
Anti-PTK7 Monoclonal Antibodies Exhibit Anti-Tumor Activity at the Cellular Level and in Mouse Xenograft Models of Esophageal Squamous Cell Carcinoma

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
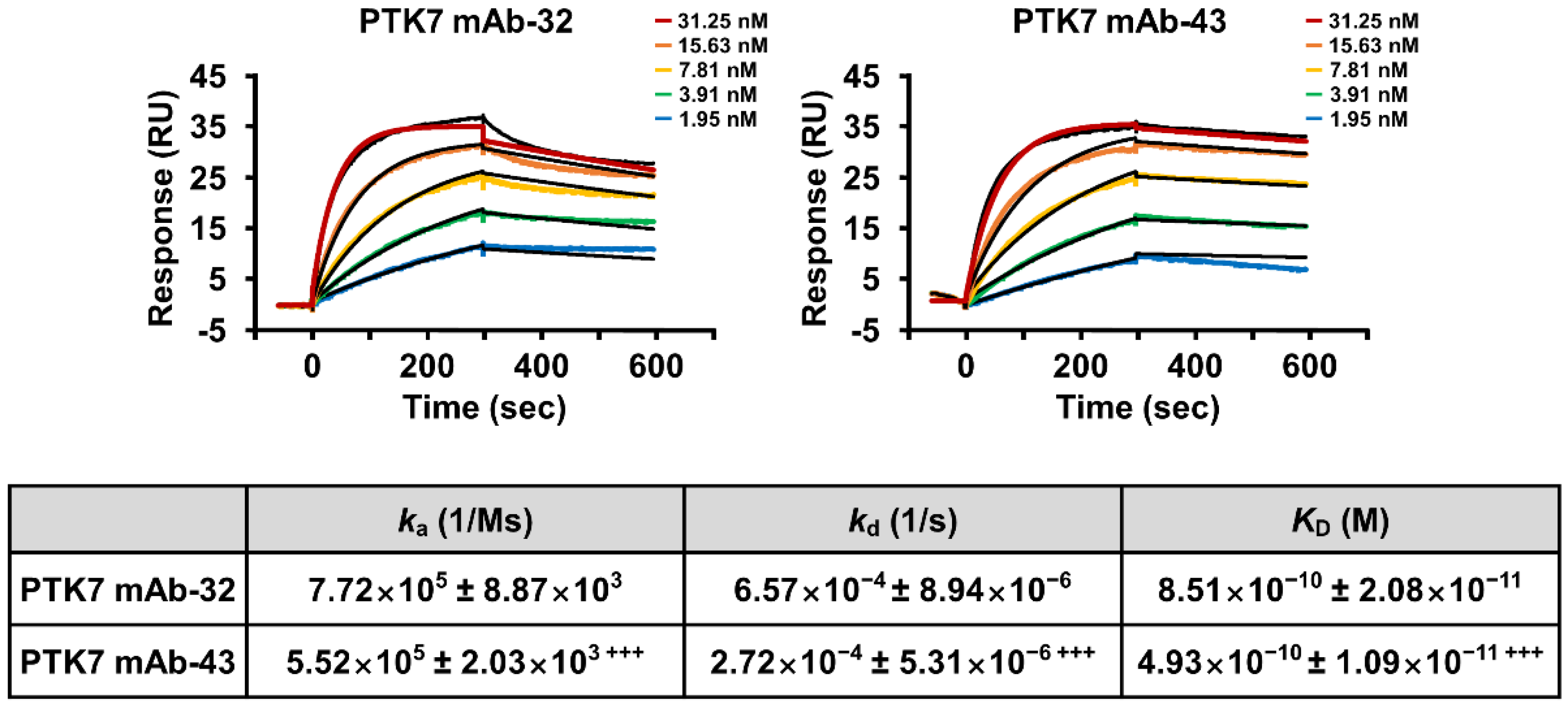
2.1. PTK7 mAbs Bind to sPTK7 with High Affinity
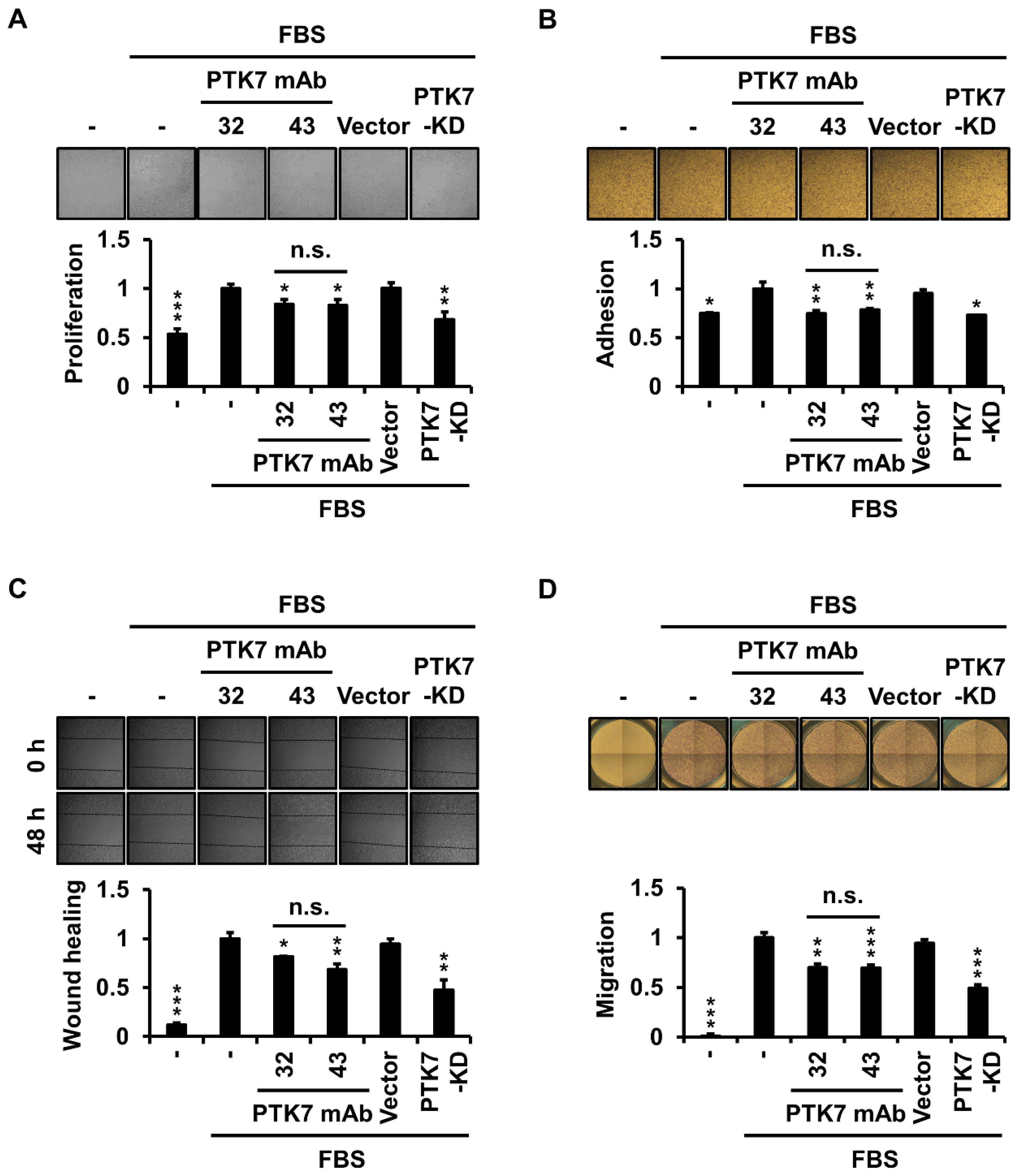
2.2. PTK7 mAbs Reduce Cell Proliferation, Adhesion, Wound Healing, and Migration in ESCC KYSE-30 Cells
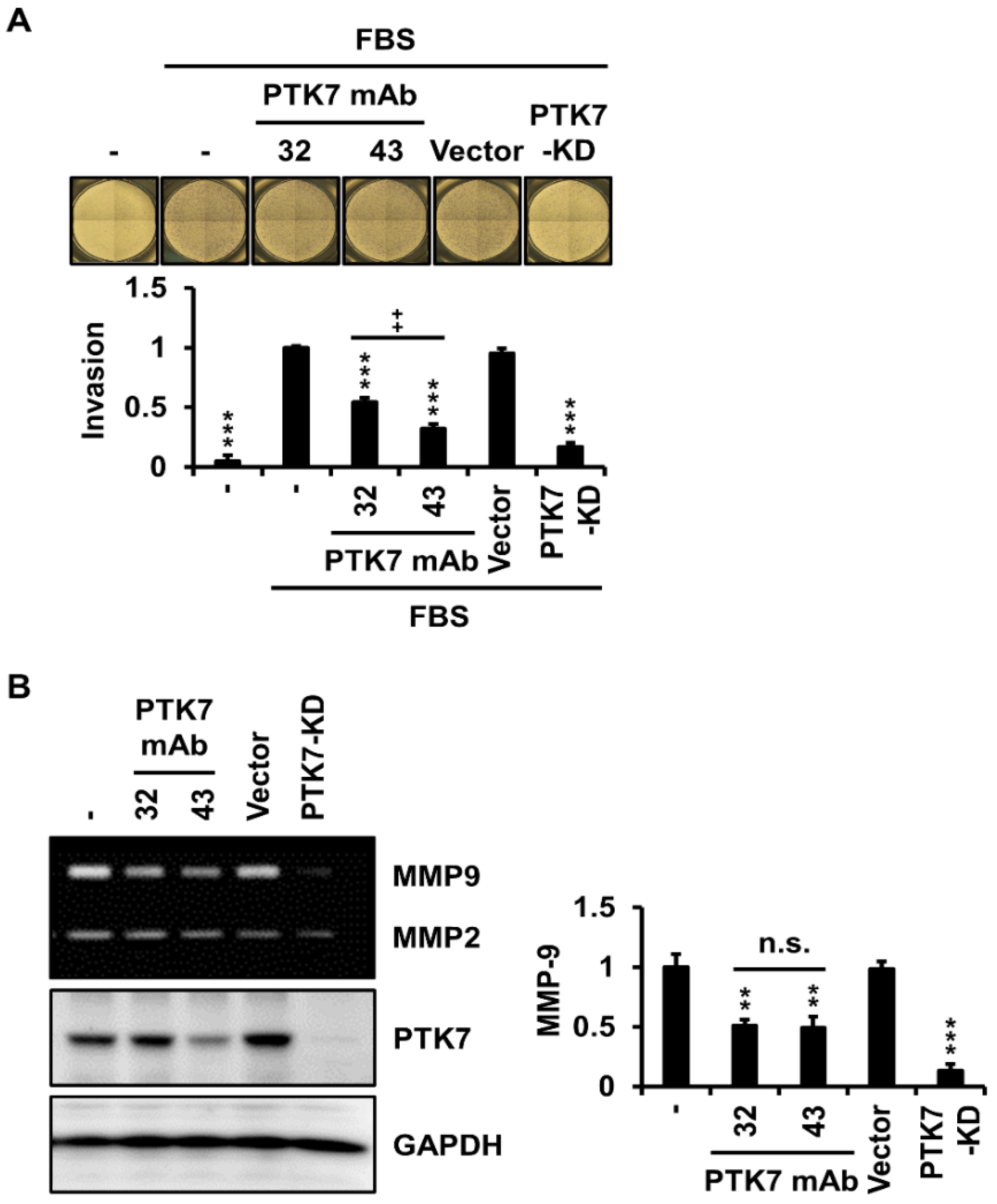
2.3. PTK7 mAbs Reduce Invasiveness and Matrix Metalloproteinase (MMP)-9 Secretion in ESCC KYSE-30 Cells
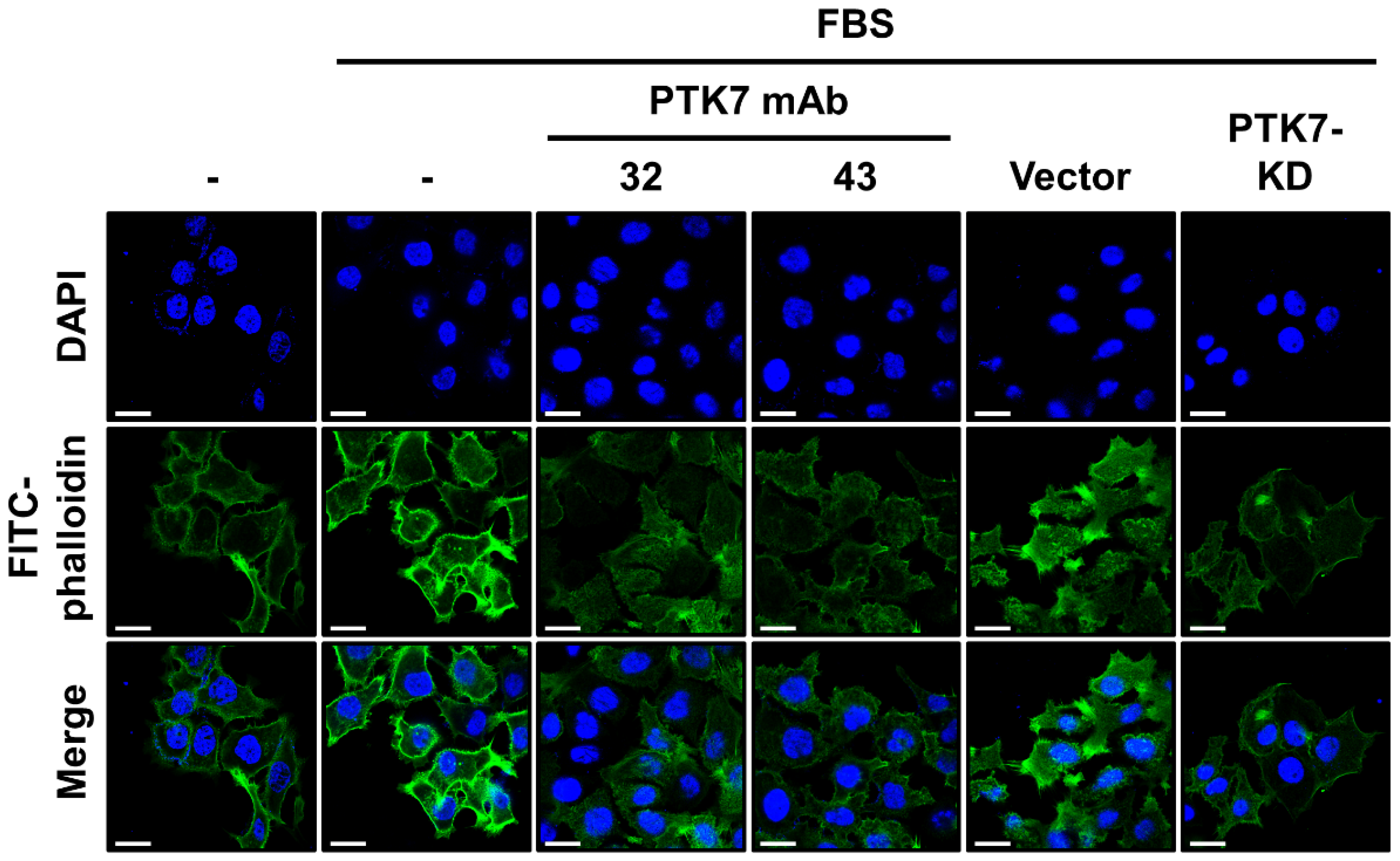
2.4. PTK7 mAbs Inhibit Actin Polymerization in the Cortical Area of KYSE-30 Cells
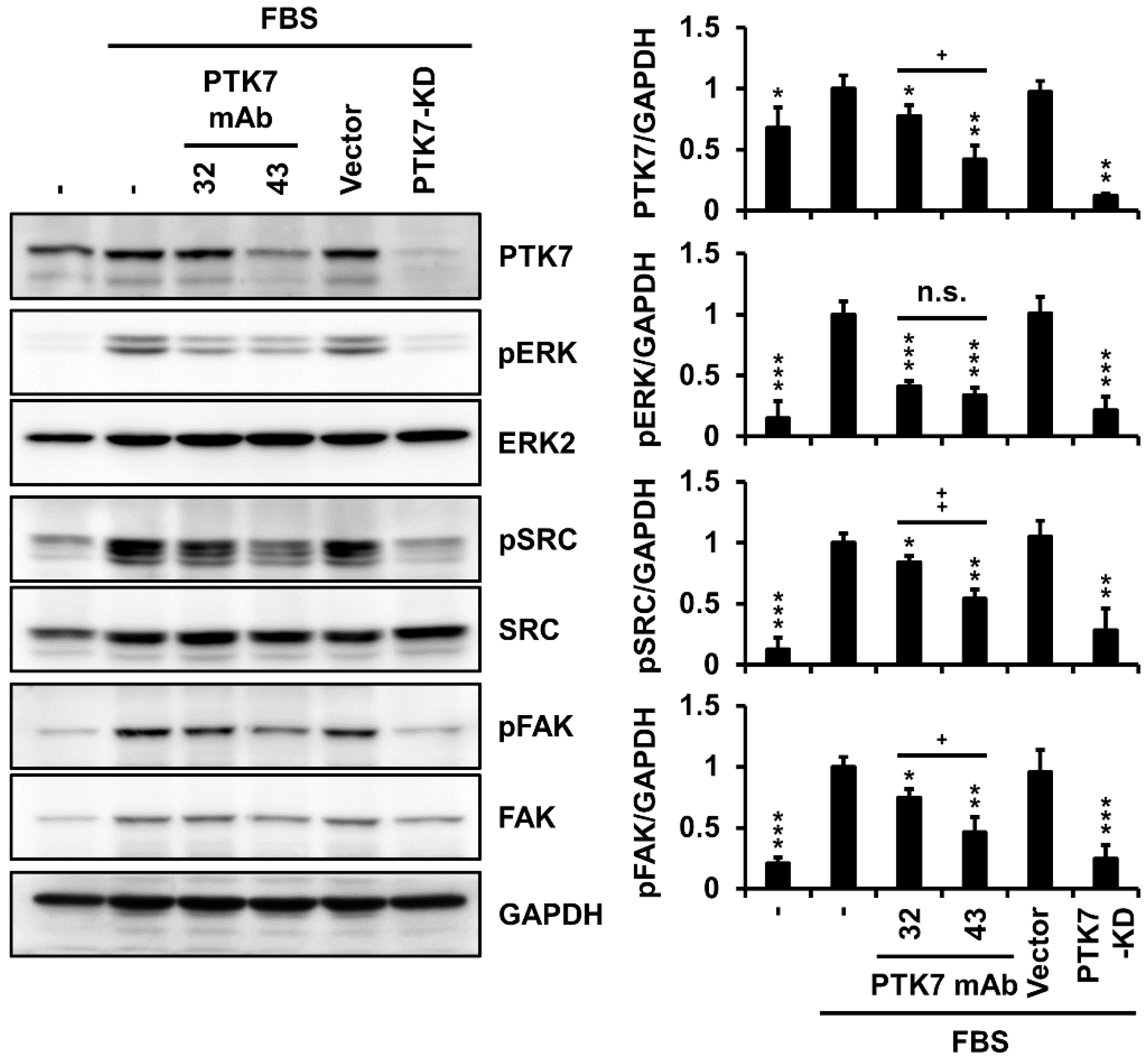
2.5. PTK7 mAbs Not Only Inhibit Activation of Oncogenic Signaling Proteins but Also Reduce PTK7 Levels in KYSE-30 Cells
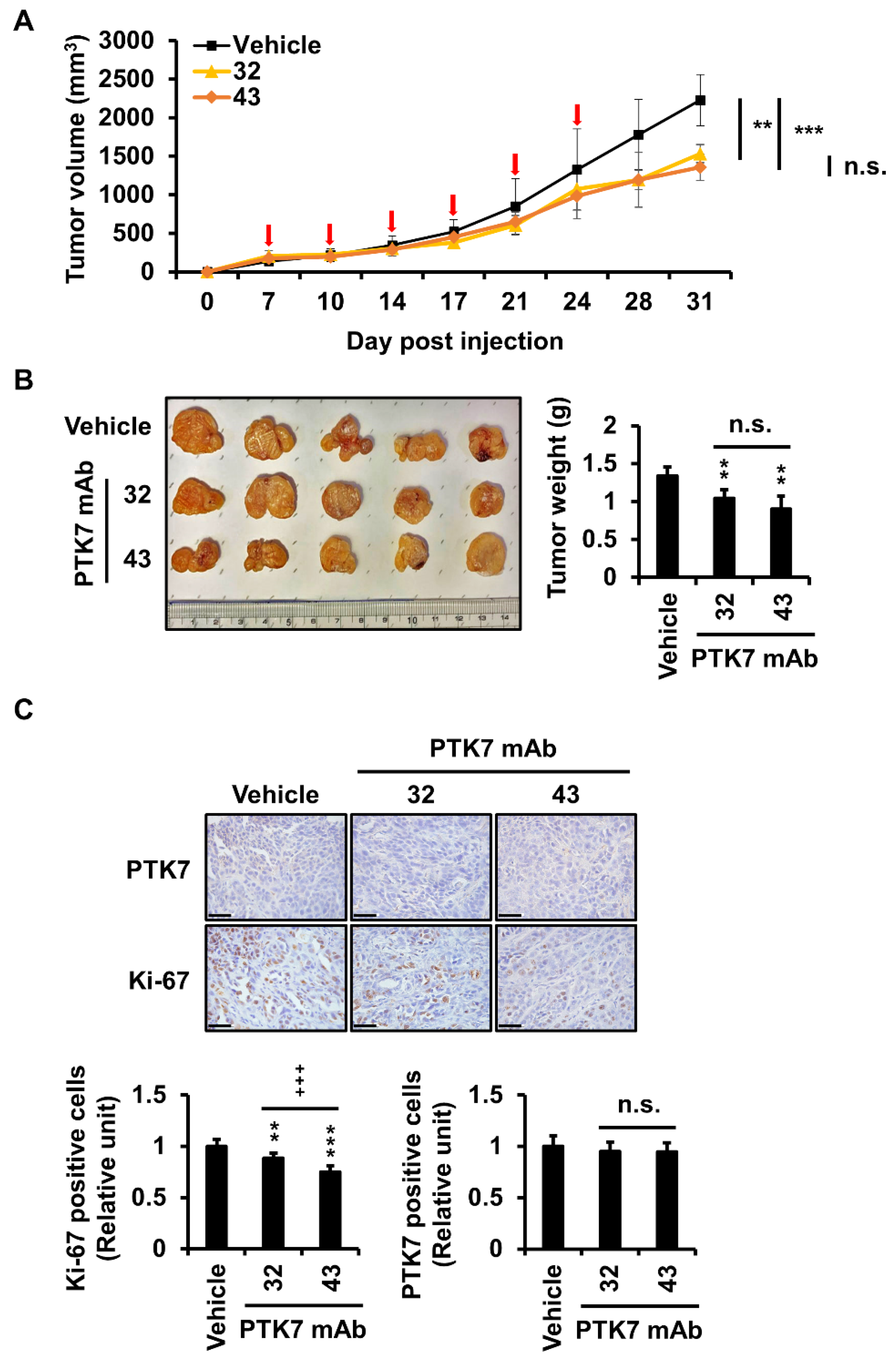
2.6. PTK7 mAbs Reduce Tumor Volume, Weight, and Number of Proliferating Cells in a Xenograft Mouse Model of ESCC
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Generation of PTK7 Knockdown Lentiviruses and Infection of KYSE-30 Cells
4.3. Antibodies
4.4. Production and Purification of Anti-PTK7 mAbs
4.5. Binding Affinity Analysis of PTK7 mAbs Using SPR Method
4.6. Cell Proliferation Assay in 3D Matrigel Culture
4.7. Cell Adhesion Assay
4.8. Wound-Healing Assay
4.9. Chemotactic Migration and Invasion Assays
4.10. Gelatin Zymography
4.11. Analysis of Actin Filament Distribution in Cells
4.12. Western Blot Analysis
4.13. Analysis of Tumor Growth in Xenograft Mouse Model of ESCC
4.14. IHC Staining
4.15. Ethics Statement
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobsen, A.V.; Murphy, J.M. The secret life of kinases: Insights into non-catalytic signalling functions from pseudokinases. Biochem. Soc. Trans. 2017, 45, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Kung, J.E.; Jura, N. Prospects for pharmacological targeting of pseudokinases. Nat. Rev. Drug Discov. 2019, 18, 501–526. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.-S.; Kwon, J.; Lee, H.W.; Kang, M.C.; Na, H.W.; Lee, S.-T.; Park, J.H. Oncogenic role of protein tyrosine kinase 7 in esophageal squamous cell carcinoma. Cancer Sci. 2013, 104, 1120–1126. [Google Scholar] [CrossRef]
- Damelin, M.; Bankovich, A.; Bernstein, J.; Lucas, J.; Chen, L.; Williams, S.; Park, A.; Aguilar, J.; Ernstoff, E.; Charati, M.; et al. A PTK7-targeted antibody-drug conjugate reduces tumor-initiating cells and induces sustained tumor regressions. Sci. Transl. Med. 2017, 9, eaag2611. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ma, W.; Li, Q.; Zhao, D.; Shao, X.; Huang, Q.; Hao, L.; Lin, Y. Aptamer-targeted DNA nanostructures with doxorubicin to treat protein tyrosine kinase 7-positive tumours. Cell Prolif. 2019, 52, e12511. [Google Scholar] [CrossRef] [Green Version]
- Schoeberl, B.; Faber, A.C.; Li, D.; Liang, M.C.; Crosby, K.; Onsum, M.; Burenkova, O.; Pace, E.; Walton, Z.; Nie, L.; et al. An ErbB3 antibody, MM-121, is active in cancers with ligand-dependent activation. Cancer Res. 2010, 70, 2485–2494. [Google Scholar] [CrossRef] [Green Version]
- Shin, W.-S.; Park, M.K.; Lee, Y.H.; Kim, K.W.; Lee, H.; Lee, S.-T. The catalytically defective receptor protein tyrosine kinase EphA10 promotes tumorigenesis in pancreatic cancer cells. Cancer Sci. 2020, 111, 3292–3302. [Google Scholar] [CrossRef]
- Gärtner, S.; Gunesch, A.; Knyazeva, T.; Wolf, P.; Högel, B.; Eiermann, W.; Ullrich, A.; Knyazev, P.; Ataseven, B. PTK 7 is a transforming gene and prognostic marker for breast cancer and nodal metastasis involvement. PLoS ONE 2014, 9, e84472. [Google Scholar] [CrossRef]
- Bie, J.; Hu, X.; Yang, M.; Shi, X.; Zhang, X.; Wang, Z. PTK7 promotes the malignant properties of cancer stem-like cells in esophageal squamous cell lines. Hum. Cell 2020, 33, 356–365. [Google Scholar] [CrossRef]
- Ataseven, B.; Angerer, R.; Kates, R.; Gunesch, A.; Knyazev, P.; Högel, B.; Becker, C.; Eiermann, W.; Harbeck, N. PTK7 expression in triple-negative breast cancer. Anticancer Res. 2013, 33, 3759–3763. [Google Scholar]
- Shin, W.-S.; Hong, Y.; Lee, H.W.; Lee, S.-T. Catalytically defective receptor protein tyrosine kinase PTK7 enhances invasive phenotype by inducing MMP-9 through activation of AP-1 and NF-κB in esophageal squamous cell carcinoma cells. Oncotarget 2016, 7, 73242–73256. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.-S.; Park, M.K.; Kim, J.H.; Oh, S.W.; Jang, J.Y.; Lee, H.; Lee, S.-T. PTK7, a Catalytically Inactive Receptor Tyrosine Kinase, Increases Oncogenic Phenotypes in Xenograft Tumors of Esophageal Squamous Cell Carcinoma KYSE-30 Cells. Int. J. Mol. Sci. 2022, 23, 2391. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.P.; Qiao, S.; Jiang, S.; Hu, J.L.; Wang, T.T.; Liu, W.W.; Qin, Y.; Wang, Y.N.; Zheng, L.S.; Zhang, J.C.; et al. Protein Tyrosine Kinase 7 Regulates EGFR/Akt Signaling Pathway and Correlates With Malignant Progression in Triple-Negative Breast Cancer. Front. Oncol. 2021, 11, 699889. [Google Scholar] [CrossRef] [PubMed]
- Calzada, V.; Moreno, M.; Newton, J.; González, J.; Fernández, M.; Gambini, J.P.; Ibarra, M.; Chabalgoity, A.; Deutscher, S.; Quinn, T.; et al. Development of new PTK7-targeting aptamer-fluorescent and -radiolabelled probes for evaluation as molecular imaging agents: Lymphoma and melanoma in vivo proof of concept. Bioorg. Med. Chem. 2017, 25, 1163–1171. [Google Scholar] [CrossRef]
- Yan, M.; Li, G.; An, J. Discovery of small molecule inhibitors of the Wnt/β-catenin signaling pathway by targeting β-catenin/Tcf4 interactions. Exp. Biol. Med. 2017, 242, 1185–1197. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Onoyama, H.; Kamiya, M.; Kuriki, Y.; Komatsu, T.; Abe, H.; Tsuji, Y.; Yagi, K.; Yamagata, Y.; Aikou, S.; Nishida, M.; et al. Rapid and sensitive detection of early esophageal squamous cell carcinoma with fluorescence probe targeting dipeptidylpeptidase IV. Sci. Rep. 2016, 6, 26399. [Google Scholar] [CrossRef] [Green Version]
- Hirano, H.; Kato, K. Systemic treatment of advanced esophageal squamous cell carcinoma: Chemotherapy, molecular-targeting therapy and immunotherapy. Jpn. J. Clin. Oncol. 2019, 49, 412–420. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Cao, S.; Dong, X.; Rao, S.; Cai, K. Understanding Esophageal Cancer: The Challenges and Opportunities for the Next Decade. Front. Oncol. 2020, 10, 1727. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, A.; Qi, S.; Cheng, S.; Yao, B.; Xu, Y. Protein tyrosine kinase 7 (PTK7) as a predictor of lymph node metastases and a novel prognostic biomarker in patients with prostate cancer. Int. J. Mol. Sci. 2014, 15, 11665–11677. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Yan, L.; Zhang, D.; Guan, X.; Dong, B.; Zhao, M.; Hao, C. PTK7 overexpression in colorectal tumors: Clinicopathological correlation and prognosis relevance. Oncol. Rep. 2016, 36, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Odintsov, I.; Lui, A.J.W.; Sisso, W.J.; Gladstone, E.; Liu, Z.; Delasos, L.; Kurth, R.I.; Sisso, E.M.; Vojnic, M.; Khodos, I.; et al. The Anti-HER3 mAb Seribantumab Effectively Inhibits Growth of Patient-Derived and Isogenic Cell Line and Xenograft Models with Oncogenic NRG1 Fusions. Clin. Cancer Res. 2021, 27, 3154–3166. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.H.; Chan, L.C.; Wang, Y.N.; Chu, Y.Y.; Wang, C.H.; Lee, H.H.; Xia, W.; Shyu, W.C.; Liu, S.P.; Yao, J.; et al. Ephrin receptor A10 monoclonal antibodies and the derived chimeric antigen receptor T cells exert an antitumor response in mouse models of triple-negative breast cancer. J. Biol. Chem. 2022, 298, 101817. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Chang, D.; Cui, Y.; Hu, J.; Gong, M.; Ma, K.; Ding, F.; Liu, Z.H.; Wang, T.Y. New orthotopic implantation model of human esophageal squamous cell carcinoma in athymic nude mice. Thorac. Cancer 2014, 5, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.L.; Zhao, L.; Xi, M.; He, L.R.; Shen, J.X.; Li, Q.Q.; Liu, S.L.; Zhang, P.; Xie, D.; Liu, M.Z. Association of insulin-like growth factor-binding protein-3 with radiotherapy response and prognosis of esophageal squamous cell carcinoma. Chin. J. Cancer 2015, 34, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Boyer, J.Z.; Phillips, G.D.L.; Nitta, H.; Garsha, K.; Admire, B.; Kraft, R.; Dennis, E.; Vela, E.; Towne, P. Activity of trastuzumab emtansine (T-DM1) in 3D cell culture. Breast Cancer Res. Treat. 2021, 188, 65–75. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Geraldo, S.; Simon, A.; Elkhatib, N.; Louvard, D.; Fetler, L.; Vignjevic, D.M. Do cancer cells have distinct adhesions in 3D collagen matrices and in vivo? Eur. J. Cell Biol. 2012, 91, 930–937. [Google Scholar] [CrossRef]
- Sun, J.J.; Li, H.L.; Guo, S.J.; Ma, H.; Liu, S.J.; Liu, D.; Xue, F.X. The Increased PTK7 Expression Is a Malignant Factor in Cervical Cancer. Dis. Markers 2019, 2019, 5380197. [Google Scholar] [CrossRef] [Green Version]
- Lhoumeau, A.C.; Martinez, S.; Boher, J.M.; Monges, G.; Castellano, R.; Goubard, A.; Doremus, M.; Poizat, F.; Lelong, B.; de Chaisemartin, C.; et al. Overexpression of the Promigratory and Prometastatic PTK7 Receptor Is Associated with an Adverse Clinical Outcome in Colorectal Cancer. PLoS ONE 2015, 10, e0123768. [Google Scholar] [CrossRef]
- Meng, L.; Sefah, K.; O’Donoghue, M.B.; Zhu, G.; Shangguan, D.; Noorali, A.; Chen, Y.; Zhou, L.; Tan, W. Silencing of PTK7 in colon cancer cells: Caspase-10-dependent apoptosis via mitochondrial pathway. PLoS ONE 2010, 5, e14018. [Google Scholar] [CrossRef] [PubMed]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In vitro cell migration and invasion assays. J. Vis. Exp. 2014, 88, e51046. [Google Scholar] [CrossRef] [Green Version]
- Friedl, P.; Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef]
- Barillari, G. The Impact of Matrix Metalloproteinase-9 on the Sequential Steps of the Metastatic Process. Int. J. Mol. Sci. 2020, 21, 4526. [Google Scholar] [CrossRef]
- Merino, F.; Pospich, S.; Raunser, S. Towards a structural understanding of the remodeling of the actin cytoskeleton. Semin. Cell Dev. Biol. 2020, 102, 51–64. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis (Review). Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.S.; Song, X.H.; Gan, H.M.; Geng, D.Z.; Yang, J. Activation of Src tyrosine kinase in esophageal carcinoma cells in different regulatory environments and corresponding occurrence mechanism. Genet. Mol. Res. 2016, 15, 15. [Google Scholar] [CrossRef]
- Miyazaki, T.; Kato, H.; Nakajima, M.; Sohda, M.; Fukai, Y.; Masuda, N.; Manda, R.; Fukuchi, M.; Tsukada, K.; Kuwano, H. FAK overexpression is correlated with tumour invasiveness and lymph node metastasis in oesophageal squamous cell carcinoma. Br. J. Cancer 2003, 89, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Shin, W.-S.; Lee, H.W.; Lee, S.-T. Catalytically inactive receptor tyrosine kinase PTK7 activates FGFR1 independent of FGF. FASEB J. 2019, 33, 12960–12971. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Bracher-Manecke, J.C.; Zhao, X.; Davies, N.H.; Zhou, L.; Ai, R.; Oliver, L.; Vallette, F.; Hendricks, D.T. Oncogenic but non-essential role of N-myc downstream regulated gene 1 in the progression of esophageal squamous cell carcinoma. Cancer Biol. Ther. 2013, 14, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Shin, W.-S.; Na, H.W.; Lee, S.-T. Biphasic effect of PTK7 on KDR activity in endothelial cells and angiogenesis. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 2251–2260. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.W.; Shin, W.-S.; Lee, S.-T. Anti-PTK7 Monoclonal Antibodies Inhibit Angiogenesis by Suppressing PTK7 Function. Cancers 2022, 14, 4463. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.T.; Moody, A.; Schorzman, A.N.; Zamboni, W.C. Importance and Considerations of Antibody Engineering in Antibody-Drug Conjugates Development from a Clinical Pharmacologist’s Perspective. Antibodies 2021, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.-S.; Maeng, Y.S.; Jung, J.W.; Min, J.K.; Kwon, Y.G.; Lee, S.-T. Soluble PTK7 inhibits tube formation, migration, and invasion of endothelial cells and angiogenesis. Biochem. Biophys. Res. Commun. 2008, 371, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Lee, Y.H.; Lee, S.-T. Galangin and kaempferol suppress phorbol-12-myristate-13-acetate-induced matrix metalloproteinase-9 expression in human fibrosarcoma HT-1080 cells. Mol. Cells 2015, 38, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An Open Source Plugin for the Quantitative Evaluation and Automated Scoring of Immunohistochemistry Images of Human Tissue Samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Shin, W.-S.; Lee, S.-R.; Kim, S.; Choi, S.-Y.; Lee, S.-T. Anti-PTK7 Monoclonal Antibodies Exhibit Anti-Tumor Activity at the Cellular Level and in Mouse Xenograft Models of Esophageal Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 12195. https://doi.org/10.3390/ijms232012195
Kim JH, Shin W-S, Lee S-R, Kim S, Choi S-Y, Lee S-T. Anti-PTK7 Monoclonal Antibodies Exhibit Anti-Tumor Activity at the Cellular Level and in Mouse Xenograft Models of Esophageal Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2022; 23(20):12195. https://doi.org/10.3390/ijms232012195
Chicago/Turabian StyleKim, Jae Hoon, Won-Sik Shin, Se-Ra Lee, Sanggil Kim, So-Young Choi, and Seung-Taek Lee. 2022. "Anti-PTK7 Monoclonal Antibodies Exhibit Anti-Tumor Activity at the Cellular Level and in Mouse Xenograft Models of Esophageal Squamous Cell Carcinoma" International Journal of Molecular Sciences 23, no. 20: 12195. https://doi.org/10.3390/ijms232012195