
Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light †


1
Department of Plant Physiology, Faculty of Agrobiology and Food Resources, Slovak University of Agriculture, 94976 Nitra, Slovakia
2
Laboratory of Bioclimatology, Department of Ecology and Environmental Protection, Poznan University of Life Sciences, Piątkowska 94, 60-649 Poznan, Poland
3
Department of Botany and Plant Physiology, Faculty of Agrobiology, Food and Natural Resources Czech University of Life Sciences, 16500 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
†
Presented at the 1st International Electronic Conference on Plant Science, 1–15 December 2020; Available online: https://iecps2020.sciforum.net/.
Biol. Life Sci. Forum 2021, 4(1), 19; https://doi.org/10.3390/IECPS2020-08777
Published: 1 December 2020
(This article belongs to the Proceedings of The 1st International Electronic Conference on Plant Science)
{kind=link}
{kind=link}
Abstract
:Karrikins (KARs) are relatively simple molecules originating from the burning of plant material, which can enhance the germination of many species from fire-prone environments, but also for species whose life cycle is not connected with fire; for example, Arabidopsis (Arabidopsis thaliana L.). KARs not only improve germination but also alter several physiological processes and morphological features of plants. One of the changes in morphology observed on Arabidopsis seedlings is the shortening of hypocotyls when grown under continuous red light. So far, six biologically active compounds belonging to the KARs group were identified. Most of the plants showed the strongest response to the first identified KAR, KAR1, but Arabidopsis exhibited the strongest reaction to KAR2. In our experiment, we focused on seedlings’ hypocotyl length of rapeseed (Brassica napus napus L.), an economically important plant from the same family as Arabidopsis, Brassicaceae. Our results show that the hypocotyl of eight-day-old seedlings of rapeseed grown under continuous red light was significantly shortened by both KARs examined by us, KAR1, as well as by KAR2. Therefore, we can conclude that, similarly to Arabidopsis, rapeseed seedlings possess the light-dependent response of development to KARs. This proves that the role of these compounds is not yet fully understood in plant life, as well as in the germination induction of fire-following species. Moreover, it is very unlikely that the combustion of plant material is the only source of KARs occurring in nature.
1. Introduction
Karrikins (KARs) are molecules present in the smoke produced by burning plant material [1]. Their chemical structure is relatively simple, as they consist of a five-membered butanolide and a six-membered pyran ring and contain only carbon, hydrogen, and oxygen atoms [2]. The first identified member of the KAR family, which was later named KAR1, was identified as an effective germination promotor of lettuce (Lactuca sativa L.) and smoke-responsive Australian species [3]. To date, six biologically active compounds belonging to the group of KARs have been identified [4]. KARs do not improve only the germination rate of species originating from the fire-prone environment, but also for the species whose life cycle is not connected to fire—one example being the model plant, Arabidopsis (Arabidopsis thaliana L.) [5]. Arabidopsis and its mutants enabled the recognition of other effects of KARs on the plant’s physiology and morphology. One of these changes is a light-dependent shortening of seedling hypocotyls when grown under continuous red light conditions [6]. Despite the majority of the KAR-responsive plants showing the strongest reaction to KAR1, which is present in the smoke in the highest amount, Arabidopsis reacts the most readily to KAR2 [5,6].
Rapeseed (Brassica napus napus L.) is the second-most produced oilseed crop worldwide, which is used for the production of vegetable oil for human consumption as well as for the production of biofuels. Rapeseed and Arabidopsis both belong to the same plant family, Brassicaceae [7,8].
Despite the response of Arabidopsis seedlings to KARs being known for years, the response of the seedlings’ hypocotyl length to KARs under continuous red light was not studied in another species. Therefore, we investigated the effect of two KARs, KAR1 and KAR2, on the length of rapeseed seedlings’ hypocotyl under continuous red and white light and constant darkness conditions.
2. Methods
2.1. Plant Material
Seeds of rapeseed (Brassica napus napus L.) variety LG Architect treated by insecticide Lumiposa were obtained from a local farm, Klasy s. r. o. (Kuralany, Slovakia).
KAR1 and KAR2 were purchased from OlChemIm s. r. o. (Olomouc, Czech Republic). Stock solutions with the amount of KAR 1 mM were prepared by dissolving in acetone and stored at −20 °C until further use. Experimental1 μM solutions were prepared from the stock solutions by dissolving in distilled water on the day of the experiment’s initiation. The control solution was prepared by dissolving the equivalent amount of acetone in distilled water.
Rapeseed seeds were placed in a transparent plastic box with proportions 100 × 90 × 80 mm (length × width × height) designated for in vitro cultivation on four filter papers wetted by 15 mL of either control or experimental (KAR1 or KAR2) solution. Six boxes for each treatment (control, KAR1, and KAR2) with 20 seeds each were prepared. Boxes were placed for eight days in a growth chamber with a temperature of 23 °C and relative air humidity of 60%. Two boxes of each treatment were placed in light-proof boxes (darkness), two to white light, and two to red light. Spectra of the light conditions were measured by SpectraPen mini (Photon Systems Instruments s. r. o., Drasov, Czech Republic) spectroradiometer. The white light of growth chamber LED tubes was of photon flux density ~100 μmol m−2 s−1. The red light was obtained by installing a light filter, effectively filtering out all wavelengths under 570 nm, with the main peak of the transmitted spectrum at ~610 nm and a secondary peak at ~710 nm. The photon flux density of red light was ~20 μmol m−2 s−1.
2.2. Hypocotyl Length Measurement
Fifteen seedlings from each box were stuck to transparent tape after eight days, covered by another piece of transparent tape, and scanned by an office scanner on a black background. The hypocotyl length of 30 seedlings from each treatment and each light condition was measured from pictures using Fiji software (LOCI, University of Wisconsin).
2.3. Statistical Analysis
Analysis of variance and Tukey’s HSD test were performed to establish statistically significant differences between treatments for each light condition. The level of statistical significance was established to p < 0.05. Statistical analysis was performed by the use of RStudio version 1.1.456 (RStudio Inc., Boston, MA, USA).
3. Results and Discussion
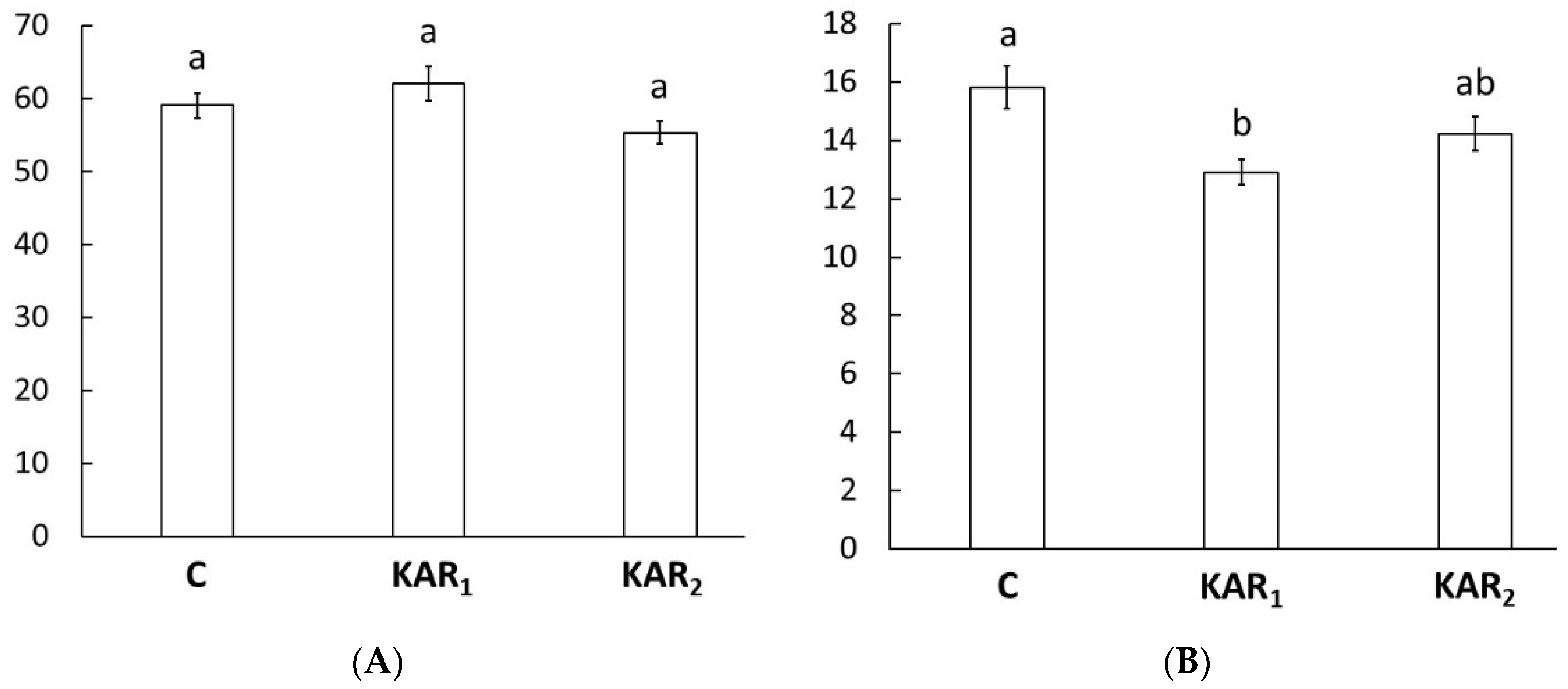
The growth of hypocotyl to length is a basic skotomorphogenic response that enables young seedlings to push through the soil to get exposure to light [9]. The response of seedling growth to KARs is independent of KARs’ effect on seed germination [6]. The results of our experiment show that neither KAR1 nor KAR2 significantly altered the hypocotyl length of rapeseed seedlings (Figure 1A). These results are the opposite of results obtained earlier in an experiment with tomatoes (Lycopersicon esculentum Mill.), okra (Abelmoschus esculentus L.), and beans (Phaseolus vulgaris L.), in which longer hypocotyls of crops grown in darkness were found after the use of KAR1 [10]. Our results are in concordance with the results obtained with Arabidopsis seedlings grown in darkness, which revealed no change of hypocotyl length after the use of KAR1, KAR2, KAR3, or KAR4. However, the same experiment reports hypocotyl elongation inhibition in the case of wild turnip (Brassica tournefortii Gouan.) [6]. The exact reason for such different responses of seedling growth in darkness to KARs remains elusive and needs more examination.
White light is composed of lights of all wavelengths, including blue light, which is a strong hypocotyl elongation inhibitor [11]. The hypocotyl of rapeseed seedlings grown under white light was more than two times shorter than hypocotyl of seedlings grown in red light and almost four times shorter than those grown in darkness (Figure 1 and Figure 2). Regardless of the inhibitory effect of white light, KAR1 further significantly hindered the growth of hypocotyls (Figure 1B). However, the effect of KAR2 on hypocotyl length was insignificant.
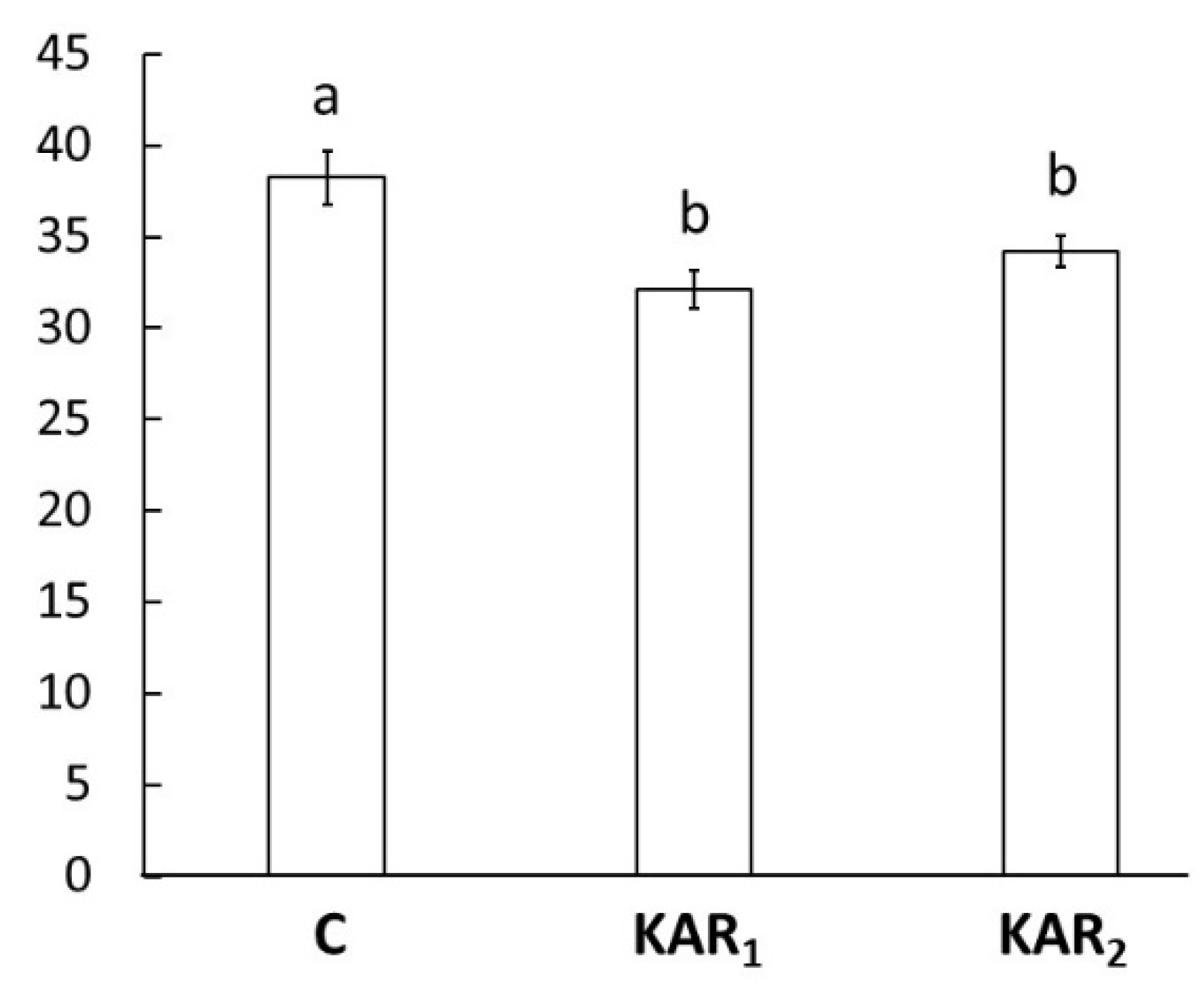
It has been shown that seedlings of Arabidopsis grown under continuous red light in the presence of KARs in the medium have shorter hypocotyl than control seedlings. This response was observed for different ecotypes, and it is independent of germination stimulation reaction [6]. Our results manifest shorter hypocotyls of seedlings grown under continuous red light in the presence of KAR1 or KAR2, which is in accordance with results obtained with the model plant Arabidopsis (Figure 2).
The upregulation of different transcription factors involved in the transduction of signals connected to light conditions was found in Arabidopsis [6]. As we did not perform transcriptional analysis of rapeseed seedlings, we can hypothesize that the mechanism of the response of rapeseed is like Arabidopsis due to their taxonomical relatedness and similar morphology. The exact mechanism needs to be examined by future experiments.
4. Conclusions
Neither of the KARs we examined significantly affected the hypocotyl length of rapeseed seedlings grown in constant darkness. Seedlings grown under continuous white light in the presence of KAR1 had significantly shorter hypocotyls, while KAR2 did not alter the length of the hypocotyl. When grown under continuous red light KAR1 and KAR2 caused the hypocotyls to be significantly shorter compared to the control. Therefore, we can conclude that, similarly to Arabidopsis, rapeseed seedlings possess a light-dependent development response to KARs. This proves that the role of these compounds is not yet fully understood in plant life, as well as the germination induction of fire-following species. Thus, it is very unlikely that the combustion of plant material is the only source of KARs occurring in nature. There remains a need for more studies about KARs as plant growth and development regulators as well as searching for KAR-like compounds produced by plants or possibly microorganisms.
Author Contributions
M.A. and M.B. designed the experiment and conceptualized the work. M.A. performed the experiment, analyzed the data, and wrote the draft of the paper. M.B. supervised the work, reviewed, and revised the draft. Both authors have read and agreed to the final version of the paper.
Funding
This work was financially supported by research project APVV-18-0465: Use of Advanced Phenomic Approaches to Exploit Variation in Photosynthetic Efficiency to Increase Yield under Fluctuating and Stress Environments.
Acknowledgments
This work was financially supported by research project APVV-18-0465: Use of Advanced Phenomic Approaches to Exploit Variation in Photosynthetic Efficiency to Increase Yield under Fluctuating and Stress Environments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nelson, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Regulation of Seed Germination and Seedling Growth by Chemical Signals from Burning Vegetation. Ann. Rev. Plant Biol. 2012, 63, 107–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flematti, G.; Dixon, K.; Smith, S.M. What are karrikins and how were they ‘discovered’ by plants? BMC Biol. 2015, 13, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. A Compound from Smoke That Promotes Seed Germination. Science 2004, 305, 977. [Google Scholar] [CrossRef] [PubMed]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. Identification of Alkyl Substituted 2H-Furo[2,3-c]pyran-2-ones as Germination Stimulants Present in Smoke. J. Agric. Food Chem. 2009, 57, 9475–9480. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C.; Risenborough, J.-A.; Flematti, G.R.; Stevens, J.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins Discovered in Smoke Trigger Arabidopsis Seed Germination by a Mechanism Requiring Gibberellic Acid Synthesis and Light. Plant Physiol. 2009, 149, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.C.; Flematti, G.R.; Riseborough, J.A.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins enhance light responses during germination and seedling development in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 7095–7100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.N.; Khan, Z.; Luo, T.; Liu, J.; Rizwan, M.; Zhang, J.; Xu, Z.; Wu, H.; Hu, L. Seed priming with gibberellic acid and melatonin in rapeseed: Consequences for improving yield and seed quality under drought and non-stress conditions. Ind. Crops Prod. 2020, 156, 112850. [Google Scholar] [CrossRef]
- FAO. Oilcrops, Oils and Meals; FAO: Rome, Italy, 2018; Available online: http://www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Oilcrops/Documents/Food_outlook_oilseeds/FO_Oilcrops.pdf (accessed on 25 October 2020).
- Wang, L.; Xu, Q.; Yu, H.; Ma, H.; Li, X.; Yang, J.; Chu, J.; Xie, Q.; Wang, Y.; Smith, S.M.; et al. Strigolactone and Karrikin Signaling Pathways Elicit Ubiquitination and Proteolysis of SMXL2 to Regulate Hypocotyl Elongation in Arabidopsis thaliana. Plant Cell 2020, 32, 2251–2270. [Google Scholar] [CrossRef] [PubMed]
- Van Staden, J.; Sparg, S.G.; Kulkarni, M.G.; Light, M.E. Post-germination effects of the smoke-derived compound3-methyl-2H-furo[2,3-c]pyran-2-one, and its potential as a preconditioning agent. Field Crops Res. 2006, 98, 98–105. [Google Scholar] [CrossRef]
- Folta, K.M.; Spalding, E.P. Unexpected roles for cryptochrome 2 and phototropin revealed by high-resolution analysis of blue light-mediated hypocotyl growth inhibition. Plant J. 2001, 26, 471–478. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Hypocotyl length [mm] of eight-day-old rapeseed (Brassica napus napus L.) seedlings treated by 1 μM karrikin1 (KAR1), 1 μM karrikin2 (KAR2), or control solution (C) grown under (A) constant darkness; (B) continuous white light. Bars represent the mean of 30 seedlings ± standard error. Different letters indicate statistically significant differences at the level p < 0.05.
Figure 1.
Hypocotyl length [mm] of eight-day-old rapeseed (Brassica napus napus L.) seedlings treated by 1 μM karrikin1 (KAR1), 1 μM karrikin2 (KAR2), or control solution (C) grown under (A) constant darkness; (B) continuous white light. Bars represent the mean of 30 seedlings ± standard error. Different letters indicate statistically significant differences at the level p < 0.05.

Figure 2.
Hypocotyl length [mm] of eight-day-old rapeseed (Brassica napus napus L.) seedlings treated by 1 μM karrikin1 (KAR1), 1 μM karrikin2 (KAR2), or control solution (C) grown under continuous red light. Bars represent the mean of 30 seedlings ± standard error. Different letters indicate statistically significant differences at the level p < 0.05.
Figure 2.
Hypocotyl length [mm] of eight-day-old rapeseed (Brassica napus napus L.) seedlings treated by 1 μM karrikin1 (KAR1), 1 μM karrikin2 (KAR2), or control solution (C) grown under continuous red light. Bars represent the mean of 30 seedlings ± standard error. Different letters indicate statistically significant differences at the level p < 0.05.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Antala, M.; Brestic, M. Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light. Biol. Life Sci. Forum 2021, 4, 19. https://doi.org/10.3390/IECPS2020-08777
AMA Style
Antala M, Brestic M. Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light. Biology and Life Sciences Forum. 2021; 4(1):19. https://doi.org/10.3390/IECPS2020-08777
Chicago/Turabian StyleAntala, Michal, and Marian Brestic. 2021. "Karrikins Reduce the Hypocotyl Length of Rapeseed (Brassica napus napus L.) under Continuous Red Light" Biology and Life Sciences Forum 4, no. 1: 19. https://doi.org/10.3390/IECPS2020-08777