
SERS-PLSR Analysis of Vaginal Microflora: Towards the Spectral Library of Microorganisms

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
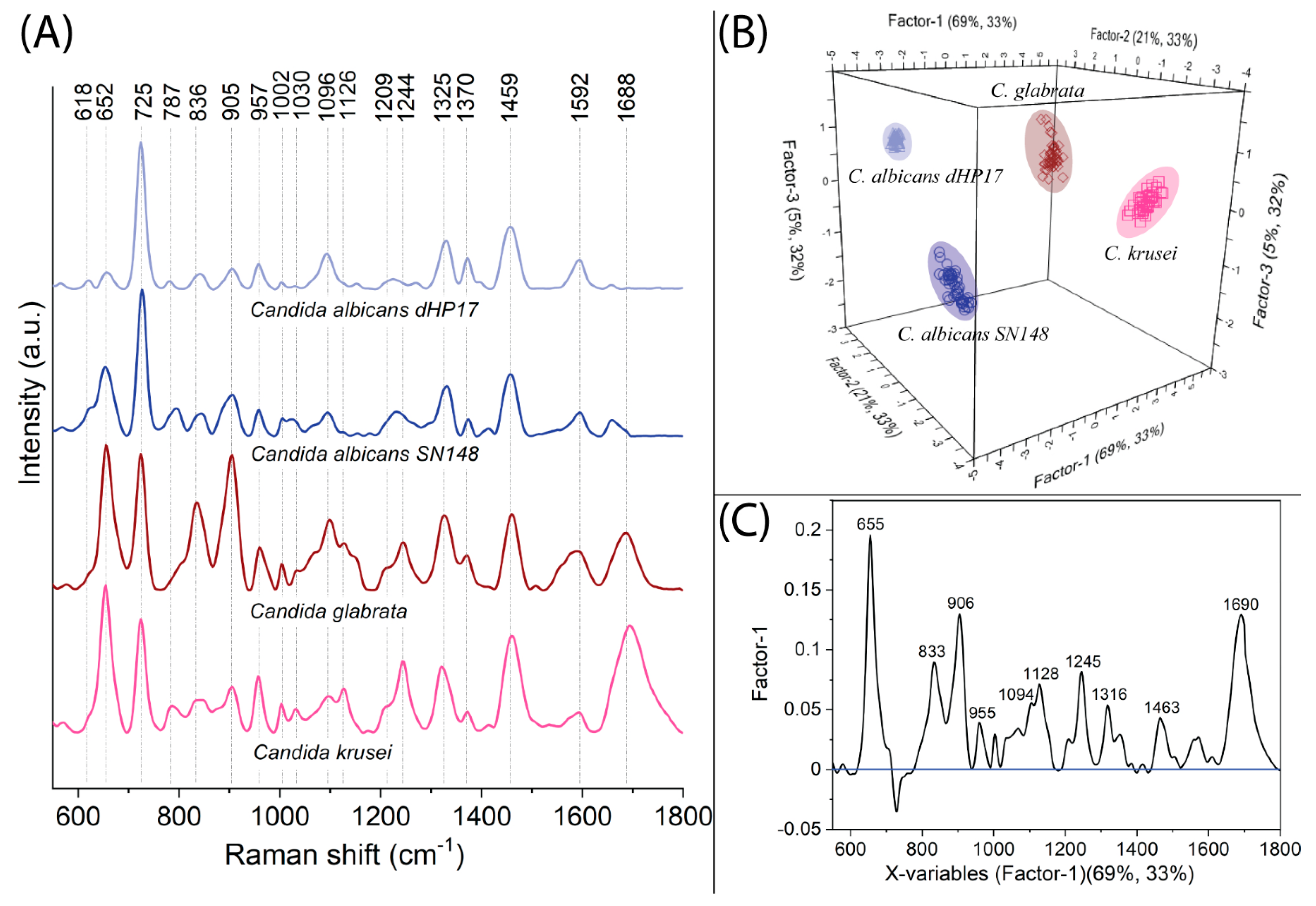
2.1. SERS-PLSR Analysis of Candida spp.—Vulvovaginal candidiasis (VVC) Disorder
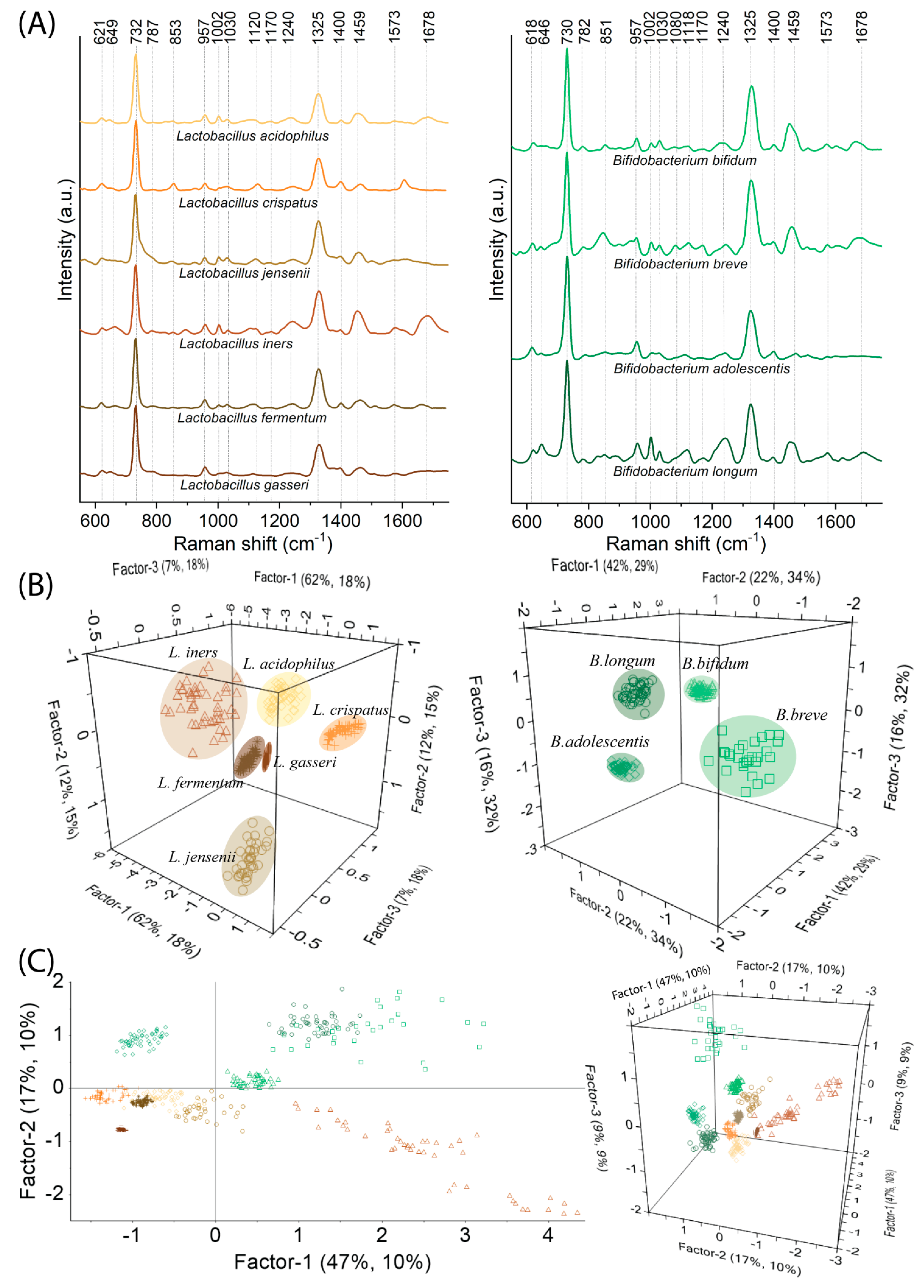
2.2. SERS-PLSR Analysis of Lactobacillus spp. and Bifidobacterium spp.—Natural Vaginal Flora
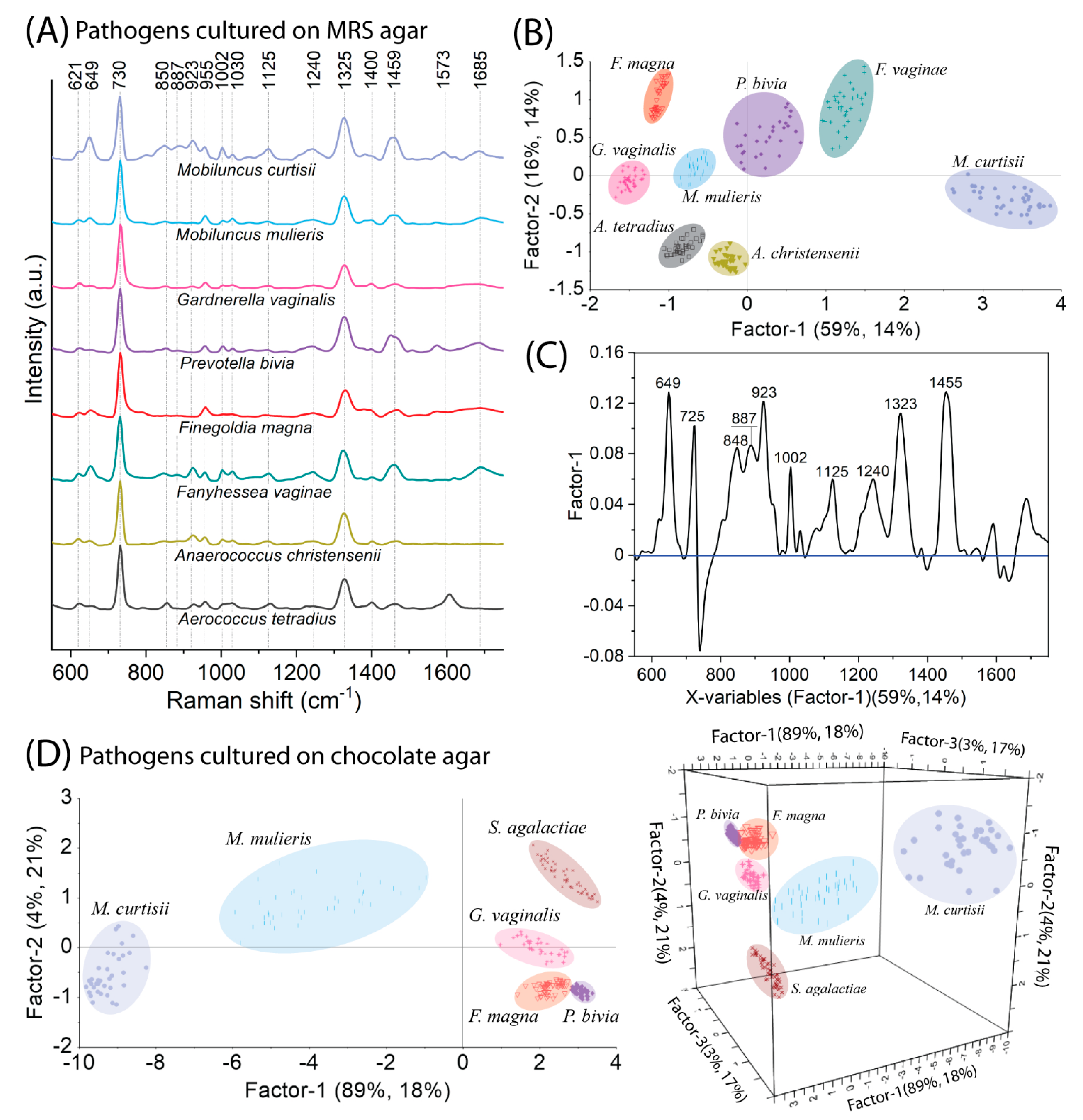
2.3. SERS-PLSR Analysis of the Most Prevalent Bacteria Responsible for BV
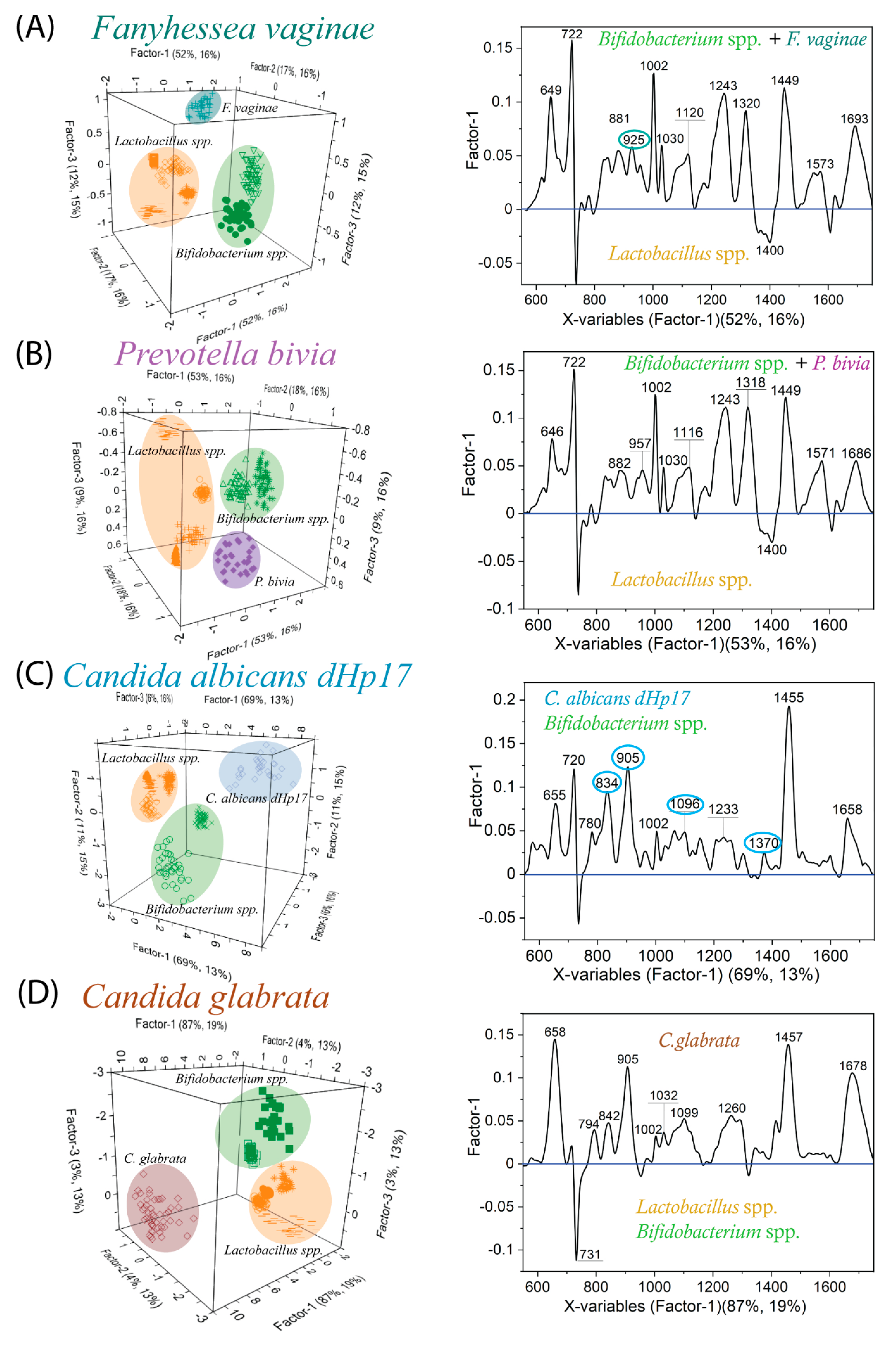
2.4. Bacterial Vaginosis vs. Vulvovaginal Candidiasis—SERS-Based Diagnosis
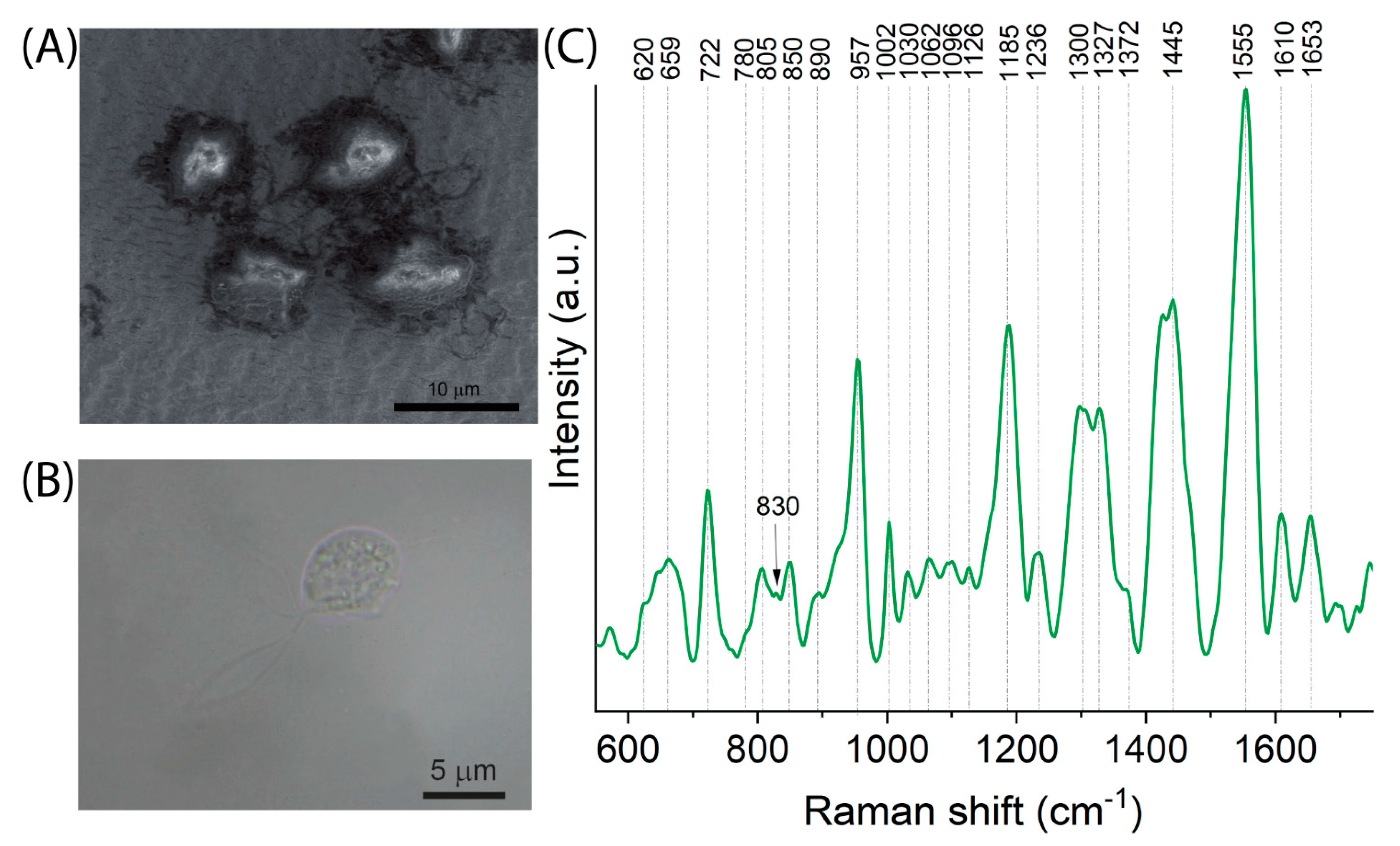
2.5. The Spectral Fingerprint of Trichomonas vaginalis—A Parasite That Leads to Trichomoniasis
3. Materials and Methods
3.1. Bacteria Species and the Culture Conditions
3.2. Sample Preparation for SERS Measurements
3.3. SERS Substrate Preparation
3.4. SERS Spectroscopy and Chemometric Analysis
3.5. Microscopic Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Redondo-lopez, V.; Cook, R.L.; Sobel, J.D. Emerging Role of Lactobacilli in the Control and Maintenance of the Vaginal Bacterial Microflora. Rev. Infect. Dis. 1990, 12, 856–872. [Google Scholar] [CrossRef] [PubMed]
- Hillier, S.L.; Krohn, M.A.; Rabe, L.K.; Klebanoff, S.J.; Eschenbach, D.A.; Hillier, S.L.; Krohn, M.A.; Rabe, L.K.; Klebanoff, S.J.; Eschenbach, D.A. The Normal Vaginal Flora, H2O2-Producing Lactobacilli, and Bacterial Vaginosis in Pregnant Women. Clin. Infect. Dis. 1993, 16, S273–S281. [Google Scholar] [CrossRef] [PubMed]
- Verstraelen, H.; Verhelst, R.; Claeys, G.; De Backer, E.; Temmerman, M.; Vaneechoutte, M. Longitudinal Analysis of the Vaginal Microflora in Pregnancy Suggests That L. Crispatus Promotes the Stability of the Normal Vaginal Microflora and That L. Gasseri and/or L. Iners Are More Conducive to the Occurrence of Abnormal Vaginal Microflora. BMC Microbiol. 2009, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawes, S.E.; Hillier, S.L.; Benedetti, J.; Stevens, C.E.; Koutsky, L.A.; Wølner-Hanssen, P.; Holmes, K.K. Hydrogen Peroxide—Producing Lactobacilli and Acquisition of Vaginal Infections. J. Infect. Dis. 1996, 174, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Verhelst, R.; Verstraelen, H.; Claeys, G.; Verschraegen, G.; Van Simaey, L.; De Ganck, C.; De Backer, E.; Temmerman, M.; Vaneechoutte, M. Comparison between Gram Stain and Culture for the Characterization of Vaginal Microflora: Definition of a Distinct Grade That Resembles Grade I Microflora and Revised Categorization of Grade I Microflora. BMC Microbiol. 2005, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Shipitsyna, E.; Roos, A.; Datcu, R.; Hallén, A.; Fredlund, H.; Jensen, J.S.; Engstrand, L.; Unemo, M. Composition of the Vaginal Microbiota in Women of Reproductive Age—Sensitive and Specific Molecular Diagnosis of Bacterial Vaginosis Is Possible? PLoS ONE 2013, 8, e60670. [Google Scholar] [CrossRef] [Green Version]
- Freitas, A.C.; Hill, J.E. Quantification, Isolation and Characterization of Bifidobacterium from the Vaginal Microbiomes of Reproductive Aged Women. Anaerobe 2017, 47, 145–156. [Google Scholar] [CrossRef]
- Chaban, B.; Links, M.G.; Jayaprakash, T.P.; Wagner, E.C.; Bourque, D.K.; Lohn, Z.; Albert, A.Y.; van Schalkwyk, J.; Reid, G.; Hemmingsen, S.M.; et al. Characterization of the Vaginal Microbiota of Healthy Canadian Women through the Menstrual Cycle. Microbiome 2014, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J.D. Desquamative Inflammatory Vaginitis: A New Subgroup of Purulent Vaginitis Responsive to Topical 2% Clindamycin Therapy. Am. J. Obstet. Gynecol. 1994, 171, 1215–1220. [Google Scholar] [CrossRef]
- Johnston, V.J.; Mabey, D.C. Global Epidemiology and Control of Trichomonas Vaginalis. Curr. Opin. Infect. Dis. 2008, 21, 56–64. [Google Scholar] [CrossRef]
- Toresani, I.; Limansky, A.; Bogado, I.; Guardati, M.C.; Viale, A.; Sutich, E.G.; Disease, P.; Group, S. Phenotypic and Genotypic Study of Streptococcus Agalactiae in Vagina of Pregnant Women in Argentina. Medicina 2001, 61, 295–300. [Google Scholar] [PubMed]
- El Aila, N.A.; Tency, I.; Claeys, G.; Saerens, B.; De Backer, E.; Temmerman, M.; Verhelst, R.; Vaneechoutte, M. Genotyping of Streptococcus Agalactiae(Group B Streptococci) Isolated from Vaginal and Rectal Swabs of Women at 35–37 Weeks of Pregnancy. BMC Infect. Dis. 2009, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smayevsky, J.; Canigia, L.F.; Lanza, A.; Bianchini, H. Vaginal Microflora Associated with Bacterial Vaginosis in Nonpregnant Women: Reliability of Sialidase Detection. Infect. Dis. Obstet. Gynecol. 2001, 9, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Verstraelen, H.; Verhelst, R.; Claeys, G.; Temmerman, M.; Vaneechoutte, M. Culture-Independent Analysis of Vaginal Microflora: The Unrecognized Association of Atopobium Vaginae with Bacterial Vaginosis. Am. J. Obstet. Gynecol. 2004, 191, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Mashburn, J. Vaginal Infections Update. J. Midwifery Women’s Health 2012, 57, 629–634. [Google Scholar] [CrossRef]
- Ison, C.A.; Hay, P.E. Validation of a Simplified Grading of Gram Stained Vaginal Smears for Use in Genitourinary Medicine Clinics. Sex. Transm. Infect. 2002, 78, 413–415. [Google Scholar] [CrossRef]
- De Backer, E.; Verhelst, R.; Verstraelen, H.; Alqumber, M.A.; Burton, J.P.; Tagg, J.R.; Temmerman, M.; Vaneechoutte, M. Quantitative Determination by Real-Time PCR of Four Vaginal Lactobacillus Species, Gardnerella Vaginalis and Atopobium Vaginae Indicates an Inverse Relationship between L. Gasseri and L. Iners. BMC Microbiol. 2007, 7, 115. [Google Scholar] [CrossRef] [Green Version]
- Eerola, E.; Lehtonen, O.P. Optimal Data Processing Procedure for Automatic Bacterial Identification by Gas-Liquid Chromatography of Cellular Fatty Acids. J. Clin. Microbiol. 1988, 26, 1745–1753. [Google Scholar] [CrossRef] [Green Version]
- Delaney, M.L.; Onderdonk, A.B. Nugent Score Related to Vaginal Culture in Pregnant Women. Obstet. Gynecol. 2001, 98, 79–84. [Google Scholar] [CrossRef]
- Mott, T.M.; Everley, R.A.; Wyatt, S.A.; Toney, D.M.; Croley, T.R. Comparison of MALDI-TOF/MS and LC-QTOF/MS Methods for the Identification of Enteric Bacteria. Int. J. Mass Spectrom. 2010, 291, 24–32. [Google Scholar] [CrossRef]
- Hanna, S.E.; Connor, C.J.; Wang, H.H. Real-Time Polymerase Chain Reaction for the Food Microbiologist: Technologies, Applications, and Limitations. J. Food Sci. 2005, 70, R49–R53. [Google Scholar] [CrossRef]
- Baele, M.; Vaneechoutte, M.; Verhelst, R.; Vancanneyt, M.; Devriese, L.A.; Haesebrouck, F. Identification of Lactobacillus Species Using TDNA-PCR. J. Microbiol. Methods 2002, 50, 263–271. [Google Scholar] [CrossRef]
- Cotton, T.M.; Kim, J.-H.; Chumanov, G.D. Application of Surface-Enhanced Raman Spectroscopy to Biological Systems. J. Raman Spectrosc. 1991, 22, 729–742. [Google Scholar] [CrossRef]
- Koglin, E.; Séquaris, J.-M. Surface Enhanced Raman Scattering of Biomolecules; Springer: Berlin/Heidelberg, Germany, 1986; Volume 134, pp. 1–57. [Google Scholar] [CrossRef]
- Herne, T.M.; Garrell, R.L. Borate Interference in Surface-Enhanced Raman Spectroscopy of Amines. Anal. Chem. 1991, 63, 2290–2294. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.S.; Suh, S.W.; Kim, M.S. Surface-Enhanced Raman Scattering of Nucleic Acid Components in Silver Sol: Uracil and Its Derivatives. J. Raman Spectrosc. 1988, 19, 261–265. [Google Scholar] [CrossRef]
- Liu, S.; Hu, Q.; Li, C.; Zhang, F.; Gu, H.; Wang, X.; Li, S.; Xue, L.; Madl, T.; Zhang, Y.; et al. Wide-Range, Rapid, and Specific Identification of Pathogenic Bacteria by Surface-Enhanced Raman Spectroscopy. ACS Sens. 2021, 6, 2911–2919. [Google Scholar] [CrossRef]
- Kashif, M.; Majeed, M.I.; Nawaz, H.; Rashid, N.; Abubakar, M.; Ahmad, S.; Ali, S.; Hyat, H.; Bashir, S.; Batool, F.; et al. Surface-Enhanced Raman Spectroscopy for Identification of Food Processing Bacteria. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 261, 119989. [Google Scholar] [CrossRef]
- Lim, J.; Nam, J.; Shin, H.; Park, J.; Song, H.; Kang, M.; Lim, K.; Choi, Y. Identification of Newly Emerging Influenza Viruses by Detecting the Virally Infected Cells Based on Surface Enhanced Raman Spectroscopy and Principal Component Analysis. Anal. Chem. 2019, 91, 5677–5684. [Google Scholar] [CrossRef]
- Lim, J.; Nam, J.; Yang, S.; Shin, H.; Jang, Y.; Bae, G.-U.; Kang, T.; Lim, K.; Choi, Y. Identification of Newly Emerging Influenza Viruses by Surface-Enhanced Raman Spectroscopy. Anal. Chem. 2015, 87, 11652–11659. [Google Scholar] [CrossRef]
- Dina, N.E.; Gherman, A.M.R.; Chiş, V.; Sârbu, C.; Wieser, A.; Bauer, D.; Haisch, C. Characterization of Clinically Relevant Fungi via SERS Fingerprinting Assisted by Novel Chemometric Models. Anal. Chem. 2018, 90, 2484–2492. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, M.; Barimah, A.O.; Chen, Q.; Li, H.; Shi, J.; El-Seedi, H.R.; Zou, X. Label-Free Surface Enhanced Raman Scattering Spectroscopy for Discrimination and Detection of Dominant Apple Spoilage Fungus. Int. J. Food Microbiol. 2021, 338, 108990. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Choo-Smith, L.P.; Endtz, H.P.; Bruining, H.A.; Puppels, G.J. Rapid Identification of Candida Species by Confocal Raman Microspectroscopy. J. Clin. Microbiol. 2002, 40, 594–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayin, I.; Kahraman, M.; Sahin, F.; Yurdakul, D.; Culha, M. Characterization of Yeast Species Using Surface-Enhanced Raman Scattering. Appl. Spectrosc. 2009, 63, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Witkowska, E.; Jagielski, T.; Kamińska, A.; Kowalska, A.; Hryncewicz-Gwóźdź, A.; Waluk, J. Detection and Identification of Human Fungal Pathogens Using Surface-Enhanced Raman Spectroscopy and Principal Component Analysis. Anal. Methods 2016, 8, 8427–8434. [Google Scholar] [CrossRef]
- Gherman, A.M.R.; Dina, N.E.; Chiș, V.; Wieser, A.; Haisch, C. Yeast Cell Wall—Silver Nanoparticles Interaction: A Synergistic Approach between Surface-Enhanced Raman Scattering and Computational Spectroscopy Tools. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2019, 222, 117223. [Google Scholar] [CrossRef]
- Pan, Y.L.; Yang, T.S.; Chang, T.C.; Chang, H.C. Rapid Identification of Candida Albicans Based on Raman Spectral Biosensing Technology. In Proceedings of the 2009 IEEE 3rd International Conference Nano/Molecular Medicine and Engineering, Tainan, Taiwan, 18–21 October 2009; pp. 120–124. [Google Scholar] [CrossRef]
- Keleştemur, S.; Çulha, M. Understanding and Discrimination of Biofilms of Clinically Relevant Microorganisms Using Surface-Enhanced Raman Scattering. Appl. Spectrosc. 2016, 71, 1180–1188. [Google Scholar] [CrossRef]
- Keleştemur, S.; Çobandede, Z.; Çulha, M. Biofilm Formation of Clinically Important Microorganisms on 2D and 3D Poly (Methyl Methacrylate) Substrates: A Surface-Enhanced Raman Scattering Study. Colloids Surf. B Biointerfaces 2020, 188, 110765. [Google Scholar] [CrossRef]
- Cui, L.; Chen, S.; Zhang, K. Effect of Toxicity of Ag Nanoparticles on SERS Spectral Variance of Bacteria. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 137, 1061–1066. [Google Scholar] [CrossRef]
- Berus, S.M.; Adamczyk-Popławska, M.; Młynarczyk-Bonikowska, B.; Witkowska, E.; Szymborski, T.; Waluk, J.; Kamińska, A. SERS-Based Sensor for the Detection of Sexually Transmitted Pathogens in the Male Swab Specimens: A New Approach for Clinical Diagnosis. Biosens. Bioelectron. 2021, 189, 113358. [Google Scholar] [CrossRef]
- Szymborski, T.; Stepanenko, Y.; Niciński, K.; Piecyk, P.; Berus, S.M.; Adamczyk-Popławska, M.; Kamińska, A. Ultrasensitive SERS Platform Made via Femtosecond Laser Micromachining for Biomedical Applications. J. Mater. Res. Technol. 2021, 12, 1496–1507. [Google Scholar] [CrossRef]
- Fujisawa, T.; Adachi, S.; Toba, T.; Arihara, K.; Mitsuoka, T. Lactobacillus Kefiranofaciens Sp. Nov. Isolated from Kefir Grains. Int. J. Syst. Bacteriol. 1988, 38, 12–14. [Google Scholar] [CrossRef]
- Gomes, A.M.P.; Malcata, F.X. Bifidobacterium Spp. and Lactobacillus Acidophilus: Biological, Biochemical, Technological and Therapeutical Properties Relevant for Use as Probiotics. Trends Food Sci. Technol. 1999, 10, 139–157. [Google Scholar] [CrossRef]
- Salman, S.A.; Baharoon, S.A. Septic Arthritis of the Knee Joint Secondary to Prevotella Bivia. Saudi Med. J. 2009, 30, 426–428. [Google Scholar] [PubMed]
- Sudbery, P.; Gow, N.; Berman, J. The Distinct Morphogenic States of Candida Albicans. Trends Microbiol. 2004, 12, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Kubryk, P.; Niessner, R.; Ivleva, N.P. The Origin of the Band at around 730 Cm-1 in the SERS Spectra of Bacteria: A Stable Isotope Approach. Analyst 2016, 141, 2874–2878. [Google Scholar] [CrossRef] [Green Version]
- Gieroba, B.; Sroka-Bartnicka, A.; Kazimierczak, P.; Kalisz, G.; Pieta, I.S.; Nowakowski, R.; Pisarek, M.; Przekora, A. Effect of Gelation Temperature on the Molecular Structure and Physicochemical Properties of the Curdlan Matrix: Spectroscopic and Microscopic Analyses. Int. J. Mol. Sci. 2020, 21, 6154. [Google Scholar] [CrossRef]
- Noothalapati, H.; Sasaki, T.; Kaino, T.; Kawamukai, M.; Ando, M.; Hamaguchi, H.; Yamamoto, T. Label-Free Chemical Imaging of Fungal Spore Walls by Raman Microscopy and Multivariate Curve Resolution Analysis. Sci. Rep. 2016, 6, 27789. [Google Scholar] [CrossRef] [Green Version]
- Sujith, A.; Itoh, T.; Abe, H.; Anas, A.A.; Yoshida, K.; Biju, V.; Ishikawa, M. Surface Enhanced Raman Scattering Analyses of Individual Silver Nanoaggregates on Living Single Yeast Cell Wall. Appl. Phys. Lett. 2008, 92, 103901. [Google Scholar] [CrossRef]
- Singh, B.N.; Hayes, G.R.; Lucas, J.J.; Sommer, U.; Viseux, N.; Mirgorodskaya, E.; Trifonova, R.T.; Sassi, R.R.S.; Costello, C.E.; Fichorova, R.N. Structural Details and Composition of Trichomonas Vaginalis Lipophosphoglycan in Relevance to the Epithelial Immune Function. Glycoconj. J. 2009, 26, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Ghonaim, H.M.; Periasamy, N.; Noro, M.G.; Anwar, J. Towards a Simplified Model Membrane of Skin Lipids: Preparation and Characterisation of a Ternary Lipid Mixture. Available online: https://eprints.lancs.ac.uk/id/eprint/73840/ (accessed on 8 August 2022).
- Talari, A.C.S.; Movasaghi, Z.; Rehman, S.; Rehman, I.U. Raman Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2015, 50, 46–111. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raman Shifts (cm−1) for Candida spp. | Assignment for Candida spp. | Raman Shifts (cm−1) for Bacteria | Assignment for Bacteria | Raman Shifts (cm−1) for T. vaginalis | Assignment for T. vaginalis |
|---|---|---|---|---|---|
| 618 | C-C twisting mode of phenylalanine (protein) | 621 a 620 b 622 c 622 d 624 e 622 f | C-C twisting mode of phenylalanine (protein) | 620 | C-C twisting mode of phenylalanine (protein) |
| 654 | C-S stretching, C-C twisting of proteins (tyrosine) COO− deformation in amino acids, guanine and thymine (ring breathing modes) | 649–664 a 645–652 b 649 c 649–665 d 647 e 649 f | C-S stretching, C-C twisting of proteins (tyrosine) COO− deformation in amino acids, guanine and thymine (ring breathing modes) | 659 | C-S stretching, C-C twisting of proteins (tyrosine) COO− deformation in amino acids, guanine and thymine (ring breathing modes) |
| 725 | Adenine (FAD, NAD, ATP, DNA) | 732 a 730 b 730 c 730 d 730 e 730 f | Adenine (FAD, NAD, ATP, DNA) | 722 | Adenine (FAD, NAD, ATP, DNA) |
| 787 | Ring breathing mode of cytosine, uracil, thymine, (O−P−O) symmetric stretching of nucleic acid | 782–787 a 781–794 b 790–800 c 790–800 d 787 e 784–791 f | Ring breathing mode of cytosine, uracil, thymine, (O−P−O) symmetric stretching of nucleic acid | 780 | Ring breathing mode of cytosine, uracil, thymine, (O−P−O) symmetric stretching of nucleic acid |
| - | - | - | - | 805 | Uracil-based ring breathing mode |
| 834 | Stretching O-P-O of DNA deformative vibrations of amine groups, tyrosine | - | - | 830 | Stretching O-P-O of DNA, deformative vibrations of amine groups, tyrosine |
| - | - | 847–855 a 846–853 b 845–847 c 853 d 853 e 853 f | Thymine, ring breathing mode of tyrosine, C-C stretch of proline ring, single bond stretching vibrations for the amino acids and valine and polysaccharides | 850 | Thymine, ring breathing mode of tyrosine, C-C stretch of proline ring, single bond stretching vibrations for the amino acids and valine and polysaccharides |
| - | - | - | - | 890 | Proteins, saccharides |
| 905 | Chitin, monosaccharide (B glucose) C-C skeletal stretching, tyrosine | - | - | - | - |
| 957 | Chitin, C=C deformation, C-N stretching, C−O stretching, CH3 symmetric stretching of proteins (α-helix) | 957 a 956 b 955 c 956 d 956 e 956 f | C=C deformation, C-N stretching, C−O stretching, CH3 symmetric stretching of proteins (α-helix) | 957 | C=C deformation, C-N stretching, C−O stretching, CH3 symmetric stretching of proteins (α-helix) |
| 1002 | Phenylalanine, C-C aromatic ring stretching | 1002 a 1002 b 1002 c 1002 d 1002 e 1002 f | Phenylalanine, C-C aromatic ring stretching | 1002 | Phenylalanine, C-C aromatic ring stretching |
| 1030 | C-N stretching, C-C stretching (phospholipids, carbohydrates e.g., chitin), C-H in-plane bending mode of phenylalanine | 1030 a 1030 b 1030 c 1030 d 1030 e 1030 f | C-N stretching, C-C stretching (phospholipids, carbohydrates), C-H in-plane bending mode of phenylalanine | 1030 | C-N stretching, C-C stretching (phospholipids, carbohydrates), C-H in-plane bending mode of phenylalanine |
| - | - | - | - | 1062 | Acyl group of ceramides stretching O-P-O of DNA, skeletal C-C stretching (lipids), C-O, C-C stretching (carbohydrates) |
| 1096 | 1,3-β-D-glucan, PO2- symmetric stretching, C–O–C stretching modes in polysaccharides (e.g., chitin), adenine, polyadenine ν(C-C), ν(C-O), phospholipids | 1070–1080 b 1072 c 1080–1096 d 1088–1097 e 1096 f | PO2- symmetric stretching, C–O–C stretching modes in polysaccharides, adenine, polyadenine, ν(C-C), ν(C-O), phospholipids | 1096 | PO2- symmetric stretching, C–O–C stretching modes in polysaccharides, adenine, polyadenine, ν(C-C), ν(C-O), phospholipids |
| 1126 | =C-O-C= (unsaturated fatty acids in lipids), C-O-C stretching modes in nucleic acids, PO2_ stretching in nucleic acid, C-O and C-C stretching in carbohydrates, C-N stretching in proteins, 1,3-β-D-glucan, N-acetylglucosamine | 1120 a 1112–1126 b 1124 c 1122 d 1124 e 1124 f | =C-O-C= (unsaturated fatty acids in lipids), C-O-C stretching modes in nucleic acids, PO2_ stretching in nucleic acid, C-O and C-C stretching in carbohydrates, C-N stretching in proteins | 1126 | =C-O-C= (unsaturated fatty acids in lipids), C-O-C stretching modes in nucleic acids, PO2_ stretching in nucleic acid, C-O and C-C stretching in carbohydrates, C-N stretching in proteins Acyl group of ceramides |
| - | - | 1170 a 1170 b | C-H in-plane bending mode of tyrosine, ν(C=C) δ(COH) (lipid) | - | - |
| - | - | - | - | 1185 | Cytosine, guanine, adenine |
| 1209 | Chitin | - | - | - | - |
| 1244 | Amide III (arising from coupling of C-N stretching and N-H bonding—can be mixed with vibrations of side chains), β-1,3-glucan | 1230–1248 a 1230–1248 b 1243 c 1240 d 1243 e 1245 f | Amide III (arising from coupling of C-N stretching and N-H bonding—can be mixed with vibrations of sidechains) | 1236 | Amide III (arising from coupling of C-N stretching and N-H bonding—can be mixed with vibrations of side chains) |
| - | - | - | - | 1300 | CH2 deformation twisting and wagging (lipids), phospholipids, fatty acids, Acyl group of ceramides |
| 1325 | Chitin, β-1,3-glucan, mannose, CH3CH2 wagging in purine bases, NH2 stretching in adenine and polyadenine, phospholipids | 1325 a 1325 b 1325 c 1325 d 1325 f | CH3CH2 wagging in purine bases, NH2 stretching in adenine and polyadenine, phospholipids | 1327 | CH3CH2 wagging in purine bases, NH2 stretching in adenine and polyadenine, phospholipids |
| 1370–1380 | Chitin, 1,3-β-D-glucan, ring breathing modes of the DNA/RNA bases, COO− stretching, C-H bending of proteins, saccharides | - | - | 1372 | Ring breathing modes of the DNA/RNA bases, COO− stretching, C-H bending of proteins, saccharides |
| - | - | 1400 a 1400 b 1400 c 1400 d 1400 e 1400 f | C=O symmetric stretch, CH2 deformation, N-H in plane deformation | - | - |
| 1459 | Chitin, mannose, 1,3-β-D-glucan, CH2 deformation (protein, lipids), CH2 bending mode of protein and lipids, C-H vibrations proteins and lipids CH2 wagging, CH2/CH3 deformation, CH2 bending and scissoring of phospholipids | 1449–1468 a 1449–1468 b 1449–1468 c 1449–1468 d 1449–1468 e 1449–1463 f | CH2 deformation (protein, lipids), CH2 bending mode of protein and lipids, C-H vibrations proteins and lipids, CH2 wagging, CH2/CH3 deformation, CH2 bending and scissoring of phospholipids | 1444 | CH2 deformation (protein, lipids), CH2 bending mode of protein and lipids, C-H vibrations proteins and lipids, CH2 wagging, CH2/CH3 deformation, CH2 bending and scissoring of phospholipids |
| - | - | - | - | 1555 | Stretching C-N and deformation N-H of Amide II |
| 1584–1598 | Amide I band of proteins (mannans), due to C=O stretching C=C bending mode (phenylalanine), guanine, adenine, tryptophan | 1570–1578 a 1570–1576 b 1573 c 1570 d 1590 e 1578 f | Amide I band of proteins, due to C=O stretching C=C bending mode (phenylalanine), guanine, adenine, tryptophan | - | - |
| - | - | - | - | 1610 | Cytosine NH2, tyrosine, phenylalanine ring vibration |
| 1663–1693 | Amide I of proteins, ν(C=C) cis, lipids, fatty acids Carbonyl stretch (C=O) | 1660–1680 a 1660–1690 b 1680–1689 c 1670–1700 d 1680–1700 e 1664–1694 f | Amide I of proteins, ν(C=C) cis, lipids, fatty acids Carbonyl stretch (C=O) | 1653 | Ceramide backbone, Amide I of proteins, ν(C=C) cis, lipids, fatty acids Carbonyl stretch (C=O) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berus, S.M.; Adamczyk-Popławska, M.; Goździk, K.; Przedpełska, G.; Szymborski, T.R.; Stepanenko, Y.; Kamińska, A. SERS-PLSR Analysis of Vaginal Microflora: Towards the Spectral Library of Microorganisms. Int. J. Mol. Sci. 2022, 23, 12576. https://doi.org/10.3390/ijms232012576
Berus SM, Adamczyk-Popławska M, Goździk K, Przedpełska G, Szymborski TR, Stepanenko Y, Kamińska A. SERS-PLSR Analysis of Vaginal Microflora: Towards the Spectral Library of Microorganisms. International Journal of Molecular Sciences. 2022; 23(20):12576. https://doi.org/10.3390/ijms232012576
Chicago/Turabian StyleBerus, Sylwia Magdalena, Monika Adamczyk-Popławska, Katarzyna Goździk, Grażyna Przedpełska, Tomasz R. Szymborski, Yuriy Stepanenko, and Agnieszka Kamińska. 2022. "SERS-PLSR Analysis of Vaginal Microflora: Towards the Spectral Library of Microorganisms" International Journal of Molecular Sciences 23, no. 20: 12576. https://doi.org/10.3390/ijms232012576