
Soil Bacteria Mediate Soil Organic Carbon Sequestration under Different Tillage and Straw Management in Rice-Wheat Cropping Systems
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
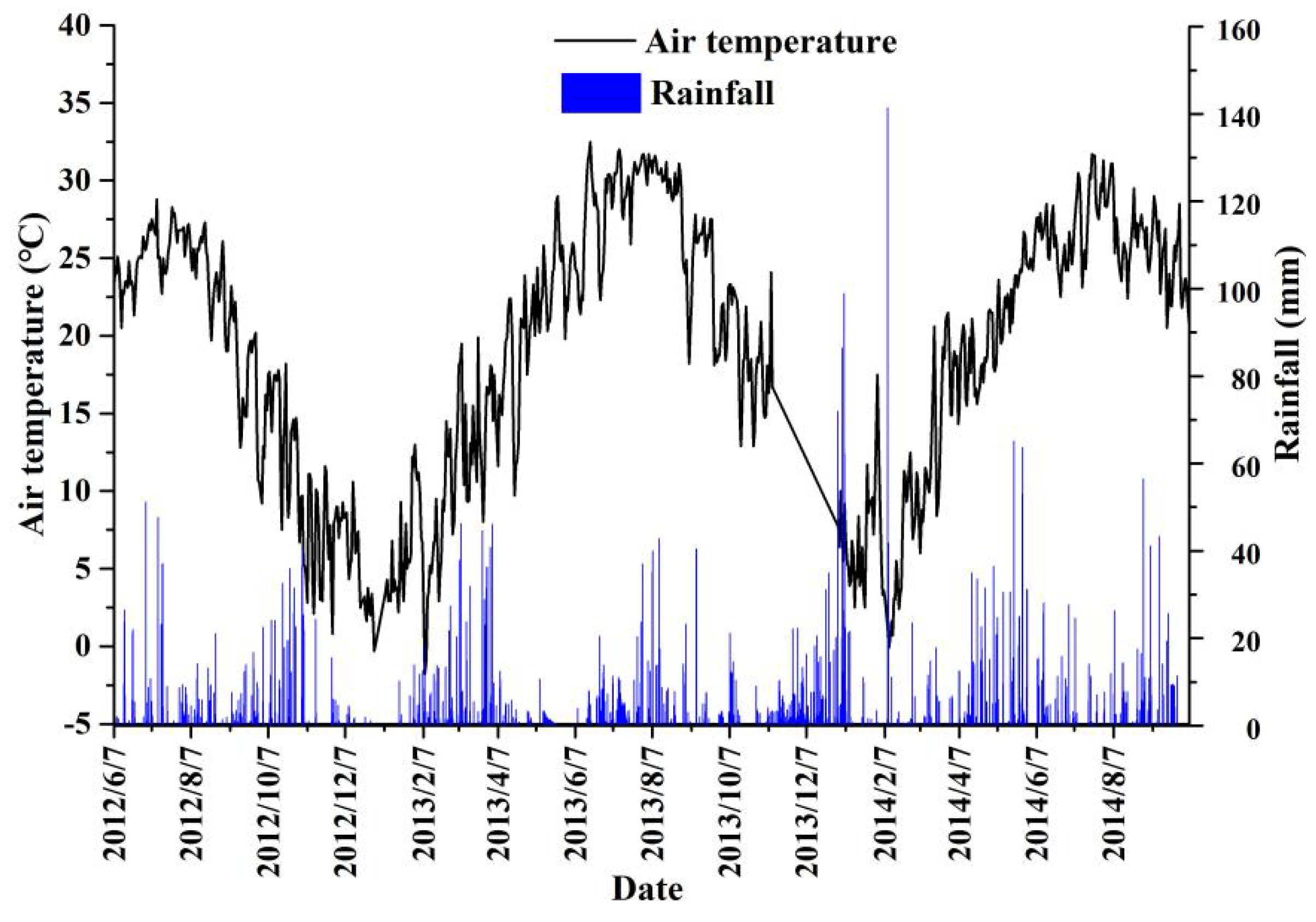
2.1. Experimental Site
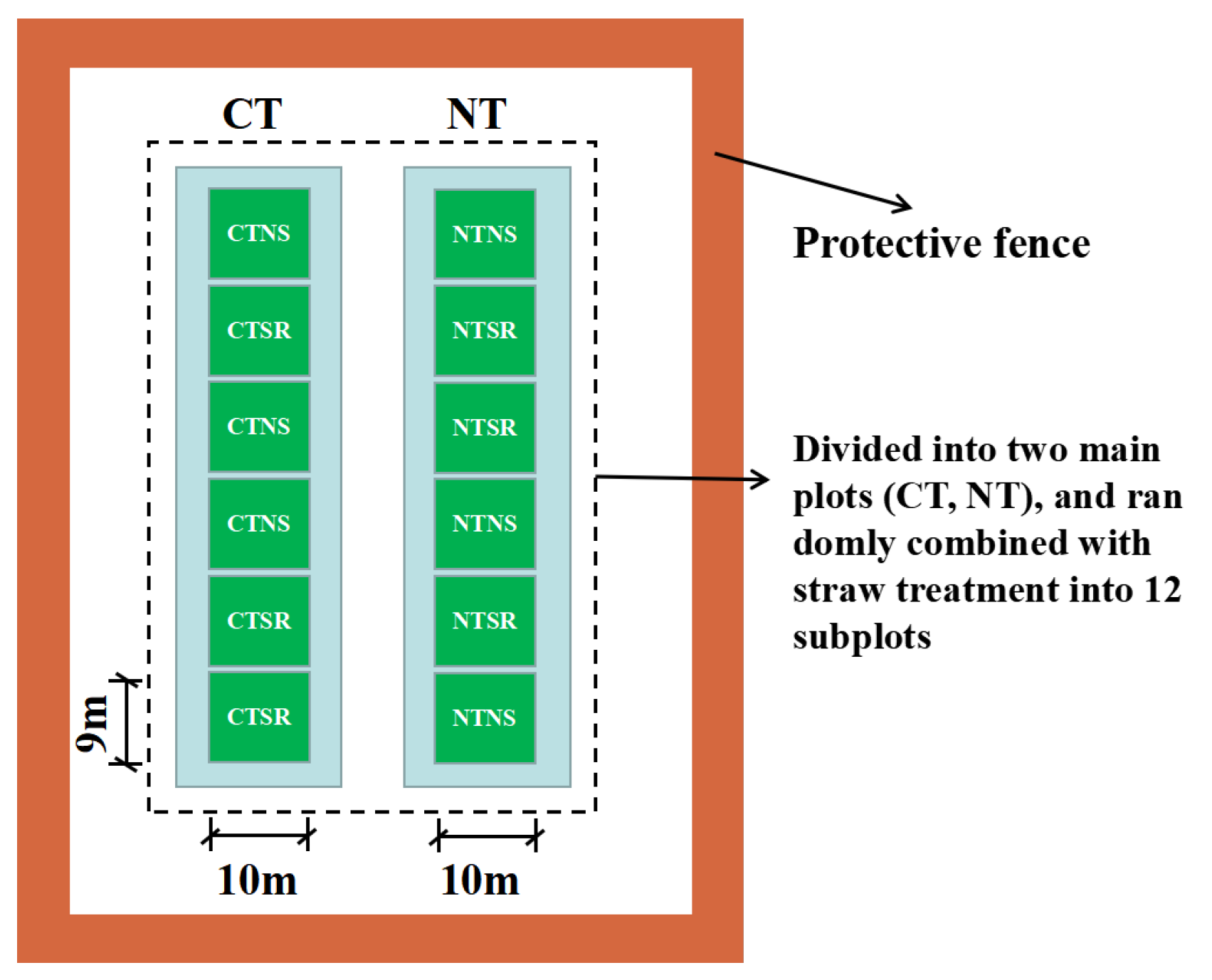
2.2. Experimental Design
2.3. Soil Sampling and Physicochemical Analysis
2.4. Phospholipid Fatty Acid Pattern
2.5. Measurement of Crop Grain Yields
2.6. Measurement of CH4 and CO2 Emissions
2.7. High-Throughput Sequencing
2.8. Statistical Analysis
3. Results
3.1. Soil Organic Carbon
3.2. Distribution of Soil Aggregates
3.3. Soil Organic Carbon Content within Aggregates
3.4. Soil Dissolved Organic Carbon and Microbial Biomass Carbon
3.5. Greenhouse Gas Emissions
3.6. Soil Microbial Community
3.7. Soil Bacterial Community
3.8. Crop Grain Yields and Their Relationship with Soil Properties
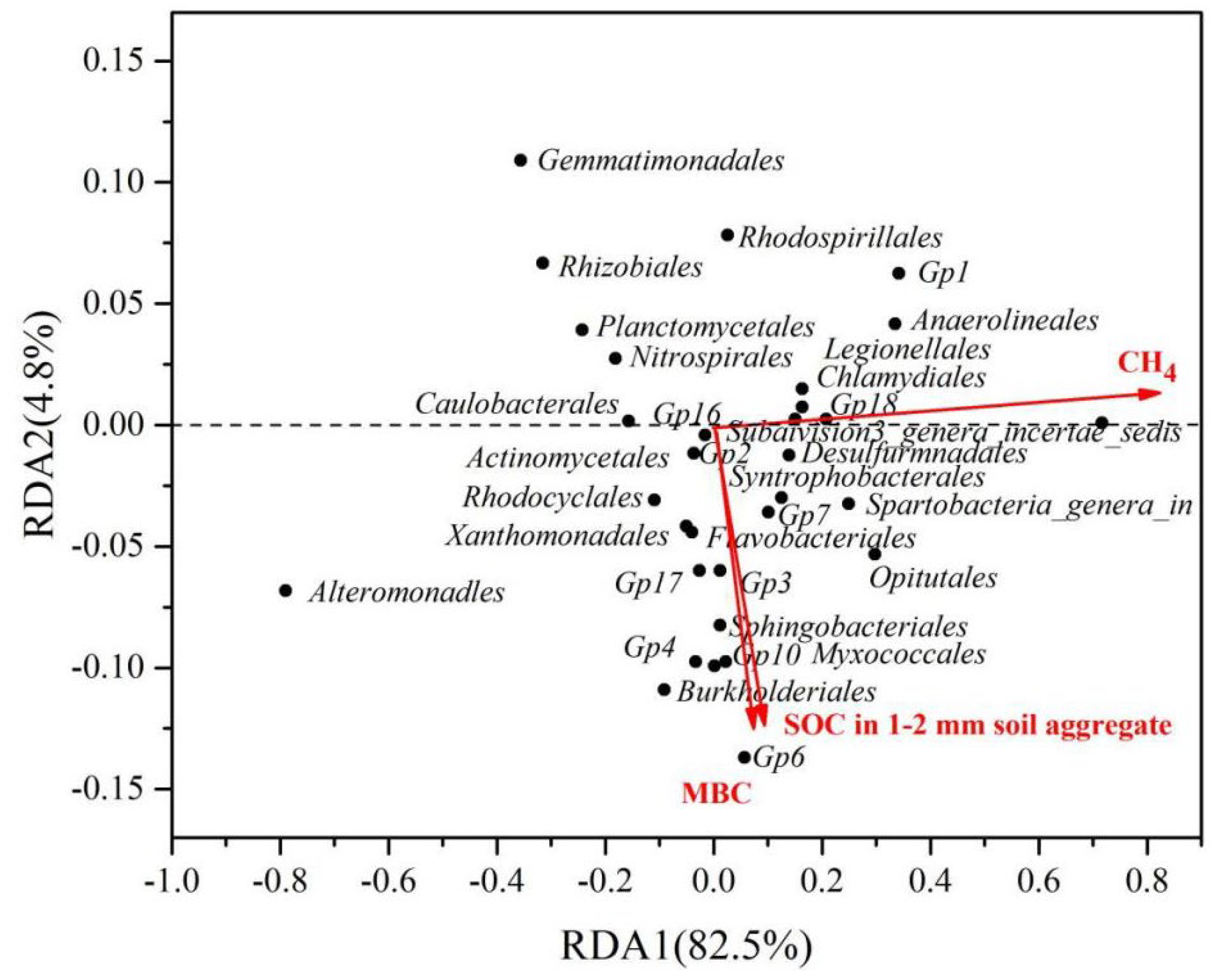
3.9. Relationship of Bacterial Community with Yield, Soil Aggregates and Soil Organic Carbon Fractions
4. Discussion
4.1. Impact of NT and Straw Return on SOC Content in Aggregates
4.2. Effect of NT and Straw Return on Greenhouse Gas Emissions
4.3. Effects of NT and Straw Return on Bacterial Community
4.4. Effect of NT and Straw Return on Crop Yields
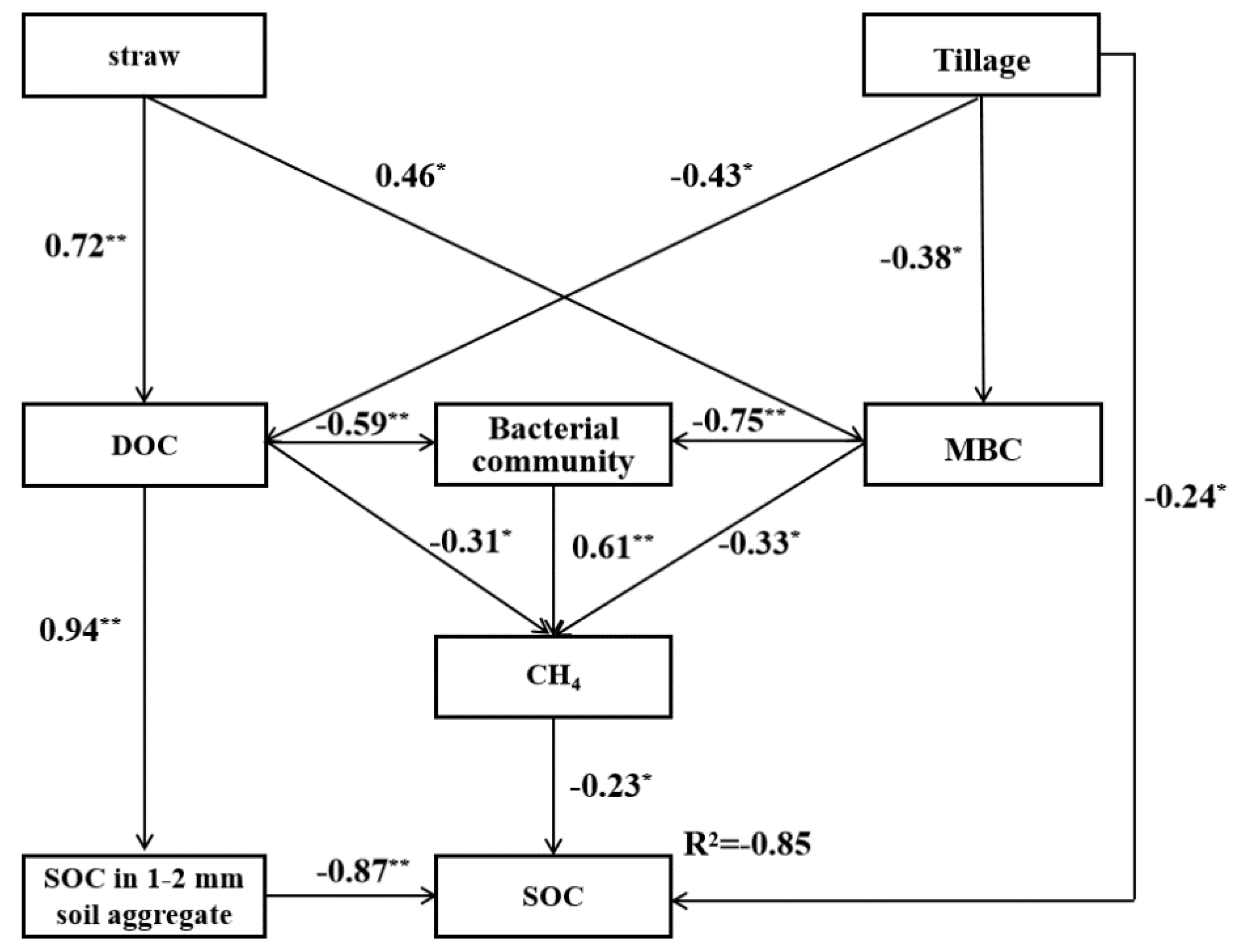
4.5. Relationships between Soil Organic Carbon and Bacterial Community under Different Tillage and Straw Return Practices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fiorini, A.; Boselli, R.; Maris, S.C.; Santelli, S.; Perego, A.; Acutis, M.; Brenna, S.; Tabaglio, V. Soil type and cropping system as drivers of soil quality indicators response to no-till: A 7-year field study. Appl. Soil Ecol. 2020, 155, 103646. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, L.; Liu, L.; Sheng, F.; Cao, C.; Li, C. Effects of long-term no tillage and straw return on greenhouse gas emissions and crop yields from a rice-wheat system in central China. Agric. Ecosyst. Environ. 2021, 322, 107650. [Google Scholar] [CrossRef]
- Berhane, M.; Xu, M.; Liang, Z.; Shi, J.; Wei, G.; Tian, X. Effects of long-term straw return on soil organic carbon storage and sequestration rate in North China upland crops: A meta-analysis. Glob. Chang. Biol. 2020, 26, 2686–2701. [Google Scholar] [CrossRef] [PubMed]
- Zeraatpisheh, M.; Ayoubi, S.; Mirbagheri, Z.; Mosaddeghi, M.R.; Xu, M. Spatial prediction of soil aggregate stability and soil organic carbon in aggregate fractions using machine learning algorithms and environmental variables. Geoderma Reg. 2021, 27, e00440. [Google Scholar] [CrossRef]
- Ajami, M.; Heidari, A.; Khormali, F.; Gorji, M.; Ayoubi, S. Environmental factors controlling soil organic carbon storage in loess soils of a subhumid region, northern Iran. Geoderma 2016, 281, 1–10. [Google Scholar] [CrossRef]
- Christopher, S.F.; Lal, R.; Mishra, U. Regional Study of No-Till Effects on Carbon Sequestration in the Midwestern United States. Soil Sci. Soc. Am. J. 2009, 73, 207–216. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K. Aggregate-associated soil organic matter as an ecosystem property and a measurement tool. Soil Biol. Biochem. 2014, 68, A4–A9. [Google Scholar] [CrossRef]
- Wu, Q.-X.; Du, B.; Jiang, S.-C.; Zhang, H.-W.; Zhu, J.-Q. Side Deep Fertilizing of Machine-Transplanted Rice to Guarantee Rice Yield in Conservation Tillage. Agriculture 2022, 12, 528. [Google Scholar] [CrossRef]
- Akhtar, K.; Wang, W.; Ren, G.; Khan, A.; Feng, Y.; Yang, G. Changes in soil enzymes, soil properties, and maize crop productivity under wheat straw mulching in Guanzhong, China. Soil Tillage Res. 2018, 182, 94–102. [Google Scholar] [CrossRef]
- Modak, K.; Biswas, D.R.; Ghosh, A.; Pramanik, P.; Das, T.K.; Das, S.; Kumar, S.; Krishnan, P.; Bhattacharyya, R. Zero tillage and residue retention impact on soil aggregation and carbon stabilization within aggregates in subtropical India. Soil Tillage Res. 2020, 202, 104649. [Google Scholar] [CrossRef]
- Mathew, R.P.; Feng, Y.; Githinji, L.; Ankumah, R.; Balkcom, K.S. Impact of No-Tillage and Conventional Tillage Systems on Soil Microbial Communities. Appl. Environ. Soil Sci. 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Jin, V.L.; Schmer, M.R.; Stewart, C.E.; Sindelar, A.J.; Varvel, G.E.; Wienhold, B. Long-term no-till and stover retention each decrease the global warming potential of irrigated continuous corn. Glob. Chang. Biol. 2017, 23, 2848–2862. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, A.; Shahbaz, M.; Farooq, T.H.; Sahar, N.E.; Shahzad, S.M.; Altaf, M.M.; Ashraf, M. A global meta-analysis of greenhouse gases emission and crop yield under no-tillage as compared to conventional tillage. Sci. Total Environ. 2021, 750, 142299. [Google Scholar] [CrossRef]
- Li, G.; Bian, S.; Lu, X.; Yang, T.; Lei, G.H. Field test on extruding soil caused of PHC pipe pile driving by static pressure for improving soft foundation of widened embankment. Rock Soil Mech. 2013, 34, 1089–1096. [Google Scholar]
- Kan, Z.-R.; Liu, Q.-Y.; Virk, A.L.; He, C.; Qi, J.-Y.; Dang, Y.P.; Zhao, X.; Zhang, H.-L. Effects of experiment duration on carbon mineralization and accumulation under no-till. Soil Tillage Res. 2021, 209, 104939. [Google Scholar] [CrossRef]
- Bandyopadhyay, P.K.; Saha, S.; Mani, P.K.; Mandal, B. Effect of organic inputs on aggregate associated organic carbon concentration under long-term rice–wheat cropping system. Geoderma 2010, 154, 379–386. [Google Scholar] [CrossRef]
- Liu, C.; Lu, M.; Cui, J.; Li, B.; Fang, C. Effects of Straw Carbon Input on Carbon Dynamics in Agricultural Soils: A Meta-Analysis. Glob. Chang. Biol. 2014, 20, 1366–1381. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, C.; Merino, A.; Osorio-Hernández, V.; Etchevers, J.D.; Figueroa, B.; Limon-Ortega, A.; Aguirre, E. Physical and chemical processes determining soil organic matter dynamics in a managed vertisol in a tropical dryland area. Soil Tillage Res. 2019, 194, 104348. [Google Scholar] [CrossRef]
- Regina, K.; Alakukku, L. Greenhouse gas fluxes in varying soils types under conventional and no-tillage practices. Soil Tillage Res. 2010, 109, 144–152. [Google Scholar] [CrossRef]
- Ruan, L.; Philip Robertson, G. Initial nitrous oxide, carbon dioxide, and methane costs of converting conservation reserve program grassland to row crops under no-till vs. conventional tillage. Glob. Chang. Biol. 2013, 19, 2478–2489. [Google Scholar] [CrossRef]
- Guo, L.-J.; Lin, S.; Liu, T.-Q.; Cao, C.-G.; Li, C.-F. Effects of Conservation Tillage on Topsoil Microbial Metabolic Characteristics and Organic Carbon within Aggregates under a Rice (Oryza sativa L.)–Wheat (Triticum aestivum L.) Cropping System in Central China. PLoS ONE 2016, 11, e0146145. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Virk, A.L.; Ma, S.-T.; Kan, Z.-R.; Qi, J.-Y.; Pu, C.; Yang, X.-G.; Zhang, H.-L. Dynamics in soil organic carbon of wheat-maize dominant cropping system in the North China Plain under tillage and residue management. J. Environ. Manag. 2020, 265, 110549. [Google Scholar] [CrossRef] [PubMed]
- Ussiri, D.A.; Lal, R.; Jarecki, M.K. Nitrous oxide and methane emissions from long-term tillage under a continuous corn cropping system in Ohio. Soil Tillage Res. 2009, 104, 247–255. [Google Scholar] [CrossRef]
- Pareja-Sánchez, E.; Cantero-Martínez, C.; Álvaro-Fuentes, J.; Plaza-Bonilla, D. Tillage and nitrogen fertilization in irrigated maize: Key practices to reduce soil CO2 and CH4 emissions. Soil Tillage Res. 2019, 191, 29–36. [Google Scholar] [CrossRef]
- Duan, M.; Long, Y.; Fan, H.; Ma, L.; Han, S.; Li, S.; Wei, B.; Wang, L. Fenlong-Ridging Promotes Microbial Activity in Sugarcane: A Soil and Root Metabarcoding Survey. Agriculture 2022, 12, 244. [Google Scholar] [CrossRef]
- Gartzia-Bengoetxea, N.; González-Arias, A.; Merino, A.; de Arano, I.M. Soil organic matter in soil physical fractions in adjacent semi-natural and cultivated stands in temperate Atlantic forests. Soil Biol. Biochem. 2009, 41, 1674–1683. [Google Scholar] [CrossRef]
- Jiang, P.; Xu, Q.; Xu, Z.; Cao, Z. Seasonal changes in soil labile organic carbon pools within a Phyllostachys praecox stand under high rate fertilization and winter mulch in subtropical China. For. Ecol. Manag. 2006, 236, 30–36. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Phospholipid Fatty Acid Composition, Biomass, and Activity of Microbial Communities from Two Soil Types Experimentally Exposed to Different Heavy Metals. Appl. Environ. Microbiol. 1993, 59, 3605–3617. [Google Scholar] [CrossRef]
- Neue, H.U.; Sass, R.L. Global Measurement Standardization of Methane Emissions from Irrigated Rice Cultivation: A Report of the Rice Cultivation and Trace Gas Exchange Activity (RICE) of the International Global Atmospheric Chemistry (IGAC) Project; Massachusetts Institute of Technology: Cambridge, MA, USA, 1994; pp. 1–6. Available online: http://www.worldcat.org/title/197230180?__cf_chl_tk=DkYxejo5hvH01DTFE3XL_W_Aq7RcOSe3nCs2nFFzE9E-1664118918-0-gaNycGzNBqU (accessed on 10 August 2022).
- Buendia, L.; Neue, H.; Wassmann, R.; Lantin, R.; Javellana, A.; Arah, J.; Wang, Z.; Wanfang, L.; Makarim, A.; Corton, T.; et al. An efficient sampling strategy for estimating methane emission from rice field. Chemosphere 1998, 36, 395–407. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Z.; Guo, L.; Cai, M.; Cao, C. Emissions of CH4 and CO2 from double rice cropping systems under varying tillage and seeding methods. Atmos. Environ. 2013, 80, 438–444. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, L.; Liu, T.; Li, C.; Cao, C. Effects of tillage practices and straw returning methods on greenhouse gas emissions and net ecosystem economic budget in rice–wheat cropping systems in central China. Atmos. Environ. 2015, 122, 636–644. [Google Scholar] [CrossRef]
- Zhao, J.; Carmody, L.A.; Kalikin, L.M.; Li, J.; Petrosino, J.F.; Schloss, P.; Young, V.; Lipuma, J.J. Impact of Enhanced Staphylococcus DNA Extraction on Microbial Community Measures in Cystic Fibrosis Sputum. PLoS ONE 2012, 7, e33127. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows integration and analysis of high-7 throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Hamady, M.; Lauber, C.L.; Knight, R. The influence of sex, handedness, and washing on the diversity of hand surface bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 17994–17999. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M.A.S.S. The vegan package. Community Ecol. Package 2007, 10, 719. [Google Scholar]
- Grace, J.B.; Michael Anderson, T.; Smith, M.D.; Seabloom, E.; Andelman, S.J.; Meche, G.; Weiher, E.; Allain, L.K.; Jutila, H.; Sankaran, M.; et al. Does species diversity limit productivity in natural grassland communities? Ecol. Lett. 2007, 10, 680–689. [Google Scholar] [CrossRef]
- Borie, F.; Rubio, R.; Rouanet, J.; Morales, A.; Borie, G.; Rojas, C. Effects of tillage systems on soil characteristics, glomalin and mycorrhizal propagules in a Chilean Ultisol. Soil Tillage Res. 2006, 88, 253–261. [Google Scholar] [CrossRef]
- Biswakarma, N.; Pooniya, V.; Zhiipao, R.; Kumar, D.; Verma, A.; Shivay, Y.; Lama, A.; Choudhary, A.; Meena, M.; Bana, R.J.A.; et al. Five years integrated crop management in direct seeded rice–zero till wheat rotation of north-western India: Effects on soil carbon dynamics, crop yields, water productivity and economic profitability. Agric. Ecosyst. Environ. 2021, 318, 107492. [Google Scholar] [CrossRef]
- Balota, E.L.; Colozzi Filho, A.; Andrade, D.S.; Dick, R.P. Long-term tillage and crop rotation effects on microbial biomass and C and N mineralization in a Brazilian Oxisol. Soil Tillage Res. 2004, 77, 137–145. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Jeffries, T.C.; Trivedi, C.; Anderson, I.C.; Lai, K.; McNee, M.; Flower, K.; Pal Singh, B.; Minkey, D.; et al. Soil aggregation and associated microbial communities modify the impact of agricultural management on carbon content. Environ. Microbiol. 2017, 19, 3070–3086. [Google Scholar] [CrossRef]
- Jia, S.; Liang, A.; Zhang, S.; Chen, X.; McLaughlin, N.B.; Sun, B.; Zhang, X.; Wu, D. Effect of tillage system on soil CO2 flux, soil microbial community and maize (Zea mays L.) yield. Geoderma 2021, 384, 114813. [Google Scholar] [CrossRef]
- Kan, Z.-R.; He, C.; Liu, Q.-Y.; Liu, B.-Y.; Virk, A.L.; Qi, J.-Y.; Zhao, X.; Zhang, H.-L. Carbon mineralization and its temperature sensitivity under no-till and straw returning in a wheat-maize cropping system. Geoderma 2020, 377, 114610. [Google Scholar] [CrossRef]
- Kan, Z.-R.; Ma, S.-T.; Liu, Q.-Y.; Liu, B.-Y.; Virk, A.L.; Qi, J.-Y.; Zhao, X.; Lal, R.; Zhang, H.-L. Carbon sequestration and mineralization in soil aggregates under long-term conservation tillage in the North China Plain. CATENA 2020, 188, 104428. [Google Scholar] [CrossRef]
- Emde, D.; Hannam, K.D.; Most, I.; Nelson, L.M.; Jones, M.D. Soil organic carbon in irrigated agricultural systems: A meta-analysis. Glob. Chang. Biol. 2021, 27, 3898–3910. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Wu, D.; Bol, R.; Wu, W.; Meng, F. Conservation farming practices in winter wheat–summer maize cropping reduce GHG emissions and maintain high yields. Agric. Ecosyst. Environ. 2019, 272, 266–275. [Google Scholar] [CrossRef]
- Ma, J.; Xu, H.; Yagi, K.; Cai, Z. Methane emission from paddy soils as affected by wheat straw returning mode. Plant Soil 2008, 313, 167–174. [Google Scholar] [CrossRef]
- Akter, M.; Deroo, H.; Kamal, A.M.; Kader, M.A.; Verhoeven, E.; Decock, C.; Boeckx, P.; Sleutel, S. Impact of irrigation management on paddy soil N supply and depth distribution of abiotic drivers. Agric. Ecosyst. Environ. 2018, 261, 12–24. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, S.; Li, Y.; Riaz, L.; Yu, F.; Yang, Q.; Han, S.; Ma, J. Effects and mechanisms of land-types conversion on greenhouse gas emissions in the Yellow River floodplain wetland. Sci. Total Environ. 2022, 813, 152406. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Dowd, S.; Sun, Y.; Allen, V. Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol. Biochem. 2008, 40, 2762–2770. [Google Scholar] [CrossRef]
- Ondreičková, K.; Piliarová, M.; Bušo, R.; Hašana, R.; Schreiber, L.; Gubiš, J.; Kraic, J. The Structure and Diversity of Bacterial Communities in Differently Managed Soils Studied by Molecular Fingerprinting Methods. Sustainability 2018, 10, 1095. [Google Scholar] [CrossRef]
- Lewin, G.R.; Carlos, C.; Chevrette, M.G.; Horn, H.A.; McDonald, B.R.; Stankey, R.J.; Fox, B.G.; Currie, C.R. Evolution and ecology of Actinobacteria and their bioenergy applications. Annu. Rev. Microbiol. 2016, 70, 235. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. Organic Amendments: Microbial Community Structure, Activity and Abundance of Agriculturally Relevant Microbes Are Driven by Long-Term Fertilization Strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Peng, H.; Ji, X.; Li, C.; Li, S. Effects of reduced inorganic fertilization and rice straw recovery on soil enzyme activities and bacterial community in double-rice paddy soils. Eur. J. Soil Biol. 2019, 94, 103116. [Google Scholar] [CrossRef]
- Luo, Y.; Iqbal, A.; He, L.; Zhao, Q.; Wei, S.; Ali, I.; Ullah, S.; Yan, B.; Jiang, L. Long-Term No-Tillage and Straw Retention Management Enhances Soil Bacterial Community Diversity and Soil Properties in Southern China. Agronomy 2020, 10, 1233. [Google Scholar] [CrossRef]
- Miura, T.; Niswati, A.; Swibawa, I.G.; Haryani, S.; Gunito, H.; Arai, M.; Yamada, K.; Shimano, S.; Kaneko, N.; Fujie, K. Shifts in the composition and potential functions of soil microbial communities responding to a no-tillage practice and bagasse mulching on a sugarcane plantation. Biol. Fertil. Soils 2016, 52, 307–322. [Google Scholar] [CrossRef]
- Yu, C.; Hu, X.; Deng, W.; Li, Y.; Han, G.; Xiong, C. Response of Bacteria Community to Long-Term Inorganic Nitrogen Application in Mulberry Field Soil. PLoS ONE 2016, 11, e0168152. [Google Scholar] [CrossRef]
- Dong, W.; Liu, E.; Yan, C.; Tian, J.; Zhang, H.; Zhang, Y. Impact of no tillage vs. conventional tillage on the soil bacterial community structure in a winter wheat cropping succession in northern China. Eur. J. Soil Biol. 2017, 80, 35–42. [Google Scholar] [CrossRef]
- Sun, M.; Chen, B.; Wang, H.; Wang, N.; Ma, T.; Cui, Y.; Luan, T.; Chun, S.; Liu, C.; Wang, L. Microbial Interactions and Roles in Soil Fertility in Seasonal Freeze-Thaw Periods under Different Straw Returning Strategies. Agriculture 2021, 11, 779. [Google Scholar] [CrossRef]
- Yamada, T.; Imachi, H.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y.; Sekiguchi, Y. Bellilinea caldifistulae gen. nov., sp. nov. and Longilinea arvoryzae gen. nov., sp. nov., strictly anaerobic, filamentous bacteria of the phylum Chloroflexi isolated from methanogenic propionate-degrading consortia. Int. J. Syst. Evol. Microbiol. 2007, 57, 2299–2306. [Google Scholar] [CrossRef]
- Xia, L.; Lam, S.K.; Wolf, B.; Kiese, R.; Chen, D.; Butterbach-Bahl, K. Trade-offs between soil carbon sequestration and reactive nitrogen losses under straw return in global agroecosystems. Glob. Chang. Biol. 2018, 24, 5919–5932. [Google Scholar] [CrossRef]
- Arunrat, N.; Sansupa, C.; Kongsurakan, P.; Sereenonchai, S.; Hatano, R. Soil Microbial Diversity and Community Composition in Rice–Fish Co-Culture and Rice Monoculture Farming System. Biology 2022, 11, 1242. [Google Scholar] [CrossRef]
- Rezgui, C.; Trinsoutrot-Gattin, I.; Benoit, M.; Laval, K.; Wassila, R.A. Linking changes in the soil microbial community to C and N dynamics during crop residue decomposition. J. Integr. Agric. 2021, 20, 3039–3059. [Google Scholar] [CrossRef]
- Guo, L.-J.; Zhang, Z.-S.; Wang, D.-D.; Li, C.-F.; Cao, C.-G. Effects of short-term conservation management practices on soil organic carbon fractions and microbial community composition under a rice-wheat rotation system. Biol. Fertil. Soils 2015, 51, 65–75. [Google Scholar] [CrossRef]
- Dubey, S.K.; Singh, A.; Singh, R.; Upadhyay, S. Changes in methanogenic population size and CH4 production potential in response to crop phenology in tropical rice field. Soil Biol. Biochem. 2013, 57, 972–978. [Google Scholar] [CrossRef]
- Fan, X.; Yu, H.; Wu, Q.; Ma, J.; Xu, H.; Yang, J.; Zhuang, Y. Effects of fertilization on microbial abundance and emissions of greenhouse gases (CH4 and N2O) in rice paddy fields. Ecol. Evol. 2016, 6, 1054–1063. [Google Scholar] [CrossRef]
- Glissmann, K.; Weber, S.; Conrad, R. Localization of processes involved in methanogenic degradation of rice straw in anoxic paddy soil. Environ. Microbiol. 2001, 3, 502–511. [Google Scholar] [CrossRef]
- Ortiz-Cornejo, N.L.; Romero-Salas, E.A.; Navarro-Noya, Y.E.; González-Zúñiga, J.C.; Ramirez-Villanueva, D.A.; Vásquez-Murrieta, M.S.; Verhulst, N.; Govaerts, B.; Dendooven, L.; Luna-Guido, M. Incorporation of bean plant residue in soil with different agricultural practices and its effect on the soil bacteria. Appl. Soil Ecol. 2017, 119, 417–427. [Google Scholar] [CrossRef]
- Conrad, R. Microbial ecology of methanogens and methanotrophs. Adv. Agron. 2007, 96, 1–63. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop Season | Soil Aggregate Fraction | CTNS | CTSR | NTNS | NTSR | T | SR | T × SR |
|---|---|---|---|---|---|---|---|---|
| 2012 rice season | 1–2 mm | 17.03 ± 1.05 a | 18.97 ± 0.60 a | 18.61 ± 2.39 a | 21.14 ± 1.70 a | ns | ns | ns |
| 0.25–1 mm | 15.60 ± 0.32 a | 17.07 ± 0.76 a | 14.16 ± 0.03 a | 16.94 ± 1.66 a | ns | ns | ns | |
| 0.053–0.25 mm | 19.67 ± 1.08 a | 16.49 ± 1.72 a | 20.62 ± 0.3 a | 16.85 ± 2.00 a | ns | ns | ns | |
| <0.053 mm | 17.97 ± 1.09 a | 19.49 ± 1.04 a | 18.72 ± 0.43 a | 18.00 ± 0.38 a | ns | ns | ns | |
| 2013 wheat season | 1–2 mm | 16.32 ± 1.00 c | 18.20 ± 0.29 bc | 18.92 ± 0.91 b | 21.45 ± 0.63 a | * | ns | ns |
| 0.25–1 mm | 17.52 ± 0.55 a | 18.52 ± 0.65 a | 19.77 ± 0.12 a | 21.53 ± 1.76 a | * | ns | ns | |
| 0.053–0.25 mm | 20.79 ± 1.62 a | 21.59 ± 0.98 a | 23.42 ± 1.91 a | 26.14 ± 3.05 a | ns | ns | ns | |
| <0.053 mm | 15.02 ± 1.34 a | 16.59 ± 1.54 a | 16.97 ± 1.75 a | 19.79 ± 1.45 a | ns | ns | ns | |
| 2013 rice season | 1–2 mm | 16.14 ± 0.67 c | 18.46 ± 0.28 bc | 18.88 ± 0.43 b | 22.60 ± 1.36 a | * | * | ns |
| 0.25–1 mm | 18.06 ± 0.43 c | 19.87 ± 0.18 b | 21.04 ± 0.46 ab | 21.98 ± 0.36 a | * | * | ns | |
| 0.053–0.25 mm | 16.90 ± 0.84 a | 16.93 ± 1.82 a | 18.23 ± 0.50 a | 19.65 ± 1.00 a | ns | ns | ns | |
| <0.053 mm | 15.51 ± 1.19 a | 16.92 ± 1.95 a | 20.18 ± 0.81 a | 18.81 ± 0.77 a | ns | ns | ns | |
| 2014 wheat season | 1–2 mm | 16.25 ± 0.12 c | 18.34 ± 0.18 b | 16.30 ± 0.07 c | 19.87 ± 0.34 a | ns | * | * |
| 0.25–1 mm | 20.66 ± 0.33 c | 21.89 ± 0.19 ab | 21.40 ± 0.55 bc | 22.78 ± 0.14 a | ns | * | ns | |
| 0.053–0.25 mm | 17.49 ± 0.88 a | 18.22 ± 0.82 a | 18.75 ± 0.85 a | 19.63 ± 1.37 a | ns | ns | ns | |
| <0.053 mm | 17.72 ± 4.40 a | 17.83 ± 2.02 a | 16.99 ± 3.11 a | 17.67 ± 1.63 a | ns | ns | ns |
| Treatment | 2012 | 2013 | 2014 | ||
|---|---|---|---|---|---|
| Rice Season | Wheat Season | Rice Season | Wheat Season | Rice Season | |
| CTNS | 400 ± 7.51 b | 4.86 ± 0.98 c | 475 ± 21.7 b | 5.39 ± 0.54 c | 167 ± 11.37 b |
| CTSR | 560 ± 30.73 a | 16.91 ± 0.37 a | 645 ± 12.0 a | 15.95 ± 0.99 a | 202 ± 13.68 a |
| NTNS | 391 ± 21.16 b | 3.99 ± 0.42 c | 445 ± 7.7 b | 4.81 ± 0.46 c | 140 ± 10.60 b |
| NTSR | 632 ± 27.09 a | 12.53 ± 2.23 b | 610 ± 9.7 a | 12.33 ± 0.60 b | 162 ± 2.35 b |
| T | ns | ns | ns | ns | * |
| SR | * | * | * | * | * |
| T × SR | ns | ns | ns | ns | ns |
| Treatment | 2012 | 2013 | 2014 | ||
|---|---|---|---|---|---|
| Rice Season | Wheat Season | Rice Season | Wheat Season | Rice Season | |
| CTNS | 2230 ± 92 d | 5282 ± 123 b | 4833 ± 397 b | 3982 ± 84 bc | 4283 ± 50 c |
| CTSR | 4914 ± 27 a | 6695 ± 408 ab | 6503 ± 308 a | 4989 ± 123 a | 6332 ± 154 a |
| NTNS | 2660 ± 86 c | 6090 ± 298 ab | 3734 ± 116 c | 3799 ± 124 c | 3817 ± 88 d |
| NTSR | 4438 ± 47 b | 7193 ± 939 a | 5557 ± 265 b | 4525 ± 452 ab | 4525 ± 162 b |
| T | ns | ns | ns | * | * |
| SR | * | * | * | * | * |
| T × SR | * | ns | ns | ns | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Shi, J.; Lin, W.; Liang, J.; Lu, Z.; Tang, X.; Liu, Y.; Wu, P.; Li, C. Soil Bacteria Mediate Soil Organic Carbon Sequestration under Different Tillage and Straw Management in Rice-Wheat Cropping Systems. Agriculture 2022, 12, 1552. https://doi.org/10.3390/agriculture12101552
Guo L, Shi J, Lin W, Liang J, Lu Z, Tang X, Liu Y, Wu P, Li C. Soil Bacteria Mediate Soil Organic Carbon Sequestration under Different Tillage and Straw Management in Rice-Wheat Cropping Systems. Agriculture. 2022; 12(10):1552. https://doi.org/10.3390/agriculture12101552
Chicago/Turabian StyleGuo, Lijin, Jie Shi, Wei Lin, Jincheng Liang, Zhenhua Lu, Xuexiao Tang, Yue Liu, Purui Wu, and Chengfang Li. 2022. "Soil Bacteria Mediate Soil Organic Carbon Sequestration under Different Tillage and Straw Management in Rice-Wheat Cropping Systems" Agriculture 12, no. 10: 1552. https://doi.org/10.3390/agriculture12101552