Novel Biomarkers in the Diagnosis of Chronic Kidney Disease and the Prediction of Its Outcome

 ,
, 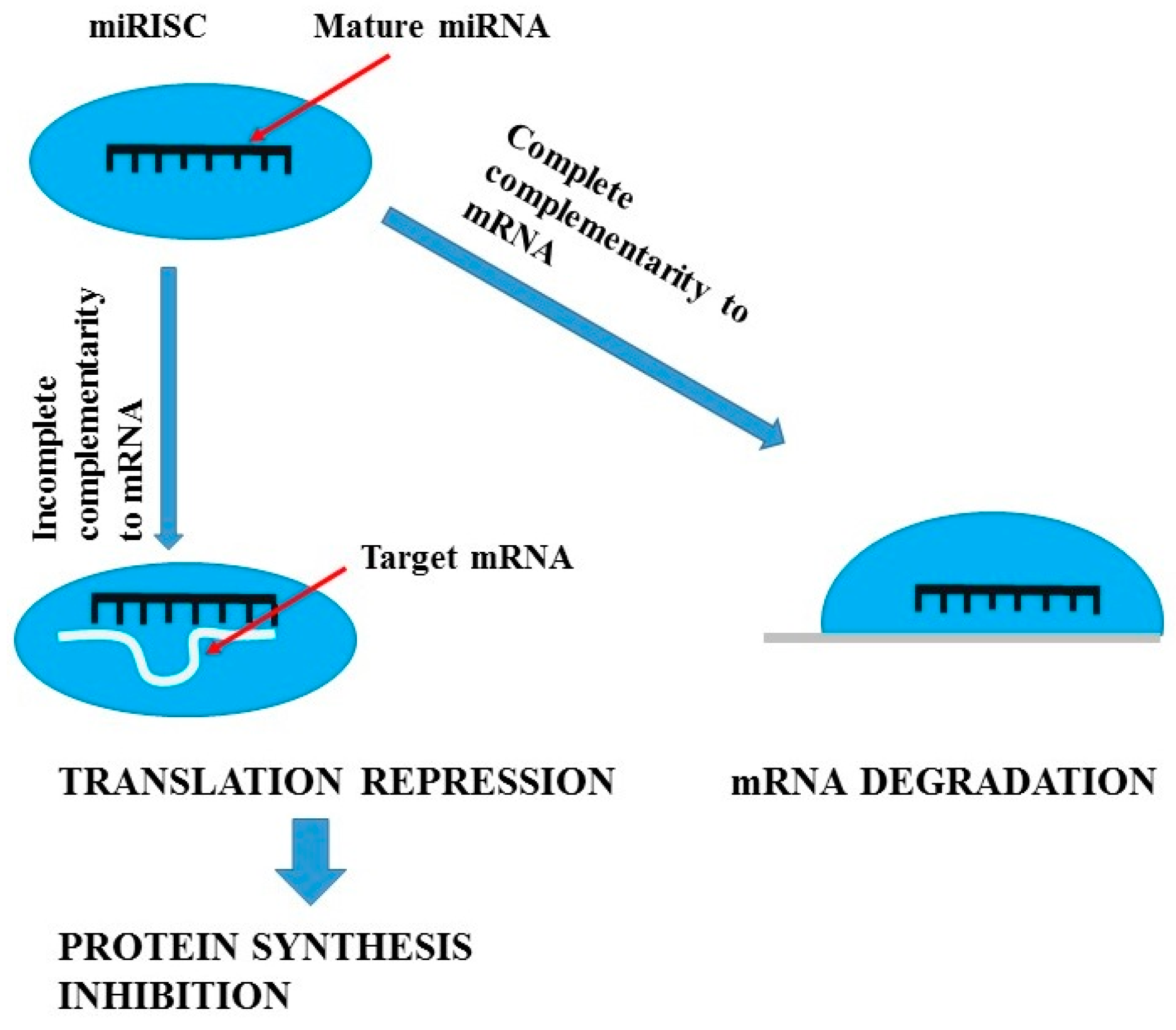{kind=link}
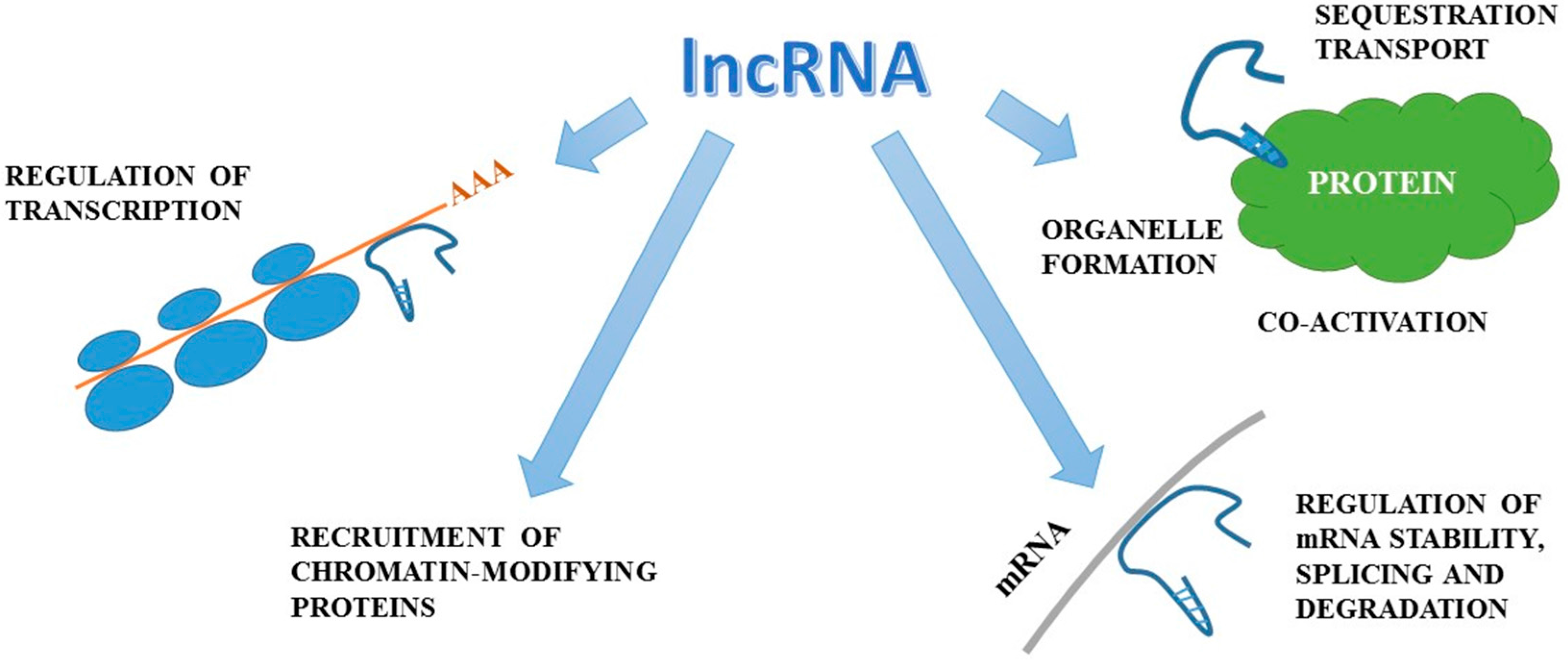{kind=link}
Abstract
:1. Introduction
2. Asymmetric Dimethylarginine (ADMA)
3. Symmetric Dimethylarginine (SDMA)
4. Uromodulin
5. Kidney Injury Molecule-1 (KIM-1)
6. Neutrophil Gelatinase-Associated Lipocalin (NGAL)
7. miRNA, ncRNA, lncRNA and lincRNA Biomarkers
8. Proteomic and Metabolomic Biomarkers
9. Conclusions
Conflicts of Interest
References
- Khan, Z.; Pandey, M. Role of kidney biomarkers of chronic kidney disease: An update. Saudi J. Biol. Sci. 2014, 21, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Viau, A.; Karoui, K.E.; Laouari, D.; Burtin, M.; Nguyen, C.; Mori, K.; Pillebout, E.; Berger, T.; Mak, T.W.; Knebelmann, B.; et al. Lipocalin 2 is essential for chronic kidney disease in mice and human. J. Clin. Investig. 2010, 120, 4065–4076. [Google Scholar] [CrossRef] [PubMed]
- Hostetter, T.H. Progression of renal disease and renal hypertrophy. Annu. Rev. Physiol. 1995, 57, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Kliem, V.; Johnson, R.J.; Alpers, C.E.; Yoshimura, A.; Couser, W.J.; Koch, K.M.; Floege, J. Mechanisms involved in the pathogenesis of tubulointerstitial fibrosis in 5/6-nephrectomized rats. Kidney Int. 1996, 49, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Pillebout, E.; Weitzman, J.B.; Burtin, M.; Martino, C.; Federici, P.; Yaniv, M.; Friedlander, G.; Terzi, F. JunD protects against chronic kidney disease by regulating paracrine mitogens. J. Clin. Investig. 2003, 112, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Quaggin, S.E.; Kreidberg, J.A. Development of the renal glomerulus: Good neighbours and good fences. Development 2008, 135, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Wasung, M.E.; Chawla, L.S.; Madero, M. Biomarkers of renal function, which and when? Clin. Chim. Acta 2015, 438, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Steubl, D.; Block, M.; Herbst, V.; Nockher, W.A.; Schlumberger, W.; Satanovskij, R.; Angermann, S.; Hasenau, A.L.; Stecher, L.; Heemann, U.; et al. Plasma uromodulin correlates with kidney function and identifies early stages in chronic kidney disease patients. Medicine 2016, 95, e3011. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.; Ranches, G.; Schafferer, S.; Lukasser, M.; Rudnicki, M.; Mayer, G.; Hüttenhofer, A. Identification of urinary exosomal noncoding RNAs as novel biomarkers in chronic kidney disease. RNA 2017, 23, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Mayer, G. Capillary rarefaction, hypoxia, VEGF and angiogenesis in chronic renal disease. Nephrol. Dial. Transplant. 2011, 26, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Fink, H.A.; Ishani, A.; Taylor, B.C.; Greer, N.L.; MacDonald, R.; Rossini, D.; Sadiq, S.; Lankireddy, S.; Kane, R.L.; Wilt, T.J. Chronic Kidney Disease Stages 1–3: Screening, Monitoring, and Treatment; Agency for Healthcare Research and Quality: Rockville, MD, USA, 2012. [Google Scholar]
- Yasuda, Y.; Cohen, C.D.; Henger, A.; Kretzler, M. Gene expression profiling analysis in nephrology: Towards molecular definition of renal disease. Clin. Exp. Nephrol. 2006, 10, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Gentile, G.; Remuzzi, G. Novel biomarkers for renal diseases? None for the moment (but one). J. Biomol. Screen. 2016, 21, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Mok, C.C. Biomarkers for lupus nephritis: A critical appraisal. J. Biomed. Biotech. 2010, 2010, 638413. [Google Scholar] [CrossRef] [PubMed]
- Good, D.M.; Zürbig, P.; Argiles, A.; Bauer, H.W.; Behrens, G.; Coon, J.J.; Dakna, M.; Decramer, S.; Delles, C.; Dominiczak, A.F.; et al. Naturally occurring human urinary peptides for use in diagnosis of chronic kidney disease. Mol. Cell. Proteom. 2010, 9, 2424–2437. [Google Scholar] [CrossRef] [PubMed]
- Kolch, W.; Neusüss, C.; Pelzing, M.; Mischak, H. Capillary electrophoresis-mass spectrometry as a powerful tool in clinical diagnosis and biomarker discovery. Mass Spectrom. Rev. 2005, 24, 959–977. [Google Scholar] [CrossRef] [PubMed]
- National Kidney Foundation. Tests to Measure Kidney Function, Damage and Detect Abnormalities. Available online: https://www.kidney.org/atoz/content/kidneytests (accessed on 29 April 2017).
- Calice-Silva, V.; Vieira, M.A.; Raimann, J.G.; Carter, M.; Callegari, J.; Levin, N.W.; Kotanko, P.; Pecoits-Filho, R. Saliva urea nitrogen dipstick—A novel bedside diagnostic tool for acute kidney injury. Clin. Nephrol. 2014, 82, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Raimann, J.G.; Kirisits, W.; Gebetsroither, E.; Carter, M.; Callegari, J.; Rosales, L.; Levin, N.W.; Kotanko, P. Saliva urea dipstick test: Application in chronic kidney disease. Clin. Nephrol. 2011, 76, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.; Calice-Silva, V.; Raimann, J.G.; Hemmila, U.; Craik, A.; Mtekateka, M.; Hamilton, F.; Kawale, Z.; Dobbie, H.; Dreyer, G.; et al. Diagnostic performance of a saliva urea nitrogen dipstick to detect kidney disease in Malawi. Kidney Int. Rep. 2016, 2, 219–227. [Google Scholar] [CrossRef]
- Zeng, Q.; Li, P.; Cai, Y.; Zhou, W.; Wang, H.; Luo, J.; Ding, J.; Chen, H. Detection of creatinine in exhaled breath of humans with chronic kidney disease by extractive electrospray ionization mass spectrometry. J. Breath Res. 2016, 10, 016008. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Snyder, M. Systems biology: Personalized medicine for the future? Curr. Opin. Pharmacol. 2012, 12, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Snyder, M.; Du, J.; Gerstein, M. Personal genome sequencing: Current approaches and challenges. Genes Dev. 2010, 5, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Su, M.W.; Tung, K.Y.; Liang, P.H.; Tsai, C.H.; Kuo, N.W.; Lee, Y.L. Gene–gene and gene–environmental interactions of childhood asthma: A multifactor dimension reduction approach. PLoS ONE 2012, 7, e30694. [Google Scholar] [CrossRef] [PubMed]
- Kinross, J.M.; Darzi, A.W.; Nicholson, J.K. Gut microbiome–host interactions in health and disease. Genome Med. 2011, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Fliser, D.; Kronemberg, F.; Kielstein, J.T.; Morath, C.; Bode-Böger, S.M.; Haller, H.; Ritz, E. Asymmetric dimethylarginine and progression of chronic kidney disease: The mild to moderate kidney disease study. J. Am. Soc. Nephrol. 2005, 16, 2456–2461. [Google Scholar] [CrossRef] [PubMed]
- Ravani, P.; Tripepi, G.; Malberti, F.; Testa, S.; Mallamaci, F.; Zoccali, C. Asymmetrical dimethylarginine predicts progression to dialysis and death in patients with chronic kidney disease: A competing risks modeling approach. J. Am. Soc. Nephrol. 2005, 16, 2449–2455. [Google Scholar] [CrossRef] [PubMed]
- Matsuguma, K.; Ueda, S.; Yamagishi, S.; Matsumoto, Y.; Kaneyuki, U.; Shibata, R.; Fujimura, T.; Matsuoka, H.; Kimoto, M.; Kato, S.; et al. Molecular mechanism for elevation of asymmetric dimethylarginine and its role for hypertension in chronic kidney disease. J. Am. Soc. Nephrol. 2006, 17, 2176–2183. [Google Scholar] [CrossRef] [PubMed]
- Kajimoto, H.; Kai, H.; Aoki, H.; Yasuoka, S.; Anegawa, T.; Aoki, Y.; Ueda, S.; Okuda, S.; Imaizumi, T. Inhibition of eNOS phosphorylation mediates endothelial dysfunction in renal failure: New effect of asymmetric dimethylarginine. Kidney Int. 2012, 81, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, D.; Zheng, J.; Feng, Y.; Zhang, Y.; Liu, W. Actin cytoskeleton dependent pathways for ADMA-induced NF-κB activation and TGF-β high expression in human renal glomerular endothelial cells. Acta Biochim. Biophys. Sin. 2012, 44, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Yamagishi, S.; Kaida, Y.; Okuda, S. Asymmetric dimethylarginine may be a missing link between cardiovascular disease and chronic kidney disease. Nephrology 2007, 12, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Yamagishi, S.; Okuda, S. New pathways to renal damage: Role of ADMA in retarding renal disease progression. J. Nephrol. 2010, 23, 377–386. [Google Scholar] [PubMed]
- Miyazaki, H.; Matsuoka, H.; Cooke, J.P.; Usui, M.; Ueda, S.; Okuda, S.; Imaizumi, T. Endogenous nitric oxide synthase inhibitor: A novel marker of atherosclerosis. Circulation 1999, 99, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Tripepi, G.; Mattace Raso, F.; Sijbrands, E.; Seck, M.S.; Maas, R.; Boger, R.; Witteman, J.; Rapisarda, F.; Malatino, L.; Mallamaci, F.; et al. Inflammation and asymmetric dimethylarginine for predicting death and cardiovascular events in ESRD patients. Clin. J. Am. Soc. Nephrol. 2011, 6, 1714–1721. [Google Scholar] [CrossRef] [PubMed]
- Vallance, P.; Leone, A.; Calver, A.; Collier, J.; Moncada, S. Accumulation of an endogenous inhibitor of nitric oxide synthesis in chronic renal failure. Lancet 1992, 339, 572–575. [Google Scholar] [PubMed]
- Martens-Lobenhoffer, J.; Bode-Boger, S.M. Amino acid N-acetylation: Metabolic elimination of symmetric dimethylarginine as symmetric N(α)-acetyldimethylarginine, determined in human plasma and urine by LC–MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015, 975, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Marescau, B.; Nagels, G.; Possemiers, I.; De Broe, M.E.; Because, I.; Billiouw, J.M.; Lornoy, W.; De Deyn, P.P. Guanidino compounds in serum and urine of nondialyzed patients with chronic renal insufficiency. Metabolism 1997, 46, 1024–1031. [Google Scholar] [CrossRef]
- Kielstein, J.T.; Salpeter, S.R.; Bode-Boeger, S.M.; Cooke, J.P.; Fliser, D. Symmetric dimethylarginine (SDMA) as endogenous marker of renal function—A meta-analysis. Nephrol. Dial. Transplant. 2006, 21, 2445–2451. [Google Scholar] [CrossRef] [PubMed]
- Hokamp, J.A.; Nabity, M.B. Renal biomarkers in domestic species. Vet. Clin. Pathol. 2016, 45, 28–56. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, S.; O’Reilly, D.S.; Reid, D.; Talwar, D. Plasma dimethylarginines during the acute inflammatory response. Eur. J. Clin. Investig. 2011, 4, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Lluch, P.; Mauricio, M.D.; Vila, J.M.; Segarra, G.; Medina, P.; Del Olmo, J.A.; Rodrigo, J.M.; Serra, M.A. Accumulation of symmetric dimethylarginine in hepatorenal syndrome. Exp. Biol. Med. 2006, 23, 70–75. [Google Scholar] [CrossRef]
- Meinitzer, A.; Kielstein, J.T.; Pilz, S.; Drechsler, C.; Ritz, E.; Boehm, B.O.; Winkelmann, B.R.; März, W. Symmetrical and asymmetrical dimethylarginine as predictors for mortality in patients referred for coronary angiography: The Ludwigshafen risk and cardiovascular health study. Clin. Chem. 2011, 57, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Cavalca, V.; Veglia, F.; Squellerio, I.; De Metrio, M.; Rubino, M.; Porro, B.; Moltrasio, M.; Tremoli, E.; Marenzi, G. Circulating levels of dimethylarginines, chronic kidney disease and long-term clinical outcome in non-ST-elevation myocardial infarction. PLoS ONE 2012, 7, e48499. [Google Scholar] [CrossRef] [PubMed]
- Krzyzanowska, K.; Mittermayer, F.; Shnawa, N.; Hofer, M.; Schnabler, J.; Etmüller, Y.; Kapiotis, S.; Wolzt, M.; Schernthaner, G. Asymmetrical dimethylarginine is related to renal function, chronic inflammation and macroangiopathy in patients with type 2 diabetes and albuminuria. Diabet. Med. 2007, 24, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, S.; O’Reilly, D.S.; Talwar, D. Biological variation of asymmetric dimethylarginine and related arginine metabolites and analytical performance goals for their measurement in human plasma. Eur. J. Clin. Investig. 2007, 37, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Reinhard, M.; Erlandsen, E.J.; Randers, E. Biological variation of cystatin C and creatinine. Scand. J. Clin. Lab. Investig. 2009, 69, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Kielstein, J.T.; Veldink, H.; Martens-Lobenhoffer, J.; Haller, H.; Burg, M.; Lorenzen, J.M.; Lichtinghagen, R.; Bode-Böger, S.M.; Kliem, V. SDMA is an early marker of change in GFR after living-related kidney donation. Nephrol. Dial. Transplant. 2011, 26, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Säemann, M.D.; Weichhart, T.; Hörl, W.H.; Zlabinger, G.J. Tamm–Horsfall protein: A multilayered defence molecule against urinary tract infection. Eur. J. Clin. Investig. 2005, 35, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Andriole, V.T. The role of Tamm–Horsfall protein in the pathogenesis of reflux nephropathy and chronic pyelonephritis. Yale J. Biol. Med. 1985, 58, 91–100. [Google Scholar] [PubMed]
- Hess, B. Tamm–Horsfall glycoprotein and calcium nephrolithiasis. Miner Electrolyte Metab. 1994, 30, 393–398. [Google Scholar]
- Serafini-Cessi, F.; Malagolini, N.; Cavallone, D. Tamm–Horsfall glycoprotein: Biology and clinical relevance. Am. J. Kidney Dis. 2003, 42, 658–676. [Google Scholar] [CrossRef]
- Malagolini, N.; Cavallone, D.; Serafini-Cessi, F. Intracellular transport, cell-surface exposure and release of recombinant Tamm–Horsfall glycoprotein. Kidney Int. 1997, 52, 1340–1350. [Google Scholar] [CrossRef] [PubMed]
- El-Achkar, T.M.; Wu, X.R. Uromodulin in kidney injury: An instigator, bystander, or protector? Am. J. Kidney Dis. 2012, 59, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Prajczer, S.; Heidenreich, U.; Pfaller, W.; Kotanko, P.; Lhotta, K.; Jennings, P. Evidence for a role of uromodulin in chronic kidney disease progression. Nephrol. Dial. Transplant. 2010, 25, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- De Silva, P.M.C.S.; Mohammed Abdul, K.S.; Eakanayake, E.M.; Jayasinghe, S.S.; Jayasumana, C.; Asanthi, H.B.; Perera, H.S.; Chaminda, G.G.; Chandana, E.P.; Siribaddana, S.H. Urinary biomarkers KIM-1 and NGAL for detection of chronic kidney disease of uncertain etiology (CKDu) among agricultural communities in Sri Lanka. PLoS Negl. Trop. Dis. 2016, 10, e0004979. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, B.D.; Xu, F.; Sabbisetti, V.; Grgic, I.; Naini, S.M.; Wang, N.; Chen, G.; Xiao, S.; Patel, D.; Henderson, J.M.; et al. Chronic epithelial kidney injury molecule-1 expression causes murine kidney fibrosis. J. Clin. Investig. 2013, 123, 4023–4035. [Google Scholar] [CrossRef] [PubMed]
- Sayanthooran, S.; Magana-Arachchi, D.N.; Gunerathne, L.; Abeysekera, T. Potential diagnostic biomarkers for chronic kidney disease of unknown etiology (CKD) in Sri Lanka: A pilot study. BMC Nephrol. 2017, 18, 31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, Y.; Li, L.; Mann, D.; Imig, J.D.; Emmett, N.; Gibbons, G.; Jin, L.M. Glomerular expression of kidney injury molecule-1 and podocytopenia in diabetic glomerulopathy. Am. J. Nephrol. 2011, 34, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Nickolas, T.L.; O’Rourke, M.J.; Yang, J.; Sise, M.E.; Canetta, P.A.; Barasch, N.; Buchen, C.; Khan, F.; Mori, K.; Giglio, J. Sensitivity and specificity of a single emergency department measurement of urinary neutrophil gelatinase-associated lipocalin for diagnosing acute kidney injury. Ann. Intern. Med. 2008, 148, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Han, W.K.; Bailly, V.; Abichandani, R.; Thadhani, R.; Bonventre, J.V. Kidney injury molecule-1 (KIM-1): A novel biomarker for human renal proximal tubule injury. Kidney Int. 2002, 62, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, T.; Bonventre, J.V.; Bailly, V.; Wei, H.; Hession, C.A.; Cate, R.L.; Sanicola, M. Kidney injury molecule-1 (KIM-1), a putative epithelial cell adhesion molecule containing a novel immunoglobulin domain, is up-regulated in renal cells after injury. J. Biol. Chem. 1998, 273, 4135–4142. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, M.; Neilson, E.G. Mechanisms of tubulointerstitial fibrosis. J. Am. Soc. Nephrol. 2010, 21, 1819–1834. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, T.; Asseldonk, E.J.; Humphreys, B.D.; Gunaratnam, L.; Duffield, J.S.; Bonventre, J.V. Kidney injury molecule-1 is a phosphatidylserine receptor that confers a phagocytic phenotype on epithelial cells. J. Clin. Invest. 2008, 118, 1657–1668. [Google Scholar] [CrossRef] [PubMed]
- Bonventre, J.V.; Yang, L. Kidney injury molecule-1. Curr. Opin. Crit. Care 2010, 16, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Blum, A.; Novak, T.; Levinson, R.; Lai, E.; Barasch, J. An epithelial precursor is regulated by the ureteric bud and by the renal stroma. Dev. Biol. 2002, 246, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Goetz, D.; Li, J.Y.; Wang, W.; Mori, K.; Setlik, D.; Du, T.; Erdjument-Bromage, H.; Tempst, P.; Strong, R.; et al. An iron delivery pathway mediated by a lipocalin. Mol. Cell 2002, 10, 1045–1056. [Google Scholar] [CrossRef]
- Kuncio, G.S.; Neilson, E.G.; Haverty, T. Mechanisms of tubulointerstitial fibrosis. Kidney Int. 1991, 39, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Mitsnefes, M.M.; Kathman, T.S.; Mishra, J.; Kartal, J.; Khoury, P.R.; Nickolas, T.L.; Barasch, J.; Devarajan, P. Serum neutrophil gelatinase-associated lipocalin as a marker of renal function in children with chronic kidney disease. Pediatr. Nephrol. 2007, 22, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Nassirpour, R.; Raj, D.; Townsend, R.; Argyropoulos, C. MicroRNA biomarkers in clinical renal disease: From diabetic nephropathy renal transplantation and beyond. Food Chem. Toxicol. 2016, 98, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.Y.; Ma, Y.; Ding, R.; Fu, B.; Shi, S.; Chen, X.M. miR-335 and miR-34a promote renal senescence by suppressing mitochondrial antioxidative enzymes. J. Am. Soc. Nephrol. 2011, 22, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Chung, A.C.K.; Huang, X.R.; Meng, X.; Lan, H.Y. miR-192 mediates TGF-β/Smad3-driven renal fibrosis. J. Am. Soc. Nephrol. 2010, 21, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Yu, X.; Liu, Y.; Kriegel, A.J.; Heng, Y.; Xu, X.; Liang, M.; Ding, X. miR-29c is downregulated in renal interstitial fibrosis in humans and rats and restored by HIF-α activation. Am. J. Physiol. Ren. Physiol. 2013, 304, F1274–F1282. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Rao, D.S.; Chaudhuri, A.A.; Baltimore, D. Physiological and pathological roles for microRNAs in the immune system. Nat. Rev. Immunol. 2010, 10, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Arce, L.; Natarajan, R. MicroRNAs and their role in progressive kidney diseases. Clin. J. Am. Soc. Nephrol. 2009, 4, 1255–1266. [Google Scholar] [CrossRef] [PubMed]
- Taïbi, F.; Metzinger-Le Meuth, V.; M’Baya-Moutoula, E.; Djelouat, M.; Louvet, L.; Bugnicourt, J.M.; Poirot, S.; Bengrine, A.; Chillon, J.M.; Massy, Z.A.; et al. Possible involvement of microRNAs in vascular damage in experimental chronic kidney disease. Biochim. Biophys. Acta 2014, 1842, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Harvey, S.J.; Jarad, G.; Cunningham, J.; Goldberg, S.; Schermer, B.; Harfe, B.D.; McManus, M.T.; Benzing, T.; Miner, J.H. Podocyte-specific deletion of dicer alters cytoskeletal dynamics and causes glomerular disease. J. Am. Soc. Nephrol. 2008, 19, 2150–2158. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Ng, K.H.; Rosen, S.; Dostal, A.; Gregory, R.I.; Kreidberg, J.A. Podocyte-specific loss of functional MicroRNAs leads to rapid glomerular and tubular injury. J. Am. Soc. Nephrol. 2008, 19, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.Y.S.; Sims-Lucas, S.; Bushnell, D.S.; Bodnar, A.J.; Kreidberg, J.A.; Ho, J. Dicer function is required in the metanephric mesenchyme for early kidney development. Am. J. Physiol. Ren. Physiol. 2014, 306, F764–F772. [Google Scholar] [CrossRef] [PubMed]
- Bartram, M.P.; Dafinger, C.; Habbig, S.; Benzing, T.; Schermer, B.; Müller, R.U. Loss of DGCR8-mediated microRNA expression in the kidney results in hydronephrosis and renal malformation. BMC Nephrol. 2015, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Lal, M.A.; Young, K.W.; Andag, U. Targeting the podocyte to treat glomerular kidney disease. Drug Discov. Today 2015, 20, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- Merscher, S.; Pedigo, C.E.; Mendez, A.J. Metabolism, energetics, and lipid biology in the podocyte e cellular cholesterol-mediated glomerular injury. Front. Endocrinol. 2014, 5, 169. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, S.; Shu, Y.; Yoshida, F.; Inagaki, T.; Sonoda, J.; Watanabe, T.; Nomoto, K.; Nagata, M. Podocyte injury promotes progressive nephropathy in zucker diabetic fatty rats. Lab. Investig. 2002, 82, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.X.; Kiattisunthorn, K.; O’Neill, K.D.; Chen, X.; Moorthi, R.N.; Gattone, V.H., 2nd; Allen, M.R.; Moe, S.M. Decreased microRNA is involved in the vascular remodeling abnormalities in chronic kidney disease (CKD). PLoS ONE 2013, 8, e64558. [Google Scholar]
- Lovren, F.; Pan, Y.; Quan, A.; Singh, K.K.; Shukla, P.C.; Gupta, N.; Steer, B.M.; Ingram, A.J.; Gupta, M.; Al-Omran, M.; et al. MicroRNA-145 targeted therapy reduces atherosclerosis. Circulation 2012, 126, S81–S90. [Google Scholar] [CrossRef] [PubMed]
- McClelland, A.; Hagiwara, S.; Kantharidis, P. Where are we in diabetic nephropathy: MicroRNAs and biomarkers? Curr. Opin. Nephrol. Hypertens. 2014, 23, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Trionfini, P.; Benigni, A.; Remuzzi, G. MicroRNAs in kidney physiology and disease. Nat. Rev. Nephrol. 2015, 11, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, K.; Karolina, D.S.; Sepramaniam, S.; Armugam, A.; Wintour, E.M.; Bertram, J.F.; Jeyaseelan, K. Role of microRNAs in kidney homeostasis and disease. Kidney Int. 2012, 81, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Guay, C.; Regazzi, R. Circulating microRNAs as novel biomarkers for diabetes mellitus. Nat. Rev. Endocrinol. 2013, 9, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Paralkar, V.R.; Weiss, M.J. Long noncoding RNAs in biology and hematopoiesis. Blood 2013, 121, 4842–4846. [Google Scholar] [CrossRef] [PubMed]
- Matas, A.J.; Smith, J.M.; Skeans, M.A.; Thompson, B.; Gustafson, S.K.; Schnitzler, M.A.; Stewart, D.E.; Cherikh, W.S.; Wainright, J.L.; Snyder, J.J.; et al. OPTN/SRTR 2012 annual data report: Kidney. Am. J. Transplant. 2014, 14, 11–44. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dov, I.Z.; Muthukumar, T.; Morozov, P.; Mueller, F.B.; Tuschl, T.; Suthanthiran, M. MicroRNA sequence profiles of human kidney allografts with or without tubulointerstitial fibrosis. Transplantation 2012, 94, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Maluf, D.G.; Dumur, C.I.; Suh, J.L.; Scian, M.J.; King, A.L.; Cathro, H.; Lee, J.K.; Gehrau, R.C.; Brayman, K.L.; Gallon, L.; et al. The urine microRNA profile may help monitor post-transplant renal graft function. Kidney Int. 2014, 85, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Scian, M.J.; Maluf, D.G.; David, K.G.; Archer, K.J.; Suh, J.L.; Wolen, A.R.; Mba, M.U.; Massey, H.D.; King, A.L.; Gehr, T.; et al. MicroRNA profiles in allograft tissues and paired urines associate with chronic allograft dysfunction with IF/TA. Am. J. Transplant. 2011, 11, 2110–2122. [Google Scholar] [CrossRef] [PubMed]
- Glowacki, F.; Savary, G.; Gnemmi, V.; Buob, D.; van der Hauwaert, C.; Lo-Guidice, J.M.; Bouy e, S.; Hazzan, M.; Pottier, N.; Perrais, M.; et al. Increased circulating miR-21 levels are associated with kidney fibrosis. PLoS ONE 2013, 8, e58014. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.L.; Cao, Y.H.; Ni, H.F.; Xu, M.; Liu, D.; Liu, H.; Chen, P.S.; Liu, B.C. MicroRNA-29c in urinary exosome/microvesicle as a biomarker of renal fibrosis. Am. J. Physiol. Ren. Physiol. 2013, 305, F1220–F1227. [Google Scholar] [CrossRef] [PubMed]
- Hessels, D.; Klein Gunnewiek, J.M.T.; van Oort, I.; Karthaus, H.F.M.; van Leenders, G.J.L.; van Balken, B.; Kiemeney, L.A.; Witjes, J.A.; Schalken, J.A. DD3 (PCA3)-based molecular urine analysis for the diagnosis of prostate cancer. Eur. Urol. 2003, 44, 8–16. [Google Scholar] [CrossRef]
- Gibb, E.A.; Brown, C.J.; Lam, W.L. The functional role of long non-coding RNA in human carcinomas. Mol. Cancer 2011, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Tinzl, M.; Marberger, M.; Horvath, S.; Chypre, C. DD3 PCA3 RNA analysis in urine—A new perspective for detecting prostate cancer. Eur. Urol. 2004, 46, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Newbury, L.; Drago, G.; Bowen, T.; Redman, J.E. Electrochemical detection of urinary microRNAs via sulfonamide-bound antisense hybridisation. Sens. Actuators B Chem. 2017, 253, 335–341. [Google Scholar] [CrossRef]
- Kashi, K.; Henderson, L.; Bonetti, A.; Carninci, P. Discovery and functional analysis of lncRNAs: Methodologies to investigate an uncharacterized transcriptome. Biochim. Biophys. Acta 2016, 1859, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Siwy, J.; Zürbig, P.; Argiles, A.; Beige, J.; Haubitz, M.; Jankowski, J.; Julian, B.A.; Linde, P.G.; Marx, D.; Mischak, H.; et al. Noninvasive diagnosis of chronic kidney diseases using urinary proteome analysis. Nephrol. Dial. Transplant. 2016, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schanstra, J.P.; Zurbig, P.; Alkhalaf, A.; Argiles, A.; Bakker, S.J.L.; Beige, J.; Bilo, H.J.G.; Chatzikyrkou, C.; Dakna, M.; Dawson, J.; et al. Diagnosis and prediction of CKD progression by assessment of urinary peptides. J. Am. Soc. Nephrol. 2015, 26, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Malaga-Dieguez, L.; Bouhassira, D.; Gipson, D.; Trachtman, H. Novel therapies for FSGS: Preclinical and clinical studies. Adv. Chronic Kidney Dis. 2015, 22, e1–e6. [Google Scholar] [CrossRef] [PubMed]
- Argiles, A.; Mourad, G.; Mion, C.; Atkins, R.C.; Haiech, J. Two-dimensional gel electrophoresis of urinary proteins in kidney diseases. Contrib. Nephrol. 1990, 83, 1–8. [Google Scholar] [PubMed]
- Smith, A.; L’Imperio, V.; De Sio, G.; Ferrario, F.; Scalia, C.; Dell’Antonio, G.; Pieruzzi, F.; Pontillo, C.; Filip, S.; Markoska, K.; et al. α-1-Antitrypsin detected by MALDI imaging in the study of glomerulonephritis: Its relevance in chronic kidney disease progression. Proteomics 2016, 16, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Soyfoo, M.S.; Roth, J.; Vogl, T.; Pochet, R.; Decaux, G. Phagocyte-specific S100A8/22 protein levels during disease exacerbations and infections in systemic lupus erythematosus. J. Rheumatol. 2009, 36, 2190–2194. [Google Scholar] [CrossRef] [PubMed]
- Pontillo, C.; Jacobs, L.; Staessen, J.A.; Schanstra, J.P.; Rossing, P.; Heerspink, H.J.; Siwy, J.; Mullen, W.; Vlahou, A.; Mischak, H.; et al. A urinary proteome-based classifier for the early detection of decline in glomerular filtration. Nephrol. Dial. Transplant. 2016. [Google Scholar] [CrossRef]
- Nkuipou-Kenfack, E.; Duranton, F.; Gayrard, N.; Argilés, À.; Lundin, U.; Weinberger, K.M.; Dakna, M.; Delles, C.; Mullen, W.; Husi, H.; et al. Assessment of metabolomic and proteomic biomarkers in detection and prognosis of progression of renal function in chronic kidney disease. PLoS ONE 2014, 9, e96955. [Google Scholar] [CrossRef] [PubMed]
- Zürbig, P.; Jerums, G.; Hovind, P.; MacIsaac, R.; Mischak, H.; Nielsen, S.E.; Panagiotopoulos, S.; Persson, F.; Rossing, P. Urinary proteomics for early diagnosis in diabetic nephropathy. Diabetes 2012, 61, 3304–3313. [Google Scholar] [CrossRef] [PubMed]
- Roscioni, S.S.; de Zeeuw, D.; Hellemons, M.E.; Mischak, H.; Zürbig, P.; Bakker, S.J.; Gansevoort, R.T.; Reinhard, H.; Persson, F.; Lajer, M.; et al. A urinary peptide biomarker set predicts worsening of albuminuria in type 2 diabetes mellitus. Diabetologia 2012, 56, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Candiano, G.; Musante, L.; Bruschi, M.; Petretto, A.; Santucci, L.; del Boccio, P.; Pavone, B.; Perfumo, F.; Urbani, A.; Scolari, F.; et al. Repetitive fragmentation products of albumin and α1-antitrypsin in glomerular diseases associated with nephrotic syndrome. J. Am. Soc. Nephrol. 2006, 17, 3139–3148. [Google Scholar] [CrossRef] [PubMed]
- Rossing, K.; Mischak, H.; Rossing, P.; Schanstra, J.P.; Wiseman, A.; Maahs, D.M. The urinary proteome in diabetes and diabetes-associated complications: New ways to assess disease progression and evaluate therapy. Proteom. Clin. 2008, 2, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, L.U.; Schiffer, E.; Zürbig, P.; Good, D.M.; Kellmann, M.; Mouls, L.; Pitt, A.R.; Coon, J.J.; Schmieder, R.E.; Peter, K.H.; et al. Urinary proteomics biomarkers in coronary artery disease. Mol. Cell Proteom. 2008, 7, 290–298. [Google Scholar] [CrossRef] [PubMed]
- von Zur Muhlen, C.; Schiffer, E.; Zuerbig, P.; Kellmann, M.; Brasse, M.; Meert, N.; Vanholder, R.C.; Dominiczak, A.F.; Chen, Y.C.; Mischak, H.; et al. Evaluation of urine proteome pattern analysis for its potential to reflect coronary artery atherosclerosis in symptomatic patients. J. Proteome Res. 2009, 8, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, Y.; Liu, Y.; Shi, S.; Wang, S.; Li, X.; Zhang, H.; Wang, H. Urinary uromodulin excretion predicts progression of chronic kidney disease resulting from IgA nephropathy. PLoS ONE 2013, 8, e71023. [Google Scholar] [CrossRef] [PubMed]
- Neilson, E.G. Mechanisms of disease: Fibroblasts—A new look at an old problem. Nat. Clin. Pract. Nephrol. 2006, 2, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Boor, P.; Ostendorf, T.; Floege, J. Renal fibrosis: Novel insights into mechanisms and therapeutic targets. Nat. Rev. Nephrol. 2010, 6, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Ruster, C.; Wolf, G. Renin–angiotensin–aldosterone system and progression of renal disease. J. Am. Soc. Nephrol. 2006, 17, 2985–2991. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D. Causes of dysregulation of lipid metabolism in chronic renal failure. Semin. Dial. 2009, 22, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Fouque, D.; Holt, S.; Guebre-Egziabher, F.; Nakamura, K.; Vianey-Saban, C.; Hadj-Aïssa, A.; Hoppel, C.L.; Kopple, J.D. Relationship between serum carnitine, acylcarnitines, and renal function in patients with chronic renal disease. J. Ren. Nutr. 2006, 16, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Norris, K.; Vaziri, N.D. Dysregulation of hepatic fatty acid metabolism in chronic kidney disease. Nephrol. Dial. Transplant. 2013, 28, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Drosatos, K.; Schulze, P.C. Cardiac lipotoxicity: Molecular pathways and therapeutic implications. Curr. Heart Fail. Rep. 2013, 10, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Li, L.O.; Klett, E.L.; Coleman, R.A. Acyl-CoA synthesis, lipid metabolism and lipotoxicity. Biochim. Biophys. Acta 2010, 1801, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Duranton, F.; Lundin, U.; Gayrard, N.; Mischak, H.; Aparicio, M.; Mourad, G.; Daurès, J.P.; Weinberger, K.M.; Argilés, A. Plasma and urinary amino acid metabolomic profiling in patients with different levels of kidney function. Clin. J. Am. Soc. Nephrol. 2014, 9, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, K.; Tankiewicz, J.; Mysliwiec, M.; Pawlak, D. Tissue factor/its pathway inhibitor system and kynurenines in chronic kidney disease patients on conservative treatment. Blood Coagul. Fibrinolysis 2009, 20, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolsky, V.N.; Bucci, T.; Heflich, R.H.; Desjardins, J.; Richardson, F.C. Mice deficient for cytosolic thymidine kinase gene develop fatal kidney disease. Mol. Genet. Metab. 2003, 78, 1–10. [Google Scholar] [CrossRef]
- Dobrovolsky, V.N.; Bowyer, J.F.; Pabarcus, M.K.; Heflich, R.H.; Williams, L.D.; Doerge, D.R.; Arvidsson, B.; Bergquist, J.; Casida, J.E. Effect of arylformamidase (kynurenine formamidase) gene inactivation in mice on enzymatic activity, kynurenine pathway metabolites and phenotype. Biochim. Biophys. Acta 2005, 1724, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Korstanje, R.; Deutsch, K.; Bolanos-Palmieri, P.; Hanke, N.; Schroder, P. Loss of kynurenine 3-mono-oxygenase causes proteinuria. J. Am. Soc. Nephrol. 2016, 27, 3271–3277. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Cui, J.; Gong, H.; Xi, C.; Zhang, T.M. Effect of NAD on PARP-mediated insulin sensitivity in oleic acid treated hepatocytes. J. Cell Physiol. 2015, 230, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- Welsh, G.I.; Hale, L.J.; Eremina, V.; Jeansson, M.; Maezawa, Y.; Lennon, R.; Pons, D.A.; Owen, R.J.; Satchell, S.C.; Miles, M.J.; et al. Insulin signaling to the glomerular podocyte is critical for normal kidney function. Cell Metab. 2010, 12, 329–340. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rysz, J.; Gluba-Brzózka, A.; Franczyk, B.; Jabłonowski, Z.; Ciałkowska-Rysz, A. Novel Biomarkers in the Diagnosis of Chronic Kidney Disease and the Prediction of Its Outcome. Int. J. Mol. Sci. 2017, 18, 1702. https://doi.org/10.3390/ijms18081702
Rysz J, Gluba-Brzózka A, Franczyk B, Jabłonowski Z, Ciałkowska-Rysz A. Novel Biomarkers in the Diagnosis of Chronic Kidney Disease and the Prediction of Its Outcome. International Journal of Molecular Sciences. 2017; 18(8):1702. https://doi.org/10.3390/ijms18081702
Chicago/Turabian StyleRysz, Jacek, Anna Gluba-Brzózka, Beata Franczyk, Zbigniew Jabłonowski, and Aleksandra Ciałkowska-Rysz. 2017. "Novel Biomarkers in the Diagnosis of Chronic Kidney Disease and the Prediction of Its Outcome" International Journal of Molecular Sciences 18, no. 8: 1702. https://doi.org/10.3390/ijms18081702