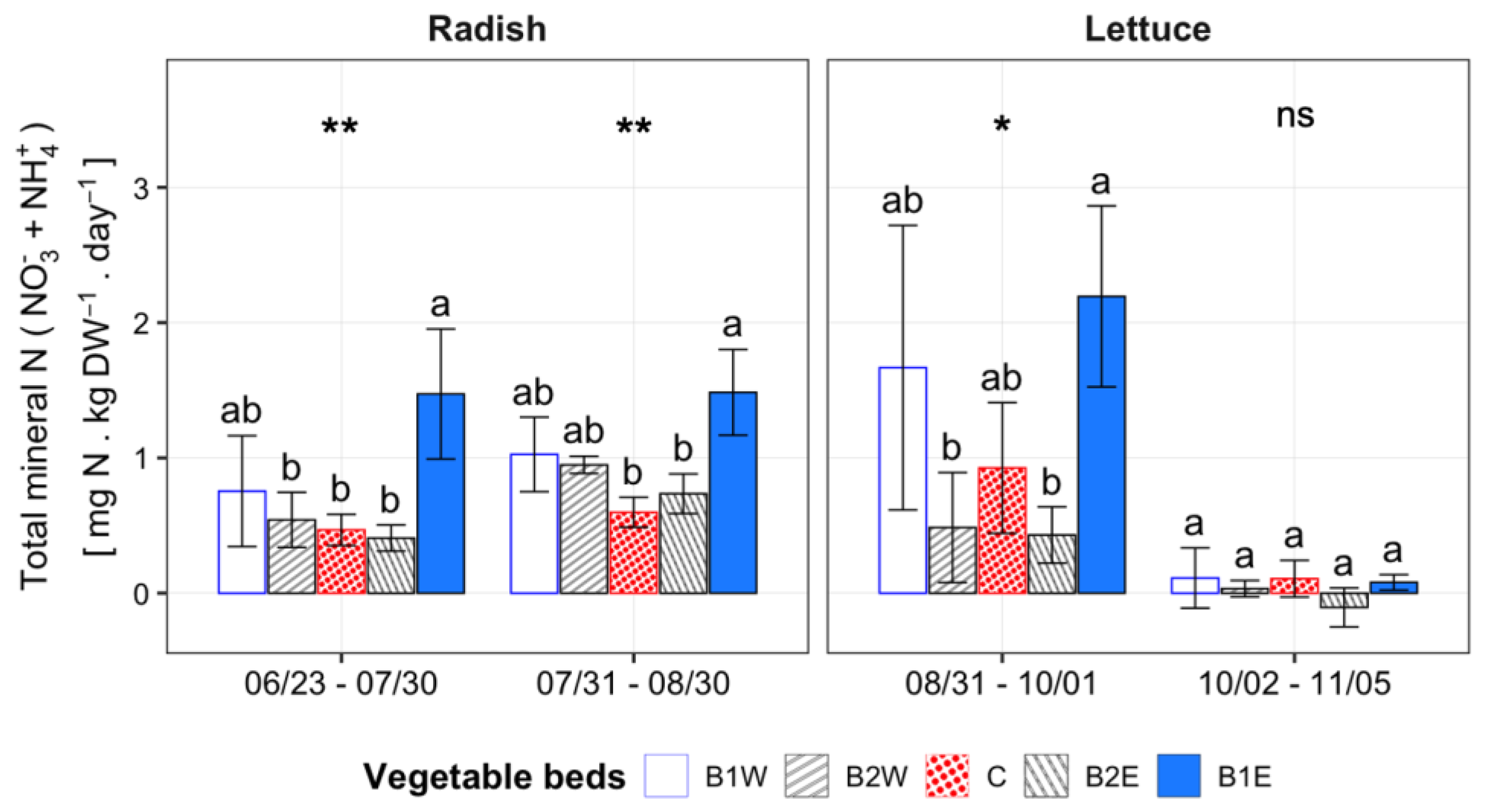
3.2.3. N Mineralization
3.2.4. Soil Mineral Nitrogen
3.2.5. Mineral Nitrogen in Soil Solution
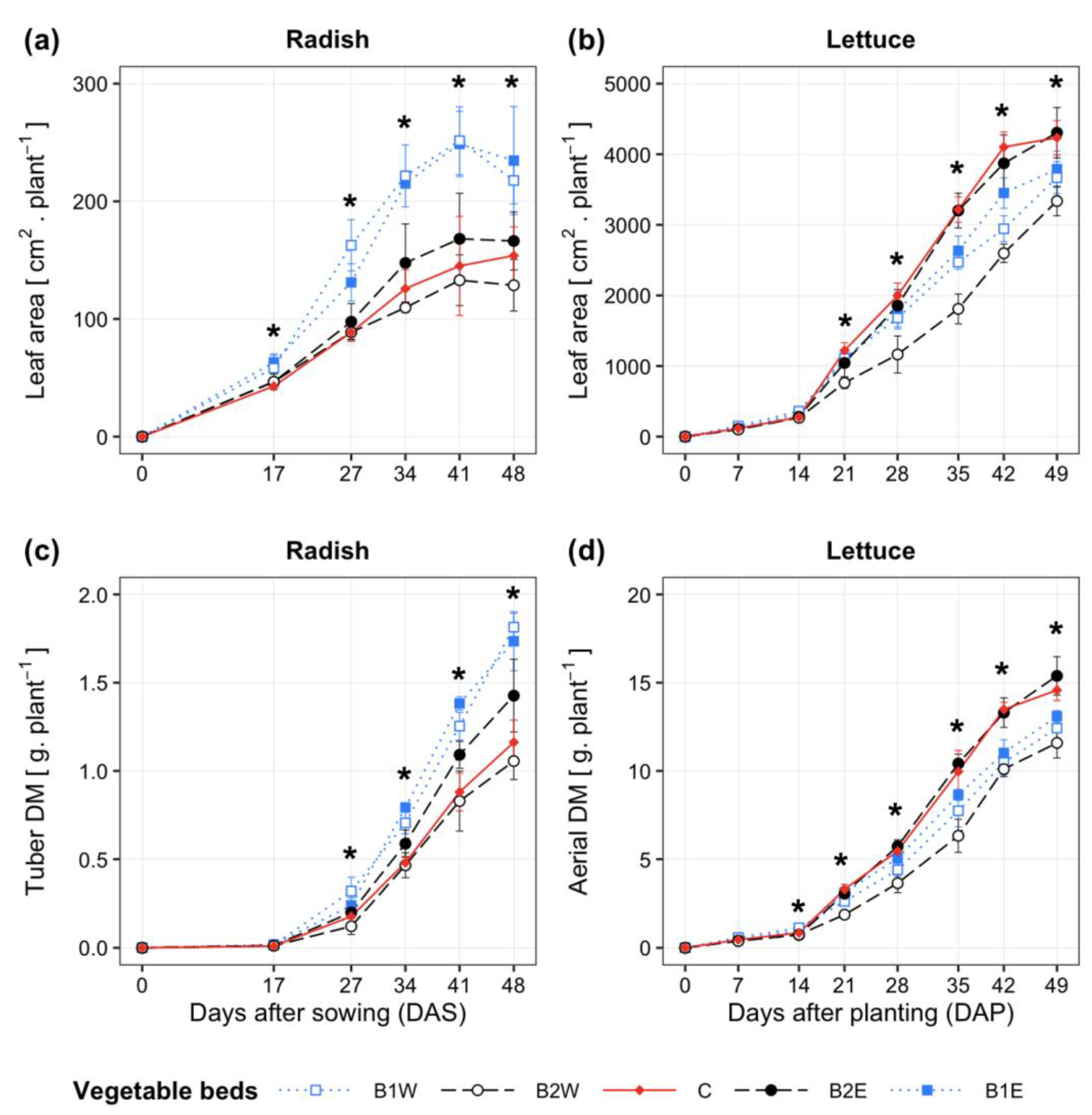
3.3. Crop Growth Properties
3.4. Correlations between Variables
4. Discussion
4.1. Effects of Apple Trees on the Spatial and Temporal Variations of Microclimatic Conditions
4.2. Effects of Apple Trees on the Spatial Variation of Soil Properties
4.2.1. Soil Water Content, Soil Organic Matter and Bulk Density
4.2.2. N mineralization and Soil Mineral Nitrogen
4.3. Effects of Microclimatic Conditions and Soil Properties on Vegetable Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bado, B.V.; Whitbread, A.; Sanoussi Manzo, M.L. Improving Agricultural Productivity Using Agroforestry Systems: Performance of Millet, Cowpea, and Ziziphus-Based Cropping Systems in West Africa Sahel. Agric. Ecosyst. Environ. 2021, 305, 107175. [Google Scholar] [CrossRef]
- Schwendenmann, L.; Pendall, E.; Sanchez-Bragado, R.; Kunert, N.; Hölscher, D. Tree Water Uptake in a Tropical Plantation Varying in Tree Diversity: Interspecific Differences, Seasonal Shifts and Complementarity. Ecohydrology 2015, 8, 1–12. [Google Scholar] [CrossRef]
- Paut, R.; Sabatier, R.; Tchamitchian, M. Modelling Crop Diversification and Association Effects in Agricultural Systems. Agric. Ecosyst. Environ. 2020, 288, 106711. [Google Scholar] [CrossRef]
- Temani, F.; Bouaziz, A.; Daoui, K.; Wery, J.; Barkaoui, K. Olive Agroforestry Can Improve Land Productivity Even under Low Water Availability in the South Mediterranean. Agric. Ecosyst. Environ. 2021, 307, 107234. [Google Scholar] [CrossRef]
- Lehmann, J.; Weigl, D.; Droppelmann, K.; Huwe, B.; Zech, W. Nutrient Cycling in an Agroforestry System with Runoff Irrigation in Northern Kenya. Agrofor. Syst. 1998, 43, 49–70. [Google Scholar] [CrossRef]
- Ling, Q.; Gao, X.; Zhao, X.; Huang, J.; Li, H.; Li, L.; Sun, W.; Wu, P. Soil Water Effects of Agroforestry in Rainfed Jujube (Ziziphus jujube Mill.) Orchards on Loess Hillslopes in Northwest China. Agric. Ecosyst. Environ. 2017, 247, 343–351. [Google Scholar] [CrossRef]
- FranceAgriMer. Pratiques et Habitudes de Consommation des Fruits et Légumes. 2016. Available online: https://www.franceagrimer.fr/content/download/43681/416109/file/Synthèse_pratiques_habitudes_F&L.pdf (accessed on 10 April 2019).
- Ouma, G.; Jeruto, P. Sustainable Horticultural Crop Production through Intercropping: The Case of Fruits and Vegetable Crops: A Review. Agric. Biol. J. N. Am. 2010, 1, 1098–1105. [Google Scholar] [CrossRef]
- Ahmed, F.; Monika, A.; Hossain, M.; Wadud, M. Performance of Guava Fruit Tree-Based Agroforestry Practice during Summer Season in Charland and Plainland Ecosystems. J. Agrofor. Environ. 2018, 12, 1–8. [Google Scholar]
- Bellow, J.G.; Hudson, R.F.; Nair, P.K.R. Adoption Potential of Fruit-Tree-Based Agroforestry on Small Farms in the Subtropical Highlands. Agrofor. Syst. 2008, 73, 23–36. [Google Scholar] [CrossRef]
- Bellow, J.G.; Nair, P.K.R.; Martin, T.A. Tree–Crop Interactions in Fruit Tree-based Agroforestry Systems in the Western Highlands of Guatemala: Component Yields and System Performance. In Toward Agroforestry Design: An Ecological Approach; Jose, S., Gordon, A.M., Eds.; Advances in Agroforestry; Springer: Dordrecht, The Netherlands, 2008; pp. 111–131. ISBN 978-1-4020-6572-9. [Google Scholar]
- Do, V.H.; La, N.; Mulia, R.; Bergkvist, G.; Dahlin, A.S.; Nguyen, V.T.; Pham, H.T.; Öborn, I. Fruit Tree-Based Agroforestry Systems for Smallholder Farmers in Northwest Vietnam—A Quantitative and Qualitative Assessment. Land 2020, 9, 451. [Google Scholar] [CrossRef]
- Hossain, J.; Ahmed, T.; Hasnat, M.Z.; Karim, D. Screening of Tomato Varieties for Fruit Tree Based Agroforestry System. Int. J. Agric. Res. Innov. Technol. 2014, 4, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Kassa, G. Profitability Analysis and Determinants of Fruit Tree Based Agroforestry System in Wondo District, Ethiopia. Afr. J. Agric. Res. 2015, 10, 1273–1280. [Google Scholar] [CrossRef]
- Rahman, G.M.M.; Wadud, M.A.; Shahjahan, M.; Jewel, K.N.A. Fruit Tree Based Agroforestry Practices in Char Land Farming System. J. Agrofor. Environ 2014, 8, 1–6. [Google Scholar]
- Karki, U.; Goodman, M.S. Microclimatic Differences between Mature Loblolly-Pine Silvopasture and Open-Pasture. Agrofor. Syst. 2015, 89, 319–325. [Google Scholar] [CrossRef]
- Cannavo, P.; Sansoulet, J.; Harmand, J.-M.; Siles, P.; Dreyer, E.; Vaast, P. Agroforestry Associating Coffee and Inga densiflora Results in Complementarity for Water Uptake and Decreases Deep Drainage in Costa Rica. Agric. Ecosyst. Environ. 2011, 140, 1–13. [Google Scholar] [CrossRef]
- Lin, B.B. The Role of Agroforestry in Reducing Water Loss through Soil Evaporation and Crop Transpiration in Coffee Agroecosystems. Agric. For. Meteorol. 2010, 150, 510–518. [Google Scholar] [CrossRef]
- Marrou, H.; Wery, J.; Dufour, L.; Dupraz, C. Productivity and Radiation Use Efficiency of Lettuces Grown in the Partial Shade of Photovoltaic Panels. Eur. J. Agron. 2013, 44, 54–66. [Google Scholar] [CrossRef]
- Niether, W.; Schneidewind, U.; Armengot, L.; Adamtey, N.; Schneider, M.; Gerold, G. Spatial-Temporal Soil Moisture Dynamics under Different Cocoa Production Systems. CATENA 2017, 158, 340–349. [Google Scholar] [CrossRef]
- McClaugherty, C.A.; Aber, J.D.; Melillo, J.M. The Role of Fine Roots in the Organic Matter and Nitrogen Budgets of Two Forested Ecosystems. Ecology 1982, 63, 1481–1490. [Google Scholar] [CrossRef] [Green Version]
- Muchane, M.N.; Sileshi, G.W.; Gripenberg, S.; Jonsson, M.; Pumariño, L.; Barrios, E. Agroforestry Boosts Soil Health in the Humid and Sub-Humid Tropics: A Meta-Analysis. Agric. Ecosyst. Environ. 2020, 295, 106899. [Google Scholar] [CrossRef]
- Ruess, R.W.; Cleve, K.V.; Yarie, J.; Viereck, L.A. Contributions of Fine Root Production and Turnover to the Carbon and Nitrogen Cycling in Taiga Forests of the Alaskan Interior. Can. J. For. Res. 1996, 26, 1326–1336. [Google Scholar] [CrossRef]
- Leigh, M.B.; Fletcher, J.S.; Fu, X.; Schmitz, F.J. Root Turnover: An Important Source of Microbial Substrates in Rhizosphere Remediation of Recalcitrant Contaminants. Environ. Sci. Technol. 2002, 36, 1579–1583. [Google Scholar] [CrossRef]
- Sauvadet, M.; Saj, S.; Freschet, G.T.; Essobo, J.-D.; Enock, S.; Becquer, T.; Tixier, P.; Harmand, J.-M. Cocoa Agroforest Multifunctionality and Soil Fertility Explained by Shade Tree Litter Traits. J. Appl. Ecol. 2020, 57, 476–487. [Google Scholar] [CrossRef]
- Cherubin, M.R.; Chavarro-Bermeo, J.P.; Silva-Olaya, A.M. Agroforestry Systems Improve Soil Physical Quality in Northwestern Colombian Amazon. Agrofor. Syst. 2019, 93, 1741–1753. [Google Scholar] [CrossRef]
- Schroth, G. A review of belowground interactions in agroforestry, focussing on mechanisms and management options. In Agroforestry for Sustainable Land-Use Fundamental Research and Modelling with Emphasis on Temperate and Mediterranean Applications; Auclair, D., Dupraz, C., Eds.; Springer: Dordrecht, The Netherlands, 1999; Volume 60, pp. 5–34. ISBN 978-90-481-5258-2. [Google Scholar]
- Seyfried, M.S.; Rao, P.S.C. Nutrient Leaching Loss from Two Contrasting Cropping Systems in the Humid Tropics. Trop. Agric. 1990, 68, 9–18. [Google Scholar]
- Gao, X.; Liu, Z.; Zhao, X.; Ling, Q.; Huo, G.; Wu, P. Extreme Natural Drought Enhances Interspecific Facilitation in Semiarid Agroforestry Systems. Agric. Ecosyst. Environ. 2018, 265, 444–453. [Google Scholar] [CrossRef]
- Li, F.; Meng, P.; Fu, D.; Wang, B. Light Distribution, Photosynthetic Rate and Yield in a Paulownia-Wheat Intercropping System in China. Agrofor. Syst. 2008, 74, 163–172. [Google Scholar] [CrossRef]
- Guo, Z.L.; Zhong, C.; Cai, C.F.; Ding, S.W.; Wang, Z.M. Nitrogen Competition in Contour Hedgerow Systems in Subtropical China. Nutr. Cycl. Agroecosyst. 2008, 81, 71–83. [Google Scholar] [CrossRef]
- Livesley, S.J.; Gregory, P.J.; Buresh, R.J. Competition in Tree Row Agroforestry Systems. 2. Distribution, Dynamics and Uptake of Soil Inorganic N. Plant Soil 2002, 247, 177–187. [Google Scholar] [CrossRef]
- Prasad, J.V.N.S.; Korwar, G.R.; Rao, K.V.; Mandal, U.K.; Rao, C.A.R.; Rao, G.R.; Ramakrishna, Y.S.; Venkateswarlu, B.; Rao, S.N.; Kulkarni, H.D.; et al. Tree Row Spacing Affected Agronomic and Economic Performance of Eucalyptus-Based Agroforestry in Andhra Pradesh, Southern India. Agrofor. Syst. 2010, 78, 253–267. [Google Scholar] [CrossRef]
- Kohli, A.; Saini, B.C. Microclimate Modification and Response of Wheat Planted under Trees in a Fan Design in Northern India. Agrofor. Syst. 2003, 58, 109–117. [Google Scholar] [CrossRef]
- Artru, S.; Garré, S.; Dupraz, C.; Hiel, M.-P.; Blitz-Frayret, C.; Lassois, L. Impact of Spatio-Temporal Shade Dynamics on Wheat Growth and Yield, Perspectives for Temperate Agroforestry. Eur. J. Agron. 2017, 82, 60–70. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources, 2006: A Framework for International Classification, Correlation, and Communication; FAO, Ed.; World soil resources reports; 2006 (2nd ed.); Food and Agriculture Organization of the United Nations: Rome, Italy, 2006; ISBN 978-92-5-105511-3. [Google Scholar]
- De Boodt, M.; Verdonck, O.; Cappaert, I. Method for Measuring the Water Release Curve of Organic Substrates. Acta Hortic. 1974, 37, 2054–2063. [Google Scholar] [CrossRef]
- Richards, L.A. Pressure-Membrane Apparatus, Construction and Use. Agric. Eng. 1947, 28, 451–454. [Google Scholar]
- van Genuchten, M.T. A Closed-Form Equation for Predicting the Hydraulic Conductivity of Unsaturated Soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Raison, R.J.; Connell, M.J.; Khanna, P.K. Methodology for Studying Fluxes of Soil Mineral-N in Situ. Soil Biol. Biochem. 1987, 19, 521–530. [Google Scholar] [CrossRef]
- Vendrell, P.F.; Zupancic, J. Determination of Soil Nitrate by Transnitration of Salicylic Acid. Commun. Soil Sci. Plant Anal. 1990, 21, 1705–1713. [Google Scholar] [CrossRef]
- Nelson, D.W. Determination of Ammonium in KCl Extracts of Soils by the Salicylate Method. Commun. Soil Sci. Plant Anal. 1983, 14, 1051–1062. [Google Scholar] [CrossRef]
- Pau, G.; Fuchs, F.; Sklyar, O.; Boutros, M.; Huber, W. EBImage-an R Package for Image Processing with Applications to Cellular Phenotypes. Bioinformatics 2010, 26, 979–981. [Google Scholar] [CrossRef]
- McMaster, G.S.; Wilhelm, W.W. Growing Degree-Days: One Equation, Two Interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Maynard, D.N.; Hochmuth, G.J.; Knott, J.E. Knott’s Handbook for Vegetable Growers, 5th ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; ISBN 978-0-471-73828-2. [Google Scholar]
- Borrelli, K.; Koenig, R.T.; Jaeckel, B.M.; Miles, C.A. Yield of Leafy Greens in High Tunnel Winter Production in the Northwest United States. HortScience 2013, 48, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Soft. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; Fao: Rome, Italy, 1998; 300p, ISBN 978-92-5-104219-9. [Google Scholar]
- Kaufman, L.; Rousseeuw, P.J. Finding Groups in Data: An Introduction to Cluster Analysis; Applied Probability and Statistics, New York, Wiley Series in Probability and Mathematical Statistics; John Wiley & Sons, Ltd.: Chichester, UK, 1990. [Google Scholar]
- Dahl, D.B.; Scott, D.; Roosen, C.; Magnusson, A.; Swinton, J. Xtable: Export Tables to LaTeX or HTML. R package version, 2009, pp. 1–6. Available online: http://cran.pau.edu.tr/web/packages/xtable/xtable.pdf (accessed on 3 April 2021).
- R Studio Team RStudio: Integrated Development for R. RStudio 2020. Available online: http://www.rstudio.com/ (accessed on 8 June 2020).
- Monteith, J.L. Light Interception and Radiative Exchange in Crop Stands. In Physiological Aspects of Crop Yield; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1969; pp. 89–111. ISBN 978-0-89118-571-0. [Google Scholar]
- Palmer, J.W. Diurnal Light Interception and a Computer Model of Light Interception by Hedgerow Apple Orchards. J. Appl. Ecol. 1977, 14, 601–614. [Google Scholar] [CrossRef]
- Renaud, V.; Rebetez, M. Comparison between Open-Site and below-Canopy Climatic Conditions in Switzerland during the Exceptionally Hot Summer of 2003. Agric. For. Meteorol. 2009, 149, 873–880. [Google Scholar] [CrossRef]
- Breshears, D.D.; Rich, P.M.; Barnes, F.J.; Campbell, K. Overstory-Imposed Heterogeneity in Solar Radiation and Soil Moisture in a Semiarid Woodland. Ecol. Appl. 1997, 7, 1201–1215. [Google Scholar] [CrossRef]
- Monteith, J.L.; Ong, C.K.; Corlett, J.E. Microclimatic Interactions in Agroforestry Systems. For. Ecol. Manag. 1991, 45, 31–44. [Google Scholar] [CrossRef]
- Rodrigues, J.C.; Miranda, I.S.; de Sousa, A.M.L.; Rodrigues, J.C.; Miranda, I.S.; de Sousa, A.M.L. Microclimate in Understories of a Mango Orchard and a Degraded Area in the Eastern Amazon. Rev. Ambiente Água 2018, 13, e2129. [Google Scholar] [CrossRef]
- Beer, J.; Muschler, R.; Kass, D.; Somarriba, E. Shade Management in Coffee and Cacao Plantations. Agrofor. Syst. 1997, 38, 139–164. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, P.; Hu, Y.; Ouyang, L.; Zhu, L.; Ni, G. Canopy Transpiration and Its Cooling Effect of Three Urban Tree Species in a Subtropical City- Guangzhou, China. Urban For. Urban Green. 2019, 43, 126368. [Google Scholar] [CrossRef]
- Meng, L.; Yu, L.; Mao, P.; Zhang, G. Effects of Inter-Planting Cocksfoot and White Clover as Cover Crops on the Microclimate of Apple Orchard. Pratacultural Sci. 2009, 26, 132–136. [Google Scholar]
- Taha, H.; Akbari, H.; Rosenfeld, A. Heat Island and Oasis Effects of Vegetative Canopies: Micro-Meteorological Field-Measurements. Theor. Appl. Climatol. 1991, 44, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Taha, H.; Akbari, H.; Rosenfeld, A. Vegetation Canopy Micro-Climate: A Field-Project in Davis, California. 1989. Available online: https://escholarship.org/uc/item/7td0c626 (accessed on 16 September 2021).
- von Arx, G.; Dobbertin, M.; Rebetez, M. Spatio-Temporal Effects of Forest Canopy on Understory Microclimate in a Long-Term Experiment in Switzerland. Agric. For. Meteorol. 2012, 166–167, 144–155. [Google Scholar] [CrossRef]
- Souch, C.A.; Souch, C. The Effect of Trees on Summertime below Canopy Urban Climates: A Case Study Bloomington, Indiana. J. Arboric. 1993, 19, 303–312. [Google Scholar]
- Dupont, S.; Patton, E.G. Influence of Stability and Seasonal Canopy Changes on Micrometeorology within and above an Orchard Canopy: The CHATS Experiment. Agric. For. Meteorol. 2012, 157, 11–29. [Google Scholar] [CrossRef]
- Dupraz, C.; Liagre, F. Agroforesterie: Des Arbres et des Cultures; France Agricole Editions: Paris, France, 2008; ISBN 978-2-85557-150-8. [Google Scholar]
- Godefroid, S.; Koedam, N. Tree-Induced Soil Compaction in Forest Ecosystems: Myth or Reality? Eur. J. For. Res. 2010, 129, 209–217. [Google Scholar] [CrossRef]
- Eastham, J.; Rose, C.W. Pasture Evapotranspiration under Varying Tree Planting Density in an Agroforestry Experiment. Agric. Water Manag. 1988, 15, 87–105. [Google Scholar] [CrossRef]
- Jackson, N.A.; Wallace, J.S. Soil Evaporation Measurements in an Agroforestry System in Kenya. Agric. For. Meteorol. 1999, 94, 203–215. [Google Scholar] [CrossRef]
- Raz-Yaseef, N.; Rotenberg, E.; Yakir, D. Effects of Spatial Variations in Soil Evaporation Caused by Tree Shading on Water Flux Partitioning in a Semi-Arid Pine Forest. Agric. For. Meteorol. 2010, 150, 454–462. [Google Scholar] [CrossRef]
- Kormanek, M.; Głąb, T.; Banach, J.; Szewczyk, G. Effects of Soil Bulk Density on Sessile Oak Quercus petraea Liebl. Seedlings. Eur. J. For. Res. 2015, 134, 969–979. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Kukal, S.S.; Bawa, S.S.; Dhaliwal, G.S. Soil Organic Carbon and Aggregation under Poplar Based Agroforestry System in Relation to Tree Age and Soil Type. Agrofor. Syst. 2009, 76, 27–35. [Google Scholar] [CrossRef]
- Pardon, P.; Reubens, B.; Reheul, D.; Mertens, J.; De Frenne, P.; Coussement, T.; Janssens, P.; Verheyen, K. Trees Increase Soil Organic Carbon and Nutrient Availability in Temperate Agroforestry Systems. Agric. Ecosyst. Environ. 2017, 247, 98–111. [Google Scholar] [CrossRef]
- Upson, M.A.; Burgess, P.J. Soil Organic Carbon and Root Distribution in a Temperate Arable Agroforestry System. Plant Soil 2013, 373, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Cardinael, R.; Chevallier, T.; Cambou, A.; Béral, C.; Barthès, B.G.; Dupraz, C.; Durand, C.; Kouakoua, E.; Chenu, C. Increased Soil Organic Carbon Stocks under Agroforestry: A Survey of Six Different Sites in France. Agric. Ecosyst. Environ. 2017, 236, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Hendricks, J.J.; Nadelhoffer, K.J.; Aber, J.D. Assessing the Role of Fine Roots in Carbon and Nutrient Cycling. Trends Ecol. Evol. 1993, 8, 174–178. [Google Scholar] [CrossRef]
- Arvidsson, J. Influence of Soil Texture and Organic Matter Content on Bulk Density, Air Content, Compression Index and Crop Yield in Field and Laboratory Compression Experiments. Soil Tillage Res. 1998, 49, 159–170. [Google Scholar] [CrossRef]
- Logsdon, S.D.; Karlen, D.L. Bulk Density as a Soil Quality Indicator during Conversion to No-Tillage. Soil Tillage Res. 2004, 78, 143–149. [Google Scholar] [CrossRef]
- Pepper, I.L.; Brooks, J.P.; Gerba, C.P. Chapter 23—Land Application of Organic Residuals: Municipal Biosolids and Animal Manures. In Environmental and Pollution Science, 3rd ed.; Brusseau, M.L., Pepper, I.L., Gerba, C.P., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 419–434. ISBN 978-0-12-814719-1. [Google Scholar]
- Li, Y.; Liu, Y.; Wang, Y.; Niu, L.; Xu, X.; Tian, Y. Interactive Effects of Soil Temperature and Moisture on Soil N Mineralization in a Stipa Krylovii Grassland in Inner Mongolia, China. J. Arid Land 2014, 6, 571–580. [Google Scholar] [CrossRef]
- Guntiñas, M.E.; Leirós, M.C.; Trasar-Cepeda, C.; Gil-Sotres, F. Effects of Moisture and Temperature on Net Soil Nitrogen Mineralization: A Laboratory Study. Eur. J. Soil Biol. 2012, 48, 73–80. [Google Scholar] [CrossRef]
- Curtin, D.; Beare, M.H.; Hernandez-Ramirez, G. Temperature and Moisture Effects on Microbial Biomass and Soil Organic Matter Mineralization. Soil Sci. Soc. Am. J. 2012, 76, 2055–2067. [Google Scholar] [CrossRef]
- Hai, L.; Li, X.G.; Liu, X.-E.; Jiang, X.J.; Guo, R.Y.; Jing, G.B.; Rengel, Z.; Li, F.-M. Plastic Mulch Stimulates Nitrogen Mineralization in Urea-Amended Soils in a Semiarid Environment. Agron. J. 2015, 107, 921–930. [Google Scholar] [CrossRef]
- Guillot, E.; Bertrand, I.; Rumpel, C.; Gomez, C.; Arnal, D.; Abadie, J.; Hinsinger, P. Spatial Heterogeneity of Soil Quality within a Mediterranean Alley Cropping Agroforestry System: Comparison with a Monocropping System. Eur. J. Soil Biol. 2021, 105, 103330. [Google Scholar] [CrossRef]
- Yao, S.; Merwin, I.A.; Bird, G.W.; Abawi, G.S.; Thies, J.E. Orchard Floor Management Practices That Maintain Vegetative or Biomass Groundcover Stimulate Soil Microbial Activity and Alter Soil Microbial Community Composition. Plant Soil 2005, 271, 377–389. [Google Scholar] [CrossRef]
- Meier, I.C.; Finzi, A.C.; Phillips, R.P. Root Exudates Increase N Availability by Stimulating Microbial Turnover of Fast-Cycling N Pools. Soil Biol. Biochem. 2017, 106, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.; Marani, A.; Rudich, J. Effects of Temperature and Photoperiod on Assimilate Partitioning in Potato Plants. Ann. Bot. 1990, 66, 513–520. [Google Scholar] [CrossRef]
- Brouwer, R. Some Aspects of the Equilibrium between Overground and Underground Plant Parts. Jaarboek van het Instituut voor Biologisch en Scheikundig onderzoek aan Landbouwgewassen 1963, 1963, 31–39. Available online: https://edepot.wur.nl/361707#page=27 (accessed on 16 September 2021).
- Sage, R.F.; Sharkey, T.D.; Pearcy, R.W. The Effect of Leaf Nitrogen and Temperature on the CO2 Response of Photosynthesis in the C3 Dicot MChenopodium album L. Funct. Plant Biol. 1990, 17, 135–148. [Google Scholar] [CrossRef]
- Driesen, E.; Van den Ende, W.; De Proft, M.; Saeys, W. Influence of Environmental Factors Light, CO2, Temperature, and Relative Humidity on Stomatal Opening and Development: A Review. Agronomy 2020, 10, 1975. [Google Scholar] [CrossRef]
- Oh, S.; Moon, K.H.; Song, E.Y.; Son, I.-C.; Koh, S.C. Photosynthesis of Chinese Cabbage and Radish in Response to Rising Leaf Temperature during Spring. Hortic. Environ. Biotechnol. 2015, 56, 159–166. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.Z.; Hang, T.; Li, P.P. Photosynthetic Characteristics and Growth Performance of Lettuce (Lactuca sativa L.) under Different Light/Dark Cycles in Mini Plant Factories. Photosynthetica 2020, 58, 740–747. [Google Scholar] [CrossRef] [Green Version]
- Cometti, N.N.; da Silva, J.V.; Zonta, E.; Cessa, R.M. Evaluation of Photosynthetic Photon Flux in Lettuce Cultivation at Different Shading Levels. Hortic. Bras. 2020, 38, 65–70. [Google Scholar] [CrossRef]
- Wheeler, T.R.; Hadley, P.; Ellis, R.H.; Morison, J.I.L. Changes in Growth and Radiation Use by Lettuce Crops in Relation to Temperature and Ontogeny. Agric. For. Meteorol. 1993, 66, 173–186. [Google Scholar] [CrossRef]
- Jie, H.; Sing Kong, L. Growth and Photosynthetic Characteristics of Lettuce (Lactuca sativa L.) under Fluctuating Hot Ambient Temperatures with the Manipulation of Cool Root-Zone Temperature. J. Plant Physiol. 1998, 152, 387–391. [Google Scholar] [CrossRef]
- Li, R.; YueLin, Z.; Takagaki, M.; Yamori, W.; LiFei, Y. Effects of root zone temperature on the growth and mineral elements content of hydroponically-grown lettuce. Acta Agric. Shanghai 2015, 31, 48–52. [Google Scholar]
- Ouyang, Z.; Tian, J.; Yan, X.; Shen, H. Effects of Different Concentrations of Dissolved Oxygen or Temperatures on the Growth, Photosynthesis, Yield and Quality of Lettuce. Agric. Water Manag. 2020, 228, 105896. [Google Scholar] [CrossRef]
- Khan, M. Effect of High Temperature and Exposure Duration on Stem Elongation of Iceberg Lettuce. Pak. J. Agric. Sci. 2018, 55, 95–101. [Google Scholar] [CrossRef]
- Arvidsson, J.; Håkansson, I. Response of Different Crops to Soil Compaction—Short-Term Effects in Swedish Field Experiments. Soil Tillage Res. 2014, 138, 56–63. [Google Scholar] [CrossRef]
- Adekiya, A.O.; Agbede, T.M.; Aboyeji, C.M.; Dunsin, O.; Simeon, V.T. Effects of Biochar and Poultry Manure on Soil Characteristics and the Yield of Radish. Sci. Hortic. 2019, 243, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Sojka, R.E.; Busscher, W.J.; Lehrsch, G.A. In situ strength, bulk density, and water content relationships of a durinodic xeric haplocalcid soil. Soil Sci. 2001, 166, 520–529. [Google Scholar] [CrossRef] [Green Version]
- Azzi, V.; Kanso, A.; Kazpard, V.; Kobeissi, A.; Lartiges, B.; El Samrani, A. Lactuca Sativa Growth in Compacted and Non-Compacted Semi-Arid Alkaline Soil under Phosphate Fertilizer Treatment and Cadmium Contamination. Soil Tillage Res. 2017, 165, 1–10. [Google Scholar] [CrossRef]
- Baloch, P.A.; Uddin, R.; Nizamani, F.K.; Solangi, A.H.; Siddiqui, A.A. Effect of Nitrogen, Phosphorus and Potassium Fertilizers on Growth and Yield Characteristics of Radish (Raphinus sativus L.). Am.-Eurasian J. Agric. Environ. Sci. 2014, 14, 565–569. [Google Scholar]
- Kang, Y.; Wan, S. Effect of Soil Water Potential on Radish (Raphanus sativus L.) Growth and Water Use under Drip Irrigation. Sci. Hortic. 2005, 106, 275–292. [Google Scholar] [CrossRef]
- Dupraz, C.; Wolz, K.J.; Lecomte, I.; Talbot, G.; Vincent, G.; Mulia, R.; Bussière, F.; Ozier-Lafontaine, H.; Andrianarisoa, S.; Jackson, N.; et al. Hi-SAFe: A 3D Agroforestry Model for Integrating Dynamic Tree–Crop Interactions. Sustainability 2019, 11, 2293. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Layer | Ag | Eg | BTg | Cg |
|---|---|---|---|---|
| (0–30 cm) | (30–50 cm) | (50–90 cm) | (90–120 cm) | |
| Texture | ||||
| Clay [%] | 15.5 | 22 | 36.5 | 31.3 |
| Silt [%] | 42.2 | 39.6 | 31.7 | 28.3 |
| Sand [%] | 39.6 | 37.8 | 31.1 | 39.7 |
| Chemical properties | ||||
| pHH2O | 6.7 | 7.2 | 7.5 | 5.1 |
| pHKCl | 5.9 | 6.3 | 6.6 | 3.8 |
| Organic matter [g.kg−1] | 35 | 6 | <6 | <6 |
| Total C [g.kg−1] | 20.3 | 3.5 | <3 | <3 |
| Total N [g.kg−1] | 1.3 | 0.5 | 0.4 | 0.3 |
| C:N ratio | 15.6 | 7.4 | <7.4 | <10.8 |
| P2O5 [mg.kg−1] | 68 | 16 | 10 | <10 |
| K2O [mg.kg−1] | 258 | 140 | 111 | 71 |
| MgO [mg.kg−1] | 165 | 157 | 168 | 125 |
| Na2O [mg.kg−1] | 14 | 12 | 27 | 21 |
| CaO [mg.kg−1] | 1757 | 1324 | 2528 | 1149 |
| CaCO3 [g.kg−1] | <1 | <1 | < 1 | <1 |
| CEC [meq.100 g−1] | 7.5 | 6.3 | 7.7 | 7.2 |
| Hydraulic properties (mean ± SD, n = 3) | ||||
| Bulk density [g.cm−3] | 1.37 ± 0.11 | 1.65 ± 0.06 | 1.55 ± 0.07 | 1.61 ± 0.06 |
| Field capacity [% volumetric water content] | 24.4 ± 1.4 | 28.7 ± 4.3 | 30.1 ± 6.3 | 29.1 ± 5.1 |
| Wilting point [% volumetric water content] | 17.7 ± 0.6 | 23.2 ± 3.7 | 24.4 ± 7.2 | 22.4 ± 4.6 |
| Saturated hydraulic conductivity [10−4 cm.s−1] | 4.33 ± 0.47 | 1.23 ± 0.13 | 3.03 ± 0.50 | 2.33 ± 0.13 |
| Bed | Radish | Lettuce | ||||||
|---|---|---|---|---|---|---|---|---|
| Bulk Density [g DW.cm−3] | SOM [g.kg−1 DW] | Bulk Density [g DW.cm−3] | SOM [g.kg−1 DW] | |||||
| B1E | 1.26 ± 0.07 | b | 37.36 ± 1.11 | b | 1.28 ± 0.08 | b | 37.76 ± 1.23 | ab |
| B2E | 1.45 ± 0.05 | a | 32.76 ± 1.86 | b | 1.46 ± 0.05 | a | 33.46 ± 1.14 | c |
| C | 1.46 ± 0.06 | a | 33.05 ± 2.27 | b | 1.51 ± 0.03 | a | 31.42 ± 2.80 | c |
| B2W | 1.46 ± 0.04 | a | 33.39 ± 3.67 | b | 1.49 ± 0.02 | a | 33.51 ± 1.29 | bc |
| B1W | 1.23 ± 0.02 | b | 46.46 ± 2.34 | a | 1.31 ± 0.05 | b | 41.55 ± 0.53 | a |
| Final DM | Final LA | Cumulated GR | Cumulated GDD | Bulk Density | SOM | ||
|---|---|---|---|---|---|---|---|
| Radish | Final LA | 0.77 ** | — | ||||
| Cumulated GR | −0.18 | −0.28 | — | ||||
| Cumulated GDD | −0.62 | −0.27 | 0.41 | — | |||
| Bulk density | −0.86 *** | −0.81 ** | 0.45 | 0.65 | — | ||
| SOM | 0.71 ** | 0.54 * | −0.60 * | −0.56 | −0.77 ** | — | |
| N mineralization | 0.63 * | 0.60 * | −0.61 * | −0.59 | −0.76 ** | 0.82 ** | |
| Lettuce | Final LA | 0.88 *** | — | ||||
| Cumulated GR | 0.88 *** | 0.74 ** | — | ||||
| Cumulated GDD | 0.61 | 0.59 | 0.64 | — | |||
| Bulk density | 0.48 | 0.25 | 0.51 | 0.66 | — | ||
| SOM | −0.53 * | −0.38 | −0.55 * | −0.72 * | −0.82 ** | — | |
| N mineralization | −0.28 | −0.08 | −0.37 | −0.07 | −0.55 ** | 0.59 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramananjatovo, T.; Chantoiseau, E.; Guillermin, P.; Guénon, R.; Delaire, M.; Buck-Sorlin, G.; Cannavo, P. Growth of Vegetables in an Agroecological Garden-Orchard System: The Role of Spatiotemporal Variations of Microclimatic Conditions and Soil Properties. Agronomy 2021, 11, 1888. https://doi.org/10.3390/agronomy11091888
Ramananjatovo T, Chantoiseau E, Guillermin P, Guénon R, Delaire M, Buck-Sorlin G, Cannavo P. Growth of Vegetables in an Agroecological Garden-Orchard System: The Role of Spatiotemporal Variations of Microclimatic Conditions and Soil Properties. Agronomy. 2021; 11(9):1888. https://doi.org/10.3390/agronomy11091888
Chicago/Turabian StyleRamananjatovo, Toky, Etienne Chantoiseau, Pascale Guillermin, René Guénon, Mickaël Delaire, Gerhard Buck-Sorlin, and Patrice Cannavo. 2021. "Growth of Vegetables in an Agroecological Garden-Orchard System: The Role of Spatiotemporal Variations of Microclimatic Conditions and Soil Properties" Agronomy 11, no. 9: 1888. https://doi.org/10.3390/agronomy11091888