
An Oligomannuronic Acid-Sialic Acid Conjugate Capable of Inhibiting Aβ42 Aggregation and Alleviating the Inflammatory Response of BV-2 Microglia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
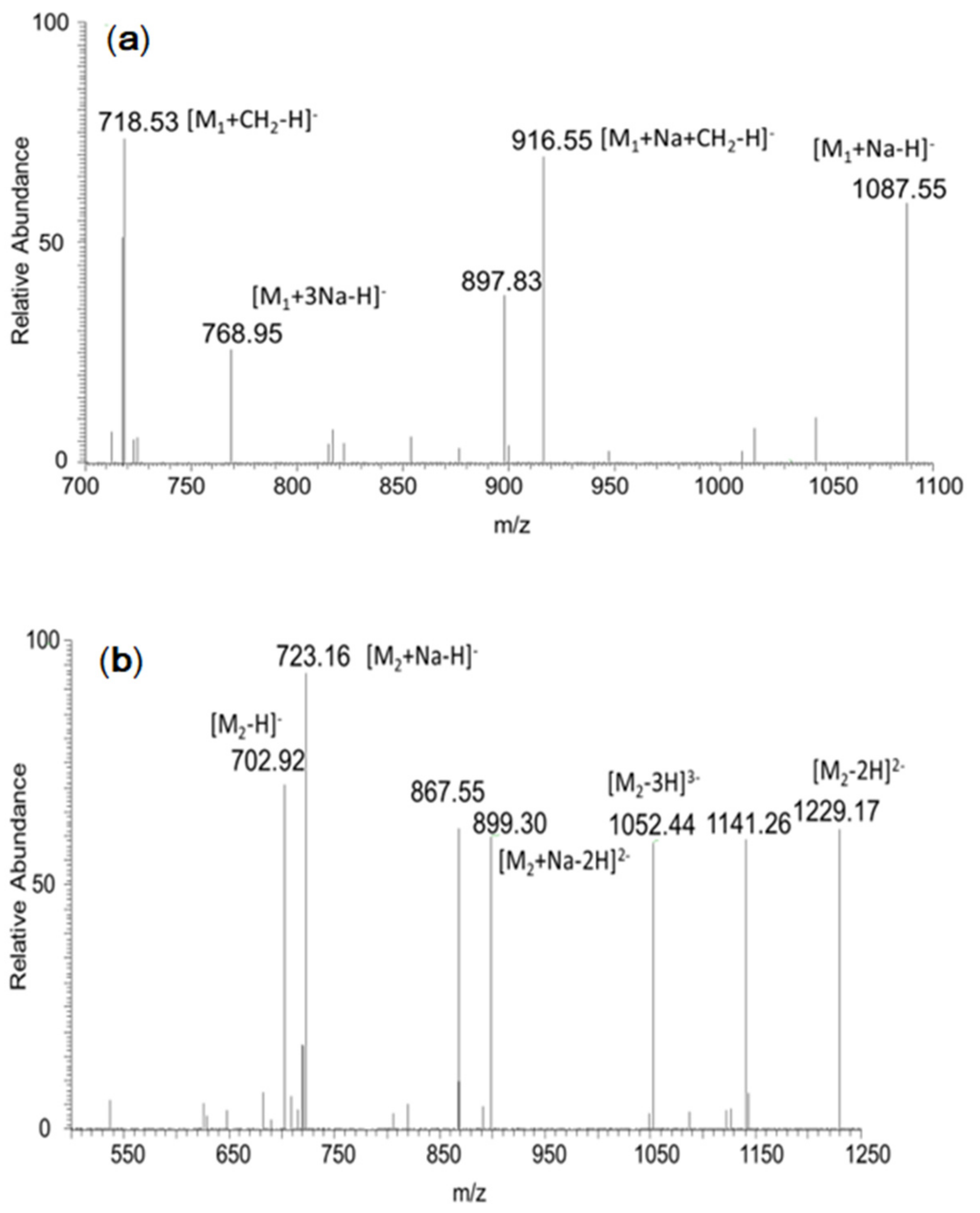
2.1. Synthesis and Characterization of MOS-Sia
2.2. Effect of MOS-Sia on the Sedimentation of Aβ42
2.3. Thioflavin T (ThT) Analysis of Interaction between Aβ42 and MOS-Sia
2.4. Circular Dichroism (CD) Analysis of Interaction between Aβ42 and MOS-Sia3
2.5. Effect of MOS-Sia3 on the Activity of BV-2 Cells Treated with Aβ42
2.6. Effect of MOS-Sia3 on the Release of Inflammatory Factors in BV-2 Cells Induced by Aβ42
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of MOS
4.3. Synthesis of MOS-Sia
4.3.1. Synthesis of MOS-Sia by Reductive Amination
4.3.2. Synthesis of MOS-Sia by Reductive Amidation
4.3.3. Synthesis of MOS-Sia by Thiol Condensation
4.4. Analytical Methods
4.5. Interaction of MOS-Sia with Aβ42 and Metal Ions
4.6. Th-T Staining Analysis
4.7. Effect of MOS-Sia3 on the Cell Viability of BV-2 Cells and Cytokine Release Induced by Aβ42
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ThT | Thioflavin T |
| AD | Alzheimer’s disease |
| MOS | oligo mannuronic acid |
| Sia | sialic acid |
| PSA | polysialic acid |
| Aβ | Amyloid β |
| CD | circular dichroism |
| PBS | phosphate buffered saline |
| IL-1β | interleukin-1β |
| IL-6 | interleukin-6 |
| TNF-α | tumor necrosis factor-α |
References
- Sošić, M.; Vuletic, V.; Tomić, Z.; Bogdanovic, N. Diagnostic and therapeutic approach to a patient with cognitive impairment. Med. Flum. 2018, 54, 140–154. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.; Lee, G.; Ritter, A.; Sabbagh, M.; Zhong, K. Alzheimer’s disease drug development pipeline: 2019. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2019, 5, 272–293. [Google Scholar] [CrossRef] [PubMed]
- Hukins, D.; MacLeod, U.; Boland, J.W. Identifying potentially inappropriate prescribing in older people with dementia: A systematic review. Eur. J. Clin. Pharmacol. 2019, 75, 467–481. [Google Scholar] [CrossRef] [Green Version]
- Tagliavini, F.; Tiraboschi, P.; Federico, A. Alzheimer’s disease: The controversial approval of Aducanumab. Neurol. Sci. 2021, 42, 3069–3070. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.A.; Pedram, K.; Malaker, S.A.; Batista, P.J.; Smith, B.A.; Johnson, A.G.; George, B.M.; Majzoub, K.; Villalta, P.W.; Carette, J.E.; et al. Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell 2021, 184, 3109–3124.e22. [Google Scholar] [CrossRef]
- Patel, D.A.; Henry, J.E.; Good, T.A. Attenuation of β-amyloid-induced toxicity by sialic-acid-conjugated dendrimers: Role of sialic acid attachment. Brain Res. 2007, 1161, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Cowan, C.B.; Coté, G.L.; Good, T.A. Development of photocrosslinked sialic acid containing polymers for use in amyloid beta toxicity attenuation. Biomaterials 2008, 29, 3408–3414. [Google Scholar] [CrossRef] [Green Version]
- Griciuc, A.; Serrano-Pozo, A.; Parrado, A.R.; Lesinski, A.N.; Asselin, C.N.; Mullin, K.; Hooli, B.; Choi, S.H.; Hyman, B.T.; Tanzi, R.E. Alzheimer’s Disease Risk Gene CD33 Inhibits Microglial Uptake of Amyloid Beta. Neuron 2013, 78, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Thiesler, H.; Beimdiek, J.; Hildebrandt, H. Polysialic acid and Siglec-E orchestrate negative feedback regulation of microglia activation. Cell. Mol. Life Sci. 2020, 78, 1637–1653. [Google Scholar] [CrossRef]
- Hu, X.; Jiang, X.; Hwang, H.; Liu, S.; Guan, H. Antitumor activities of alginate-derived oligosaccharides and their sulphated substitution derivatives. Eur. J. Phycol. 2004, 39, 67–71. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kurachi, M.; Yamaguchi, K.; Oda, T. Induction of Multiple Cytokine Secretion from RAW264.7 Cells by Alginate Oligosaccharides. Biosci. Biotechnol. Biochem. 2007, 71, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Hu, J.; Li, J.; Yang, Z.; Xin, X.; Wang, J.; Ding, J.; Geng, M. Effect of acidic oligosaccharide sugar chain on scopolamine-induced memory impairment in rats and its related mechanisms. Neurosci. Lett. 2005, 374, 222–226. [Google Scholar] [CrossRef]
- Wang, X.J.; Chen, X.H.; Yang, X.Y.; Geng, M.Y.; Wang, L.M. Acidic oligosaccharide sugar chain, a marine-derived oligosaccharide, activates human glial cell line-derived neurotrophic factor signaling. Neurosci. Lett. 2017, 417, 176–180. [Google Scholar] [CrossRef]
- Wang, X.; Sun, G.; Feng, T.; Zhang, J.; Huang, X.; Wang, T.; Xie, Z.; Chu, X.; Yang, J.; Wang, H.; et al. Sodium oligomannate therapeutically remodels gut microbiota and suppresses gut bacterial amino acids-shaped neuroinflammation to inhibit Alzheimer’s disease progression. Cell Res. 2019, 29, 787–803. [Google Scholar] [CrossRef]
- Salloway, S.; Sperling, R.; Keren, R.; Porsteinsson, A.; Van Dyck, C.H.; Tariot, P.N.; Gilman, S.; Arnold, D.; Abushakra, S.; Hernandez, C.; et al. A phase 2 randomized trial of ELND005, scyllo-inositol, in mild to moderate Alzheimer disease. Neurology 2011, 77, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, T.; Lieblein, T.; Pohland, M.; Kalden, E.; Freund, P.; Zangl, R.; Grewal, R.; Heilemann, M.; Eckert, G.P.; Morgner, N.; et al. Peptidomimetics That Inhibit and Partially Reverse the Aggregation of Aβ1–42. Biochemistry 2017, 56, 4840–4849. [Google Scholar] [CrossRef]
- Xu, P.; Zhang, M.; Sheng, R.; Ma, Y. Synthesis and biological evaluation of deferiprone-resveratrol hybrids as antioxidants, Aβ 1–42 aggregation inhibitors and metal-chelating agents for Alzheimer’s disease. Eur. J. Med. Chem. 2017, 127, 174–186. [Google Scholar] [CrossRef]
- Krishnan, H.S.; Bernard-Gauthier, V.; Placzek, M.S.; Dahl, K.; Narayanaswami, V.; Livni, E.; Chen, Z.; Yang, J.; Collier, T.L.; Ran, C.; et al. Metal Protein-Attenuating Compound for PET Neuroimaging: Synthesis and Preclinical Evaluation of [11C]PBT2. Mol. Pharm. 2018, 15, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Murthy, R.V.; Bharate, P.; Gade, M.; Sangabathuni, S.; Kikkeri, R. Effect of Transition Metals on Polysialic Acid Structure and Functions. ChemMedChem 2016, 11, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Younan, N.D.; Viles, J.H. A Comparison of Three Fluorophores for the Detection of Amyloid Fibers and Prefibrillar Oligomeric Assemblies. ThT (Thioflavin T); ANS (1-Anilinonaphthalene-8-sulfonic Acid); and bisANS (4,4′-Dianilino-1,1′-binaphthyl-5,5′-disulfonic Acid). Biochemistry 2015, 54, 4297–4306. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Zhao, Y.; Liu, X. High Aβ load may cause microglial cell dysfunction and reduced nuclear repressor element-1 silencing transcription factor (REST) expression which might be ascribed to its degradation by ubiquitination. Ann. Transl. Med. 2019, 7, 338. [Google Scholar] [CrossRef]
- Rawal, P.; Zhao, L. Sialometabolism in Brain Health and Alzheimer’s Disease. Front. Neurosci. 2021, 15, 308. [Google Scholar] [CrossRef]
- Schnaar, R.L.; Gerardy-Schahn, R.; Hildebrandt, H. Sialic Acids in the Brain: Gangliosides and Polysialic Acid in Nervous System Development, Stability, Disease, and Regeneration. Physiol. Rev. 2014, 94, 461–518. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ding, F.; Xiao, L.; Shi, R.; Wang, H.; Han, W.; Huang, Z. Food-Derived Antioxidant Polysaccharides and Their Pharmacological Potential in Neurodegenerative Diseases. Nutrients 2017, 9, 778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Jin, L. The Structure-Activity Relationship of Glycosaminoglycans and Their Analogues with β-Amyloid Peptide. Protein Pept. Lett. 2016, 23, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Paul, T.; Kelly, H.; Zuchniarz, J.; Ahmed, T.; Prabhakar, R. Design of heparin oligosaccharide based molecules for inhibition of Alzheimer amyloid beta (Aβ40) aggregation. Can. J. Chem. 2016, 94, 1090–1098. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Liu, G.; Yang, Y.; Shao, K.; Wang, Y.; Liu, W.; Han, B. N-Acetyl chitooligosaccharides attenuate amyloid β-induced damage in animal and cell models of Alzheimer’s disease. Process. Biochem. 2019, 84, 161–171. [Google Scholar] [CrossRef]
- Ai, L.; Chung, Y.-C.; Lin, S.-Y.; Jeng, K.-C.G.; Lai, P.F.-H.; Xiong, Z.-Q.; Wang, G. Carrageenan polysaccharides and oligosaccharides with distinct immunomodulatory activities in murine microglia BV-2 cells. Int. J. Biol. Macromol. 2018, 120, 633–640. [Google Scholar] [CrossRef]
- Zhu, Z.; Liu, Q.; Chen, P.; Xu, X.; Ni, J.; Yang, S.; Song, Y. Seleno-polymannuronate synthesis and resistance to oxidation and apoptosis in Alzheimer’s disease cells. Chem. J. Chin. Univ. 2013, 34, 115–122. [Google Scholar]
- Li, Q.; Zeng, Y.; Wang, L.; Guan, H.; Li, C.; Zhang, L. The heparin-like activities of negatively charged derivatives of low-molecular-weight polymannuronate and polyguluronate. Carbohydr. Polym. 2016, 155, 313–320. [Google Scholar] [CrossRef]
- Yin, T.; Yang, L.; Liu, Y.; Zhou, X.; Sun, J.; Liu, J. Sialic acid (SA)-modified selenium nanoparticles coated with a high blood–brain barrier permeability peptide-B6 peptide for potential use in Alzheimer’s disease. Acta Biomater. 2015, 25, 172–183. [Google Scholar] [CrossRef]
- Dhavale, D.D. Sialic Acid Conjugated Chitosan for the Attenuation of Amyloid-Beta Toxicity. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2009. [Google Scholar]
- Li, J.; Zhang, J.; Zhang, Q.; Bai, Z.; Zhao, Q.; He, D.; Wang, Z.; Chen, Y.; Liu, B. Syntheses and anti-cancer activity of CO-releasing molecules with targeting galactose receptors. Org. Biomol. Chem. 2018, 16, 8115–8129. [Google Scholar] [CrossRef] [PubMed]
- Hanh, V.T.; Kobayashi, Y.; Maebuchi, M.; Nakamori, T.; Tanaka, M.; Matsui, T. Quantitative mass spectrometric analysis of dipeptides in protein hydrolysate by a TNBS derivatization-aided standard addition method. Food Chem. 2016, 190, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D., Jr.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–90. [Google Scholar] [CrossRef]
- Lei, Y.; Wu, M.; Wang, J.; Zhang, H.; Zhan, X.; Sun, Z.; Wu, J. Preparation and property of a biantenna macromolecule based on polysialic acid. Int. J. Biol. Macromol. 2019, 155, 1342–1349. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Wu, M.; Zhang, H.; Zhan, X.; Wu, N. An Oligomannuronic Acid-Sialic Acid Conjugate Capable of Inhibiting Aβ42 Aggregation and Alleviating the Inflammatory Response of BV-2 Microglia. Int. J. Mol. Sci. 2021, 22, 12338. https://doi.org/10.3390/ijms222212338
Wu J, Wu M, Zhang H, Zhan X, Wu N. An Oligomannuronic Acid-Sialic Acid Conjugate Capable of Inhibiting Aβ42 Aggregation and Alleviating the Inflammatory Response of BV-2 Microglia. International Journal of Molecular Sciences. 2021; 22(22):12338. https://doi.org/10.3390/ijms222212338
Chicago/Turabian StyleWu, Jianrong, Miaosen Wu, Hongtao Zhang, Xiaobei Zhan, and Nian Wu. 2021. "An Oligomannuronic Acid-Sialic Acid Conjugate Capable of Inhibiting Aβ42 Aggregation and Alleviating the Inflammatory Response of BV-2 Microglia" International Journal of Molecular Sciences 22, no. 22: 12338. https://doi.org/10.3390/ijms222212338