
Merkel Cell Carcinoma Display PIEZO2 Immunoreactivity

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
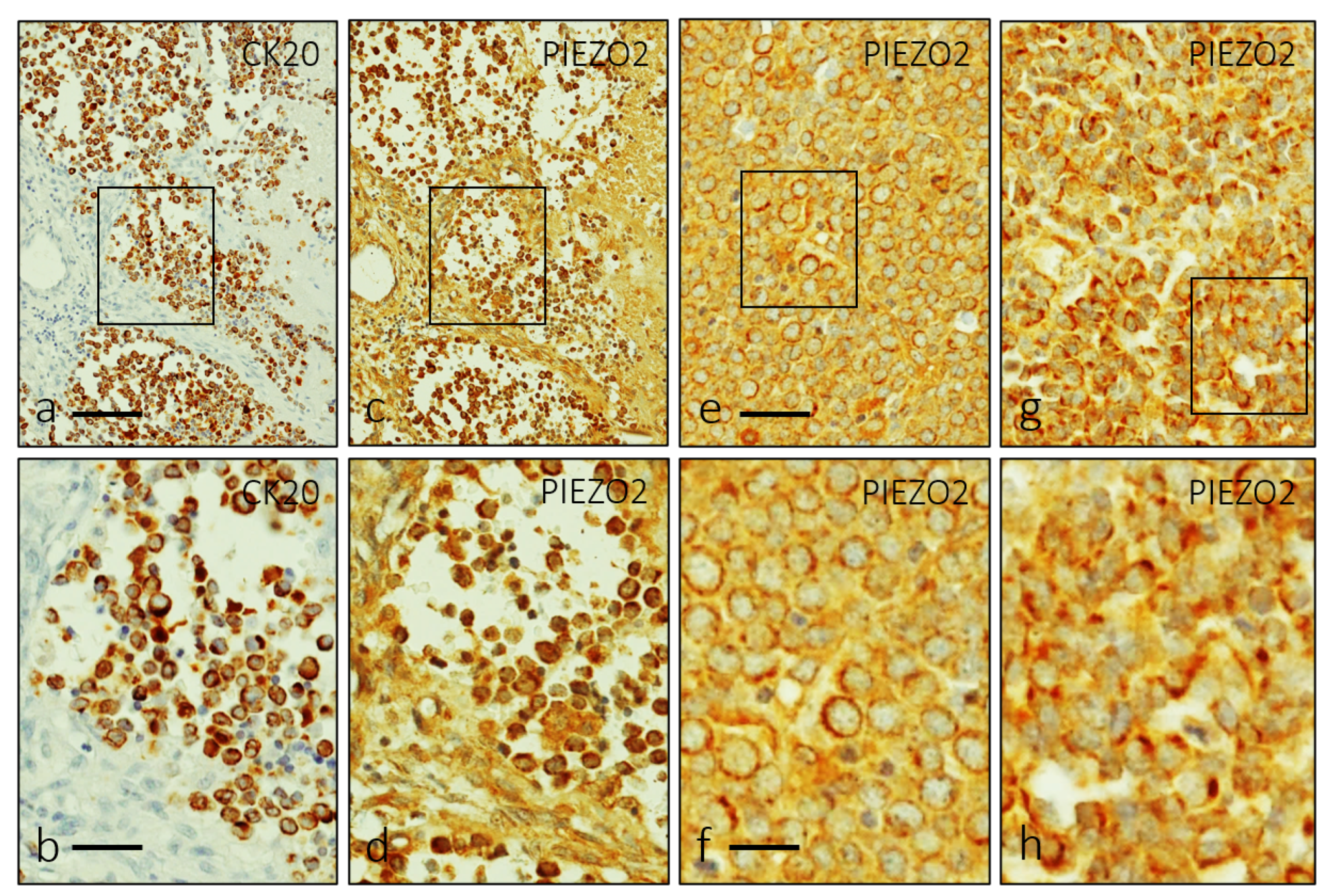
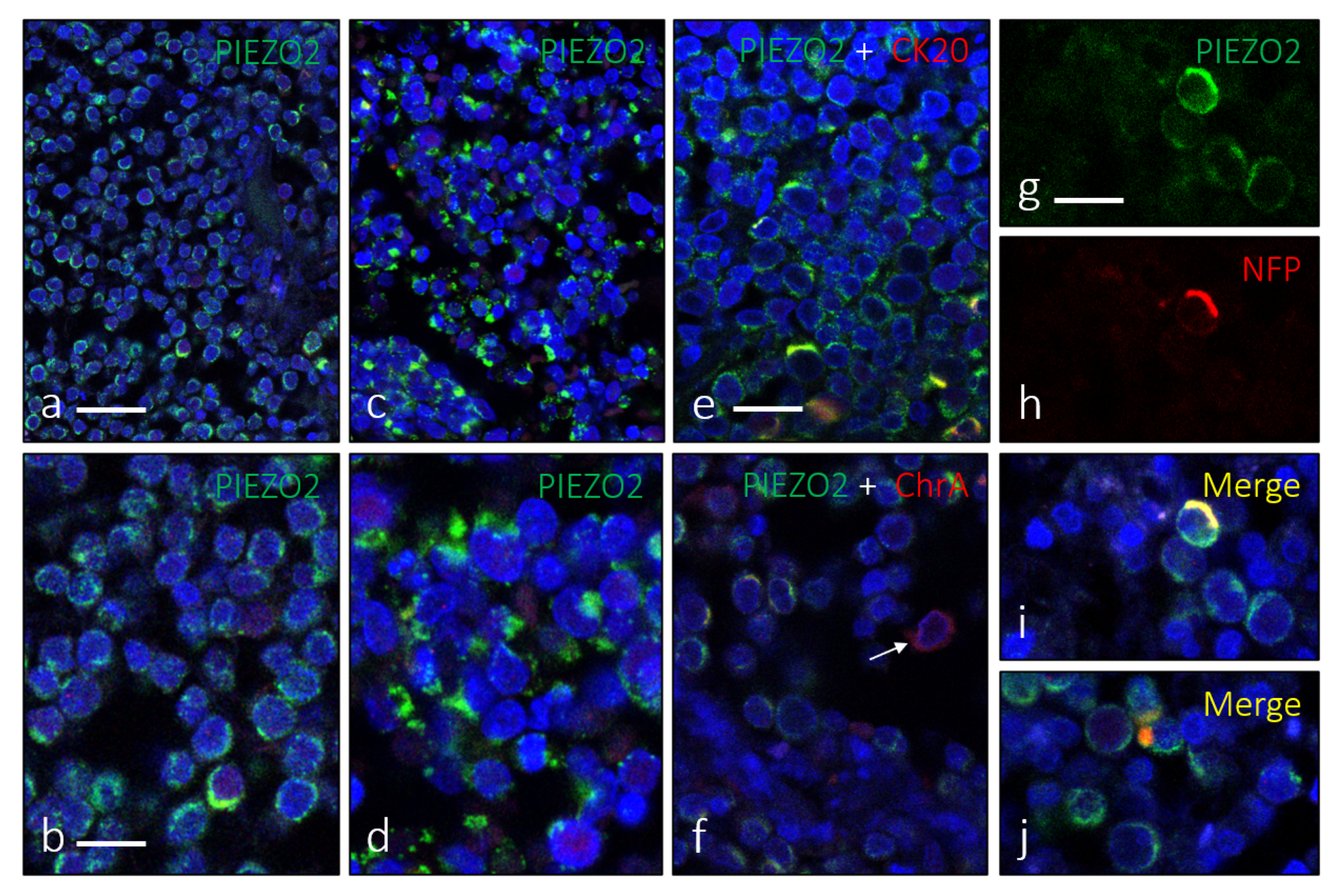
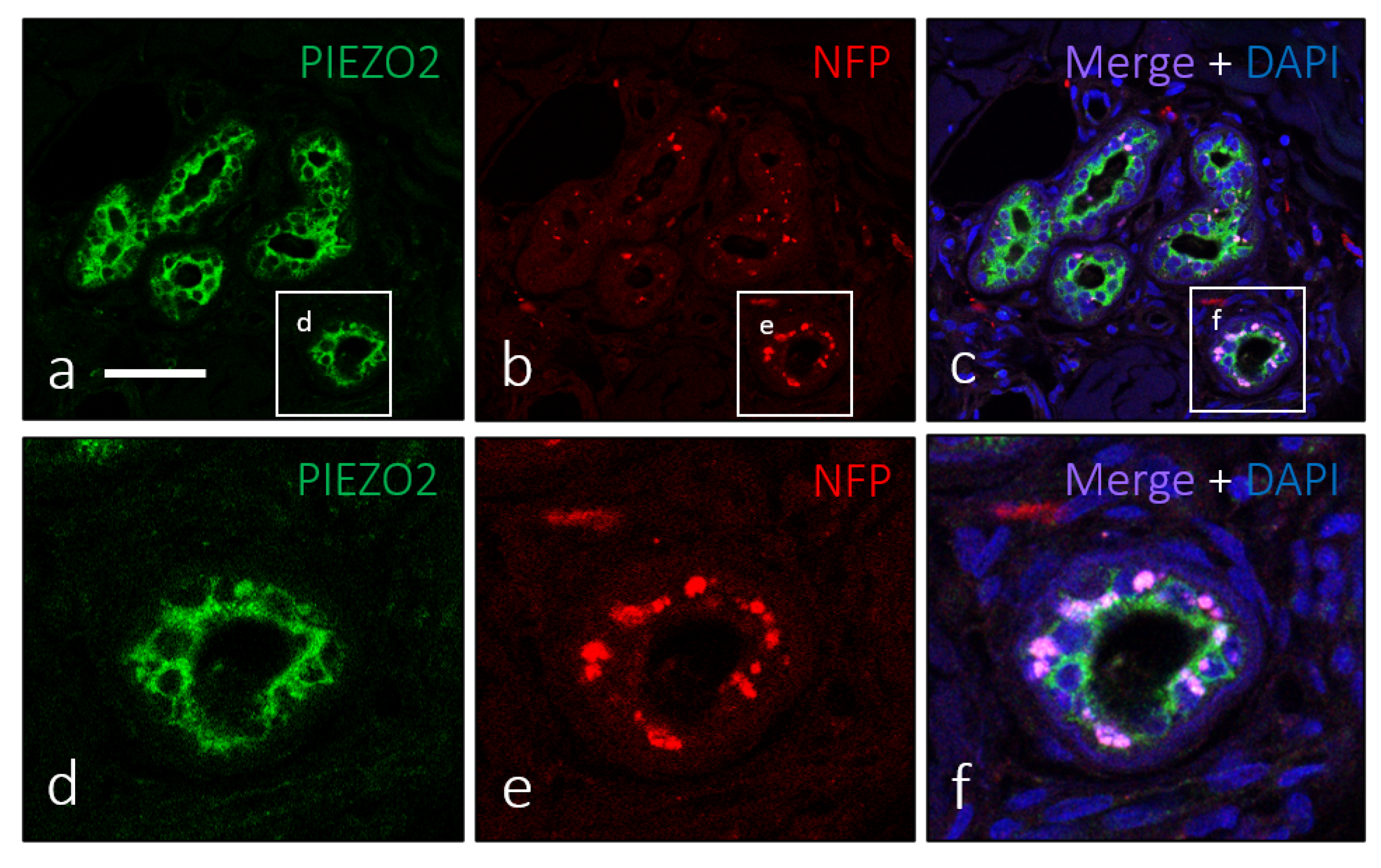
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramahi, E.; Choi, J.; Fuller, C.D.; Eng, T.Y. Merkel cell carcinoma. Am. J. Clin. Oncol. 2013, 36, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, J.C.; Stang, A.; DeCaprio, J.A.; Cerroni, L.; Lebbé, C.; Veness, M.; Nghiem, P. Merkel cell carcinoma. Nat. Rev. Dis. Primers 2017, 3, 17077. [Google Scholar] [CrossRef] [PubMed]
- Harms, P.W. Update on Merkel Cell Carcinoma. Clin. Lab. Med. 2017, 37, 485–501. [Google Scholar] [CrossRef]
- Pulitzer, M. Merkel Cell Carcinoma. Surg. Pathol. Clin. 2017, 10, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, N.C. Merkel cell carcinoma: Changing incidence trends. J. Surg. Oncol. 2005, 89, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.; Baker, K.; Redman, M.; Lachance, K.; Nguyen, M.H.; Parvathaneni, U.; Bhatia, S.; Nghiem, P.; Tseng, Y.D. Differential outcomes among immunosuppressed patients with Merkel cell carcinoma: Impact of immunosupression type on cancer-specific and overall survival. Am. J. Clin. Oncol. 2019, 42, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Sunshine, J.C.; Jahchan, N.S.; Sage, J.; Choi, J. Are there multiple cells of origin of Merkel cell carcinoma? Oncogene 2018, 37, 1409–1416. [Google Scholar] [CrossRef]
- Kervarrec, T.; Samimi, M.; Guyétant, S.; Sarma, B.; Chéret, J.; Blanchard, E.; Berthon, P.; Schrama, D.; Houben, R.; Touzé, A. Histogenesis of Merkel Cell Carcinoma: A Comprehensive Review. Front. Oncol. 2019, 9, 451. [Google Scholar] [CrossRef] [Green Version]
- Akaike, T.; Nghiem, P. Scientific and clinical developments in Merkel cell carcinoma: A polyomavirus-driven, often-lethal skin cancer. J. Dermatol. Sci. 2022, 105, 2–10. [Google Scholar] [CrossRef]
- Hernandez, L.E.; Mohsin, N.; Yaghi, M.; Frech, F.S.; Dreyfuss, I.; Nouri, K. Merkel cell carcinoma: An updated review of pathogenesis, diagnosis, and treatment options. Dermatol. Ther. 2022, 35, e15292. [Google Scholar] [CrossRef]
- Maricich, S.M.; Wellnitz, S.A.; Nelson, A.M.; Lesniak, D.R.; Gerling, G.J.; Lumpkin, E.A.; Zoghbi, H.Y. Merkel cells are essential for light-touch responses. Science 2009, 324, 1580–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, W.; Dong, P.; Fleming, M.; Luo, W. The specification and wiring of mammalian cutaneous low-threshold mechanoreceptors. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 389–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, R.; Cha, M.; Ling, J.; Jia, Z.; Coyle, D.; Gu, J.G. Merkel cells transduce and encode tactile stimuli to drive Abeta-afferent impulses. Cell 2014, 157, 664–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.; Kanda, H.; Ikeda, R.; Ling, J.; DeBerry, J.J.; Gu, J.G. Merkel disc is a serotonergic synapse in the epidermis for transmitting tactile signals in mammals. Proc. Natl. Acad. Sci. USA 2016, 113, E5491–E5500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felice, D.; Alaimo, A. Mechanosensitive Piezo channels in cancer: Focus on altered calcium signaling in cancer cells and in tumor progression. Cancers 2020, 12, 1780. [Google Scholar] [CrossRef]
- Szczot, M.; Nickolls, A.R.; Lam, R.M.; Chesler, A.T. The form and function of PIEZO2. Annu. Rev. Biochem. 2021, 90, 507–534. [Google Scholar] [CrossRef]
- Delmas, P.; Coste, B. Mechano-gated ion channels in sensory systems. Cell 2013, 155, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Anderson, E.O.; Schneider, E.R.; Bagriantsev, S.N. PIEZO2 in cutaneous and proprioceptive mechanotransduction in vertebrates. Curr. Top. Membr. 2017, 79, 197–217. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Lewis, A.H.; Grandl, J. Touch, tension, and transduction—The function and regulation of Piezo ion channels. Trends Biochem. Sci. 2017, 42, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Bagriantsev, S.N.; Gracheva, E.O.; Gallagher, P.G. Piezo proteins: Regulators of mechanosensation and other cellular processes. J. Biol. Chem. 2014, 289, 31673–31681. [Google Scholar] [CrossRef] [Green Version]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, J.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and PIEZO2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranade, S.S.; Woo, S.H.; Dubin, A.E.; Moshourab, R.A.; Wetzel, C.; Petrus, M.; Mathur, J.; Bégay, V.; Coste, B.; Mainquist, J.; et al. PIEZO2 is the major transducer of mechanical forces for touch sensation in mice. Nature 2014, 516, 121–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florez-Paz, D.; Bali, K.K.; Kuner, R.; Gomis, A. A critical role for PIEZO2 channels in the mechanotransduction of mouse proprioceptive neurons. Sci. Rep. 2016, 6, 25923. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.H.; Ranade, S.; Weyer, A.D.; Dubin, A.E.; Baba, Y.; Qui, Z.; Petrus, M.; Miyamoto, T.; Reddy, K.; Lumpkin, E.A.; et al. PIEZO2 is required for Merkel-cell mechanotransduction. Nature 2014, 509, 622–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Mesa, Y.; García-Piqueras, J.; García, B.; Feito, J.; Cabo, R.; Jobo, J.; Vega, J.A.; García-Suárez, O. Merkel cells and Meissner’s corpuscles in human digital skin display PIEZO2 immunoreactivity. J. Anat. 2017, 231, 978–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Piqueras, J.; García-Mesa, Y.; Cárcaba, L.; Feito, J.; Torres-Parejo, I.; Martín-Biedma, B.; Cobo, J.; García-Suárez, O.; Vega, J.A. Ageing of the somatosensory system at the periphery: Age-related changes in cutaneous mechanoreceptors. J. Anat. 2019, 234, 839–852. [Google Scholar] [CrossRef]
- Shin, K.C.; Park, H.J.; Kim, J.G.; Lee, I.H.; Cho, H.; Park, C.; Sung, T.S.; Koh, S.D.; Park, S.W.; Bae, Y.M. The PIEZO2 ion channel is mechanically activated by low-threshold positive pressure. Sci. Rep. 2019, 9, 6446. [Google Scholar] [CrossRef] [Green Version]
- Petho, Z.; Najder, K.; Bulk, E.; Schwab, A. Mechanosensitive ion channels push cancer progression. Cell Calcium 2019, 80, 79–90. [Google Scholar] [CrossRef]
- Morrison, K.M.; Miesegaes, G.R.; Lumpkin, E.A.; Maricich, S.M. Mammalian Merkel cells are descended from the epidermal lineage. Dev. Biol. 2009, 336, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Van Keymeulen, A.; Mascre, G.; Youseff, K.K.; Harel, I.; Michaux, C.; De Geest, N.; Szpalski, C.; Achouri, Y.; Bloch, W.; Hassan, B.A.; et al. Epidermal progenitors give rise to Merkel cells during embryonic development and adult homeostasis. J. Cell Biol. 2009, 187, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Bobos, M.; Hytiroglou, P.; Kostopoulos, I.; Karkavelas, G.; Papadimitriou, C.S. Immunohistochemical distinction between Merkel cell carcinoma and small cell carcinoma of the lung. Am. J. Dermatopathol. 2006, 28, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Gaub, B.M.; Müller, D.J. Mechanical stimulation of Piezo1 receptors depends on extracellular matrix proteins and directionality of force. Nano Lett. 2017, 17, 2064–2072. [Google Scholar] [CrossRef] [PubMed]
- McHugh, B.J.; Buttery, R.; Lad, Y.; Banks, S.; Haslett, C.; Sethi, T. Integrin activation by Fam38A uses a novel mechanism of R-Ras targeting to the endoplasmic reticulum. J. Cell Sci. 2010, 123, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kefauver, J.M.; Ward, A.B.; Patapoutian, A. Discoveries in structure and physiology of mechanically activated ion channels. Nature 2020, 587, 567–576. [Google Scholar] [CrossRef]
- Knepper, T.C.; Panchaud, R.A.; Muradova, E.; Cohen, L.; DeCaprio, J.A.; Khushalani, N.I.; Tsai, K.; Brohl, A.S. An analysis of the use of targeted therapies in patients with advanced Merkel cell carcinoma and an evaluation of genomic correlates of response. Cancer Med. 2021, 10, 5889–5896. [Google Scholar] [CrossRef]
- Das, B.K.; Kannan, A.; Nguyen, Q.; Gogoi, J.; Zhao, H.; Gao, L. Selective inhibition of Aurora kinase A by AK-01/LY3295668 attenuates MCC tumor growth by inducing MCC cell cycle arrest and apoptosis. Cancers 2021, 13, 3708. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Origin | Dilution | Supplier |
|---|---|---|---|
| Merkel cell markers | |||
| CK20 (clone Ks20.8-, IS777) | Mouse | Prediluted | Dako 1 |
| Chromogranin A | Mouse | Prediluted | Leica 2 |
| NFP (clone 2F11) | Mouse | 1:100 | Dako 1 |
| Synaptophysin (clone DAK-SYNAP) | Mouse | 1:100 | Dako 1 |
| PIEZO2 | Rabbit | 1:200 | Milipore Sigma 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Mesa, Y.; Martín-Sanz, R.; García-Piqueras, J.; Cobo, R.; Muñoz-Bravo, S.; García-Suárez, O.; Martín-Biedma, B.; Vega, J.A.; Feito, J. Merkel Cell Carcinoma Display PIEZO2 Immunoreactivity. J. Pers. Med. 2022, 12, 894. https://doi.org/10.3390/jpm12060894
García-Mesa Y, Martín-Sanz R, García-Piqueras J, Cobo R, Muñoz-Bravo S, García-Suárez O, Martín-Biedma B, Vega JA, Feito J. Merkel Cell Carcinoma Display PIEZO2 Immunoreactivity. Journal of Personalized Medicine. 2022; 12(6):894. https://doi.org/10.3390/jpm12060894
Chicago/Turabian StyleGarcía-Mesa, Yolanda, Raquel Martín-Sanz, Jorge García-Piqueras, Ramón Cobo, Saray Muñoz-Bravo, Olivia García-Suárez, Benjamín Martín-Biedma, José Antonio Vega, and Jorge Feito. 2022. "Merkel Cell Carcinoma Display PIEZO2 Immunoreactivity" Journal of Personalized Medicine 12, no. 6: 894. https://doi.org/10.3390/jpm12060894