Effect of Ethylene-Insensitive Mutation etr2b on Postharvest Chilling Injury in Zucchini Fruit

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Growth Conditions
2.2. Genotyping etr2b Mutation Using TaqMan Probes
2.3. Ethylene Production and Respiration Rate
2.4. Weight Loss, Firmness, and Chilling Injury Index
2.5. MDA and H2O2 Contents
2.6. Statistical Analysis
3. Results
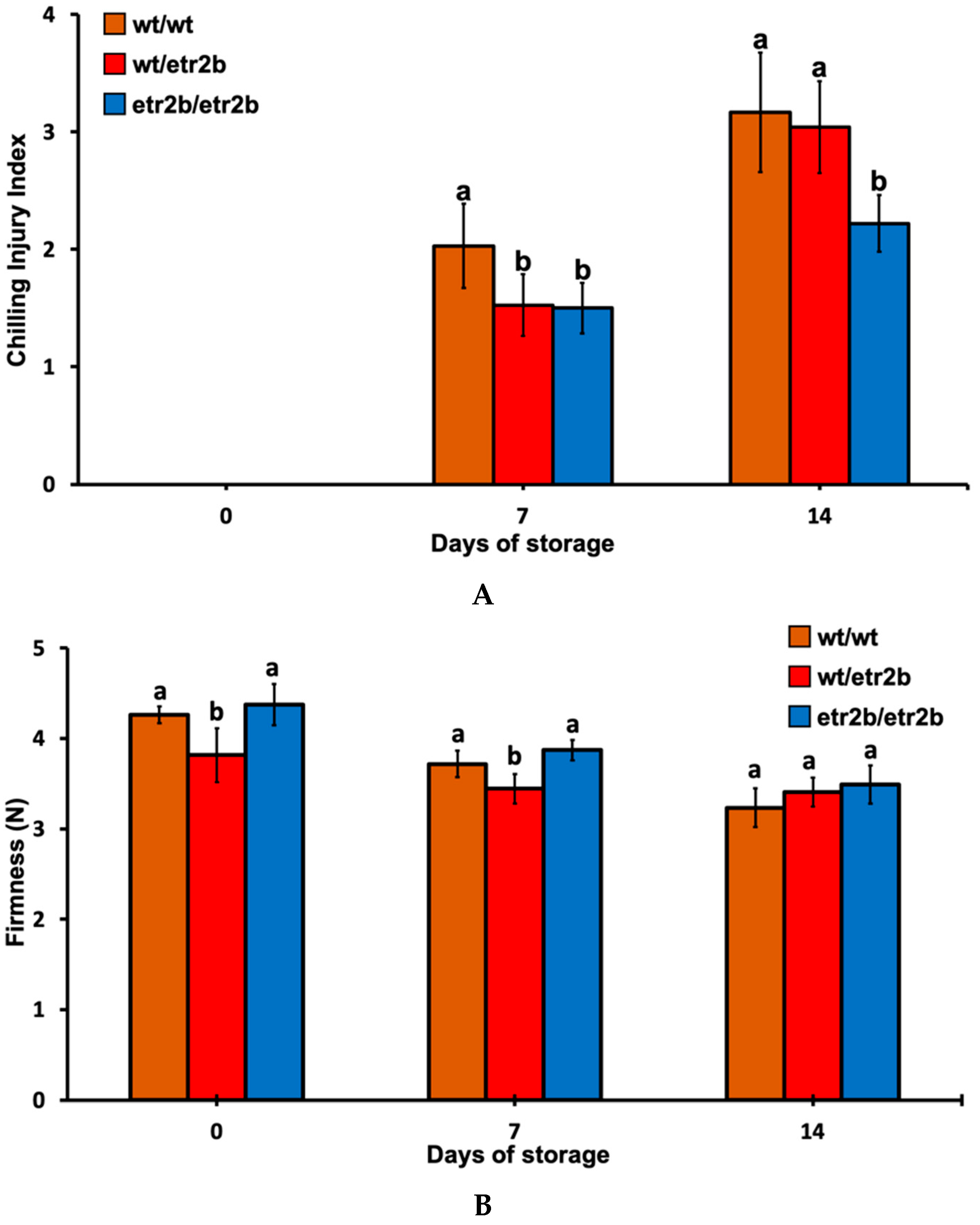
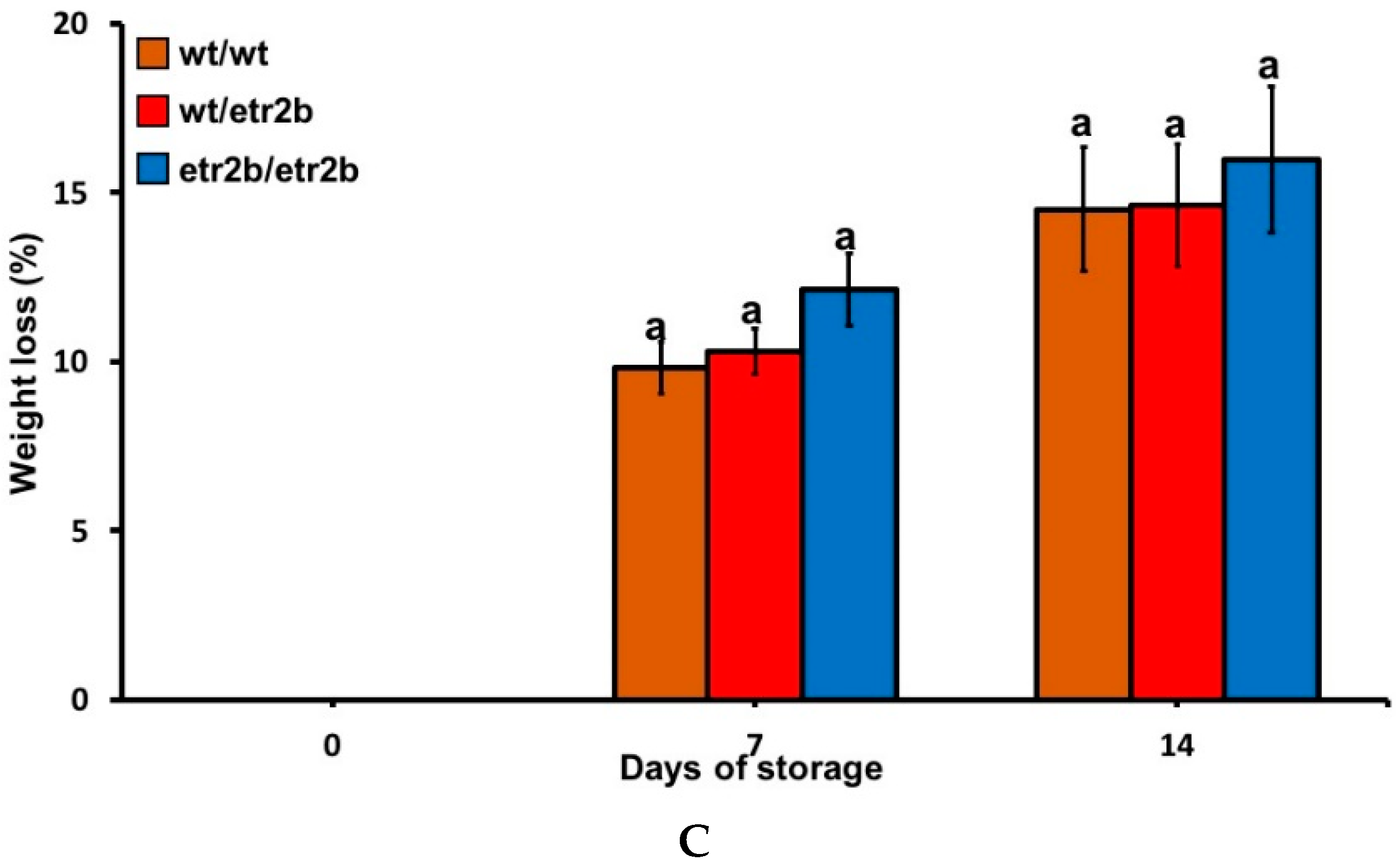
3.1. Response of Zucchini Ethylene-Insensitive Mutant Fruit to Cold Storage
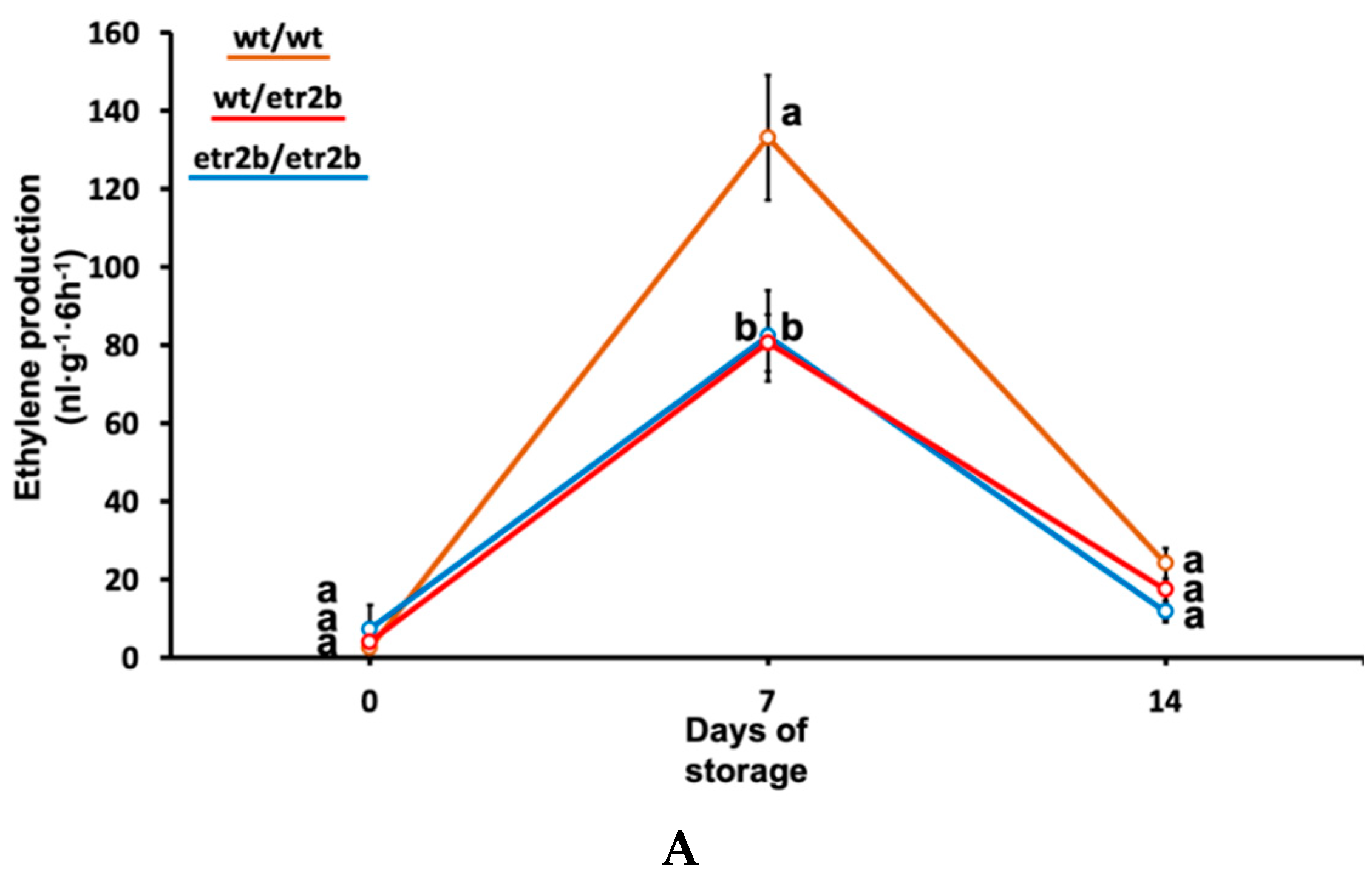
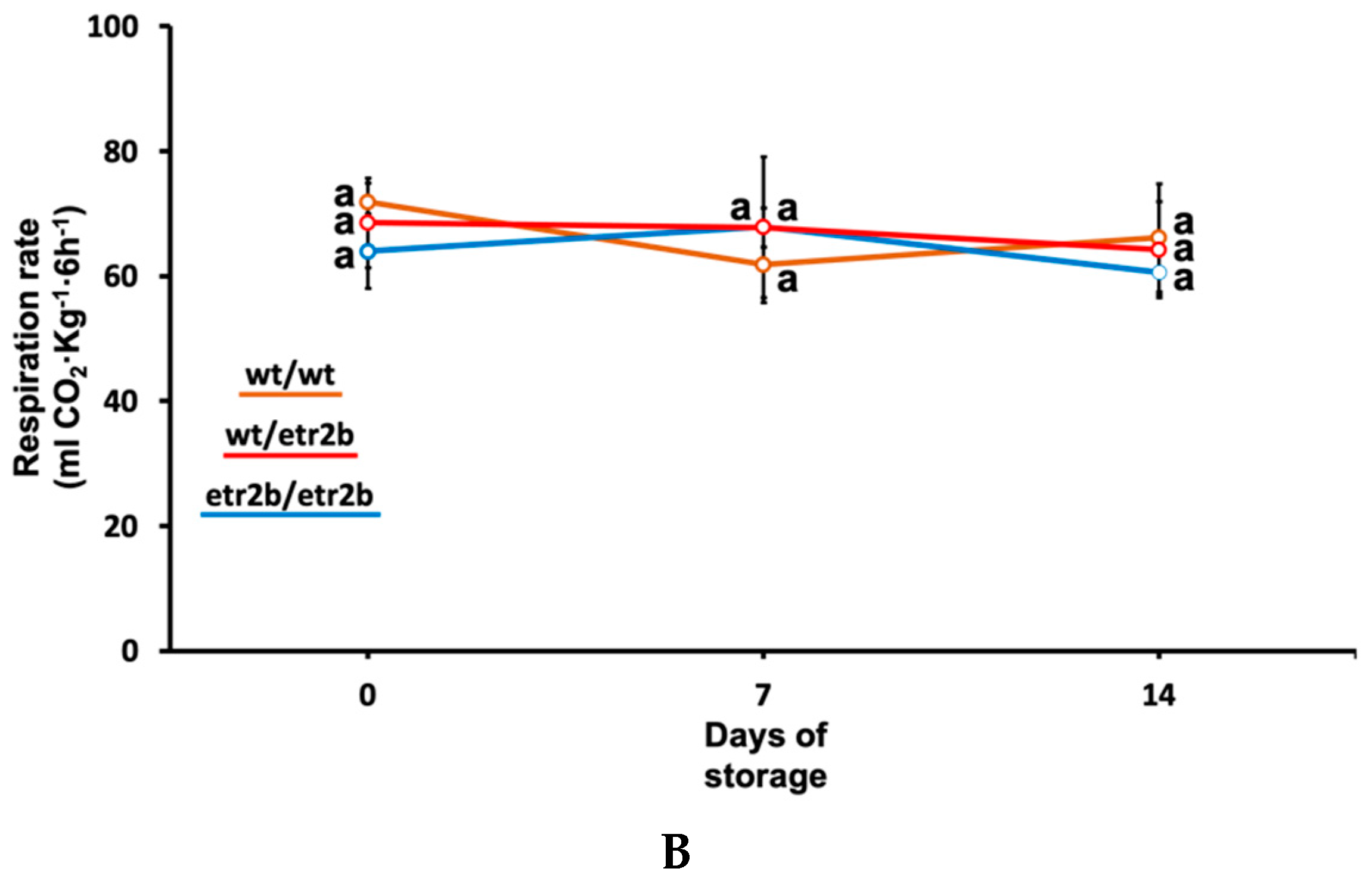
3.2. Effect of Ethylene-Insensitive Mutations on Ethylene and CO2 Production in Cold-Stored Zucchini Fruit
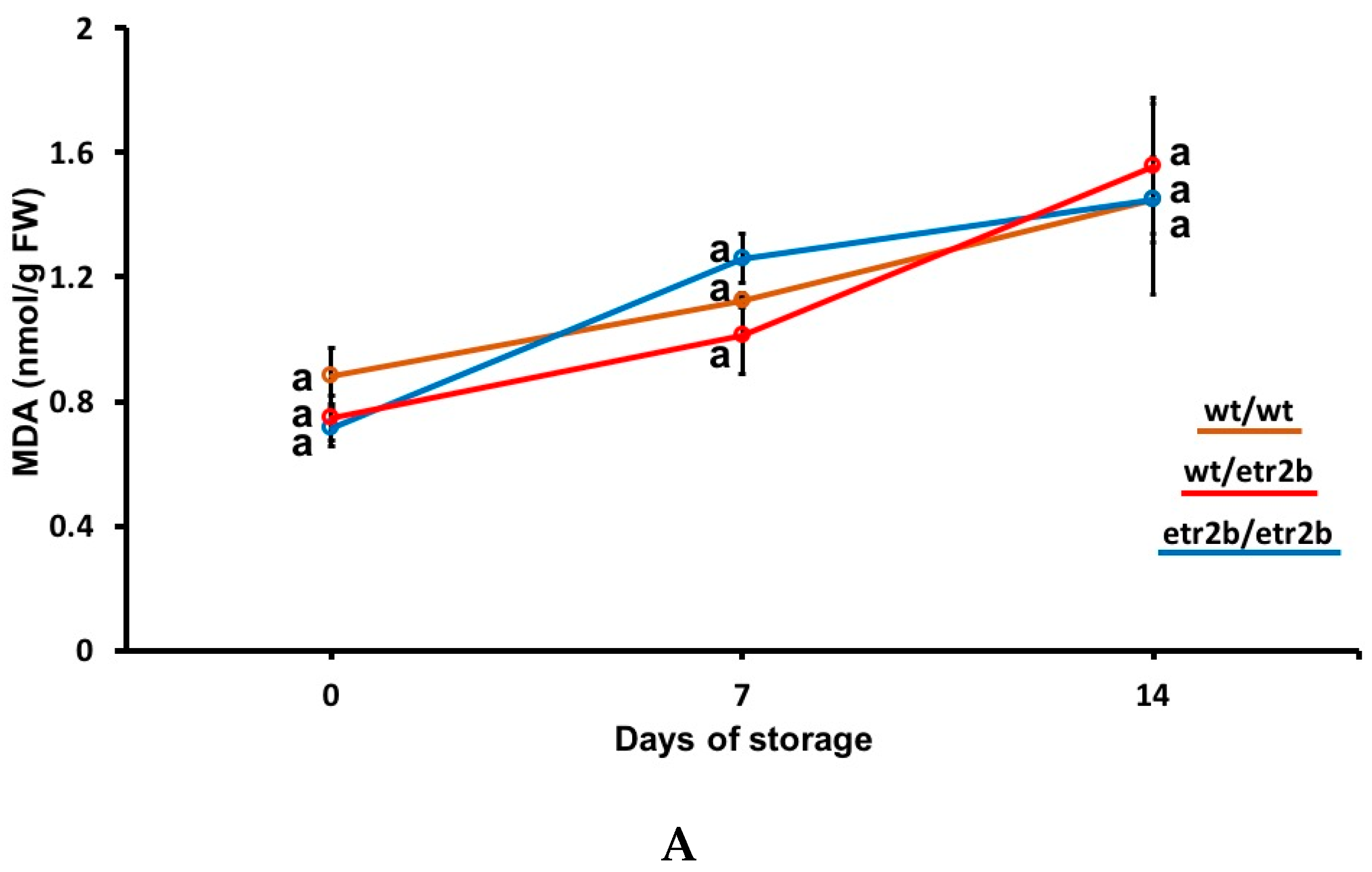
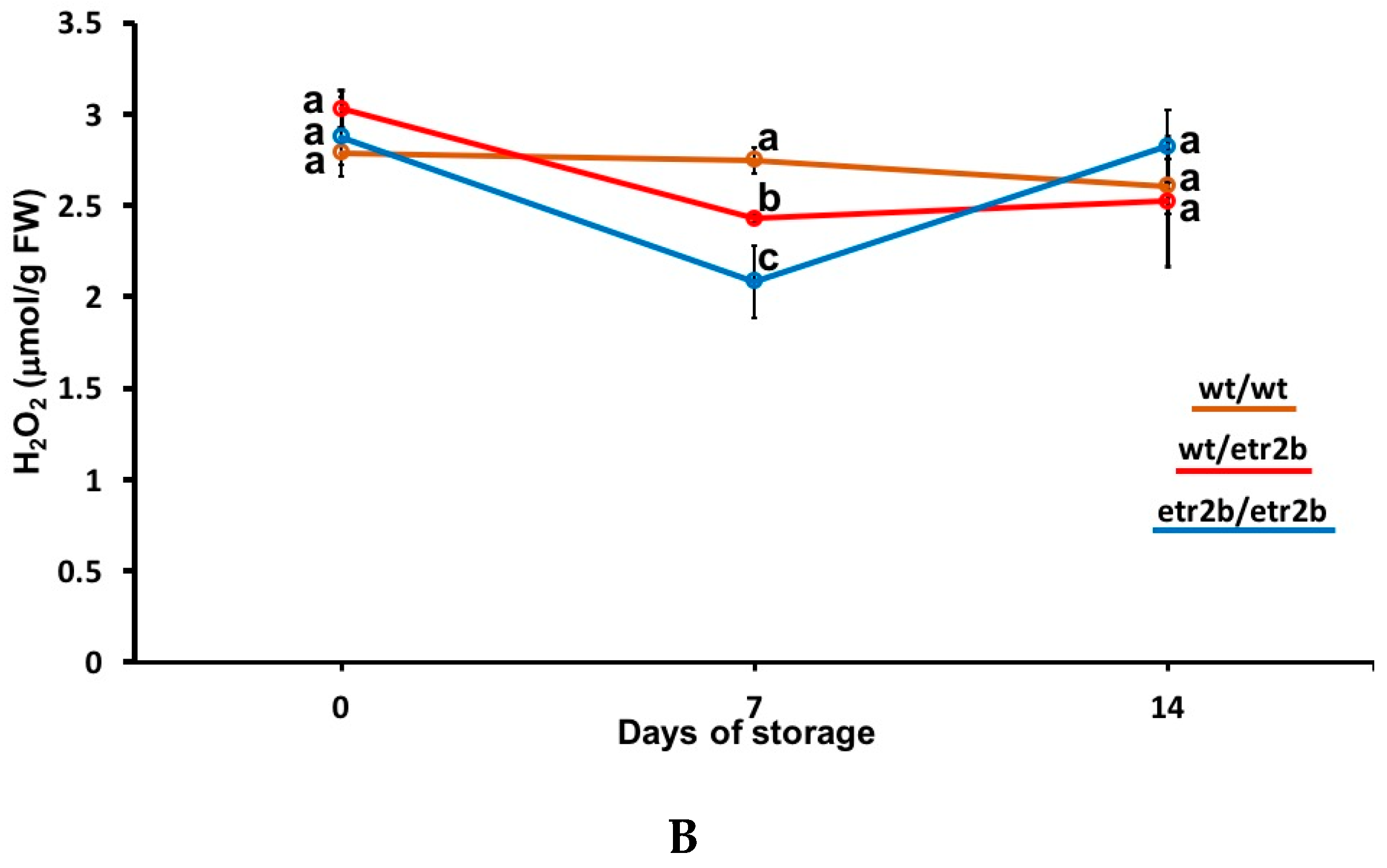
3.3. Effect of Ethylene-Insensitive Mutations on Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carvajal, F.; Martinez, C.; Jamilena, M.; Garrido, D. Differential response of zucchini varieties to low storage temperature. Sci. Hortic. 2011, 130, 90–96. [Google Scholar] [CrossRef]
- Megías, Z.; Martínez, C.; Manzano, S.; García, A.; Rebolloso-Fuentes, M.; Valenzuela, J.L.; Garrido DJamilena, M. Ethylene biosynthesis and signaling elements involved in chilling injury and other postharvest quality traits in the non-climacteric fruit of zucchini (Cucurbita pepo). Postharvest Biol. Technol. 2016, 113, 48–57. [Google Scholar] [CrossRef]
- Megías, Z.; González-Rodríguez, L.J.; Aguado, E.; Garcia, A.; Manzano, S.; Rebolloso, M.M.; Valenzuela, J.L.; Jamilena, M. Effect of cold storage time on chilling injury in two zucchini cultivars. Acta. Hortic. 2018, 1194, 479–486. [Google Scholar] [CrossRef]
- Megías, Z.; Martínez, C.; Manzano, S.; García, A.; Rebolloso-Fuentes, M.M.; Garrido, D.; Valenzuela, J.L.; Jamilena, M. Individual Shrink Wrapping of Zucchini Fruit Improves Postharvest Chilling Tolerance Associated with a Reduction in Ethylene Production and Oxidative Stress Metabolites. PLoS ONE 2015, 10, e0133058. [Google Scholar] [CrossRef] [PubMed]
- Megías, Z.; Manzano, S.; Martínez, C.; García, A.; Aguado, E.; Garrido, D.; Valenzuela, J.L.; Jamilena, M. Postharvest cold tolerance in summer squash and its association with reduced cold-induced ethylene production. Euphytica 2017, 213, 9. [Google Scholar] [CrossRef]
- Balandrán-Quintana, R.R.; Mendoza-Wilson, A.M.; Gardea-Béjar, A.A.; Vargas-Arispuro, I.; Martínez-Téllez, M.A. Irreversibility of chilling injury in zucchini squash (Cucurbita pepo L.) could be a programmed event long before the visible symptoms are evident. Biochem. Biophys. Res. Commun. 2003, 307, 553–557. [Google Scholar] [CrossRef]
- Megías, Z.; Manzano, S.; Martínez, C.; Valenzuela, J.L.; Garrido, D.; Jamilena, M. Ethylene production by fruits of zucchini cultivars differing in postharvest fruit quality and tolerance to chilling injury. In Proceedings of the 10th EUCARPIA Meet Genet Breed Cucurbitaceae, Antalya, Turkey, 15–18 October 2012; pp. 638–642. [Google Scholar]
- Valenzuela, J.; Manzano, S.; Palma, F.; Carvajal, F.; Garrido, D.; Jamilena, M. Oxidative Stress Associated with Chilling Injury in Immature Fruit: Postharvest Technological and Biotechnological Solutions. Int. J. Mol. Sci. 2017, 18, 1467. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y. Combined treatment of heat shock and low temperature conditioning reduces chilling injury in zucchini squash. Postharvest Biol. Technol. 1994, 4, 65–73. [Google Scholar] [CrossRef]
- Carvajal, F.; Palma, F.; Jamilena, M.; Garrido, D. Cell wall metabolism and chilling injury during postharvest cold storage in zucchini fruit. Postharvest Biol. Technol. 2015, 108, 68–77. [Google Scholar] [CrossRef]
- Megías, Z.; Barrera, A.; Manzano, S.; Martínez, C.; Garrido, D.; Valenzuela, J.L.; Jamilena, M. Physical and chemical treatments enhancing postharvest fruit quality in zucchini. Acta. Hortic. 2015, 1091, 141–146. [Google Scholar] [CrossRef]
- Salvador, A.; Arnal, L.; Monterde, A.; Cuquerella, J. Reduction of chilling injury symptoms in persimmon fruit cv. ‘Rojo Brillante’ by 1-MCP. Postharvest Biol. Technol. 2004, 33, 285–291. [Google Scholar] [CrossRef]
- Salvador, A.; Carvalho, C.P.; Monterde, A.; Martínez-Jávega, J.M. 1-MCP effect on chilling injury development in ‘Nova’ and ‘Ortanique’ mandarins. Food Sci. Technol. Int. 2006, 12, 165–170. [Google Scholar] [CrossRef]
- García, A.; Aguado, E.; Parra, G.; Manzano, S.; Martínez, C.; Megías, Z.; Cebrián, G.; Romero, J.; Beltrán, S.; Garrido, D. Jamilena. Phenomic and Genomic Characterization of a Mutant Platform in Cucurbita pepo. Front Plant Sci. 2018, 9, 1049. [Google Scholar] [CrossRef]
- García, A.; Aguado, E.; Martínez, C.; Loska, D.; Beltrán, S.; Valenzuela, J.L.; Garrido, D.; Jamilena, M. The ethylene receptors CpETR1A and CpETR2B cooperate in the control of sex determination in Cucurbita pepo. J. Exp. Bot. 2020, 71, 154–167. [Google Scholar] [CrossRef]
- Megías, Z.; Martínez, C.; Manzano, S.; Barrera, A.; Rosales, R.; Valenzuela, J.L.; Garrido DJamilena, M. Cold-induced ethylene in relation to chilling injury and chilling sensitivity in the non-climacteric fruit of zucchini (Cucurbita pepo L.). LWT-Food. Sci. Technol. 2014, 57, 194–199. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant. Cell. Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Lara, I.; Belge, B.; Goulao, L.F. The fruit cuticle as a modulator of postharvest quality. Postharvest Biol. Technol. 2014, 87, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Trujillo, J.P.; Martínez, J.A. Ultrastructure of the onset of chilling injury in cucumber fruit. J. Appl. Bot. Food Qual. 2006, 80, 100–110. [Google Scholar]
- Brummell, D.A.; Harpster, M.H. Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. In Plant Cell Wall; Carpita, N.C., Campbell, M., Tierney, M., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 311–340. [Google Scholar] [CrossRef]
- Guzmán, M.; Sánchez, A.; Salas, M.C.; Del Moral, F.; Valenzuela, J.L. Relationship between pectin-methyl esterase activity and softening in “Raf” tomato fruit. Acta. Hortic. 2012, 934, 1291–1296. [Google Scholar] [CrossRef]
- Rugkong, A.; Rose, J.K.; Lee, S.J.; Giovannoni, J.J.; O’Neill, M.A.; Watkins, C.B. Cell wall metabolism in cold-stored tomato fruit. Postharvest Biol. Technol. 2010, 57, 106–1013. [Google Scholar] [CrossRef]
- Wang, C.Y.; Adams, D.O. Chilling-Induced Ethylene Production in Cucumbers (Cucumis sativus L.). Plant Physiol. 1982, 69, 424–427. [Google Scholar] [CrossRef] [Green Version]
- Hyodo, H.; Fukasawa, R. Ethylene Production in Kiwifruit. J. Jpn. Soc. Hortic. Sci. 1985, 54, 209–215. [Google Scholar] [CrossRef] [Green Version]
- McCollum, T.G.; McDonald, R.E. Electrolyte leakage, respiration, and ethylene production as indices of chilling injury in grapefruit. HortScience 1991, 26, 1191–1192. [Google Scholar] [CrossRef] [Green Version]
- Lafuente, M.T.; Zacarias, L.; Martínez-Téllez, M.A.; Sánchez-Ballesta, M.T.; Dupille, E. Phenylalanine Ammonia-lyase As Related to Ethylene in the Development of Chilling Symptoms during Cold Storage of Citrus Fruits. J. Agric. Food Chem. 2001, 49, 6020–6025. [Google Scholar] [CrossRef]
- Pons, C.; Martí, C.; Forment, J.; Crisosto, C.H.; Dandekar, A.M.; Granell, A. A bulk segregant gene expression analysis of a peach population reveals components of the underlying mechanism of the fruit cold response. PLoS ONE 2014, 9, e90706. [Google Scholar] [CrossRef]
- Lado, J.; Rodrigo, M.J.; Zacarías, L. Analysis of ethylene biosynthesis and perception during postharvest cold storage of Marsh and Star Ruby grapefruits. Food Sci. Technol. Int. 2015, 21, 537–546. [Google Scholar] [CrossRef]
- Ben-Amor, M.; Flores, B.; Latché, A.; Bouzayen, M.; Pech, J.C.; Romojaro, F. Inhibition of ethylene biosynthesis by antisense ACC oxidase RNA prevents chilling injury in Charentais cantaloupe melons. Plant Cell Environ. 1999, 22, 1579–1586. [Google Scholar] [CrossRef]
- Obando, J.; Miranda, C.; Jowkar, M.M.; Moreno, E.; Sour, M.K.; Martínez, J.A.; Arús, P.; García-Mas, J.; Monforte, A.J.; Fernández-Trujillo, J.P. Creating climacteric melon fruit from nonclimacteric parentals: Postharvest quality implications. In Advances in Plant Ethylene Research; Ramina, A., Chang, C., Giovannoni, J., Klee, H., Perata, P., Woltering, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 197–205. [Google Scholar] [CrossRef]
- Lado, J.; Manzi, M.; Sainz, M.M.; Sotelo, M.; Zacarías, L. Involvement of Plant Hormones in Cold Stress Tolerance. In Plant Hormones under Challenging Environmental Factors; Ahammed, G., Yu, J.Q., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 23–49. [Google Scholar] [CrossRef]
- Huang, R.; Li, W.; Guan, X.-W.; Xie, B.; Zhang, S.-M. Molecular cloning and characterization of genes related to the ethylene signal transduction pathway in pomegranate (Punica granatum L.) under different temperature treatments. J. Biosci. 2019, 44, 137. [Google Scholar] [CrossRef]
- Martínez-Téllez, M.; Ramos-Clamont, M.; Gardea, A.; Vargas-Arispuro, I. Effect of infiltrated polyamines on polygalacturonase activity and chilling injury responses in zucchini squash (Cucurbita pepo L.). Biochem. Biophys. Res. Commun. 2002, 295, 98–101. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Ramos, J.M.; Jamilena, M.; Garrido, D. Effect of putrescine application on maintenance of zucchini fruit quality during cold storage: Contribution of GABA shunt and other related nitrogen metabolites. Postharvest Biol. Technol. 2015, 99, 131–140. [Google Scholar] [CrossRef]
- Cao, S.; Jiang, S.; Zhang, R. Evidence for a role of Ethylene-Insensitive 2 gene in the regulation of the oxidative stress response in Arabidopsis. Acta Physiol. Plant 2006, 28, 417–425. [Google Scholar] [CrossRef]
- Wang, H.; Wang, F.; Zheng, F.; Wang, L.; Pei, H.; Dong, C.-H. Ethylene-insensitive mutants of Nicotiana tabacum exhibit drought stress resistance. Plant Growth Regul. 2016, 79, 107–117. [Google Scholar] [CrossRef]
- Monteiro, C.C.; Carvalho, R.F.; Gratão, P.L.; Carvalho, G.; Tezotto, T.; Medici, L.O.; Peres, L.E.P.; Azevedo, R.A. Biochemical responses of the ethylene-insensitive Never ripe tomato mutant subjected to cadmium and sodium stresses. Environ. Exp. Bot. 2011, 71, 306–320. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García, A.; Aguado, E.; Cebrián, G.; Iglesias, J.; Romero, J.; Martínez, C.; Garrido, D.; Rebolloso, M.d.M.; Valenzuela, J.L.; Jamilena, M. Effect of Ethylene-Insensitive Mutation etr2b on Postharvest Chilling Injury in Zucchini Fruit. Agriculture 2020, 10, 532. https://doi.org/10.3390/agriculture10110532
García A, Aguado E, Cebrián G, Iglesias J, Romero J, Martínez C, Garrido D, Rebolloso MdM, Valenzuela JL, Jamilena M. Effect of Ethylene-Insensitive Mutation etr2b on Postharvest Chilling Injury in Zucchini Fruit. Agriculture. 2020; 10(11):532. https://doi.org/10.3390/agriculture10110532
Chicago/Turabian StyleGarcía, Alicia, Encarnación Aguado, Gustavo Cebrián, Jessica Iglesias, Jonathan Romero, Cecilia Martínez, Dolores Garrido, María del Mar Rebolloso, Juan Luis Valenzuela, and Manuel Jamilena. 2020. "Effect of Ethylene-Insensitive Mutation etr2b on Postharvest Chilling Injury in Zucchini Fruit" Agriculture 10, no. 11: 532. https://doi.org/10.3390/agriculture10110532