
Grapevine Rootstocks Differently Affect Physiological and Molecular Responses of the Scion under Water Deficit Condition

 , , , ,
, , , , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Leaf Physiological Measurements
2.3. Contents of Amino Acids, Total Sugars and Potassium
2.4. ABA Determination
2.5. qPCR Analysis of Gene Expression
2.6. Statistical Analysis
3. Results and Discussion
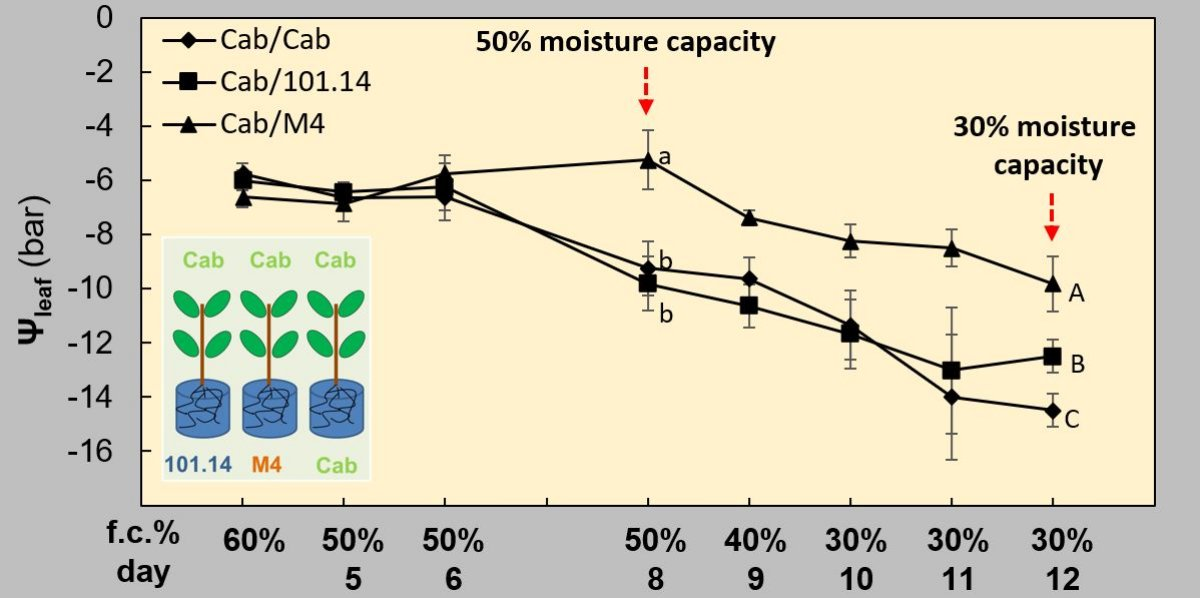
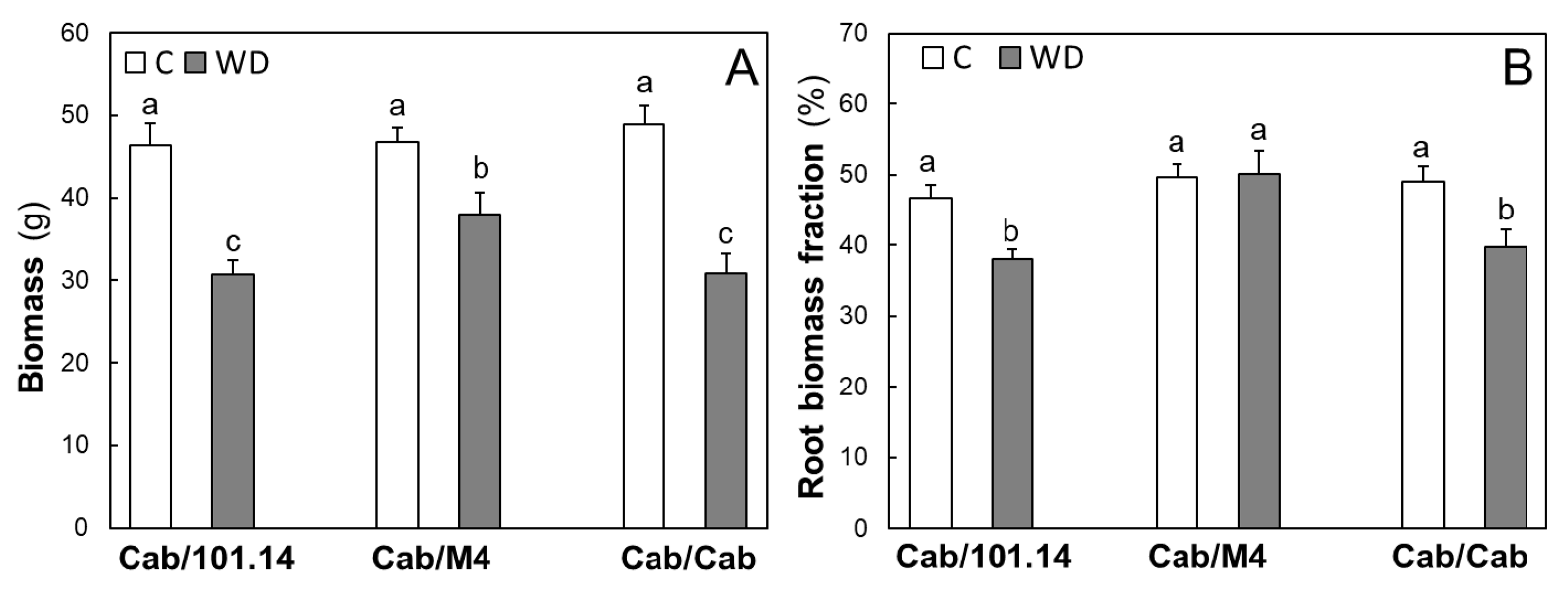
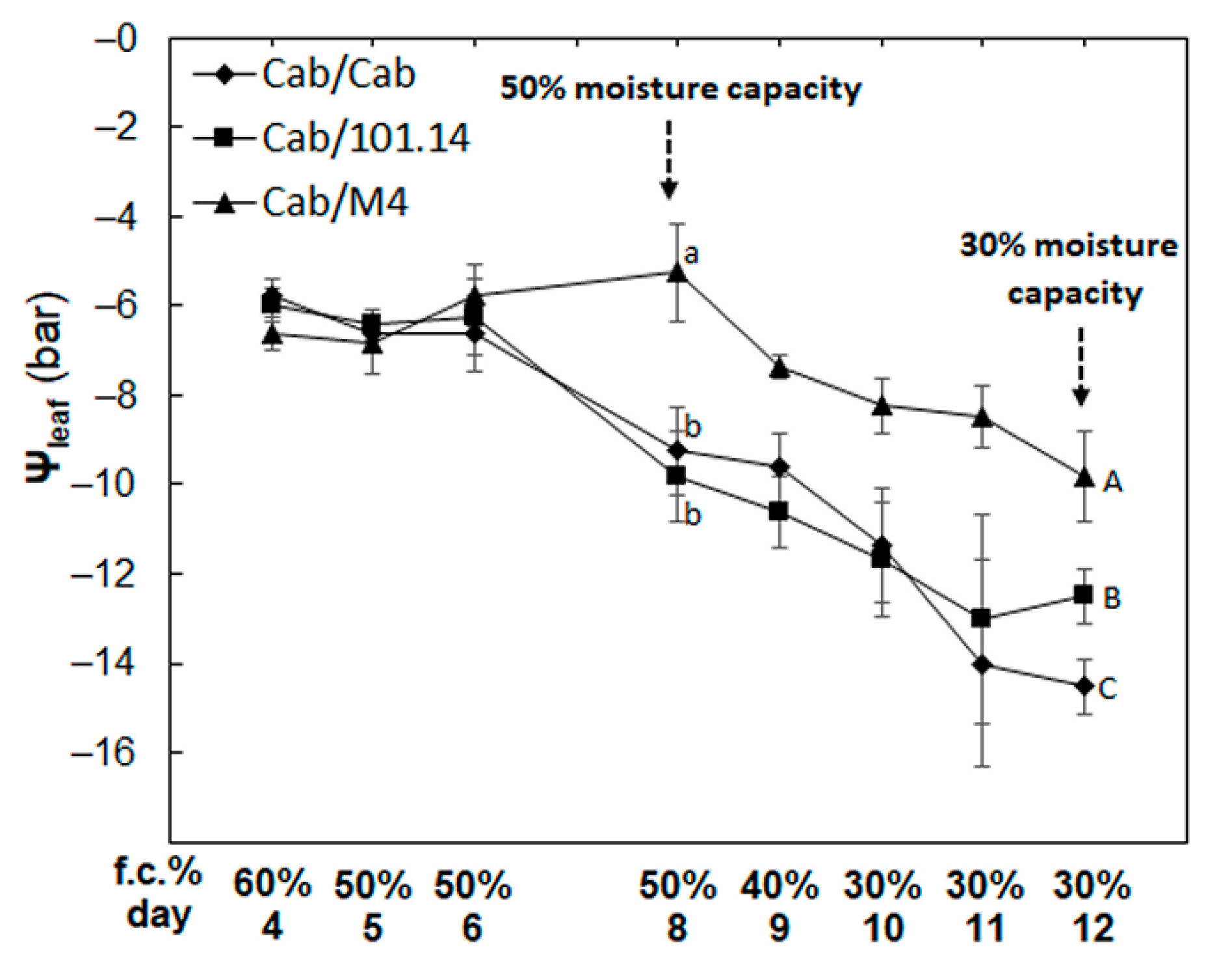
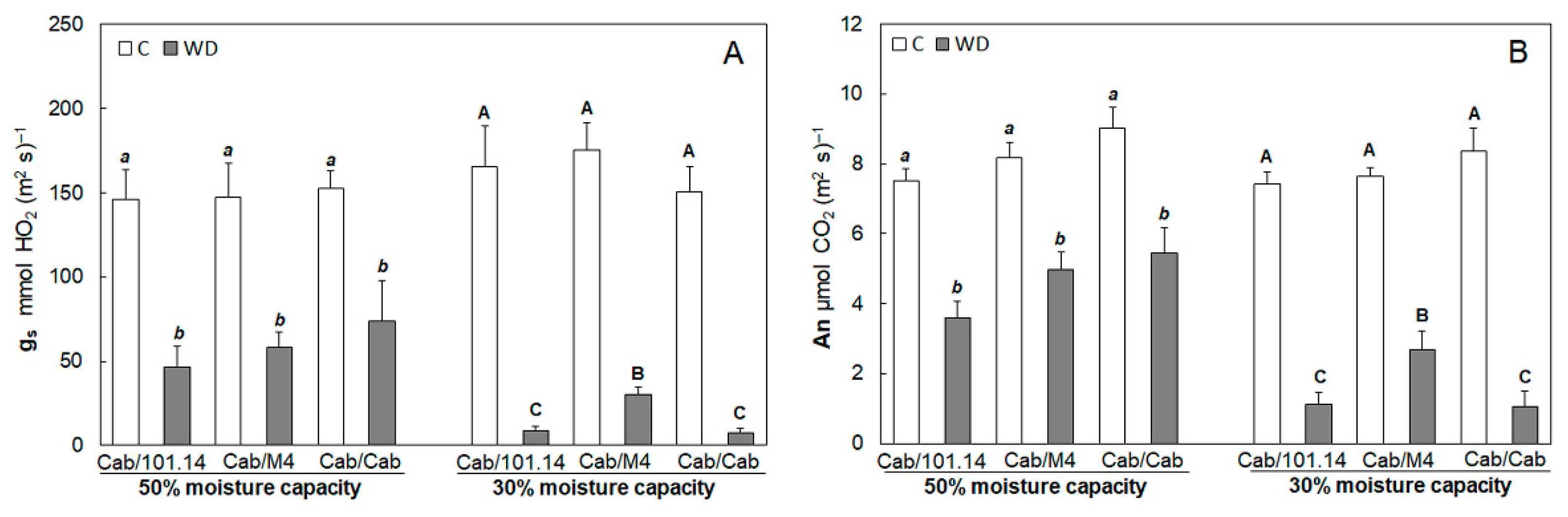
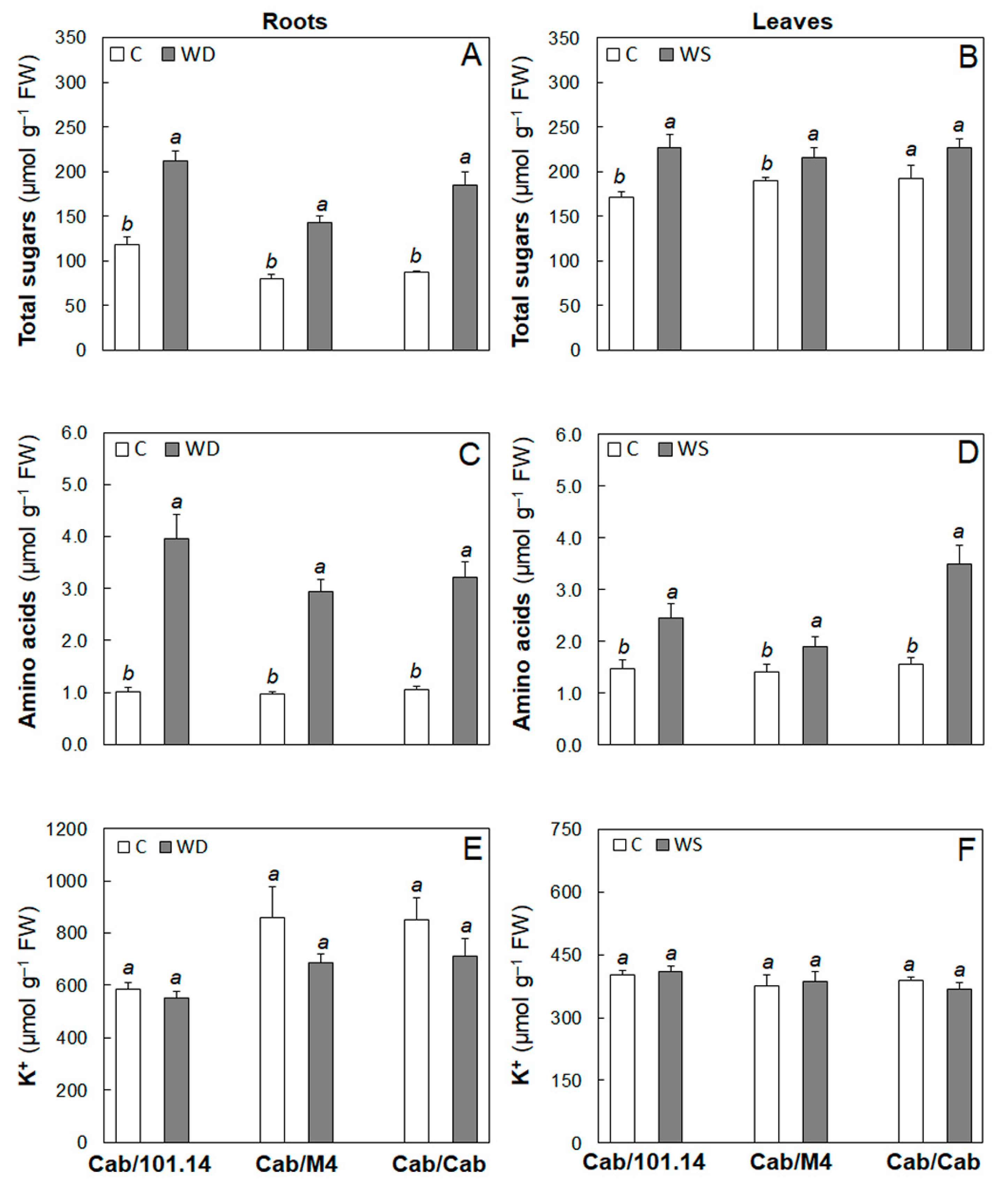
3.1. Physiological and Biochemical Parameters
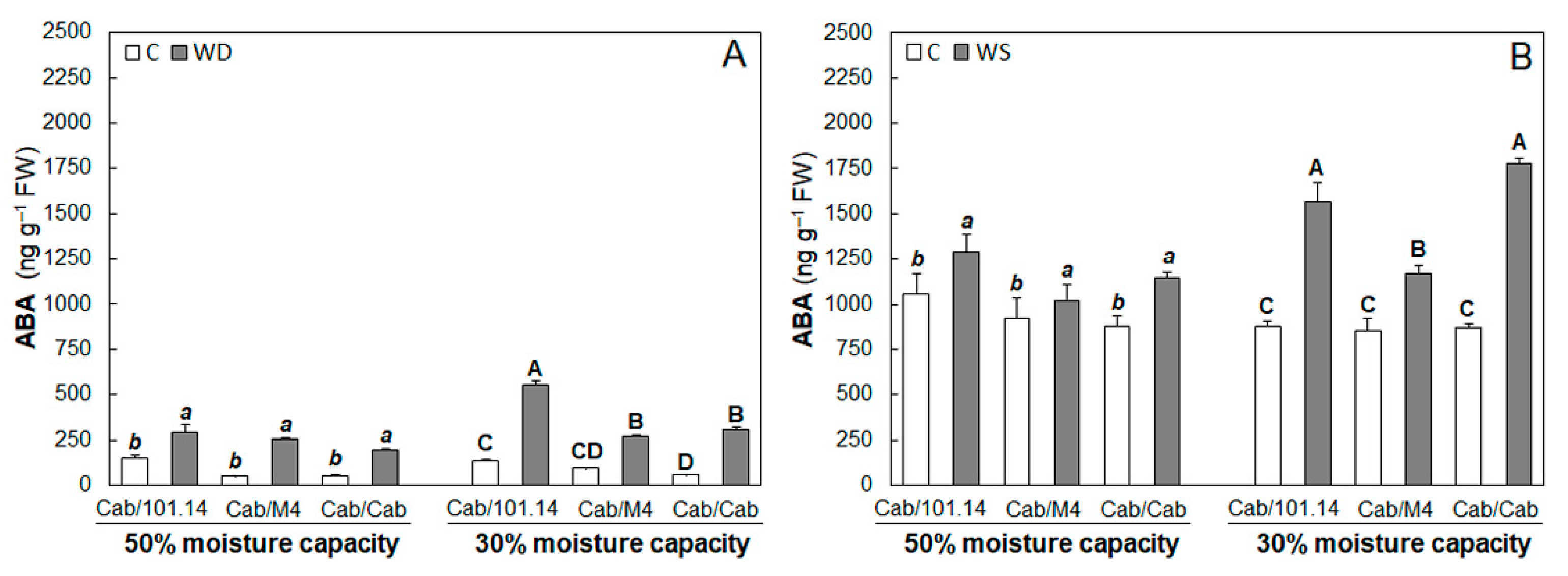
3.2. ABA Contents in Roots and Leaves
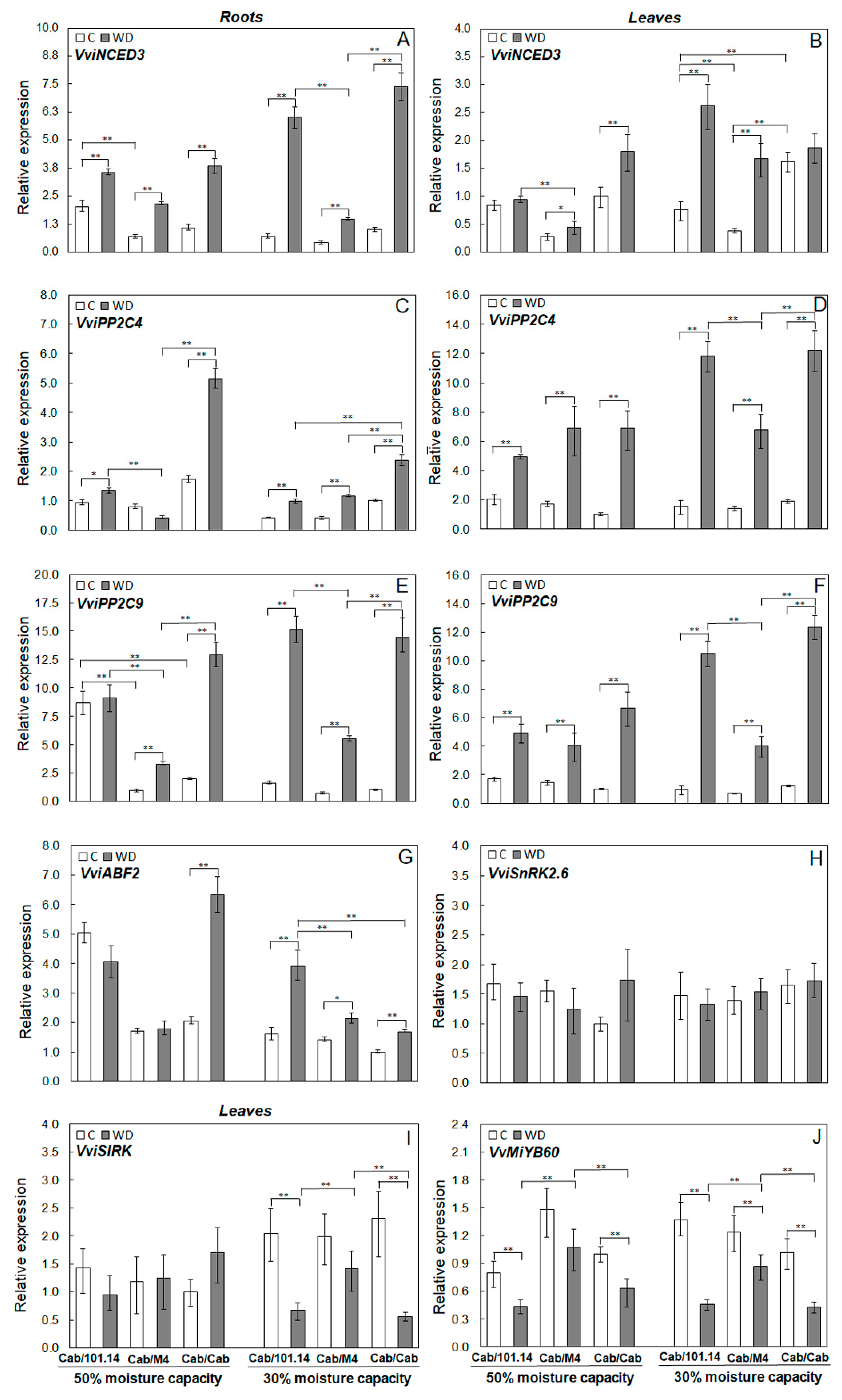
3.3. Expression of ABA-Related Genes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, A. Grapevine Breeding Programs for the Wine Industry, 1st ed.; Elsevier: Cambridge, UK, 2015. [Google Scholar]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate Change and Global Wine Quality. Clim. Chang. 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Matthews, M.A.; Di Gaspero, G.; Gambetta, G.A. Water Deficits Accelerate Ripening and Induce Changes in Gene Expression Regulating Flavonoid Biosynthesis in Grape Berries. Planta 2007, 227, 101–112. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under Deficit Irrigation: Hints from Physiological and Molecular Data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, H.R.; Stoll, M. Some Critical Issues in Environmental Physiology of Grapevines: Future Challenges and Current Limitations. Aust. J. Grape Wine Res. 2010, 16, 4–24. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Manuck, C.M.; Drucker, S.T.; Shaghasi, T.; Fort, K.; Matthews, M.A.; Walker, M.A.; McElrone, A.J. The Relationship between Root Hydraulics and Scion Vigour across Vitis Rootstocks: What Role Do Root Aquaporins Play? J. Exp. Bot. 2012, 63, 6445–6455. [Google Scholar] [CrossRef]
- Marguerit, E.; Brendel, O.; Lebon, E.; Leeuwen, C.V.; Ollat, N. Rootstock Control of Scion Transpiration and Its Acclimation to Water Deficit Are Controlled by Different Genes. New Phytol. 2012, 194, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Tramontini, S.; Vitali, M.; Centioni, L.; Schubert, A.; Lovisolo, C. Rootstock Control of Scion Response to Water Stress in Grapevine. Environ. Exp. Bot. 2013, 93, 20–26. [Google Scholar] [CrossRef]
- Corso, M.; Bonghi, C. Grapevine Rootstock Effects on Abiotic Stress Tolerance. Plant Sci. Today 2014, 1, 108–113. [Google Scholar] [CrossRef]
- Corso, M.; Vannozzi, A.; Maza, E.; Vitulo, N.; Meggio, F.; Pitacco, A.; Telatin, A.; D’Angelo, M.; Feltrin, E.; Negri, A.S.; et al. Comprehensive Transcript Profiling of Two Grapevine Rootstock Genotypes Contrasting in Drought Susceptibility Links the Phenylpropanoid Pathway to Enhanced Tolerance. J. Exp. Bot. 2015, 66, 5739–5752. [Google Scholar] [CrossRef]
- Pagliarani, C.; Vitali, M.; Ferrero, M.; Vitulo, N.; Incarbone, M.; Lovisolo, C.; Valle, G.; Schubert, A. The Accumulation of MiRNAs Differentially Modulated by Drought Stress Is Affected by Grafting in Grapevine. Plant Physiol. 2017, 173, 2180–2195. [Google Scholar] [CrossRef] [Green Version]
- Prinsi, B.; Negri, A.S.; Failla, O.; Scienza, A.; Espen, L. Root Proteomic and Metabolic Analyses Reveal Specific Responses to Drought Stress in Differently Tolerant Grapevine Rootstocks. BMC Plant Biol. 2018, 18, 126. [Google Scholar] [CrossRef]
- Skirycz, A.; Inzé, D. More from Less: Plant Growth under Limited Water. Curr. Opin. Biotechnol. 2010, 21, 197–203. [Google Scholar] [CrossRef]
- Munns, R. Chapter 1—Plant Adaptations to Salt and Water Stress: Differences and Commonalities. In Advances in Botanical Research; Turkan, I., Ed.; Plant Responses to Drought and Salinity Stress; Academic Press: Cambridge, MA, USA, 2011; Volume 57, pp. 1–32. [Google Scholar] [CrossRef]
- Kantar, M.; Lucas, S.J.; Budak, H. Chapter 13—Drought Stress: Molecular Genetics and Genomics Approaches. In Advances in Botanical Research; Turkan, I., Ed.; Plant Responses to Drought and Salinity Stress; Academic Press: Cambridge, MA, USA, 2011; Volume 57, pp. 445–493. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding Plant Responses to Drought—From Genes to the Whole Plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Lovisolo, C.; Schubert, A. Effects of Water Stress on Vessel Size and Xylem Hydraulic Conductivity in Vitis Vinifera L. J. Exp. Bot. 1998, 49, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Tombesi, S.; Johnson, R.S.; Day, K.R.; DeJong, T.M. Relationships between Xylem Vessel Characteristics, Calculated Axial Hydraulic Conductance and Size-Controlling Capacity of Peach Rootstocks. Ann. Bot. 2010, 105, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Dayer, S.; Scharwies, J.D.; Ramesh, S.A.; Sullivan, W.; Doerflinger, F.C.; Pagay, V.; Tyerman, S.D. Comparing Hydraulics Between Two Grapevine Cultivars Reveals Differences in Stomatal Regulation Under Water Stress and Exogenous ABA Applications. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of Root Water Uptake under Abiotic Stress Conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Sharp, R.E. Complexity and Coordination of Root Growth at Low Water Potentials: Recent Advances from Transcriptomic and Proteomic Analyses. Plant Cell Environ. 2010, 33, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of Osmoprotectants in Improving Salinity and Drought Tolerance in Plants: A Review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Hussain, S.; Rao, M.J.; Anjum, M.A.; Ejaz, S.; Zakir, I.; Ali, M.A.; Ahmad, N.; Ahmad, S. Oxidative Stress and Antioxidant Defense in Plants Under Drought Conditions. In Plant Abiotic Stress Tolerance: Agronomic, Molecular and Biotechnological Approaches; Hasanuzzaman, M., Hakeem, K.R., Nahar, K., Alharby, H.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 207–219. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Rajasheker, G.; Jawahar, G.; Jalaja, N.; Kumar, S.A.; Kumari, P.H.; Punita, D.L.; Karumanchi, A.R.; Reddy, P.S.; Rathnagiri, P.; Sreenivasulu, N.; et al. Chapter 27—Role and Regulation of Osmolytes and ABA Interaction in Salt and Drought Stress Tolerance. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 417–436. [Google Scholar] [CrossRef]
- Rosales, M.A.; Maurel, C.; Nacry, P. Abscisic Acid Coordinates Dose-Dependent Developmental and Hydraulic Responses of Roots to Water Deficit. Plant Physiol. 2019, 180, 2198–2211. [Google Scholar] [CrossRef] [Green Version]
- Kuromori, T.; Seo, M.; Shinozaki, K. ABA Transport and Plant Water Stress Responses. Trends Plant Sci. 2018, 23, 513–522. [Google Scholar] [CrossRef]
- Christmann, A.; Weiler, E.W.; Steudle, E.; Grill, E. A Hydraulic Signal in Root-to-Shoot Signalling of Water Shortage. Plant J. 2007, 52, 167–174. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Goodger, J.Q.D. Chemical Root to Shoot Signaling under Drought. Trends Plant Sci. 2008, 13, 281–287. [Google Scholar] [CrossRef]
- Takahashi, F.; Shinozaki, K. Long-Distance Signaling in Plant Stress Response. Curr. Opin. Plant Biol. 2019, 47, 106–111. [Google Scholar] [CrossRef]
- Rattanakon, S.; Ghan, R.; Gambetta, G.A.; Deluc, L.G.; Schlauch, K.A.; Cramer, G.R. Abscisic Acid Transcriptomic Signaling Varies with Grapevine Organ. BMC Plant Biol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Galmés, J.; Gallé, A.; Gulías, J.; Pou, A.; Ribas-Carbo, M.; Tomàs, M.; Medrano, H. Improving Water Use Efficiency in Grapevines: Potential Physiological Targets for Biotechnological Improvement. Aust. J. Grape Wine Res. 2010, 16, 106–121. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-Induced Changes in Development and Function of Grapevine (Vitis Spp.) Organs and in Their Hydraulic and Non-Hydraulic Interactions at the Whole-Plant Level: A Physiological and Molecular Update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Bota, J.; Tomás, M.; Flexas, J.; Medrano, H.; Escalona, J.M. Differences among Grapevine Cultivars in Their Stomatal Behavior and Water Use Efficiency under Progressive Water Stress. Agric. Water Manag. 2016, 164, 91–99. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Water-Use Efficiency in Grapevine Cultivars Grown under Controlled Conditions: Effects of Water Stress at the Leaf and Whole-Plant Level. Aust. J. Grape Wine Res. 2012, 18, 164–172. [Google Scholar] [CrossRef]
- Serra, I.; Strever, A.; Myburgh, P.A.; Deloire, A. Review: The Interaction between Rootstocks and Cultivars (Vitis Vinifera L.) to Enhance Drought Tolerance in Grapevine. Aust. J. Grape Wine Res. 2014, 20, 1–14. [Google Scholar] [CrossRef]
- Rossdeutsch, L.; Edwards, E.; Cookson, S.J.; Barrieu, F.; Gambetta, G.A.; Delrot, S.; Ollat, N. ABA-Mediated Responses to Water Deficit Separate Grapevine Genotypes by Their Genetic Background. BMC Plant Biol. 2016, 16, 91. [Google Scholar] [CrossRef] [Green Version]
- Carbonneau, A. The Early Selection of Grapevine Rootstocks for Resistance to Drought Conditions. Am. J. Enol. Vitic. 1985, 36, 195–198. [Google Scholar]
- Meggio, F.; Prinsi, B.; Negri, A.S.; Lorenzo, G.S.D.; Lucchini, G.; Pitacco, A.; Failla, O.; Scienza, A.; Cocucci, M.; Espen, L. Biochemical and physiological responses of two grapevine rootstock genotypes to drought and salt treatments. Aust. J. Grape Wine Res. 2014, 20, 310–323. [Google Scholar] [CrossRef]
- Galbignani, M.; Merli, M.C.; Magnanini, E.; Bernizzoni, F.; Talaverano, I.; Gatti, M.; Tombesi, S.; Palliotti, A.; Poni, S. Gas Exchange and Water-Use Efficiency of Cv. Sangiovese Grafted to Rootstocks of Varying Water-Deficit Tolerance. Irrig. Sci. 2016, 34, 105–116. [Google Scholar] [CrossRef]
- Frioni, T.; Biagioni, A.; Squeri, C.; Tombesi, S.; Gatti, M.; Poni, S. Grafting Cv. Grechetto Gentile Vines to New M4 Rootstock Improves Leaf Gas Exchange and Water Status as Compared to Commercial 1103P Rootstock. Agronomy 2020, 10, 708. [Google Scholar] [CrossRef]
- Moore, S.; Stein, W.H. A Modified Ninhydrin Reagent for the Photometric Determination of Amino Acids and Related Compounds. J. Biol. Chem. 1954, 211, 907–913. [Google Scholar] [CrossRef]
- Nelson, N. A Photometric Adaptation of the Somogyi Method for the Determination of Glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar] [CrossRef]
- Speirs, J.; Binney, A.; Collins, M.; Edwards, E.; Loveys, B. Expression of ABA synthesis and metabolism genes under different irrigation strategies and atmospheric VPDs is associated with stomatal conductance in grapevine (Vitis vinifera L. cv Cabernet Sauvignon). J. Exp. Bot. 2013, 64, 1907–1916. [Google Scholar] [CrossRef] [Green Version]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Matus, J.T.; Loyola, R.; Vega, A.; Peña-Neira, A.; Bordeu, E.; Arce-Johnson, P.; Alcalde, J.A. Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berry skins of Vitis vinifera. J. Exp. Bot. 2009, 60, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Young, P.R.; Lashbrooke, J.G.; Alexandersson, E.; Jacobson, D.; Moser, C.; Velasco, R.; Vivier, M.A. The Genes and Enzymes of the Carotenoid Metabolic Pathway in Vitis Vinifera L. BMC Genom. 2012, 13, 243. [Google Scholar] [CrossRef] [Green Version]
- Boneh, U.; Biton, I.; Zheng, C.; Schwartz, A.; Ben-Ari, G. Characterization of Potential ABA Receptors in Vitis Vinifera. Plant Cell Rep. 2012, 31, 311–321. [Google Scholar] [CrossRef]
- Boneh, U.; Biton, I.; Schwartz, A.; Ben-Ari, G. Characterization of the ABA Signal Transduction Pathway in Vitis Vinifera. Plant Sci. 2012, 187, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, P.; Lecourieux, D.; Kappel, C.; Cluzet, S.; Cramer, G.; Delrot, S.; Lecourieux, F. The Basic Leucine Zipper Transcription Factor ABSCISIC ACID RESPONSE ELEMENT-BINDING FACTOR2 Is an Important Transcriptional Regulator of Abscisic Acid-Dependent Grape Berry Ripening Processes. Plant Physiol. 2014, 164, 365–383. [Google Scholar] [CrossRef] [Green Version]
- Galbiati, M.; Matus, J.T.; Francia, P.; Rusconi, F.; Cañón, P.; Medina, C.; Conti, L.; Cominelli, E.; Tonelli, C.; Arce-Johnson, P. The grapevine guard cell-related VviMYB60 transcription factor is involved in the regulation of stomatal activity and is differentially expressed in response to ABA and osmotic stress. BMC Plant Biol. 2011, 11, 142. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, R.; Lacombe, B.; Torregrosa, L.; Gaymard, F.; Romieu, C.; Thibaud, J.-B.; Sentenac, H. A Grapevine Gene Encoding a Guard Cell K+ Channel Displays Developmental Regulation in the Grapevine Berry. Plant Physiol. 2002, 128, 564–577. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.-S.P. Response of Plants to Water Stress. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soar, C.J.; Dry, P.R.; Loveys, B.R. Scion Photosynthesis and Leaf Gas Exchange in Vitis Vinifera L. Cv. Shiraz: Mediation of Rootstock Effects via Xylem Sap ABA. Aust. J. Grape Wine Res. 2006, 12, 82–96. [Google Scholar] [CrossRef]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA Perception and Signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Mustilli, A.-C.; Merlot, S.; Vavasseur, A.; Fenzi, F.; Giraudat, J. Arabidopsis OST1 Protein Kinase Mediates the Regulation of Stomatal Aperture by Abscisic Acid and Acts Upstream of Reactive Oxygen Species Production. Plant Cell 2002, 14, 3089–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mogami, J.; Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Four Arabidopsis AREB/ABF Transcription Factors Function Predominantly in Gene Expression Downstream of SnRK2 Kinases in Abscisic Acid Signalling in Response to Osmotic Stress. Plant Cell Environ. 2015, 38, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilati, S.; Bagagli, G.; Sonego, P.; Moretto, M.; Brazzale, D.; Castorina, G.; Simoni, L.; Tonelli, C.; Guella, G.; Engelen, K.; et al. Abscisic Acid Is a Major Regulator of Grape Berry Ripening Onset: New Insights into ABA Signaling Network. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Leonhardt, N.; Dellaporta, S.L.; Tonelli, C. A Guard-Cell-Specific MYB Transcription Factor Regulates Stomatal Movements and Plant Drought Tolerance. Curr. Biol. 2005, 15, 1196–1200. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Id | Gene Function | Forward (5′-3′) | Reverse (5′-3′) | Ref. |
|---|---|---|---|---|---|
| VviNCED3 | VIT_19s0093g00550 | ABA biosynthesis | GCAGAGGACGAGAGTGTAAAGGA | GCAGAGGACGAGAGTGTAAAGGA | [47] |
| VviPP2C9 | VIT_06s0004g05460 | ABA signaling | TTAAAGCCCTTCGTGAGCTG | TTAAAGCCCTTCGTGAGCTG | [48] |
| VviPP2C4 | VIT_11s0016g03180 | ABA signaling | CACAGGATTGATGGGAAACC | CACAGGATTGATGGGAAACC | [48] |
| VviSnRk2.6 | VIT_03s0063g01080 | ABA signaling | ACTACCGGTCGGTGACTACG | TCCTCTGTGTTCCCTTCTGG | [49] |
| VviABF2 | VIT_18s0001g10450 | Regulation of gene expression | CACAGGATTGATGGGAAACC | CACAGGATTGATGGGAAACC | [50] |
| VviMYB60 | VIT_08s0056g00800 | Stomatal opening | TTGAGTACGAAAACCTGAATGAT | TTGAGTACGAAAACCTGAATGAT | [51] |
| VviSIRK | VIT_07s0031g01440 | Stomatal opening | AGTCCCCGTTACAGGGCTTGGG | AGTCCCCGTTACAGGGCTTGGG | [52] |
| VviG3PDH | VIT_01s0010g02460 | Reference gene | TTAAAGCCCTTCGTGAGCTG | TTAAAGCCCTTCGTGAGCTG | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prinsi, B.; Simeoni, F.; Galbiati, M.; Meggio, F.; Tonelli, C.; Scienza, A.; Espen, L. Grapevine Rootstocks Differently Affect Physiological and Molecular Responses of the Scion under Water Deficit Condition. Agronomy 2021, 11, 289. https://doi.org/10.3390/agronomy11020289
Prinsi B, Simeoni F, Galbiati M, Meggio F, Tonelli C, Scienza A, Espen L. Grapevine Rootstocks Differently Affect Physiological and Molecular Responses of the Scion under Water Deficit Condition. Agronomy. 2021; 11(2):289. https://doi.org/10.3390/agronomy11020289
Chicago/Turabian StylePrinsi, Bhakti, Fabio Simeoni, Massimo Galbiati, Franco Meggio, Chiara Tonelli, Attilio Scienza, and Luca Espen. 2021. "Grapevine Rootstocks Differently Affect Physiological and Molecular Responses of the Scion under Water Deficit Condition" Agronomy 11, no. 2: 289. https://doi.org/10.3390/agronomy11020289