
Exploring the Influence of Biological Traits and Environmental Drivers on Water Use Variations across Contrasting Forests




Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Sites
2.2. Sap Flow Measurements and Scaling to Stand Water Use
2.3. Data Analysis
3. Results
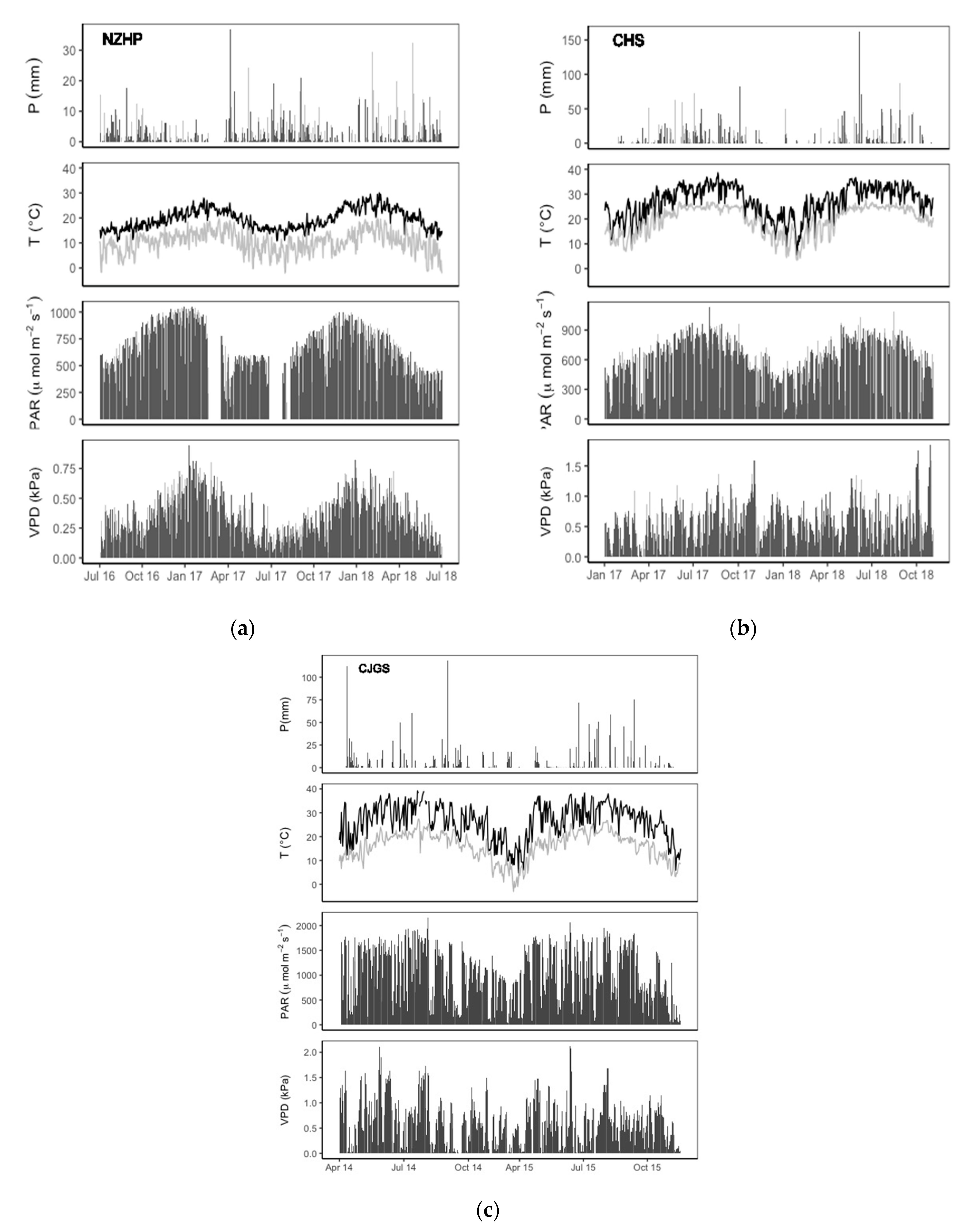
3.1. Environmental Conditions across Three Climatic Zones
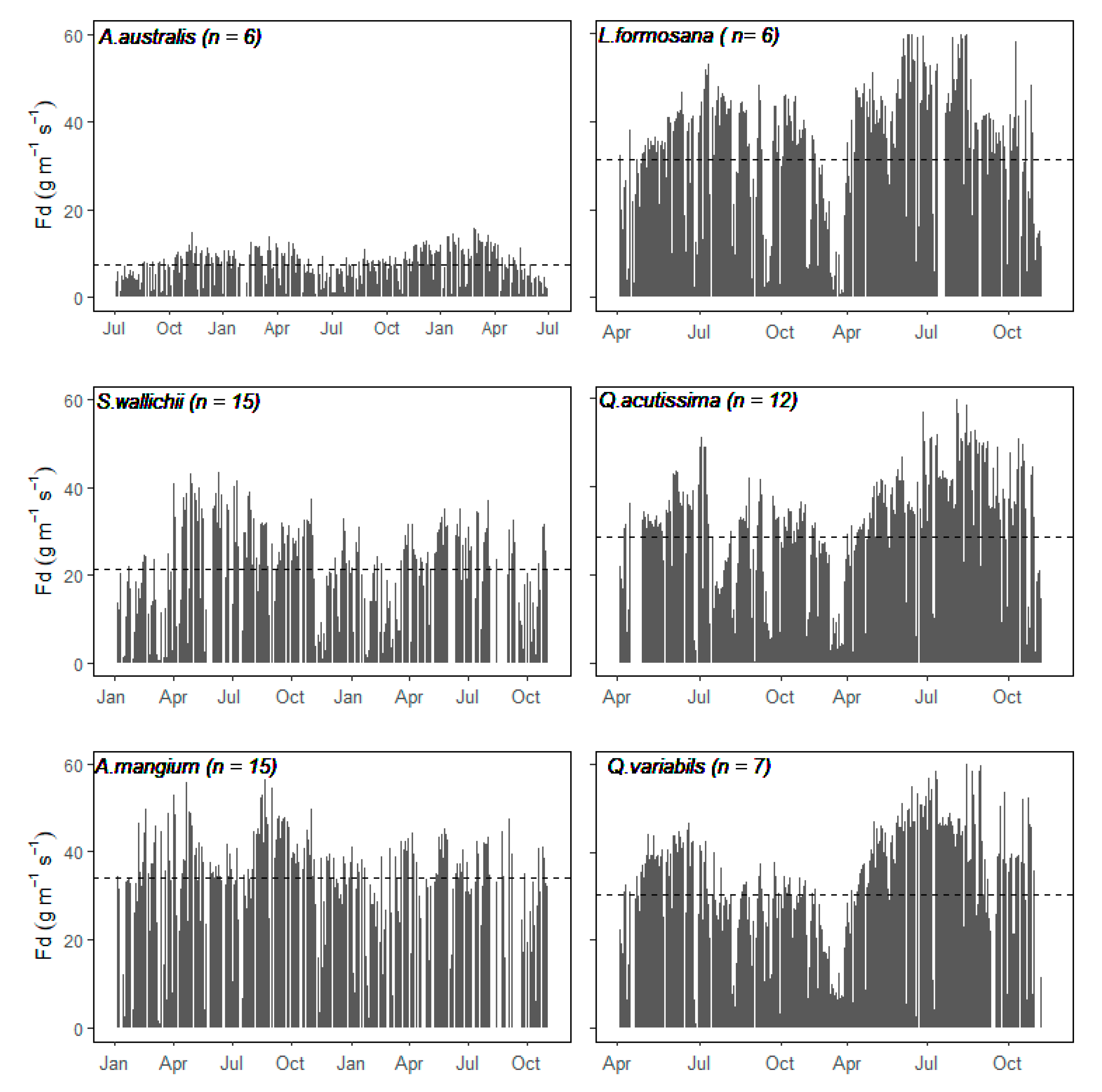
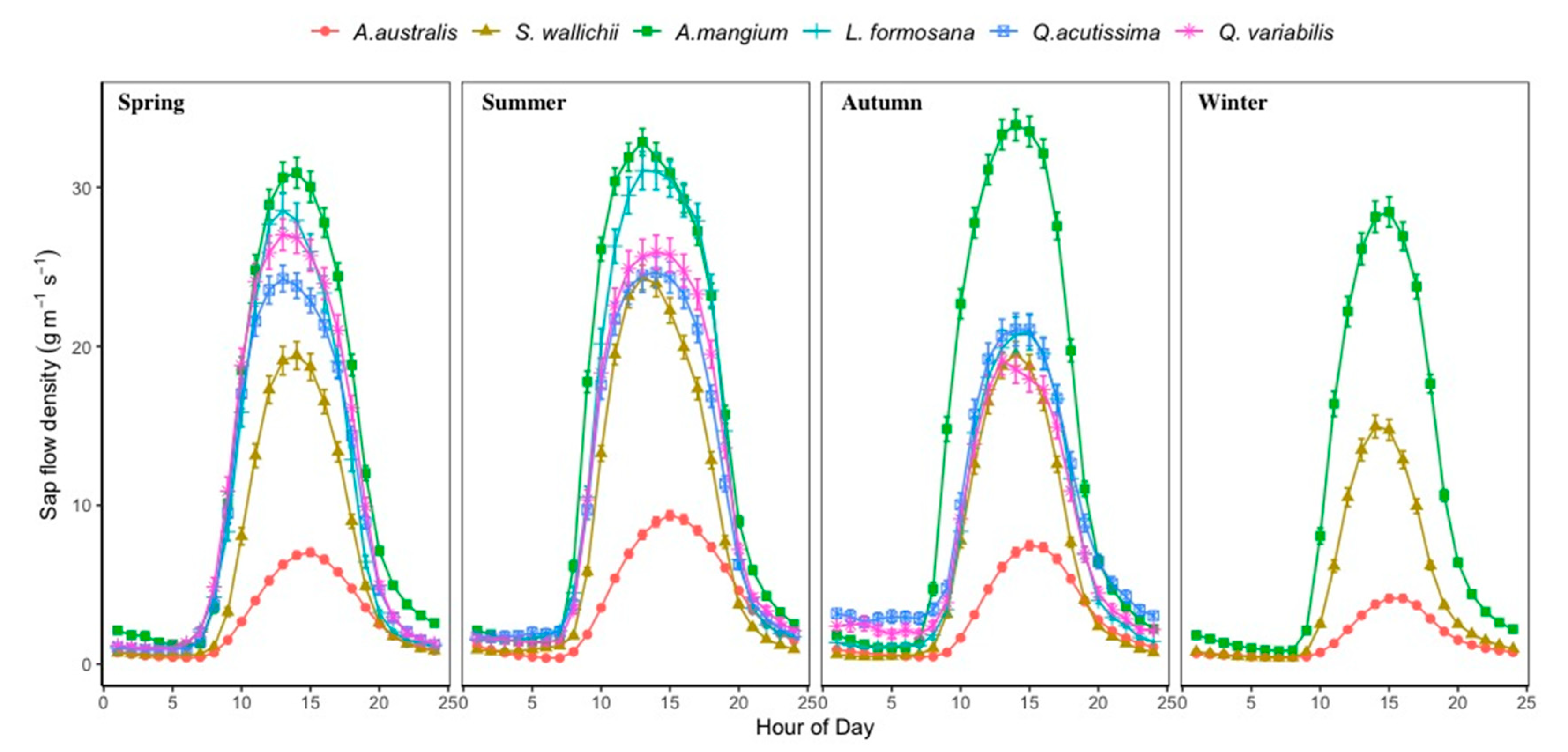
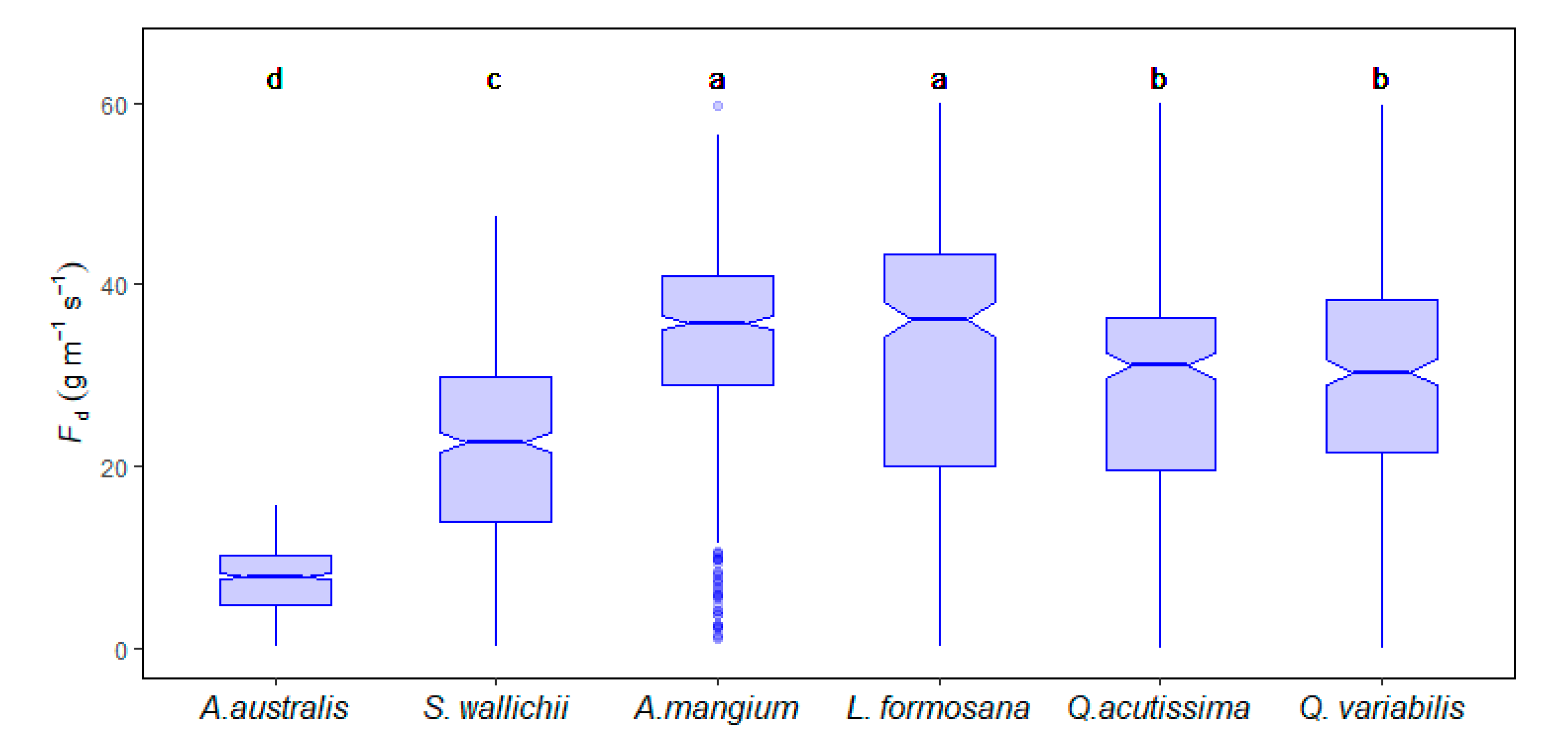
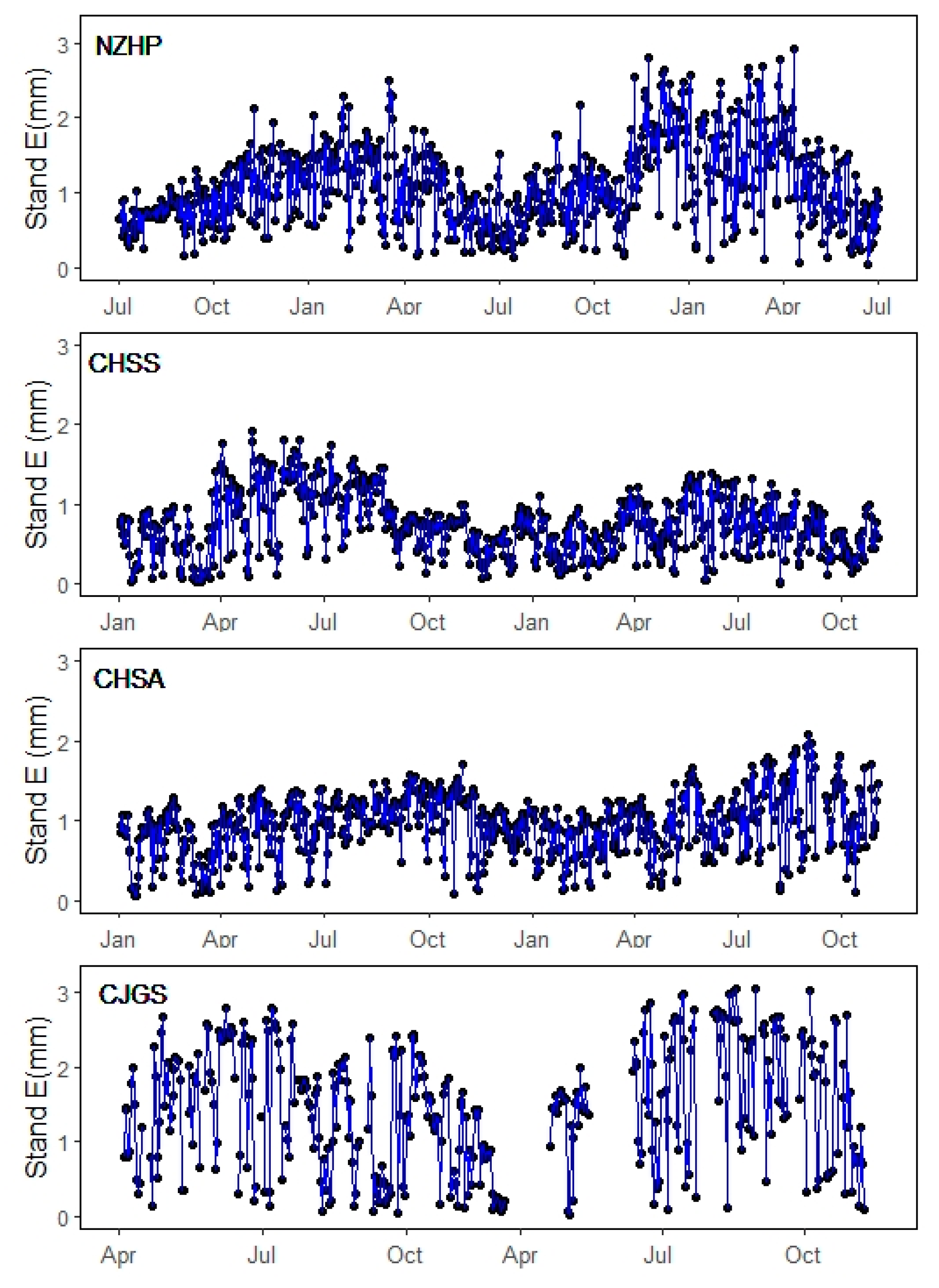
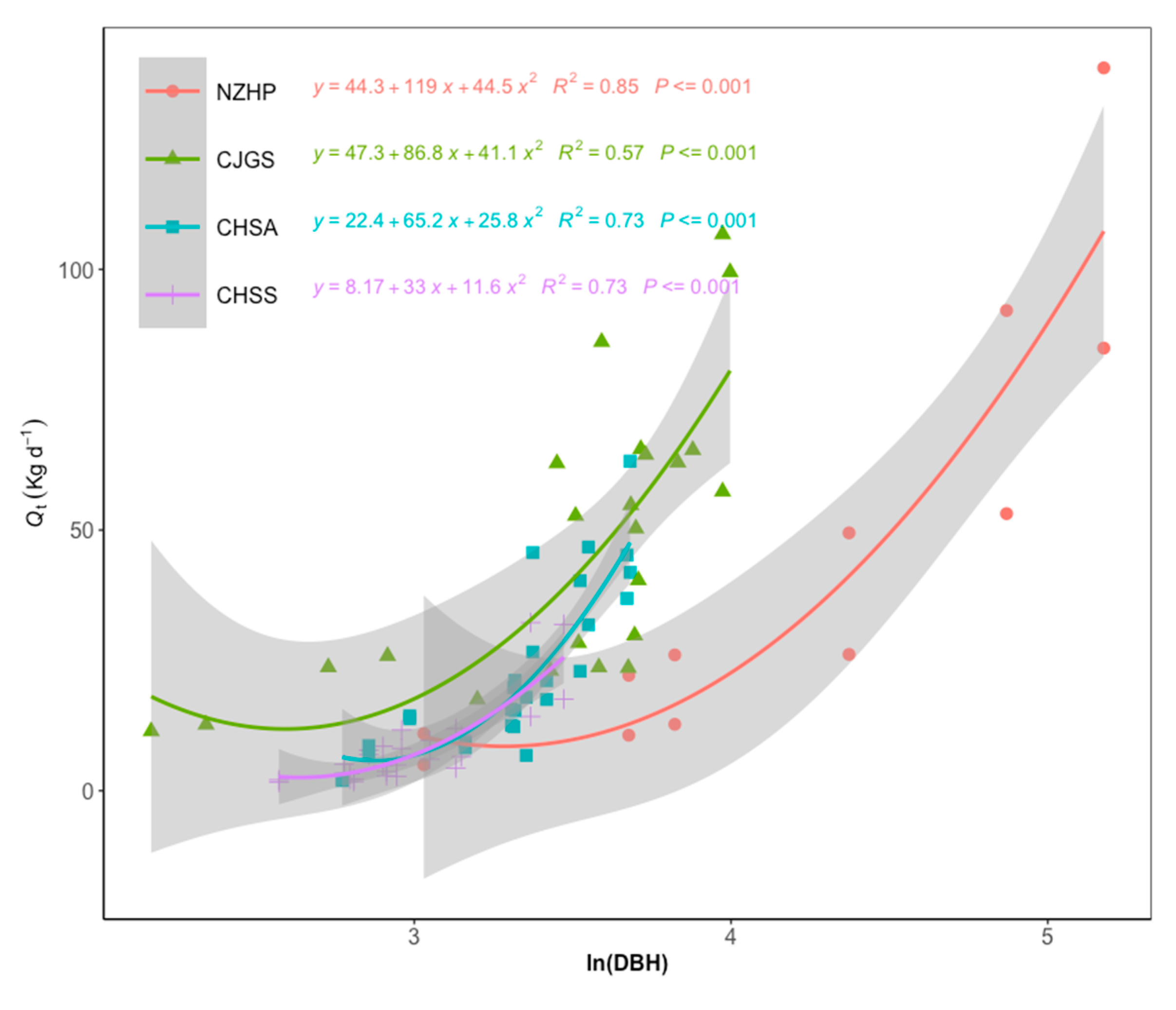
3.2. Sap Flow Density and Stand Scale Transpiration
3.3. Environmental Variables for Stand Scale and Key Drivers Affecting Tree Sap Flow
3.4. Response of Individual Daily Sap Flux to VPD and PAR for Different Species
4. Discussion
4.1. Climate—Depend Water Use Variation across Sites
4.2. Response of Tree Transpiration to Environmental Drivers
4.3. Transpiration Differences among Tree Species
4.4. Sensitivity and Vulnerability to Environmental Drivers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghimire, C.P.; Bruijnzeel, L.A.; Lubczynski, M.W.; Zwartendijk, B.W.; Odongo, V.O.; Ravelona, M.; Van Meerveld, H.J. Transpiration and stomatal conductance in a young secondary tropical montane forest: Contrasts between native trees and invasive understorey shrubs. Tree Physiol. 2018, 38, 1053–1070. [Google Scholar] [CrossRef]
- Tor-ngern, P.; Oren, R.; Oishi, A.C.; Uebelherr, J.M.; Palmroth, S.; Tarvainen, L.; Ottosson-Löfvenius, M.; Linder, S.; Domec, J.C.; Näsholm, T. Ecophysiological variation of transpiration of pine forests: Synthesis of new and published results. Ecol. Appl. 2017, 27, 118–133. [Google Scholar] [CrossRef]
- Manoli, G.; Domec, J.C.; Novick, K.; Oishi, A.C.; Noormets, A.; Marani, M.; Katul, G. Soil–plant–atmosphere conditions regulating convective cloud formation above southeastern US pine plantations. Glob. Chang. Biol. 2016, 22, 2238–2254. [Google Scholar] [CrossRef]
- Peters, E.B.; McFadden, J.P.; Montgomery, R.A. Biological and environmental controls on tree transpiration in a suburban landscape. J. Geophys. Res. 2010, 115, G04006. [Google Scholar] [CrossRef]
- Hassler, S.K.; Weiler, M.; Blume, T. Tree-, stand-and site-specific controls on landscape-scale patterns of transpiration. Hydrol. Earth Syst. Sci. 2018, 22, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Whitley, R.; Taylor, D.; Macinnis-Ng, C.; Zeppel, M.; Yunusa, I.; O’Grady, A.; Froend, R.; Medlyn, B.; Eamus, D. Developing an empirical model of canopy water flux describing the common response of transpiration to solar radiation and VPD across five contrasting woodlands and forests. Hydrol. Process. 2013, 27, 1133–1146. [Google Scholar] [CrossRef]
- Macinnis-Ng, C.; Wyse, S.; Veale, A.; Schwendenmann, L.; Clearwater, M. Sap flow of the southern conifer, Agathis australis during wet and dry summers. Trees-Struct. Funct. 2016, 30, 19–33. [Google Scholar] [CrossRef]
- Wang, H.; Wang, X.; Zhao, P.; Zheng, H.; Ren, Y.; Gao, F.; Ouyang, Z. Transpiration rates of urban trees, Aesculus Chinensis. J. Environ. Sci. 2012, 24, 1278–1287. [Google Scholar] [CrossRef]
- Hu, Y.T.; Zhao, P.; Zhu, L.W.; Zhao, X.H.; Ni, G.Y.; Ouyang, L.; Schäfer, K.V.R.; Shen, W.J. Responses of sap flux and intrinsic water use efficiency to canopy and understory nitrogen addition in a temperate broadleaved deciduous forest. Sci. Total Environ. 2019, 648, 325–336. [Google Scholar] [CrossRef]
- Sperry, J.S.; Venturas, M.D.; Todd, H.N.; Trugman, A.T.; Anderegg, W.R.; Wang, Y.; Tai, X. The impact of rising CO2 and acclimation on the response of US forests to global warming. PNAS 2019, 116, 25734–25744. [Google Scholar] [CrossRef]
- Chemura, A.; Rwasoka, D.; Mutanga, O.; Dube, T.; Mushore, T. The impact of land-use/land cover changes on water balance of the heterogeneous Buzi sub-catchment, Zimbabwe. RSASE 2020, 18, 100292. [Google Scholar] [CrossRef]
- Kume, T.; Otsuki, K.; Du, S.; Yamanaka, N.; Wang, Y.L.; Liu, G.B. Spatial variation in sap flow velocity in semiarid region trees: Its impact on stand-scale transpiration estimates. Hydrol. Process. 2012, 26, 1161–1168. [Google Scholar] [CrossRef]
- Catovsky, S.; Holbrook, N.M.; Bazzaz, F.A. Coupling whole-tree transpiration and canopy photosynthesis in coniferous and broad-leaved tree species. Can. J. Forest Res. 2002, 32, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Lee, M.; Reum Han, A.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Siddiq, Z.; Chen, Y.J.; Zhang, Y.J.; Zhang, J.L.; Cao, K.F. More sensitive response of crown conductance to VPD and larger water consumption in tropical evergreen than in deciduous broadleaf timber trees. Agric. Forest Meteorol. 2017, 247, 399–407. [Google Scholar] [CrossRef]
- Kaplick, J.; Clearwater, M.J.; Macinnis-Ng, C. Comparative water relations of co-occurring trees in a mixed podocarp-broadleaf forest. J. Plant Ecol. 2019, 12, 163–175. [Google Scholar] [CrossRef]
- Poyatos, R.; Granda, V.; Molowny-Horas, R.; Mencuccini, M.; Steppe, K.; Martínez-Vilalta, J. SAPFLUXNET: Towards a global database of sap flow measurements. Tree Physiol. 2016, 36, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Qaderi, M.M.; Martel, A.B.; Dixon, S.L. Environmental factors influence plant vascular system and water regulation. Plants 2019, 8, 65. [Google Scholar] [CrossRef] [Green Version]
- Meinzer, F.C.; Woodruff, D.R.; Eissenstat, D.M.; Lin, H.S.; Adams, T.S.; McCulloh, K.A. Above-and belowground controls on water use by trees of different wood types in an eastern US deciduous forest. Tree Physiol. 2013, 33, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Zhao, P.; Shen, W.; Rao, X.; Hu, Y. Physiological homeostasis and morphological plasticity of two tree species subjected to precipitation seasonal distribution changes. Perspect. Plant Ecol. 2017, 25, 1–19. [Google Scholar] [CrossRef]
- Tombesi, S.; Almehdi, A.; Dejong, T.M. Phenotyping vigour control capacity of new peach rootstocks by xylem vessel analysis. Sci. Hortic. 2011, 127, 353–357. [Google Scholar] [CrossRef]
- Hietz, P.; Rosner, S.; Hietz-Seifert, U.; Wright, S.J. Wood traits related to size and life history of trees in a Panamanian rainforest. New Phytol. 2017, 213, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database– enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [Green Version]
- Oliva, C.L.; Bucci, S.J.; Di, F.D.; Lezcano, O.A.; Campanello, P.I.; Scholz, F.G.; Rodríguez, S.; Madanes, N.; Cristiano, P.M.; Hao, G.Y.; et al. Water storage dynamics in the main stem of subtropical tree species differing in wood density, growth rate and life history traits. Tree Physiol. 2015, 35, 354–365. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Bhusal, S.J.; Yoon, T.M. Comparisons of physiological and anatomical characteristics between two cultivars in bileader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- von Allmen, E.I.; Sperry, J.S.; Bush, S.E. Contrasting whole-tree water use, hydraulics, and growth in a co-dominant diffuse-porous vs. ring-porous species pair. Trees-Struct. Funct. 2015, 29, 717–728. [Google Scholar] [CrossRef]
- Matheny, A.M.; Fiorella, R.P.; Bohrer, G.; Poulsen, C.J.; Morin, T.H.; Wunderlich, A.; Vogel, C.S.; Curtis, P.S. Contrasting strategies of hydraulic control in two codominant temperate tree species. Ecohydrology 2017, 10, e1815. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Garcia-Forner, N. Water potential regulation, stomatal behaviour and hydraulic transport under drought: Deconstructing the iso/anisohydric concept. Plant Cell Environ. 2017, 40, 962–976. [Google Scholar] [CrossRef] [Green Version]
- Buckley, T.N. How do stomata respond to water status? New Phytol. 2019, 224, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/anisohydry: A plant–environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef]
- Klein, T. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Funct. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef]
- Roman, D.T.; Novick, K.A.; Brzostek, E.R.; Dragoni, D.; Rahman, F.; Phillips, R.P. The role of isohydric and anisohydric species in determining ecosystem-scale response to severe drought. Oecologia 2015, 179, 641–654. [Google Scholar] [CrossRef]
- Feng, X.; Ackerly, D.D.; Dawson, T.E.; Manzoni, S.; McLaughlin, B.; Skelton, R.P.; Vico, G.; Weitz, A.P.; Thompson, S.E. Beyond isohydricity: The role of environmental variability in determining plant drought responses. Plant Cell Environ. 2019, 42, 1104–1111. [Google Scholar] [CrossRef] [Green Version]
- Macinnis-Ng, C.; Schwendenmann, L.; Clearwater, M. Radial variation of sap flow of kauri (Agathis australis) during wet and dry summers. Acta Hortic. 2013, 991, 205–214. [Google Scholar] [CrossRef]
- Chappel, P.R. The climate and weather of Auckland. NIWA Sci. Technol. Ser. 2013, 60, 40. [Google Scholar]
- Thomas, M.B.; Spurway, M.I. Nutrition of container-grown rewa-rewa (Knightia excelsa). In The International Plant Propagators’ Society Combined Proceedings; Lincoln University: Canterbury, New Zealand, 1987; Volume 36, pp. 450–455. [Google Scholar]
- Wang, Q.; Lintunen, A.; Zhao, P.; Shen, W.; Salmon, Y.; Chen, X.; Ouyang, L.; Zhu, L.; Ni, G.; Sun, D.; et al. Assessing Environmental Control of Sap Flux of Three Tree Species Plantations in Degraded Hilly Lands in South China. Forests 2020, 11, 206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Shen, W.; Zhu, S.; Wan, S.; Luo, Y.; Yan, J.; Wang, K.; Liu, L.; Dai, H.; Li, P.; et al. CAN Canopy Addition of Nitrogen Better Illustrate the Effect of Atmospheric Nitrogen Deposition on Forest Ecosystem? Sci Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Hu, Y.; Zhao, X.; Zhao, P.; Ouyang, L.; Ni, G.; Liu, N. Specific responses of sap flux and leaf functional traits to simulated canopy and understory nitrogen additions in a deciduous broadleaf forest. Funct. Plant Biol. 2019, 46, 986–993. [Google Scholar] [CrossRef]
- Zhu, L.; Hu, Y.; Zhao, X.; Zeng, X.; Zhao, P.; Zhang, Z.; Ju, Y. The impact of drought on sap flow of cooccurring Liquidambar formosana Hance and Quercus variabilis Blume in a temperate forest, Central China. Ecohydrology 2017, 10, e1828. [Google Scholar] [CrossRef]
- Zhu, L.W.; Zhao, P.; Cai, X.A.; Zeng, X.P.; Ni, G.Y.; Zhang, J.Y.; Zou, L.L.; Mei, T.T.; Yu, M.H. Effects of sap velocity on the daytime increase of stem CO 2 efflux from stems of Schima superba trees. Trees-Struct. Funct. 2012, 26, 535–542. [Google Scholar] [CrossRef]
- Kaplick, J. Water and Carbon Relations in New Zealand Kauri Forest. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2018. [Google Scholar]
- Burkhardt, T. Thirsty at Night: Patterns and Functionality of Nocturnal Transpiration in Kauri (Agathis australis). Master’s Thesis, The University of Auckland, Auckland, New Zealand, 2019. [Google Scholar]
- Karikala, C. Sapwood Area and Density of Temperate Kauri Forest Species. Honours Thesis, The University of Auckland, Auckland, New Zealand, 2015. [Google Scholar]
- Ouyang, L.; Zhao, P.; Zhu, L.; Zhang, Z.; Zhao, X.; Ni, G. Difference in response of water use to evaporative demand for codominant diffuse-porous versus ring-porous tree species under N addition in a temperate forest. Ecohydrology 2017, 10, e1829. [Google Scholar] [CrossRef]
- Pataki, D.E.; McCarthy, H.R.; Litvak, E.; Pincetl, S. Transpiration of urban forests in the Los Angeles metropolitan area. Ecol. Appl. 2011, 21, 661–677. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yu, Z. Environmental Control on Transpiration: A Case Study of a Desert Ecosystem in Northwest China. Water 2020, 12, 1211. [Google Scholar] [CrossRef]
- Wu, J.; Chavana-Bryant, C.; Prohaska, N.; Serbin, S.P.; Guan, K.; Albert, L.P.; Yang, X.; van Leeuwen, W.J.; Garnello, A.J.; Martins, G.; et al. Convergence in relationships between leaf traits, spectra and age across diverse canopy environments and two contrasting tropical forests. New Phytol. 2017, 214, 1033–1048. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.H.; Zhang, Y.P.; Deng, X.B.; Song, Q.H.; Liu, W.J.; Deng, Y.; Tang, J.W.; Liao, Z.Y.; Zhao, J.F.; Song, L.; et al. Interannual and seasonal variability of water use efficiency in a tropical rainforest: Results from a 9 year eddy flux time series. J. Geophys. Res.-Atmos. 2015, 120, 464–479. [Google Scholar] [CrossRef]
- Jonard, F.; André, F.; Ponette, Q.; Vincke, C.; Jonard, M. Sap flux density and stomatal conductance of European beech and common oak trees in pure and mixed stands during the summer drought of 2003. J. Hydrol. 2011, 409, 371–381. [Google Scholar] [CrossRef]
- Clausnitzer, F.; Köstner, B.; Schwärzel, K.; Bernhofer, C. Relationships between canopy transpiration, atmospheric conditions and soil water availability-Analyses of long-term sap-flow measurements in an old Norway spruce forest at the Ore Mountains/Germany. Agric. Forest Meteorol. 2011, 151, 1023–1034. [Google Scholar] [CrossRef]
- Zeppel, M.J.; Macinnis-Ng, C.M.; Yunusa, I.A.; Whitley, R.J.; Eamus, D. Long term trends of stand transpiration in a remnant forest during wet and dry years. J. Hydrol. 2008, 349, 200–213. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Xiao, K.; Zhao, Z.; Xiang, W.; Xu, C.; Lei, P.; Deng, X.; Li, J. Stand Transpiration Estimates from Recalibrated Parameters for the Granier Equation in a Chinese Fir (Cunninghamia lanceolata) Plantation in Southern China. Forests 2018, 9, 162. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Dickinson, R.E. A review of global terrestrial evapotranspiration: Observation, modeling, climatology, and climatic variability. Rev. Geophys. 2012, 50, RG2005. [Google Scholar] [CrossRef]
- Hu, Y.; Zhao, P.; Shen, W.; Zhu, L.; Ni, G.; Zhao, X.; Zhang, Z.; Rao, X.; Ouyang, L.; Zeng, X.; et al. Responses of tree transpiration and growth to seasonal rainfall redistribution in a subtropical evergreen broad-leaved forest. Ecosystems 2018, 21, 811–826. [Google Scholar] [CrossRef]
- Bourdier, T.; Cordonnier, T.; Kunstler, G.; Piedallu, C.; Lagarrigues, G.; Courbaud, B. Tree size inequality reduces forest productivity: An analysis combining inventory data for ten European species and a light competition model. PLoS ONE 2016, 11, e0151852. [Google Scholar] [CrossRef]
- Johnson, D.M.; McCulloh, K.A.; Woodruff, D.R.; Meinzer, F.C. Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different? Plant Sci. 2012, 195, 48–53. [Google Scholar] [CrossRef]
- Wyse, S.V.; Macinnis-Ng, C.M.; Burns, B.R.; Clearwater, M.J.; Schwendenmann, L. Species assemblage patterns around a dominant emergent tree are associated with drought resistance. Tree Physiol. 2013, 33, 1269–1283. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Zhao, P.; Shen, W.; Niu, J.; Zhu, L.; Ni, G. Biophysical limits to responses of water flux to vapor pressure deficit in seven tree species with contrasting land use regimes. Agric. Forest Meteorol. 2015, 200, 258–269. [Google Scholar] [CrossRef]
- Germon, A.; Guerrini, I.A.; Bordron, B.; Bouillet, J.P.; Nouvellon, Y.; de Moraes Gonçalves, J.L.; Jourdan, C.; Paula, R.R.; Laclau, J.P. Consequences of mixing Acacia mangium and Eucalyptus grandis trees on soil exploration by fine-roots down to a depth of 17 m. Plant Soil. 2018, 424, 203–220. [Google Scholar] [CrossRef] [Green Version]
- Siddiq, Z.; Cao, K.F. Increased water use in dry season in eight dipterocarp species in a common plantation in the northern boundary of Asian tropics. Ecohydrology 2016, 9, 871–881. [Google Scholar] [CrossRef]
- Anderegg, W.R.; Klein, T.; Bartlett, M.; Sack, L.; Pellegrini, A.F.; Choat, B.; Jansen, S. Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. PNAS 2016, 113, 5024–5029. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.; Konings, A.G.; Trugman, A.T.; Yu, K.; Bowling, D.R.; Gabbitas, R.; Karp, D.S.; Pacala, S.; Sperry, J.S.; Sulman, B.N.; et al. Hydraulic diversity of forests regulates ecosystem resilience during drought. Nature 2018, 561, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Kröber, W.; Zhang, S.; Ehmig, M.; Bruelheide, H. Linking xylem hydraulic conductivity and vulnerability to the Leaf Economics Spectrum—A cross-species study of 39 evergreen and deciduous broadleaved subtropical tree species. PLoS ONE 2014, 9, e109211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Õunapuu, E.; Sellin, A. Daily dynamics of leaf and soil-to-branch hydraulic conductance in silver birch (Betula pendula) measured in situ. Plant Physiol. Bioch. 2013, 68, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Gil-Pelegrín, E.; Peguero-Pina, J.J.; Sancho-Knapik, D. (Eds.) Oaks Physiological Ecology: Exploring the Functional Diversity of Genus Quercus L.; Springer: Berlin/Heidelberg, Germany, 2017; Volume 7. [Google Scholar]
- Köcher, P.; Horna, V.; Leuschner, C. Stem water storage in five coexisting temperate broad-leaved tree species: Significance, temporal dynamics and dependence on tree functional traits. Tree Physiol. 2013, 33, 817–832. [Google Scholar] [CrossRef]
- Richards, A.E.; Wright, I.J.; Lenz, T.I.; Zanne, A.E. Sapwood capacitance is greater in evergreen sclerophyll species growing in high compared to low-rainfall environments. Funct. Ecol. 2014, 28, 734–744. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | New Zealand NZHP | South China | Central China CJGS | |
|---|---|---|---|---|
| CHSS | CHSA | |||
| Location | 36°47.7′ S 174°29.5′ E | 22°41′ N, 112°54′ E | 31°46′–31°52′ N 114°01′–114°06′ E | |
| Study period | July 2016–June 2018 24 months | January 2017–November 2018 23 months | April–November 2014, April–October 2015 15 months | |
| Annual mean temperature (°C) | 14.2 | 21.9 | 15.2 | |
| Annual rainfall (mm) | 1231 | 1580 | 1119 | |
| Plot size (m2) | 2000 | 900 | 1600 | 1600 |
| Stand DBH (cm) | 26.2 ± 2.1 | 20.4 ± 0.8 | 23.9 ± 1.1 | 12.6 ± 0.2 |
| Stand height(m) | 19.8 ± 1.6 | 12.5 ± 0.4 | 14.8 ± 0.5 | 12.1 ± 0.2 |
| Tree density (N ha−1) | 770 | 867 | 519 | 446 |
| Stand basal area (m2 ha−1) | 94 | 31 | 26.2 | 55.6 |
| Biological Traits | A. australis | S. wallichii | A. mangium | L. formosana | Q. acutissima | Q. variabilis |
|---|---|---|---|---|---|---|
| Wood type WD (g/cm3) | c 0.45 | d 0.60 | d 0.46 | d 0.59 | r 0.91 | r 0.82 |
| N (%) | 0.73 | 1.72 | 2.53 | 1.89 | 2.34 | 2.25 |
| C (%) | 51.15 | 47.87 | 49.15 | 42.15 | 45.57 | 45.71 |
| gs (mmol/m2/s1) | 130–158 | 130 | 350 | 120(s) | × | 100–400 |
| δ13C (‰) | −26.28 | −31.40 | −33.60 | −30.0 | −30.7 | −30.0 |
| A (µmol/m2/s) | 8.20 | 7.00 | 9.67 | 12.63(s) | 9.41 | 2.56–14.54(s) |
| P50 (MPa) | −2.32 | −2.8 | × | −3.5 | −3.39 | × |
| Ψ-min (MPa) | −(1.2–1.5) | −(1.2–1.7) | −(1.6) | −(1.7–2.3) | −(1.7–1.9) | × |
| SLA (cm2/g−1) | 27.3 | × | × | 238.2 | 166.9 | 143.1 |
| LAI | × | 3.9 | 2.8 | × | × | × |
| Sites | Species | Sample Trees | Height (m) | DBH (cm) | As e Equation | Reference |
|---|---|---|---|---|---|---|
| NZHP | A. australis | 6 | 23.5 ± 0.8 | 83.6 ± 24.4 | 0.2971 × DBH1.4226 | [7,44] |
| CHS | S. wallichii | 15 | 12.85 ± 0.43 | 20.3 ± 1.3 | 0.2077 × DBH2.2168 | [37] |
| A. mangium | 15 | 14.61 ± 0.63 | 28.48 ± 1.78 | 0.4936 × DBH1.761 | ||
| CJGS | L. formosana | 6 | 17.4 ± 2.1 | 18.8 ± 3.8 | 0.0068 × exp 0.0677DBH | [39,45] |
| Q. acutissima | 11 | 25.7 ± 1.0 | 40.3 ± 2.6 | 0.0071 × exp 0.0512DBH | ||
| Q. variabilis | 7 | 24.0 ± 1.2 | 40.7 ± 2.6 | 0.0083 × exp 0.0495DBH |
| Category | NZHP | CHSS | CHSA | CJGS | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| Eigenvalue | 1.76 | 1.25 | 0.96 | 1.70 | 1.37 | 0.96 | 1.64 | 1.34 | 0.99 | 1.78 | 1.08 | 0.92 |
| Variance (%) | 44.44 | 22.24 | 23.30 | 41.27 | 26.81 | 13.11 | 38.50 | 25.58 | 14.01 | 52.76 | 19.55 | 13.90 |
| Cum.var (%) | 80.00 | 81.19 | 78.10 | 86.22 | ||||||||
| Variables Loadings | ||||||||||||
| Tem | 0.351 | 0.500 | 0.285 | 0.564 | −0.020 | −0.005 | 0.573 | 0.111 | −0.004 | 0.451 | −0.392 | 0.204 |
| PAR | 0.435 | −0.183 | −0.097 | 0.471 | −0.240 | −0.036 | 0.520 | −0.110 | −0.106 | 0.501 | −0.134 | 0.103 |
| VPD | 0.512 | 0.188 | 0.022 | 0.370 | −0.497 | −0.170 | 0.472 | −0.383 | −0.131 | 0.532 | −0.027 | 0.145 |
| Rain | −0.314 | 0.117 | 0.748 | 0.043 | 0.493 | −0.538 | −0.038 | 0.487 | −0.250 | −0.274 | −0.255 | 0.876 |
| RH | −0.396 | 0.476 | −0.002 | 0.199 | 0.587 | 0.348 | 0.072 | 0.644 | 0.292 | −0.429 | −0.339 | 0.004 |
| VMC | −0.399 | −0.218 | −0.306 | 0.341 | 0.259 | −0.530 | 0.099 | 0.307 | −0.806 | x | x | x |
| wind | 0.084 | −0.063 | 0.506 | −0.407 | −0.201 | −0.527 | −0.403 | −0.287 | −0.418 | 0.052 | 0.805 | 0.399 |
| Sites | Variables | Estimate | p Value | Partial R2 |
|---|---|---|---|---|
| NZHP | (Intercept) | −1.071 | <0.001 | 0.607 |
| PAR | 0.0406 | <0.001 | 0.386 | |
| VPD | 11.43 | <0.001 | 0.488 | |
| T RH | −0.8699 0.0797 | <0.001 <0.001 | 0.0896 0.1038 | |
| VMC | 0.5095 | <0.001 | 0.1036 | |
| P | −0.1355 | <0.001 | 0.205 | |
| Wind | −5.842 | <0.001 | 0.0004 | |
| CHSS | (Intercept) | −1.5678 | <0.05 | 0.6601 |
| PAR | 0.0073 | <0.001 | 0.4645 | |
| VPD | 5.991 | <0.001 | 0.3969 | |
| T | 0.0817 | <0.05 | 0.2372 | |
| VMC | 0.0484 | <0.05 | 0.0504 | |
| CHSA | (Intercept) | 0.8410 | 0.7400 | 0.4562 |
| PAR | 0.0039 | <0.01 | 0.2377 | |
| VPD | 4.9375 | <0.001 | 0.3023 | |
| T RH | 0.4816 −0.0108 | <0.001 <0.001 | 0.1564 0.0577 | |
| VMC | 0.2483 | <0.001 | 0.0439 | |
| CJGS | (Intercept) | 0.4163 | <0.01 | 0.4977 |
| PAR | 0.0017 | <0.001 | 0.3895 | |
| VPD | 0.4959 | <0.001 | 0.3644 | |
| T | 0.0279 | <0.001 | 0.2474 | |
| P | −0.0089 | <0.05 | 0.0952 | |
| Wind | −0.2431 | <0.001 | 0.0035 |
| Species | Fd vs. VPD | Fd vs. PAR | |||||
|---|---|---|---|---|---|---|---|
| n | a | b | p | a | b | p | |
| A. australis | 6 | 18.73 | 0.67 | <0.001 | 1.33 | 0.0016 | <0.001 |
| S. wallichii | 15 | 38.15 | 0.71 | <0.001 | 5.61 | 0.0015 | <0.001 |
| A. mangium | 15 | 41.23 | 1.89 | <0.001 | 4.26 | 0.0035 | <0.001 |
| L. formosana | 6 | 44.25 | 1.09 | <0.001 | 5.08 | 0.0011 | <0.001 |
| Q. acutissima | 12 | 35.94 | 1.68 | <0.001 | 3.81 | 0.0018 | <0.001 |
| Q. variabilis | 7 | 35.11 | 2.26 | <0.001 | 4.01 | 0.0016 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Zhao, P.; Zhu, L.; Wang, Q.; Hu, Y.; Cranston, B.M.; Kaplick, J.; Lei, O.; Chen, X.; Ni, G.; et al. Exploring the Influence of Biological Traits and Environmental Drivers on Water Use Variations across Contrasting Forests. Forests 2021, 12, 161. https://doi.org/10.3390/f12020161
Zhao X, Zhao P, Zhu L, Wang Q, Hu Y, Cranston BM, Kaplick J, Lei O, Chen X, Ni G, et al. Exploring the Influence of Biological Traits and Environmental Drivers on Water Use Variations across Contrasting Forests. Forests. 2021; 12(2):161. https://doi.org/10.3390/f12020161
Chicago/Turabian StyleZhao, Xiuhua, Ping Zhao, Liuwei Zhu, Qian Wang, Yanting Hu, Benjamin M. Cranston, Julia Kaplick, Ouyang Lei, Xia Chen, Guangyan Ni, and et al. 2021. "Exploring the Influence of Biological Traits and Environmental Drivers on Water Use Variations across Contrasting Forests" Forests 12, no. 2: 161. https://doi.org/10.3390/f12020161