
Spatial Delimitation of Genetic Diversity of Native Maize and Its Relationship with Ethnic Groups in Mexico

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Databases
2.2. Spatial Delimitation of the Regions of Genetic Richness per Maize Race
2.3. Spatial Delimitation of Intraspecific Maize Diversity
2.4. Spatial Delimitation of the Maize Diversity Center
2.5. Characterization of the Maize Diversity Center by Climate Factors
2.6. Relationship of Environmental Units with Ethnic Groups and Maize Uses
3. Results
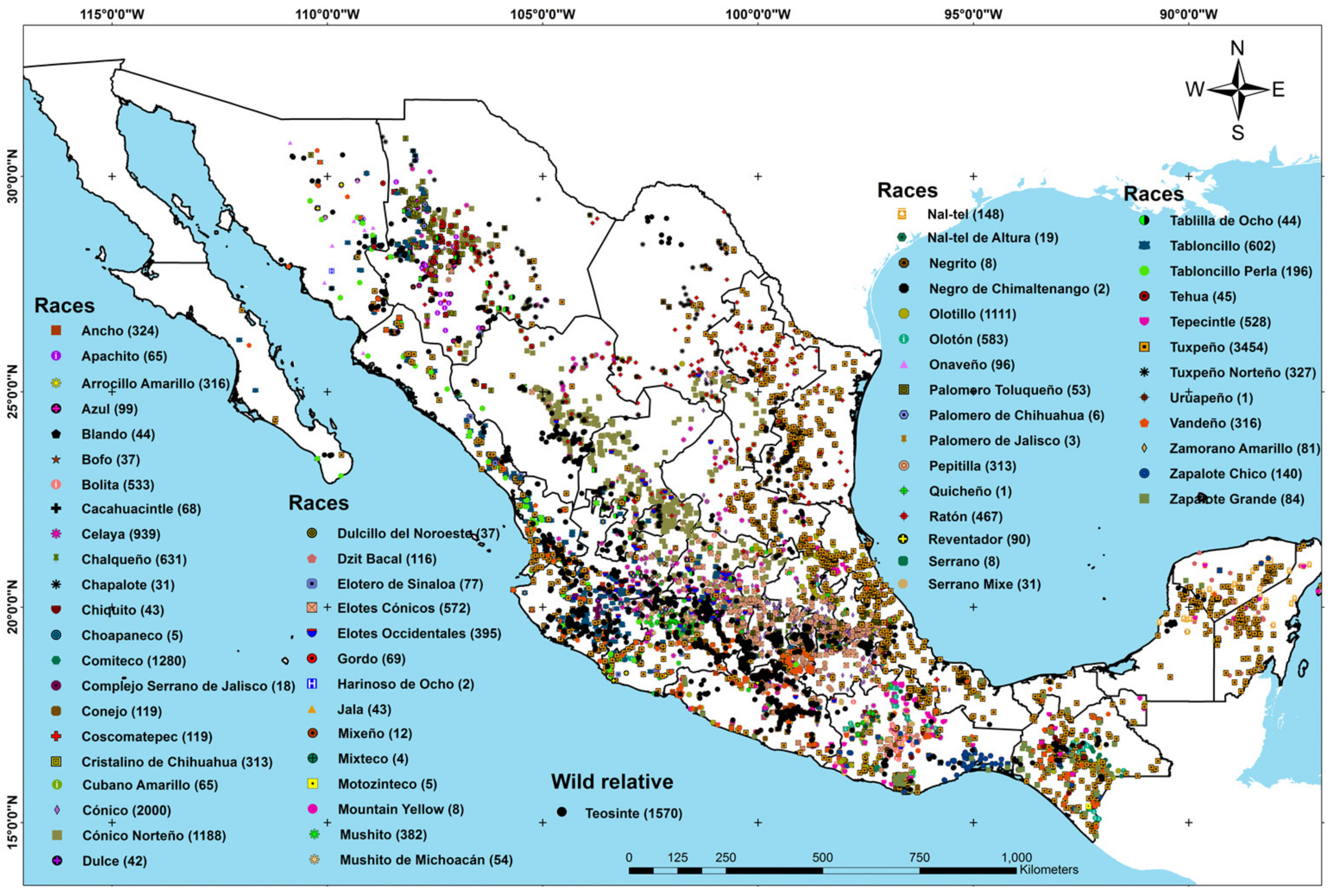
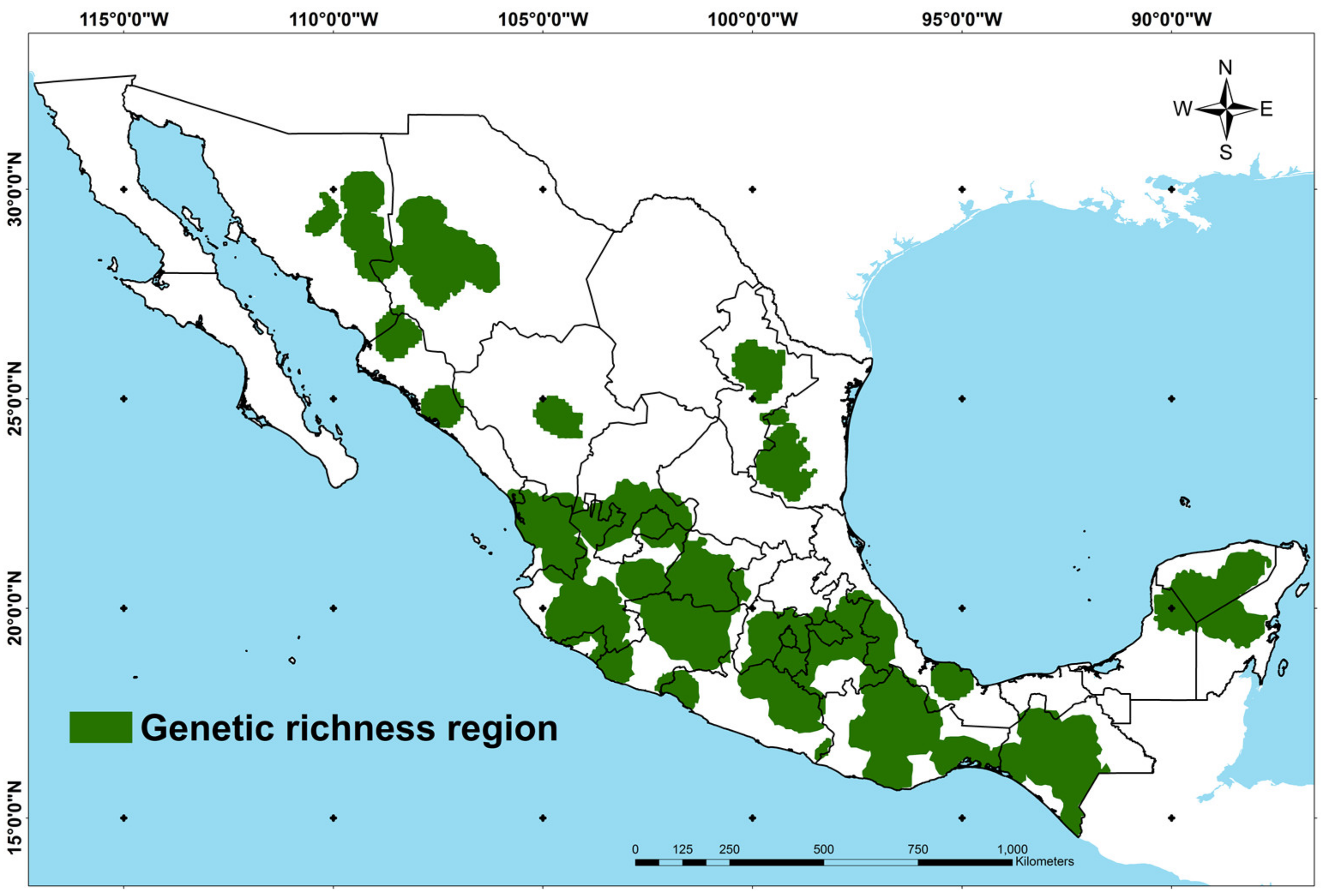
3.1. Spatial Delimitation of the Genetic Richness Regions per Maize Race
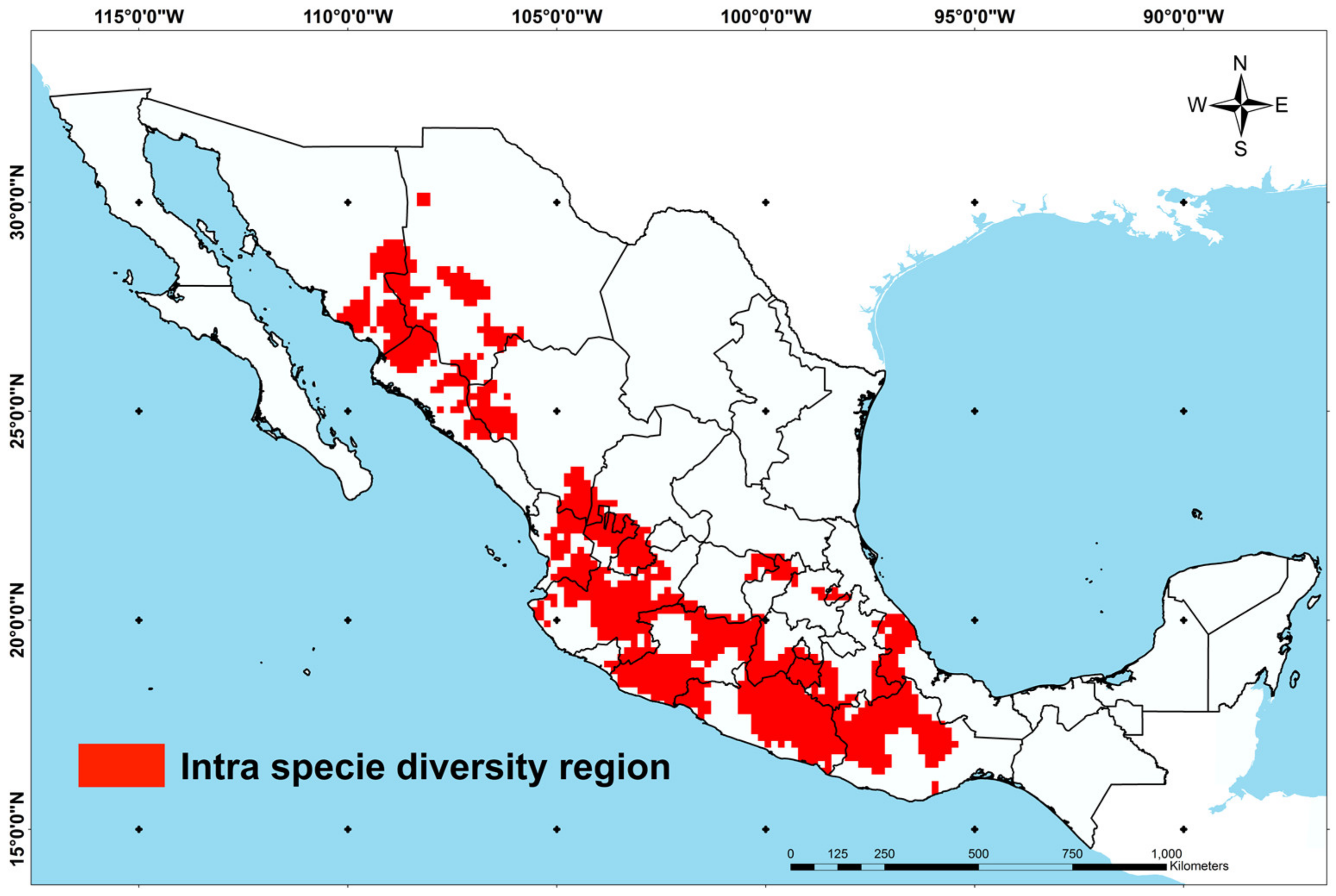
3.2. Spatial Delimitation of Intraspecific Maize Diversity
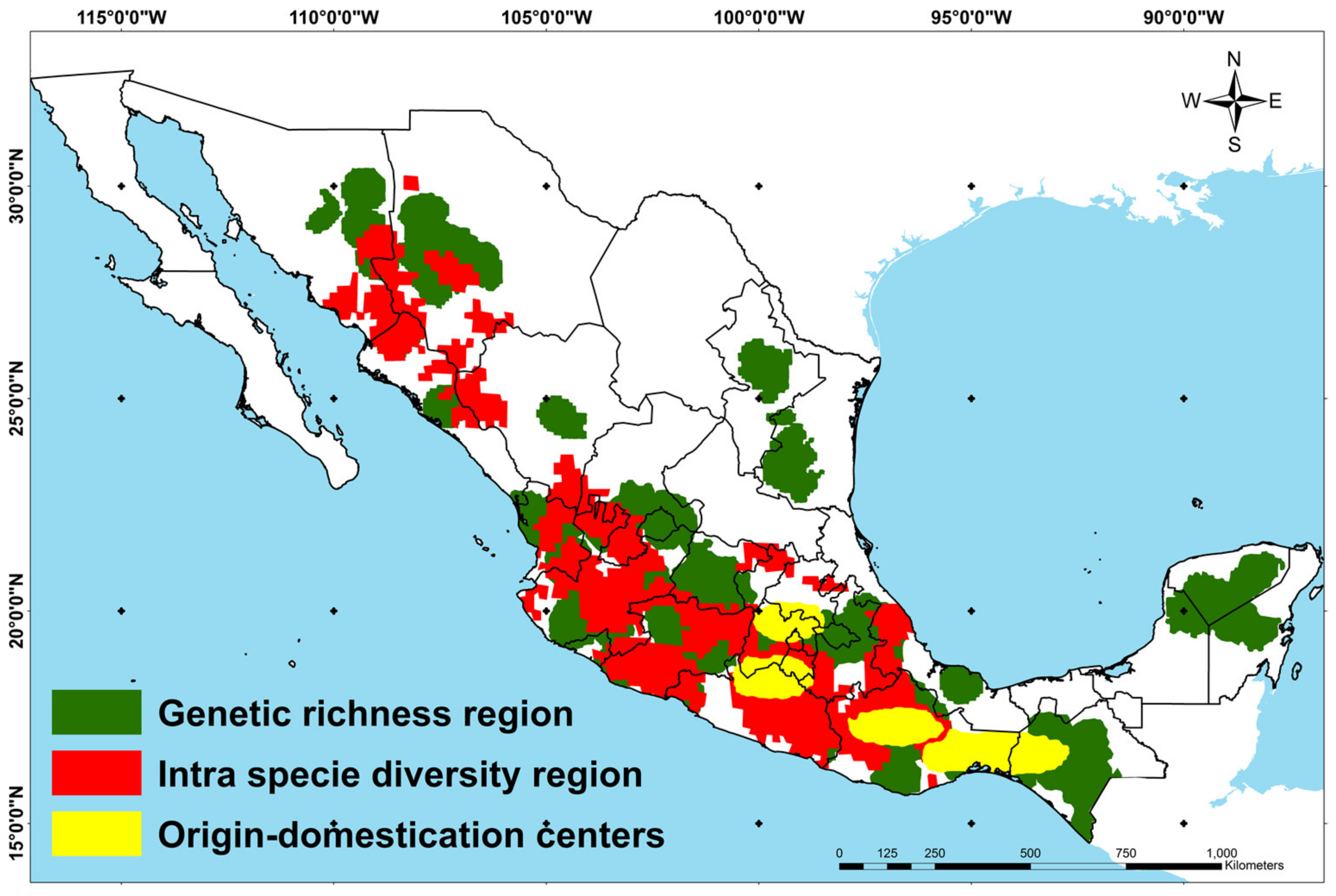
3.3. Spatial Delimitation of the Maize Diversity Center
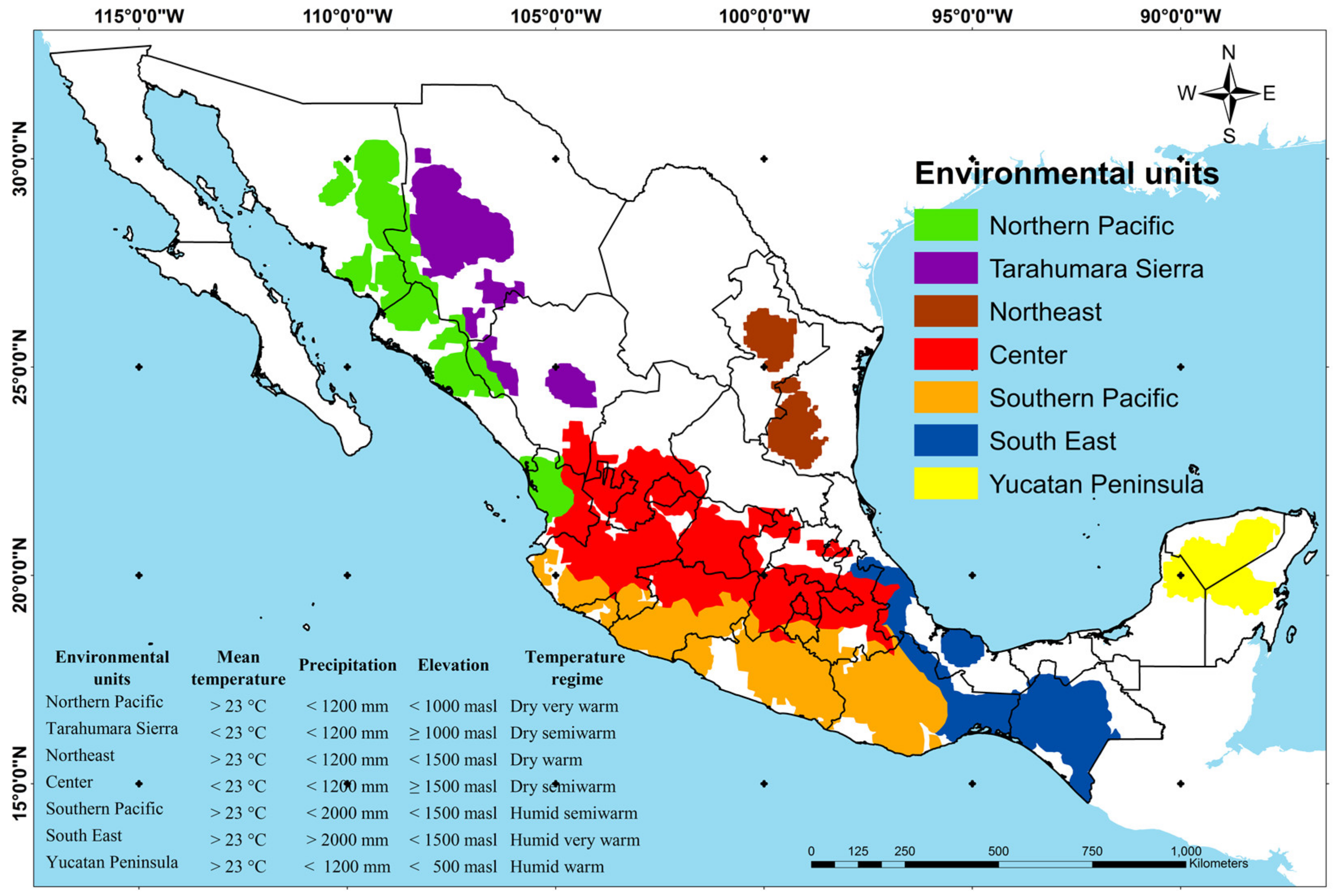
3.4. Characterization of the Maize Diversity Center by Climatic Factors
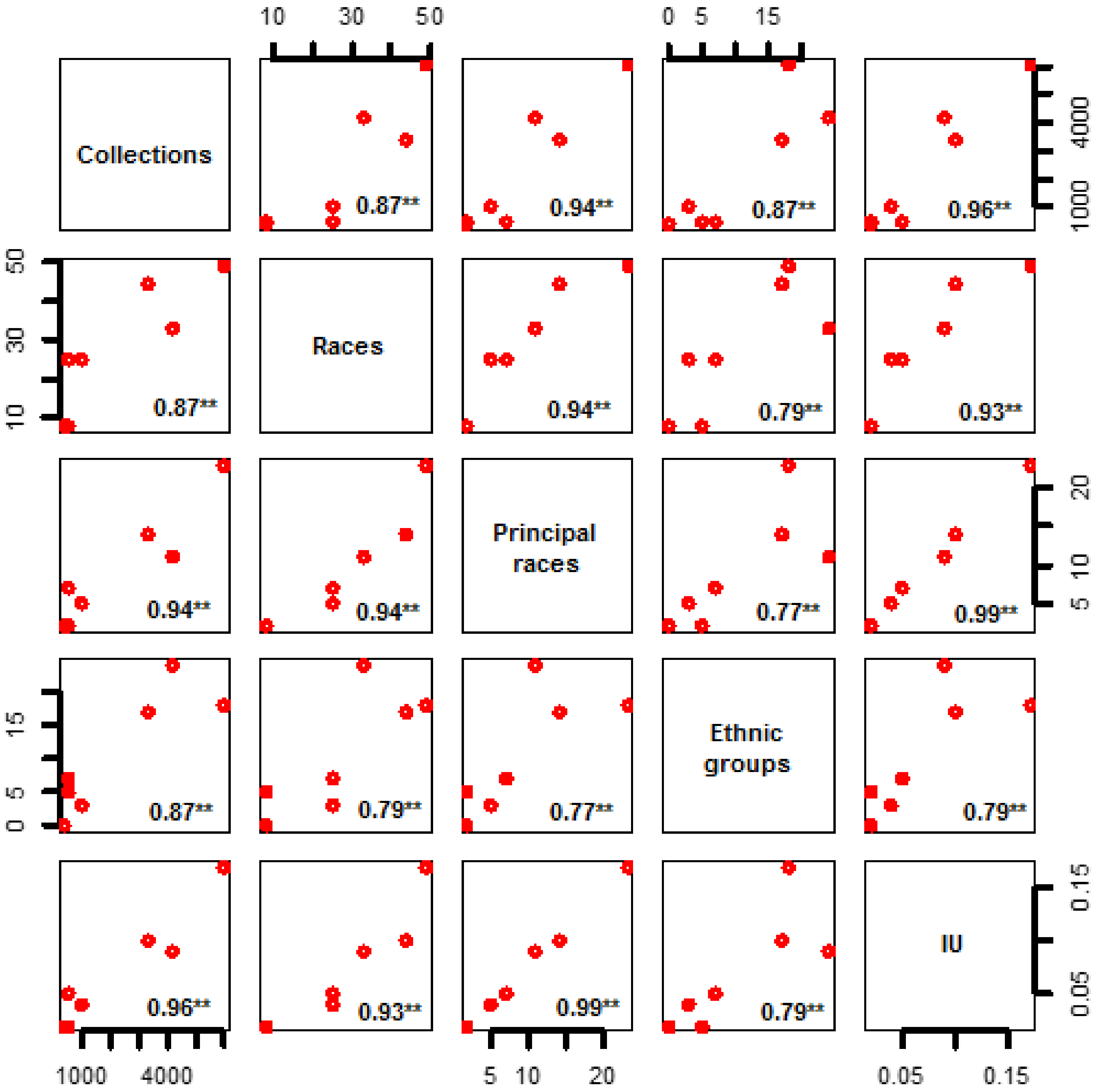
3.5. Relationship of Environmental Units with Ethnic Groups and Maize Uses
4. Discussion
4.1. Spatial Delimitation of the Genetic Richness Regions per Maize Race
4.2. Environmental Units
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruíz-Corral, J.A.; Durán-Puga, N.; Sánchez-González, J.J.; Rón-Parra, J.; González-Eguiarte, D.R.; Holland, J.B.; Medina-García, G. Climatic adaptation and ecological descriptors of 42 Mexican maize races. Crop Sci. 2008, 48, 1502–1512. [Google Scholar] [CrossRef] [Green Version]
- Dyer, G.A.; Taylor, J.E. A crop population perspective on maize seed systems in Mexico. Proc. Natl. Acad. Sci. USA 2008, 105, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Ureta, C.; González-Salazar, C.; González, E.J.; Álvarez-Buylla, E.R.; Martínez-Meyer, E. Environmental and social factors account for Mexican maize richness and distribution: A data mining approach. Agr. Ecosyst. Environ. 2013, 179, 25–34. [Google Scholar] [CrossRef]
- CONABIO. Base de Datos del Proyecto Global “Recopilación, Generación, Actualización y Análisis de Información Acerca de la Diversidad Genética de Maíces y sus Parientes Silvestres en México”. Available online: https://www.biodiversidad.gob.mx/diversidad/proyectoMaices (accessed on 3 September 2019).
- Matsuoka, Y.; Vigouroux, Y.; Goodman, M.M.; Sanchez, J.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.T.A. Cómo y dónde se originó el maíz. Investig. Cienc. 2005, 347, 68–72. [Google Scholar]
- Caballero-García, M.A.; Córdova-Téllez, L.; López-Herrera, A.D.J. Validación empírica de la teoría multicéntrica del origen y diversidad del maíz en México. Rev. Fitotec. Mex. 2019, 42, 357–366. [Google Scholar] [CrossRef]
- Piperno, D.R.; Ranere, A.J.; Holst, I.; Iriarte, J.; Dickau, R. Starch grain and phytolith evidence for early ninth millennium B.P. maize from the Central Balsas River Valley, Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 5019–5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hufford, M.B.; Martínez-Meyer, E.; Gaut, B.S.; Eguiarte, L.E.; Tenaillon, M.I. Inferences from the historical distribution of wild and domesticated maize provide ecological and evolutionary insight. PLoS ONE 2012, 7, e47659. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Suárez, R.; Morales-Chávez, L.A.; Gálvez-Mariscal, A. Importancia de los maíces nativos de México en la dieta nacional: Una revisión indispensable. Revi. Fitotec. Mex. 2013, 36, 275–283. [Google Scholar] [CrossRef]
- Perales, H.R.; Benz, B.F.; Brush, S.B. Maize diversity and ethnolinguistic diversity in Chiapas, Mexico. Proc. Natl. Acad. Sci. USA 2005, 102, 949–954. [Google Scholar] [CrossRef] [Green Version]
- Brush, S.B. In situ conservation of landraces in centers of crop diversity. Crop Sci. 1995, 35, 346–354. [Google Scholar] [CrossRef]
- Thrupp, L.A. Linking agricultural biodiversity and food security: The valuable role of agrobiodiversity for sustainable agriculture. Int. Aff. 2000, 76, 283–297. [Google Scholar] [CrossRef]
- Esquinas-Alcazar, J. Protecting crop genetic diversity for food security: Political, ethical and technical challenges. Nat. Rev. Genet. 2005, 6, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Guzzon, F.; Arandia-Rios, L.W.; Caviedes-Cepeda, G.M.; Céspedes-Polo, M.; Chavez-Cabrera, A.; Muriel-Figueroa, J.; Medina-Hoyos, A.E.; Jara-Calvo, T.W.; Molnar, T.L.; Narro-León, L.A.; et al. Conservation and use of Latin American maize diversity: Pillar of nutrition security and cultural heritage of humanity. Agronomy 2021, 11, 172. [Google Scholar] [CrossRef]
- Van Etten, J.; López, M.R.F.; Monterroso, L.G.M.; Samayoa, K.M.P. Genetic diversity of maize (Zea mays L. ssp. Mays) in communities of the western highlands of Guatemala: Geographical patterns and processes. Genet. Resour. Crop Evol. 2008, 55, 303–317. [Google Scholar] [CrossRef] [Green Version]
- Fenzi, M.; Jarvis, D.I.; Reyes, L.M.A.; Moreno, L.L.; Tuxill, J. Longitudinal analysis of maize diversity in Yucatan, Mexico: Influence of agro-ecological factors on landraces conservation and modern variety introduction. Plant. Genet. Resour. Characterisation Util. 2017, 15, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Perales, H.; Golicher, D. Mapping the diversity of maize races in Mexico. PLoS ONE 2014, 9, e114657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco-Ramírez, Q.; Perales, H.; Hijmans, R.J. Geographical distribution and diversity of maize (Zea mays L. subsp. mays) races in Mexico. Genet. Resour. Crop Evol. 2017, 64, 855–865. [Google Scholar] [CrossRef]
- Orozco-Ramírez, Q.; Ross-Ibarra, J.; Santacruz-Varela, A.; Brush, S. Maize diversity associated with social origin and environmental variation in Southern Mexico. Heredity 2016, 116, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Bellon, M.R.; Hodson, D.; Hellin, J. Assessing the vulnerability of traditional maize seed systems in Mexico to climate change. Proc. Natl. Acad. Sci. USA 2011, 108, 13432–13437. [Google Scholar] [CrossRef] [Green Version]
- Mercer, K.L.; Perales, H.R.; Wainwright, J.D. Climate change and the transgenic adaptation strategy: Smallholder livelihoods, climate justice, and maize landraces in Mexico. Glob. Environ. Change 2012, 22, 495–504. [Google Scholar] [CrossRef]
- Serratos-Hernández, J.A.; Islas-Gutiérrez, F.; Buendía-Rodríguez, E.; Berthaud, J. Gene flow scenarios with transgenic maize in Mexico. Environ. Biosaf. Res. 2004, 3, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schröder, W.; Schmidt, G. Overview of principles and implementations to deal with spatial issues in monitoring environmental effects of genetically modified organisms. Environ. Sci. Eur. 2012, 24, 6. [Google Scholar] [CrossRef] [Green Version]
- Lazos-Chavero, E. Consideraciones socioeconómicas y culturales en la controvertida introducción del maíz transgénico: El caso de Tlaxcala. Sociológica 2014, 29, 201–240. [Google Scholar]
- Dyer, G.A.; López-Feldman, A.; Yúnez-Naude, A.; Taylor, J.E. Genetic erosion in maize’s center of origin. Proc. Natl. Acad. Sci. USA 2014, 111, 14094–14099. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Schreuder, M.J.C.; de la Cruz, J.; Guarino, L. Using GIS to check coordinates of genebank accessions. Genet. Resour. Crop Evol. 1999, 46, 291–296. [Google Scholar] [CrossRef]
- Guarino, L.; Jarvis, A.; Hijmans, R.J.; Maxted, N. Geographic Information Systems (GIS) and the Conservation and Use of Plant Genetic Resources. In Managing Plant Genetic Diversity; Engels, J.M.M., Ramanatha, R.V.B., Jackson, M.T., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 387–404. [Google Scholar]
- Hijmans, R.J.; Garrett, K.A.; Huaman, Z.; Zhang, D.P.; Schreuder, M.; Bonierbale, M. Assessing the geographic representativeness of genebank collections: The case of Bolivian wild potatoes. Conserv. Biol. 2000, 14, 1755–1765. [Google Scholar] [CrossRef]
- Scheldeman, X.; Willemen, L.; d’Eeckenbrugge, G.C.; Romeijn-Peeters, E.; Restrepo, M.T.; Motoche, J.R.; Jiménez, D.; Lobo, M.; Medina, C.I.; Reyes, C.; et al. Distribution, diversity and environmental adaptation of highland papayas (Vasconcellea spp.) in tropical and subtropical America. In Plant Conservation and Biodiversity; Hawksworth, D.L., Bull, A.T., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 16, pp. 293–310. [Google Scholar] [CrossRef]
- Willemen, L.; Scheldeman, X.; Cabellos, V.S.; Salazar, S.R.; Guarino, L. Spatial patterns of diversity and genetic erosion of traditional cassava (Manihot esculenta Crantz) in the Peruvian Amazon: An evaluation of socio-economic and environmental indicators. Genet. Resour. Crop Evol. 2007, 54, 1599–1612. [Google Scholar] [CrossRef]
- Sánchez González, J.D.J.; Ruiz Corral, J.A.; García, G.M.; Ojeda, G.R.; Larios, J.B.; Holland, L.D.L.C.; Medrano, R.M.; García Romero, G.E. Ecogeography of Teosinte. PLoS ONE 2018, 13, e0192676. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Guarino, L.; Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Genet. Resour. Newsl. 2001, 127, 15–19. [Google Scholar]
- Viner, B.J.; Arritt, R.W. Increased pollen viability resulting from transport to the upper boundary layer. Field Crops Res. 2010, 119, 195–200. [Google Scholar] [CrossRef]
- Hijmans, R.; Cameron, E.; Parra, L.; Jones, G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- CONABIO. Elementos para la determinación de centros de origen y centros de diversidad genética en general y el caso específico de la liberación experimental de maíz transgénico al medio ambiente en México. Rev. Geogr. Agrícola 2006, 38, 121–140. [Google Scholar]
- Diker, K.; Heermann, D.F.; Bausch, W.C.; Wright, D.K. Shannon-Wiener’s diversity index for linking yield monitor and remotely sensed data for corn. Trans. ASABE 2004, 47, 1347–1354. [Google Scholar] [CrossRef]
- Kato, T.A.; Mapes, C.; Mera, L.M.; Serratos, J.A.; Bye, R.A. Origen y Diversificación del Maíz: Una Revisión Analítica; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2009; p. 116. [Google Scholar]
- CONABIO. Portal de Geoinformación 2020: Sistema Nacional de Información sobre Biodiversidad (SNIB). Available online: http://www.conabio.gob.mx/informacion/gis/ (accessed on 25 January 2020).
- Anguix, A.; Díaz, L. GvSIG: A GIS desktop solution for an open SDI. J. Geogr. Reg. Plann. 2008, 1, 041–048. [Google Scholar]
- Gómez-Orea, D. Ordenación del Territorio. Una Aproximación desde el Medio Físico; Agrícola Española: Madrid, Spain, 1994; p. 238. [Google Scholar]
- National Commission for the Development of Indigenous Peoples. Territorios de Pueblos Indígenas. Available online: http://132.248.14.102/layers/CapaBase:ii_4_3_terri_indigena (accessed on 14 November 2020).
- Team, R. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 4 December 2020).
- Graham, C.H.; Hijmans, R.J. A comparison of methods for mapping species ranges and species richness. Glob. Ecol. Biogeogr. 2006, 15, 578–587. [Google Scholar] [CrossRef]
- Aragón-Cuevas, F.; Taba, S.; Hernández-Casillas, J.; Figueroa, C.J.; Serrano, A.B.; Castro-García, F. Catálogo de Maíces Criollos de Oaxaca; INIFAP-SAGARPA: Oaxaca, Mexico, 2006; p. 343. [Google Scholar]
- Ureta, C.; Martínez-Meyer, E.; Perales, H.R.; Álvarez-Buylla, E.R. Projecting the effects of climate change on the distribution of maize races and their wild relatives in Mexico. Glob. Chang. Biol. 2012, 18, 1073–1082. [Google Scholar] [CrossRef]
- Bonett, D.G.; Wright, T.A. Sample size requirements for estimating Pearson, Kendall and Spearman correlations. Psychometrika 2000, 65, 23–28. [Google Scholar] [CrossRef]
- Brush, S.B.; Perales, H.R. A maize landscape: Ethnicity and agrobiodiversity in Chiapas Mexico. Agric. Ecosyst. Environ. 2007, 121, 211–221. [Google Scholar] [CrossRef]
- Masas, O.; Flour, N.C. Propiedades químicas, fisicoquímicas y reológicas de masas y harinas de maíz nixtamalizado. Agrociencia 2002, 36, 319–328. [Google Scholar]
- Godoy, A.; Herrera, T.; Ulloa, M. Más Allá del Pulque y el Tepache. Las Bebidas Alcohólicas no Destiladas Indígenas de México; Universidad Nacional Autonoma de México: Mexico City, Mexico, 2003; p. 112. [Google Scholar]
- Olivares-Illana, V.; Wacher-Rodarte, C.; Le-Borgne, S.; López-Munguía, A. Characterization of a cell-associated inulosucrase from a novel source: A Leuconostoc citreum strain isolated from Pozol, a fermented corn beverage of Mayan origin. J. Ind. Microbiol. Biotechnol. 2002, 28, 112–117. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Unit | Collections | Races | Principal Races | Ethnic Groups | Index of Maize Grain and Plant Uses (IU) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Number | % Total | Number | % Total | Number | % Total | Number | % Total | ||||
| North Pacific | 516 | 2.74 | 25 | 39.06 | 7 | 10.94 | 7 | 11.11 | 45 | 960 | 0.05 |
| Tarahumara Sierras | 993 | 5.28 | 25 | 39.06 | 5 | 7.81 | 3 | 4.76 | 35 | 960 | 0.04 |
| Northeast | 379 | 2.01 | 8 | 12.50 | 2 | 3.13 | 0 | 0.00 | 16 | 960 | 0.02 |
| Center | 6010 | 31.95 | 49 | 76.56 | 23 | 35.94 | 18 | 28.57 | 162 | 960 | 0.17 |
| South Pacific | 3340 | 17.75 | 44 | 68.75 | 14 | 21.88 | 17 | 26.98 | 96 | 960 | 0.10 |
| South East | 4196 | 22.30 | 33 | 51.56 | 11 | 17.19 | 24 | 38.10 | 82 | 960 | 0.09 |
| Yucatán Peninsula | 491 | 2.61 | 8 | 12.50 | 2 | 3.13 | 5 | 7.94 | 17 | 960 | 0.02 |
| Sum | 15,925 | 84.65 | 64 | 100.00 | |||||||
| Total | 18,812 | 100.00 | 64 | 100.00 | 64 | 100.00 | 63 | 100.00 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santillán-Fernández, A.; Salinas-Moreno, Y.; Valdez-Lazalde, J.R.; Bautista-Ortega, J.; Pereira-Lorenzo, S. Spatial Delimitation of Genetic Diversity of Native Maize and Its Relationship with Ethnic Groups in Mexico. Agronomy 2021, 11, 672. https://doi.org/10.3390/agronomy11040672
Santillán-Fernández A, Salinas-Moreno Y, Valdez-Lazalde JR, Bautista-Ortega J, Pereira-Lorenzo S. Spatial Delimitation of Genetic Diversity of Native Maize and Its Relationship with Ethnic Groups in Mexico. Agronomy. 2021; 11(4):672. https://doi.org/10.3390/agronomy11040672
Chicago/Turabian StyleSantillán-Fernández, Alberto, Yolanda Salinas-Moreno, José René Valdez-Lazalde, Jaime Bautista-Ortega, and Santiago Pereira-Lorenzo. 2021. "Spatial Delimitation of Genetic Diversity of Native Maize and Its Relationship with Ethnic Groups in Mexico" Agronomy 11, no. 4: 672. https://doi.org/10.3390/agronomy11040672