
NMR Structure and Biophysical Characterization of Thermophilic Single-Stranded DNA Binding Protein from Sulfolobus Solfataricus



Abstract
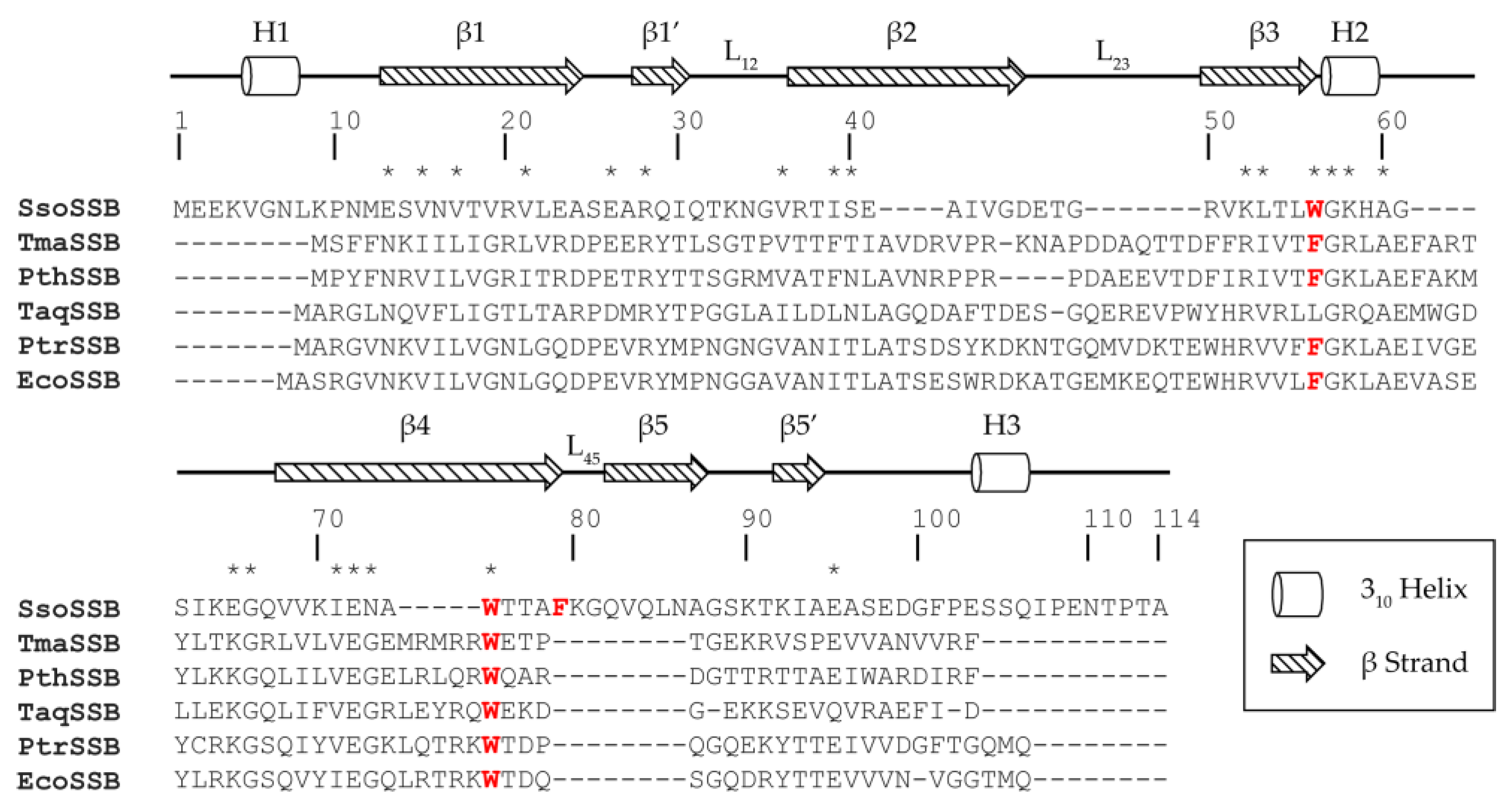
:1. Introduction
2. Results
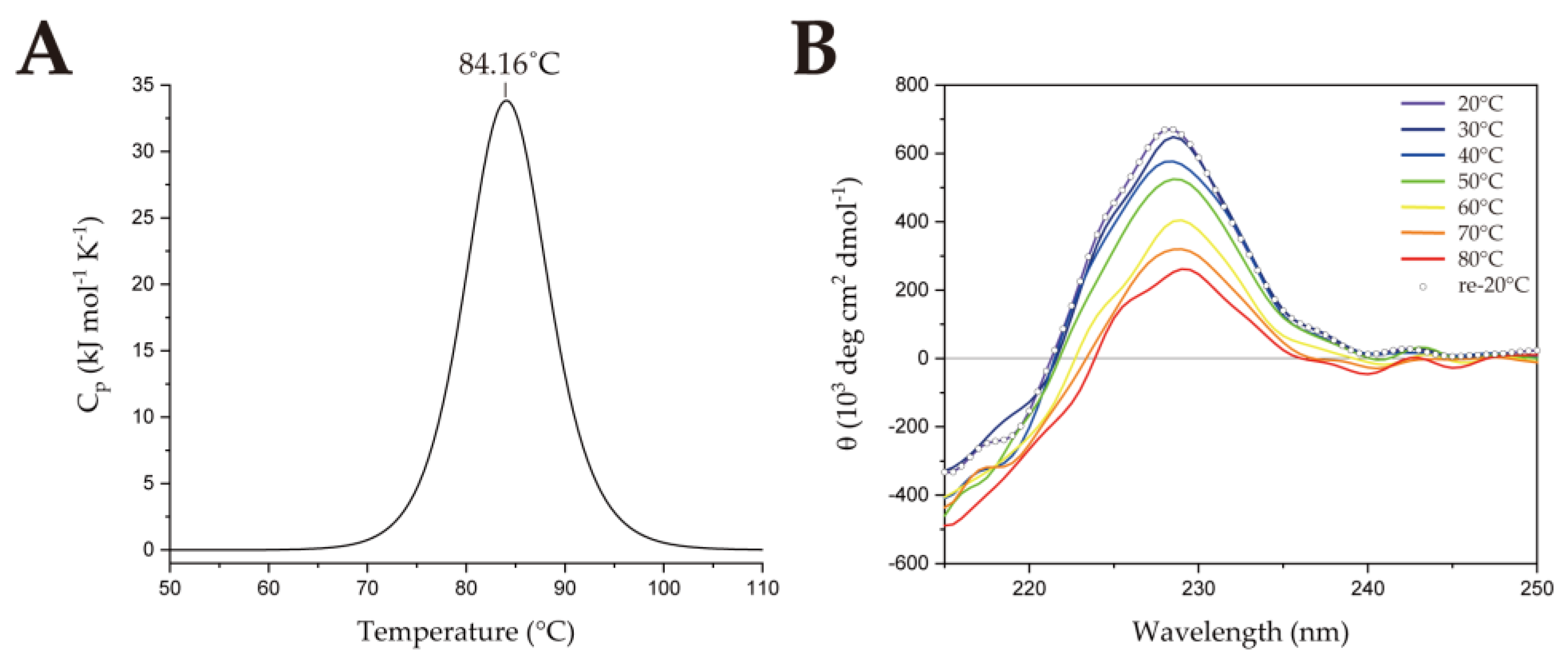
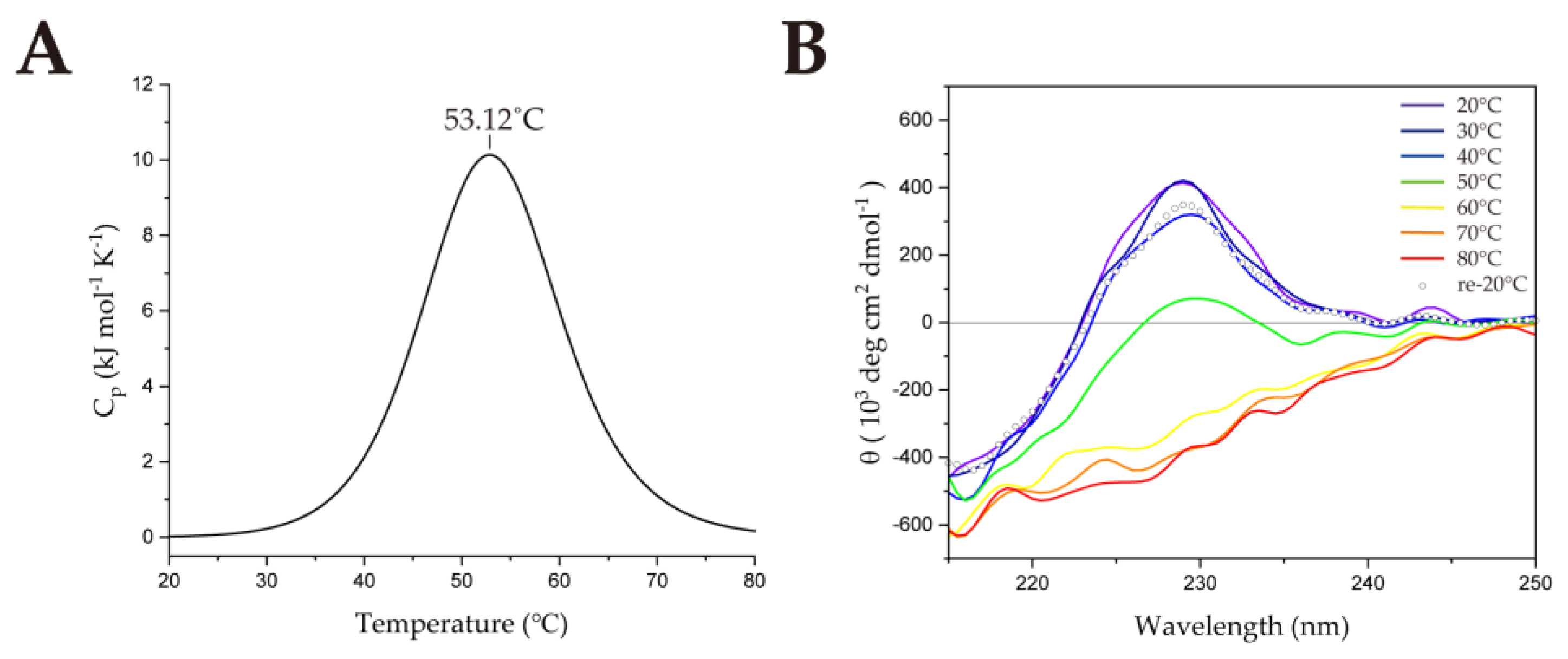
2.1. Thermostability of SsoSSB
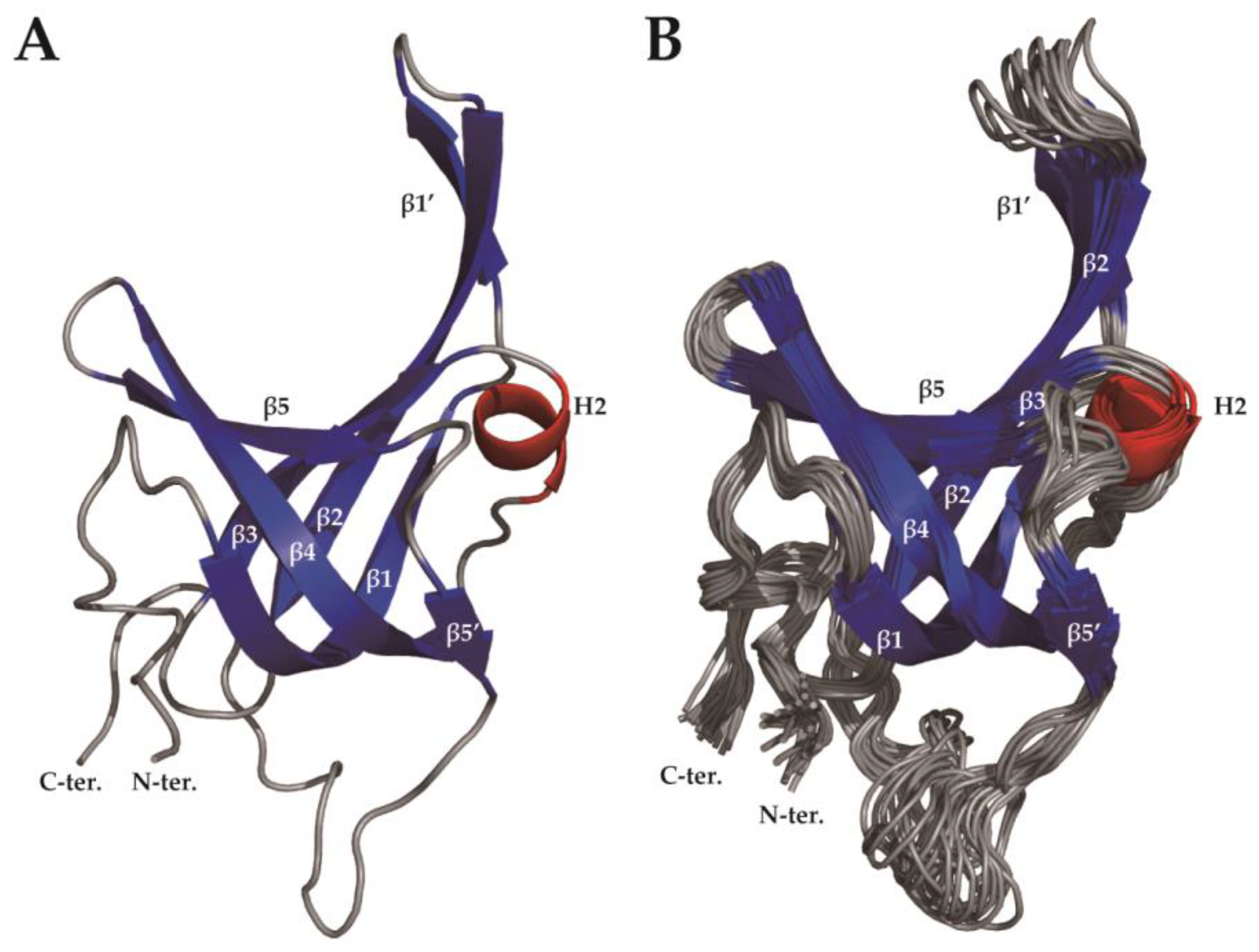
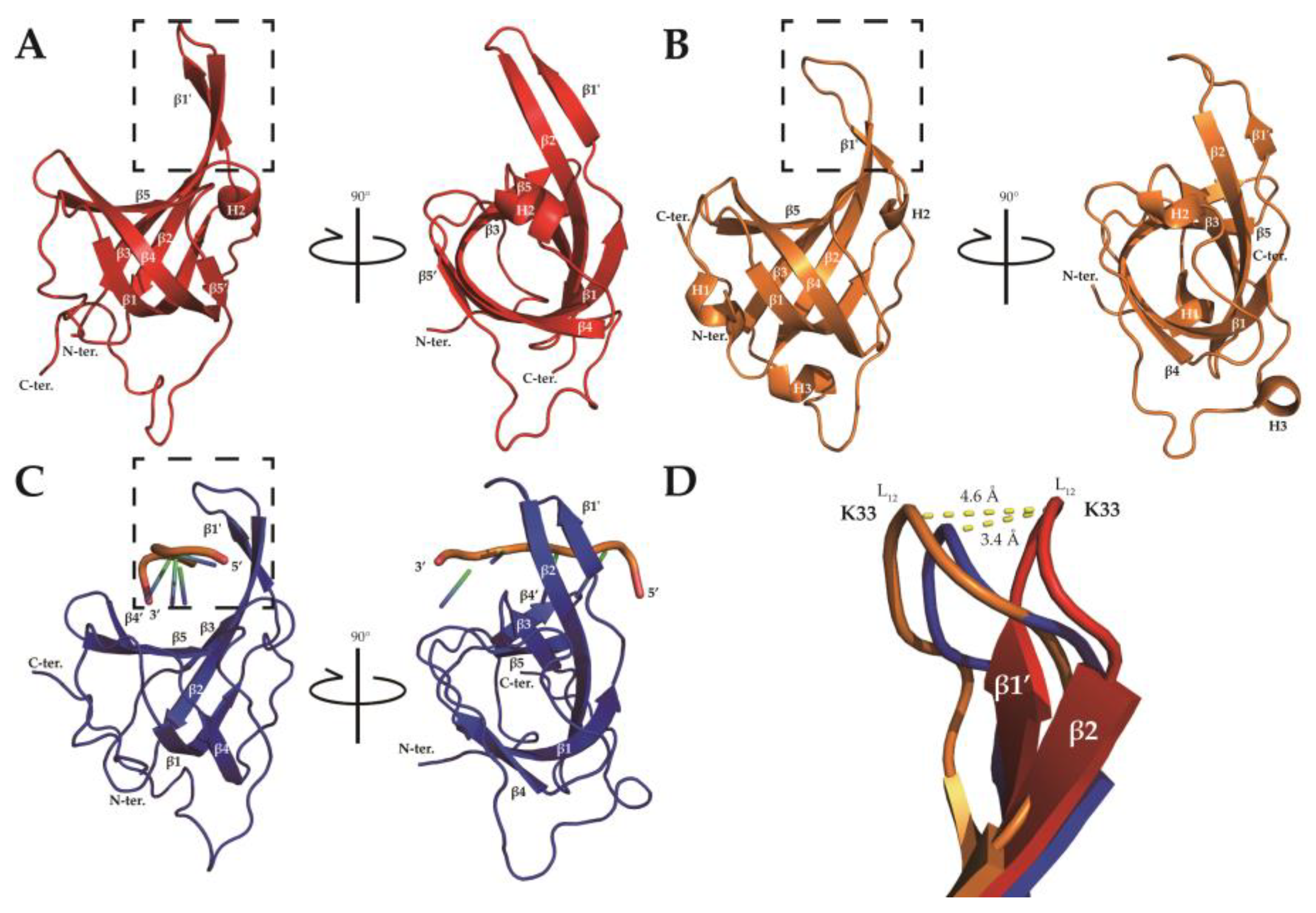
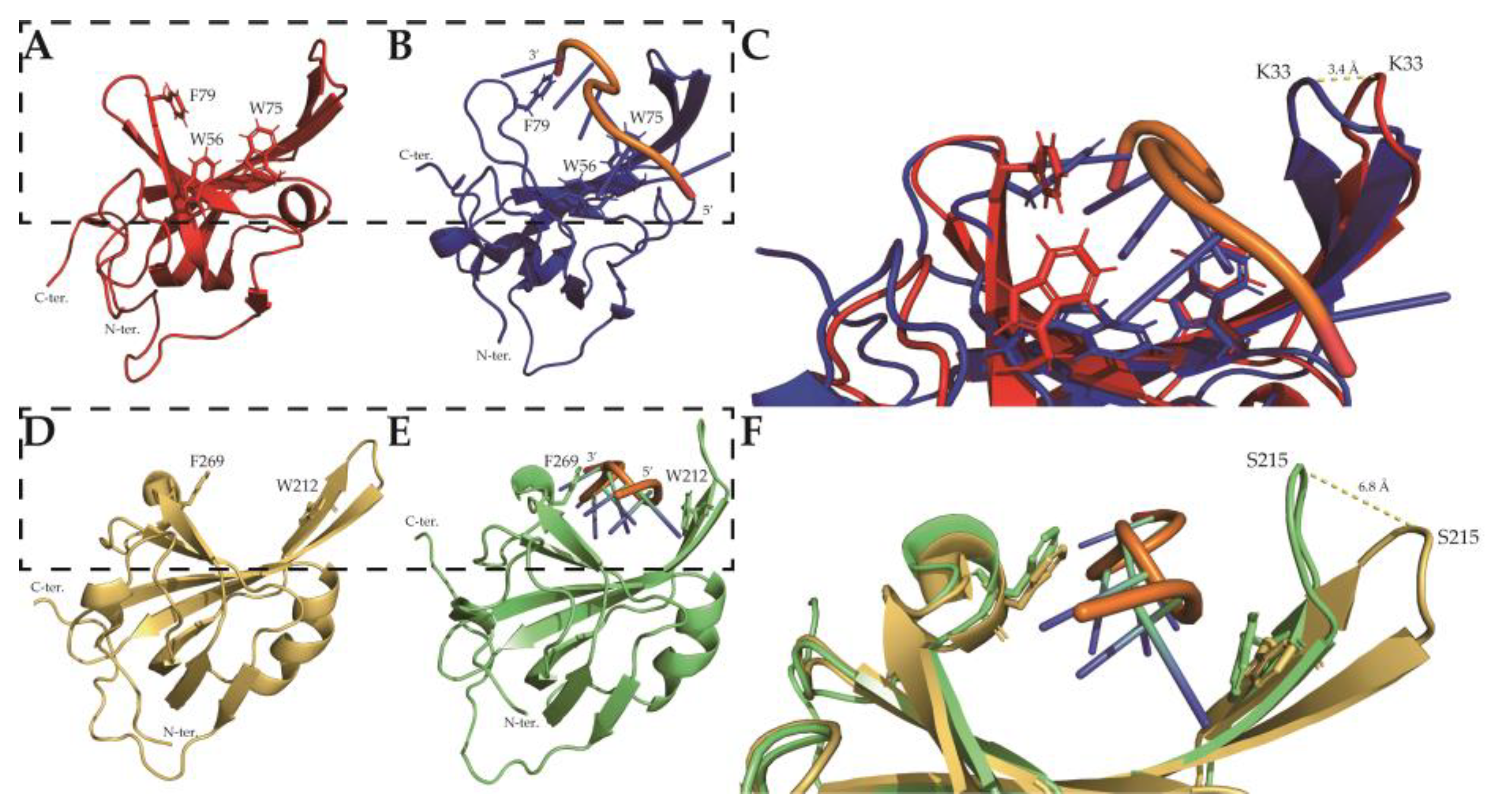
2.2. Solution Structure of SsoSSB1–114 at High Temperature
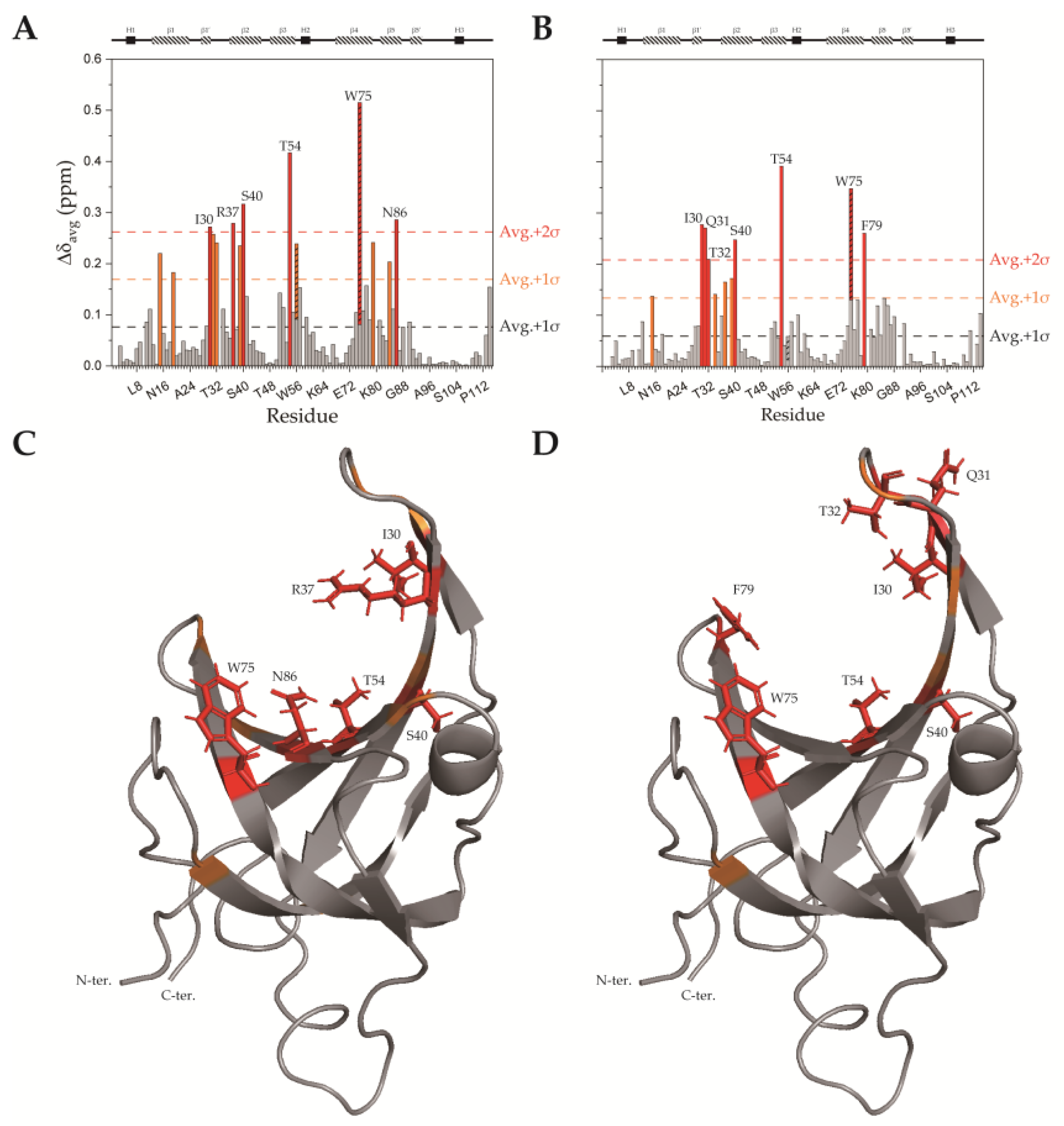
2.3. SsoSSB1–114–DNA Interaction at High Temperature by NMR CSP Analysis
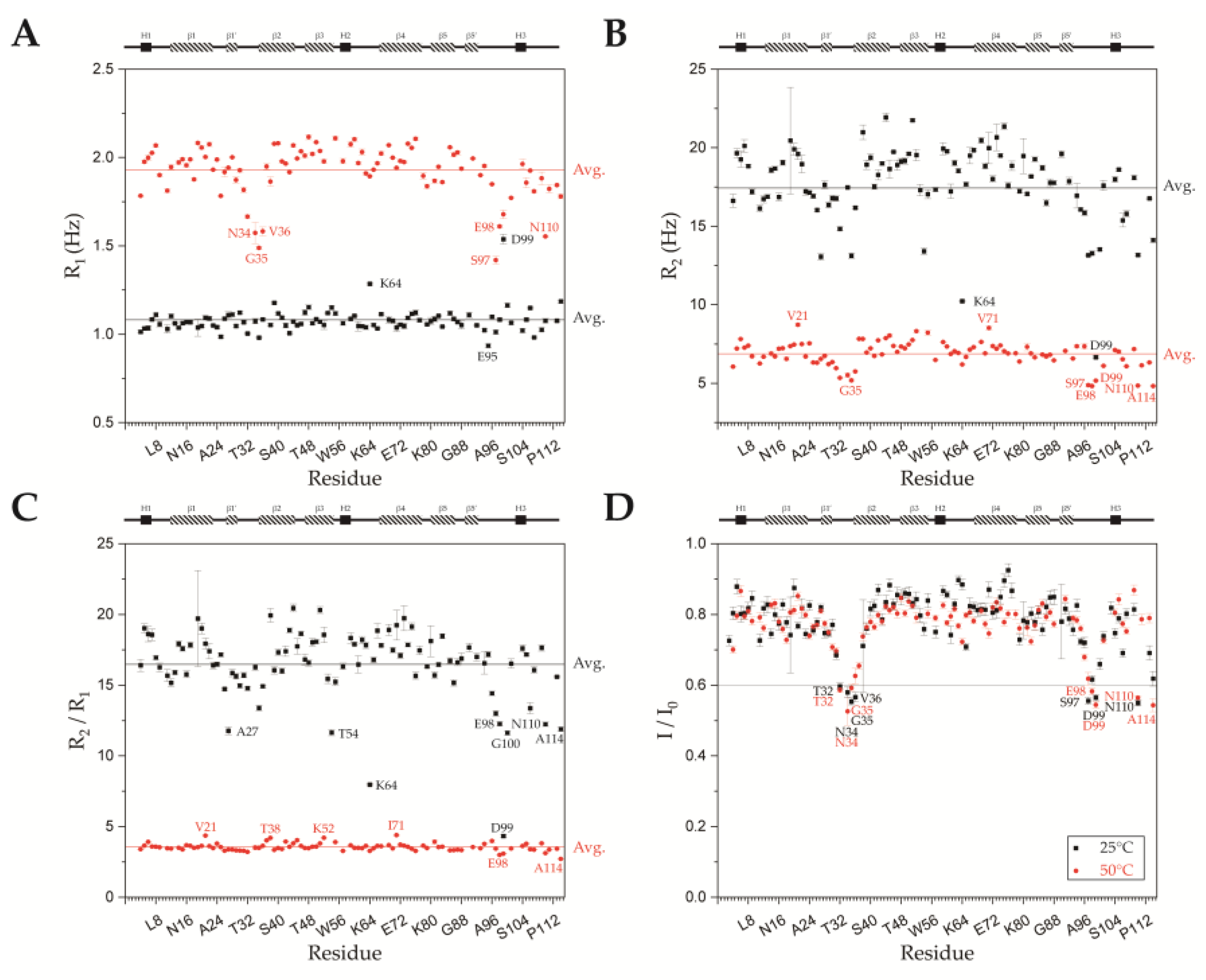
2.4. Backbone Dynamics of SsoSSB1–114 at High Temperature and Room Temperature
2.5. Thermostability and ssDNA Binding Property of SsoSSB12–114
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. NMR Experiments
4.3. Solution Structure Calculation
4.4. NMR Backbone Relaxation Experiment
4.5. Differential Scanning Calorimetry
4.6. Circular Dichroism Spectroscopy
4.7. Isothermal Titration Calorimetry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CSP | chemical shift perturbation |
| CD | circular dichroism |
| DSC | differential scanning calorimetry |
| dA(15) | single-stranded DNA composed of 15 adenines |
| dsDNA | double-stranded DNA |
| hetNOE | heteronuclear Overhauser Effect |
| HSQC | heteronuclear single-quantum coherence |
| ITC | isothermal titration calorimetry |
| NMR | nuclear magnetic resonance |
| NOE | nuclear Overhauser effect |
| OB-fold | oligonucleotide/oligosaccharide binding fold domain |
| PDB | protein data bank |
| RMSD | root-mean-square deviation |
| R1 | spin-lattice relaxation |
| R2 | spin-spin relaxation |
| SSB | single-stranded DNA binding protein |
| ssDNA | single-stranded DNA |
| Sso | Sulfolobus solfataricus |
| Tm | melting temperature |
References
- Brock, T.D.; Brock, K.M.; Belly, R.T.; Weiss, R.L. Sulfolobus: A new genus of sulfur-oxidizing bacteria living at low pH and high temperature. Arch. Mikrobiol. 1972, 84, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Zillig, W.; Stetter, K.O.; Wunderl, S.; Schulz, W.; Priess, H.; Scholz, I. The Sulfolobus-“Caldariella” group: Taxonomy on the basis of the structure of DNA-dependent RNA polymerases. Arch. Microbiol. 1980, 125, 259–269. [Google Scholar] [CrossRef]
- Grogan, D.; Palm, P.; Zillig, W. Isolate B12, which harbours a virus-like element, represents a new species of the archaebacterial genus Sulfolobus, Sulfolobus shibatae, sp. nov. Arch. Microbiol. 1990, 154, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.D.; Kurosawa, N. Saccharolobus caldissimus gen. nov., sp. nov., a facultatively anaerobic iron-reducing hyperthermophilic archaeon isolated from an acidic terrestrial hot spring, and reclassification of Sulfolobus solfataricus as Saccharolobus solfataricus comb. nov. and Sulfolobus shibatae as Saccharolobus shibatae comb. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 1271–1278. [Google Scholar]
- Quehenberger, J.; Shen, L.; Albers, S.-V.; Siebers, B.; Spadiut, O. Sulfolobus–A potential key organism in future biotechnology. Front. Microbiol. 2017, 8, 2474. [Google Scholar] [CrossRef]
- De Rosa, M.; Esposito, E.; Gambacorta, A.; Nicolaus, B.; Bu’Lock, J.D. Effects of temperature on ether lipid composition of Caldariella acidophila. Phytochemistry 1980, 19, 827–831. [Google Scholar] [CrossRef]
- Guagliardi, A.; Napoli, A.; Rossi, M.; Ciaramella, M. Annealing of complementary DNA strands above the melting point of the duplex promoted by an archaeal protein. J. Mol. Biol. 1997, 267, 841–848. [Google Scholar] [CrossRef]
- Forterre, P.; Bergerat, A.; Lopez-Garcia, P. The unique DNA topology and DNA topoisomerases of hyperthermophilic archaea. FEMS Microbiol. Rev. 1996, 18, 237–248. [Google Scholar] [CrossRef]
- Littlechild, J.A. Archaeal enzymes and applications in industrial biocatalysts. Archaea 2015, 2015, 147671. [Google Scholar] [CrossRef] [Green Version]
- Mushegian, A.R.; Koonin, E.V. A minimal gene set for cellular life derived by comparison of complete bacterial genomes. Proc. Natl. Acad. Sci. USA 2002, 93, 10268–10273. [Google Scholar] [CrossRef] [Green Version]
- Ashton, N.W.; Bolderson, E.; Cubeddu, L.; O’Byrne, K.J.; Richard, D.J. Human single-stranded DNA binding proteins are essential for maintaining genomic stability. BMC Mol. Biol. 2013, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, T.; Nyberg, B. Rate of depurination of native deoxyribonucleic acid. Biochemistry 1972, 11, 3610–3618. [Google Scholar] [CrossRef] [PubMed]
- Morten, M.J.; Gamsjaeger, R.; Cubeddu, L.; Kariawasam, R.; Peregrina, J.; Penedo, J.C.; White, M.F. High-affinity RNA binding by a hyperthermophilic single-stranded DNA-binding protein. Extremophiles 2017, 21, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, I.D.; Wadsworth, R.I.M.; Cubeddu, L.; Blankenfeldt, W.; Naismith, J.H.; White, M.F. Insights into ssDNA recognition by the OB fold from a structural and thermodynamic study of Sulfolobus SSB protein. EMBO J. 2003, 22, 2561–2570. [Google Scholar] [CrossRef] [Green Version]
- Gamsjaeger, R.; Kariawasam, R.; Gimenez, A.X.; Touma, C.; Mcllwain, E.; Bernardo, R.E.; Shepherd, N.E.; Ataide, S.F.; Dong, Q.; Richard, D.J.; et al. The structural basis of DNA binding by the single-stranded DNA-binding protein from Sulfolobus solfataricus. Biochem. J. 2015, 465, 337–346. [Google Scholar] [CrossRef]
- Murzin, A.G. OB(oligonucleotide/oligosaccharide binding)-fold: Common structural and functional solution for non-homologous sequences. EMBO J. 1993, 12, 861–867. [Google Scholar] [CrossRef]
- Theobald, D.L.; Mitton-Fry, R.M.; Wuttke, D.S. Nucleic acid recognition by OB-fold proteins. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 115–133. [Google Scholar] [CrossRef] [Green Version]
- Arunkumar, A.I.; Stauffer, M.E.; Bochkareva, E.; Bochkarev, A.; Chazin, W.J. Independent and coordinated functions of replication protein A tandem high affinity single-stranded DNA binding domains. J. Biol. Chem. 2003, 278, 41077–41082. [Google Scholar] [CrossRef] [Green Version]
- Raghunathan, S.; Kozlov, A.G.; Lohman, T.M.; Waksman, G. Structure of the DNA binding domain of E. coli SSB bound to ssDNA. Nat. Struct. Biol. 2000, 7, 648–652. [Google Scholar] [CrossRef]
- Qian, Y.; Johnson, K.A. The human mitochondrial single-stranded DNA-binding protein displays distinct kinetics and thermodynamics of DNA binding and exchange. J. Biol. Chem. 2017, 292, 13068–13084. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, M.; Grot, A.; Wojciechowski, M.; Nowak, M.; Mickiewicz, M.; Kur, J. Characterization of exceptionally thermostable single-stranded DNA-binding proteins from Thermotoga maritima and Thermotoga neapolitana. BMC Microbiol. 2010, 10, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2007, 1, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Fasman, G.D. Circular Dichroism and Conformational Analysis of Biomolecules; Plenum Press: New York, NY, USA, 1996; ISBN 978-03-0645-142-3. [Google Scholar]
- Buchtova, M.; Chaloupkova, R.; Zakrzewska, M.; Vesela, I.; Cela, P.; Barathova, J.; Gudernova, I.; Zajickova, R.; Trantirek, L.; Martin, J.; et al. Instability restricts signaling of multiple fibroblast growth factors. Cell. Mol. Life Sci. 2015, 72, 2445–2459. [Google Scholar] [CrossRef]
- Hider, R.C.; Kupryszewski, G.; Rekowski, P.; Lammek, B. Origin of the positive 225–230 nm circular dichroism band in proteins. Its application to conformational analysis. Biophys. Chem. 1988, 31, 45–51. [Google Scholar] [CrossRef]
- Kelly, S.; Price, N. The use of circular dichroism in the investigation of protein structure and function. Curr. Protein Pept. Sci. 2005, 1, 349–384. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.J.; Lee, W.; Park, C.-J. Resonance assignments and secondary structure of thermophile single-stranded DNA binding protein from Sulfolobus solfataricus at 323K. Biomol. NMR Assign. 2021, 15, 159–164. [Google Scholar] [CrossRef]
- Lee, W.; Petit, C.M.; Cornilescu, G.; Stark, J.L.; Markley, J.L. The AUDANA algorithm for automated protein 3D structure determination from NMR NOE data. J. Biomol. NMR 2016, 65, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Bax, A. Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Schwieters, C.D.; Bermejo, G.A.; Clore, G.M. Xplor-NIH for molecular structure determination from NMR and other data sources. Protein Sci. 2018, 27, 26–40. [Google Scholar] [CrossRef]
- Kremer, W.; Schuler, B.; Harrieder, S.; Geyer, M.; Gronwald, W.; Welker, C.; Jaenicke, R.; Kalbitzer, H.R. Solution NMR structure of the cold-shock protein from the hyperthermophilic bacterium Thermotoga maritima. Eur. J. Biochem. 2001, 268, 2527–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, A.; Bamann, C.; Kremer, W.; Kalbitzer, H.R.; Brunner, E. High-temperature solution NMR structure of TmCsp. Protein Sci. 2004, 13, 342–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Stark, J.L.; Markley, J.L. PONDEROSA-C/S: Client–server based software package for automated protein 3D structure determination. J. Biomol. NMR 2014, 60, 73–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, A.; Tejero, R.; Montelione, G.T. Evaluating protein structures determined by structural genomics consortia. Proteins 2007, 66, 778–795. [Google Scholar] [CrossRef]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced software for biomolecular NMR spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.; Henrick, K.; Nakamura, H. Announcing the worldwide Protein Data Bank. Nat. Struct. Biol. 2003, 10, 980. [Google Scholar] [CrossRef]
- Lee, W.; Rahimi, M.; Lee, Y.; Chiu, A. POKY: A software suite for multidimensional NMR and 3D structure calculation of biomolecules. Bioinformatics 2021, 37, 3041–3042. [Google Scholar] [CrossRef]
- de Medeiros, L.N.; Angeli, R.; Sarzedas, C.G.; Barreto-bergter, E.; Valente, A.P.; Kurtenbach, E.; Almeida, F.C.L. Backbone dynamics of the antifungal Psd1 pea defensin and its correlation with membrane interaction by NMR spectroscopy. Biochim. Biophys. Acta 2010, 1798, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Kay, L.E.; Torchia, D.A.; Bax, A. Backbone dynamics of proteins as studied by 15N inverse detected heteronuclear NMR spectroscopy: Application to staphylococcal nuclease. Biochemistry 1989, 28, 8972–8979. [Google Scholar] [CrossRef]
- Tremblay, M.-L.; Xu, L.; Sarker, M.; Liu, X.-Q.; Rainey, J.K. Characterizing aciniform silk repetitive domain backbone dynamics and hydrodynamic modularity. Int. J. Mol. Sci. 2016, 17, 1305. [Google Scholar] [CrossRef] [Green Version]
- Baber, J.L.; Szabo, A.; Tjandra, N. Analysis of slow interdomain motion of macromolecules using NMR relaxation data. J. Am. Chem. Soc. 2001, 123, 3953–3959. [Google Scholar] [CrossRef] [PubMed]
- DiDonato, M.; Krishna, S.S.; Schwarzenbacher, R.; McMullan, D.; Jaroszewski, L.; Miller, M.D.; Abdubek, P.; Agarwalla, S.; Ambing, E.; Axelrod, H.; et al. Crystal structure of a single-stranded DNA-binding protein (TM0604) from Thermotoga maritima at 2.60 Å resolution. Proteins 2006, 63, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Sugiki, T.; Kobayashi, N.; Fujiwara, T. Modern technologies of solution nuclear magnetic resonance spectroscopy for three-dimensional structure determination of proteins open avenues for life scientists. Comput. Struct. Biotechnol. J. 2017, 15, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Fowler, N.J.; Sljoka, A.; Williamson, M.P. A method for validating the accuracy of NMR protein structures. Nat. Commun. 2020, 11, 6321. [Google Scholar] [CrossRef] [PubMed]
- Bochkareva, E.; Belegu, V.; Korolev, S.; Bochkarev, A. Structure of the major single-stranded DNA-binding domain of replication protein A suggests a dynamic mechanism for DNA binding. EMBO J. 2001, 20, 612–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochkarev, A.; Pfuetzner, R.A.; Edwards, A.M.; Frappier, L. Structure of the single-stranded-DNA-binding domain of replication protein A bound to DNA. Nature 1997, 385, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Solomon, I. Relaxation processes in a system of two spins. Phys. Rev. 1955, 99, 559–565. [Google Scholar] [CrossRef]
- Lee, Y.; Jang, A.; Jeong, M.-C.; Park, N.; Park, J.; Lee, W.C.; Cheong, C.; Kim, Y. Structural characterization of an ACP from Thermotoga maritima: Insights into hyperthermal adaptation. Int. J. Mol. Sci. 2020, 21, 2600. [Google Scholar] [CrossRef] [Green Version]
- Brininger, C.; Spradlin, S.; Cobani, L.; Evilia, C. The more adaptive to change, the more likely you are to survive: Protein adaptation in extremophiles. Semin. Cell Dev. Biol. 2018, 84, 158–169. [Google Scholar] [CrossRef]
- Hait, S.; Mallik, S.; Basu, S.; Kundu, S. Finding the generalized molecular principles of protein thermal stability. Proteins 2020, 88, 788–808. [Google Scholar] [CrossRef]
- Fang, X.; Cui, Q.; Tong, Y.; Feng, Y.; Shan, L.; Huang, L.; Wang, J. A stabilizing alpha/beta-hydrophobic core greatly contributes to hyperthermostability of archaeal [P62A]Ssh10b. Biochemistry 2008, 47, 11212–11221. [Google Scholar] [CrossRef] [PubMed]
- Tych, K.M.; Batchelor, M.; Hoffmann, T.; Wilson, M.C.; Hughes, M.L.; Paci, E.; Brockwell, D.J.; Dougan, L. Differential effects of hydrophobic core packing residues for thermodynamic and mechanical stability of a hyperthermophilic protein. Langmuir 2016, 32, 7392–7402. [Google Scholar] [CrossRef] [Green Version]
- Perry, L.J.; Wetzel, R. Disulfide bond engineered into T4 lysozyme: Stabilization of the protein toward thermal inactivation. Science 1984, 226, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Gokhale, R.S.; Agarwalla, S.; Francis, V.S.; Santi, D.V.; Balaram, P. Thermal stabilization of thymidylate synthase by engineering two disulfide bridges across the dimer interface. J. Mol. Biol. 1994, 235, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Kellis, J.T.; Todd, R.J.; Arnold, F.H. Protein stabilization by engineered metal chelation. Bio/Technology 1991, 9, 994–995. [Google Scholar] [CrossRef]
- Eijsink, V.G.H.; Vriend, G.; van der Vinne, B.; Hazes, B.; van den Burg, B.; Venema, G. Effects of changing the interaction between subdomains on the thermostability of Bacillus neutral proteases. Proteins 1992, 14, 224–236. [Google Scholar] [CrossRef]
- Kwon, W.S.; Da Silva, N.A.; Kellis, J.T., Jr. Relationship between thermal stability, degradation rate and expression yield of barnase variants in the periplasm of Escherichia coli. Protein Eng. 1996, 9, 1197–1202. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, H.-F.; Liu, S.-R.; Yan, X.; Song, K.-J. Predicting protein-protein interactions from matrix-based protein sequence using convolution neural network and feature-selective rotation forest. Sci. Rep. 2019, 9, 9848. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Yarlagadda, R.; Reed, C.C. Structure based thermostability prediction models for protein single point mutations with machine learning tools. PLoS ONE 2015, 10, e0138022. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Ma, Z.; Yang, D.; Li, X.; Zhang, J.; Li, Y. A method for prediction of thermophilic protein based on reduced amino acids and mixed features. Front. Bioeng. Biotechnol. 2020, 8, 285. [Google Scholar] [CrossRef]
- Jia, L.-L.; Sun, T.-T.; Wang, Y.; Shen, Y. A machine learning study on the thermostability prediction of (R)-ω-selective amine transaminase from Aspergillus terreus. Biomed. Res. Int. 2021, 2021, 2593748. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowski, S.; Olszewski, M.; Piątek, R.; Kur, J. Novel thermostable ssDNA-binding proteins from Thermus thermophilus and T. aquaticus-expression and purification. Protein Expr. Purif. 2002, 26, 131–138. [Google Scholar] [CrossRef]
- Kang, J.; Yeom, G.; Jang, H.; Oh, J.; Park, C.-J.; Kim, M.G. Development of replication protein A-Conjugated gold nanoparticles for highly sensitive detection of disease biomarkers. Anal. Chem. 2019, 91, 10001–10007. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.N.; Roni, M.A. Challenges of storage and stability of mRNA-based COVID-19 vaccines. Vaccines 2021, 9, 1033. [Google Scholar] [CrossRef]
- Gamsjaeger, R.; Kariawasam, R.; Touma, C.; Kwan, A.H.; White, M.F.; Cubeddu, L. Backbone and side-chain 1H, 13C and 15N resonance assignments of the OB domain of the single stranded DNA binding protein from Sulfolobus solfataricus and chemical shift mapping of the DNA-binding interface. Biomol. NMR Assign. 2014, 8, 243–246. [Google Scholar] [CrossRef]
- Lee, W.; Cornilescu, B.; Dashti, H.; Eghbalnia, H.R.; Tonelli, M.; Westler, W.M.; Butcher, S.E.; Henzler-Wildman, K.A.; Markley, J.L. Integrative NMR for biomolecular research. J. Biomol. NMR 2016, 64, 307–332. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Restraints 1 | Value | ||
|---|---|---|---|
| Total NMR Constraints | 1086 | ||
| Distance Constraints | |||
| Intra Residue (|i–j| = 0) | 259 | ||
| Sequential Residue (|i–j| = 1) | 225 | ||
| Medium Range (1 < |i–j| ≤ 5) | 85 | ||
| Long Range (|i–j| > 5) | 291 | ||
| Hydrogen Bond | 33 | ||
| Dihedral Angle Constraints | |||
| Φ | 96 | ||
| Ψ | 97 | ||
| Pairwise RMSD (Å) 2 | |||
| Backbone Atoms 3 | 0.974 ± 0.044 | ||
| Heavy Atoms 3 | 1.751 ± 0.054 | ||
| Ramachandran Plot Summary from PROCHECK (%) 2 | |||
| Most Favored Regions | 94.9 | ||
| Additionally Allowed Regions | 3.9 | ||
| Generously Allowed Regions | 1.2 | ||
| Disallowed Regions | 0.0 | ||
| wwPDB NMR Structure Validation 4 | |||
| Clashscore | 8 | ||
| Ramachandran Outliers | 2.0% | ||
| Sidechain Outliers | 1.0% | ||
| Average Number of Violations Per Conformer 5 | |||
| Distance Violations (>0.5 Å) | 0 | ||
| Angle Violations (>5°) | 0 | ||
| Repulsive Violations | 0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.J.; Kim, J.; Lee, Y.; Lee, W.; Park, C.-J. NMR Structure and Biophysical Characterization of Thermophilic Single-Stranded DNA Binding Protein from Sulfolobus Solfataricus. Int. J. Mol. Sci. 2022, 23, 3099. https://doi.org/10.3390/ijms23063099
Yang MJ, Kim J, Lee Y, Lee W, Park C-J. NMR Structure and Biophysical Characterization of Thermophilic Single-Stranded DNA Binding Protein from Sulfolobus Solfataricus. International Journal of Molecular Sciences. 2022; 23(6):3099. https://doi.org/10.3390/ijms23063099
Chicago/Turabian StyleYang, Min June, Jinwoo Kim, Yeongjoon Lee, Woonghee Lee, and Chin-Ju Park. 2022. "NMR Structure and Biophysical Characterization of Thermophilic Single-Stranded DNA Binding Protein from Sulfolobus Solfataricus" International Journal of Molecular Sciences 23, no. 6: 3099. https://doi.org/10.3390/ijms23063099