Salicylate UV-Filters in Sunscreen Formulations Compromise the Preservative System Efficacy against Pseudomonas aeruginosa and Burkholderia cepacia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms Strains and Growth Conditions
2.2. Preservative System and Formulations
2.3. Antimicrobial and Antifungal Activity of the Preservative
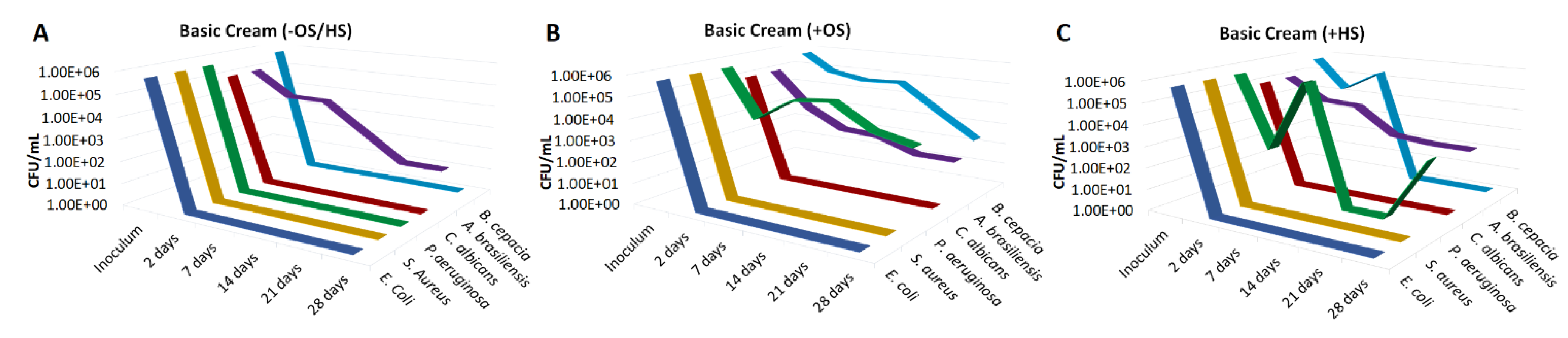
2.4. Challenge Test
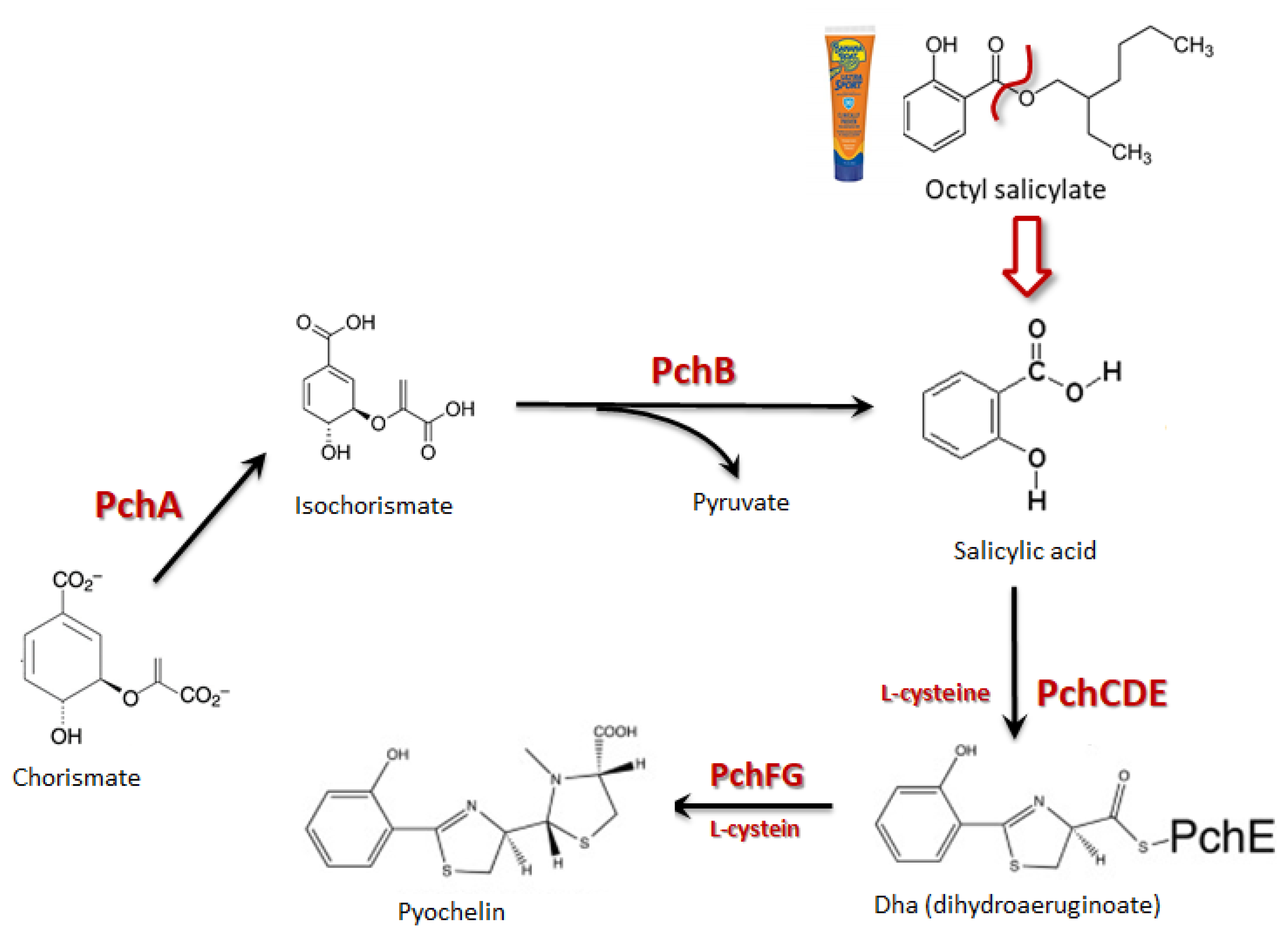
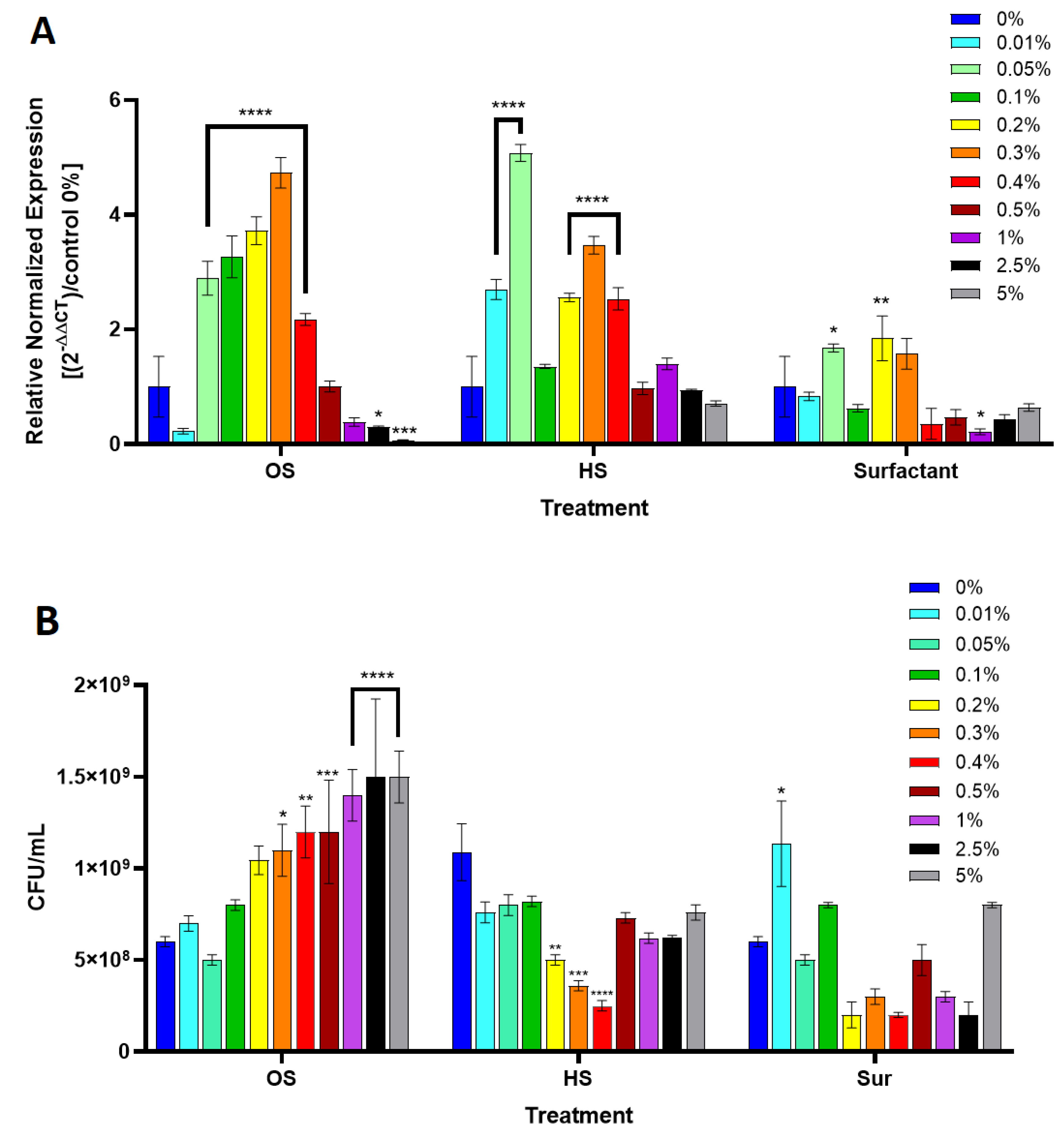
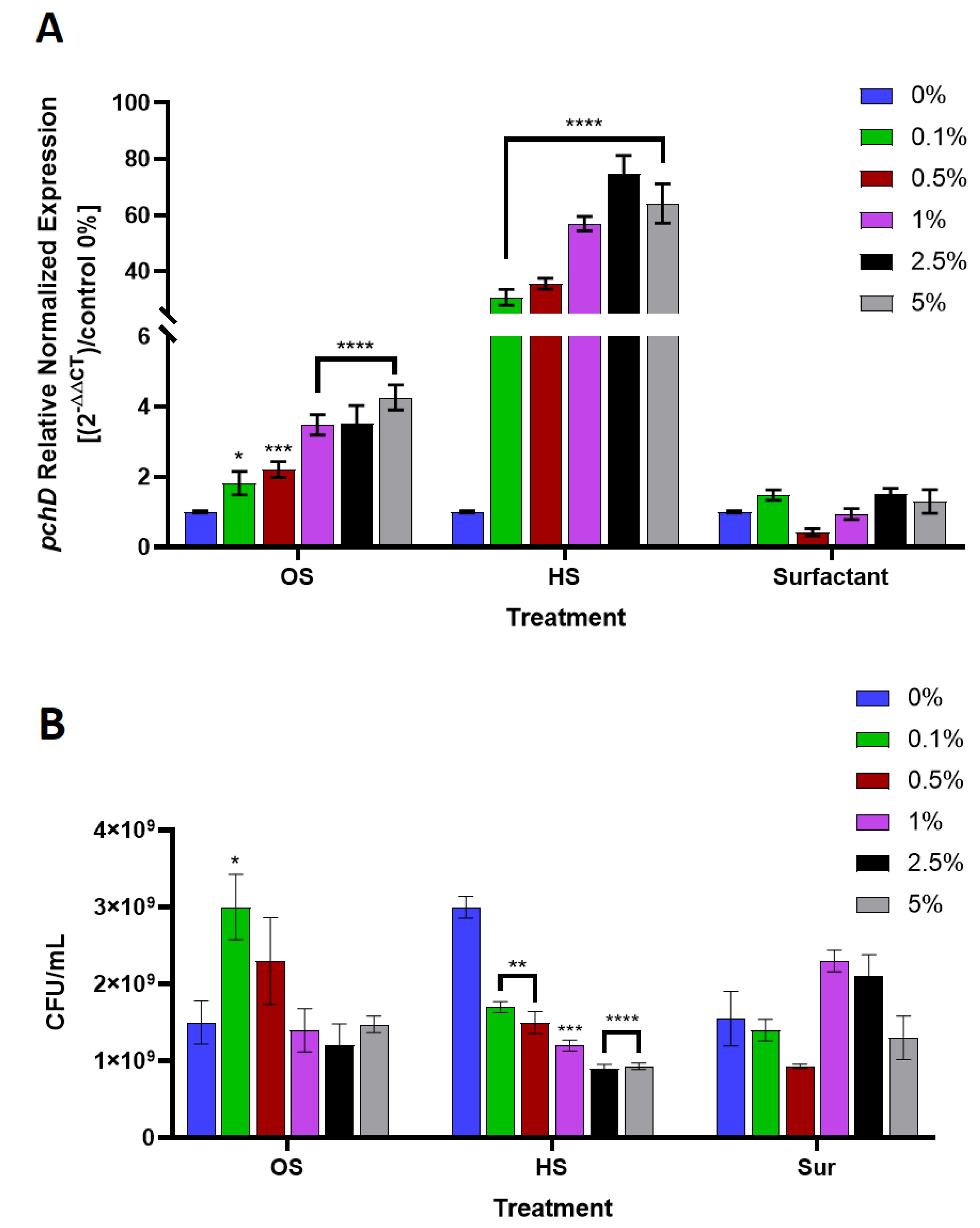
2.5. P. aeruginosa and B. cepacia Growth Culture in the Presence of Octyl Salicylate
2.6. Quantitative Polymerase Chain Reaction for pchD Gene Expression
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lundov, M.D.; Moesby, L.; Zachariae, C.; Johansen, J.D. Contamination versus preservation of cosmetics: A review on legislation, usage, infections, and contact allergy. Contact Dermat. 2009, 60, 70–78. [Google Scholar] [CrossRef]
- Vainio, H.; Bianchini, F. Chemical and physical characteristics of sunscreen constituents. IARC Handb. Cancer Prev. 2001, 5, 17–21. [Google Scholar]
- Davis, J.E. Are one or two dangerous? Methyl salicylate exposure in toddlers. J. Emerg. Med. 2007, 32, 63–69. [Google Scholar] [CrossRef]
- Hong, K.H.; Jang, W.H.; Choi, K.D.; Yoo, O.J. Characterization of Pseudomonas fluorescens Carboxylesterase: Cloning and Expression of the Esterase Gene in Escherichia coli. Agric. Biol. Chem. 1991, 55, 2839–2845. [Google Scholar] [CrossRef] [Green Version]
- Crosa, J.H.; Walsh, C.T. Genetics and Assembly Line Enzymology of Siderophore Biosynthesis in Bacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 223–249. [Google Scholar] [CrossRef] [Green Version]
- Gasser, V.; Guillon, L.; Cunrath, O.; Schalk, I.J. Cellular organization of siderophore biosynthesis in Pseudomonas aeruginosa: Evidence for siderosomes. J. Inorg. Biochem. 2015. [Google Scholar] [CrossRef]
- Serino, L.; Reimmann, C.; Visca, P.; Beyeler, M.; Chiesa, V.D.; Haas, D. Biosynthesis of pyochelin and dihydroaeruginoic acid requires the iron- regulated pchDCBA operon in Pseudomonas aeruginosa. J. Bacteriol. 1997, 179, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.S. Characterizing bacterial resistance to preservatives and disinfectants. Int. Biodeterior. Biodegrad. 1998, 41, 241–245. [Google Scholar] [CrossRef]
- Valkova, N.; Lépine, F.; Bollet, C.; Dupont, M.; Villemur, R. prbA, a Gene Coding for an Esterase Hydrolyzing Parabens in Enterobacter cloacae and Enterobacter gergoviae strains. J. Bacteriol. 2002, 184, 5011–5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiser, R.; Green, A.E.; Bull, M.J.; Cunningham-Oakes, E.; Jolley, K.A.; Maiden, M.C.J.; Hall, A.J.; Winstanley, C.; Weightman, A.J.; Donoghue, D.; et al. Not all Pseudomonas aeruginosa are equal: Strains from industrial sources possess uniquely large multireplicon genomes. Microb. Genom. 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Green, A.E.; Amézquita, A.; Le Marc, Y.; Bull, M.J.; Connor, T.R.; Mahenthiralingam, E. The consistent differential expression of genetic pathways following exposure of an industrial Pseudomonas aeruginosa strain to preservatives and a laundry detergent formulation. FEMS Microbiol. Lett. 2018, 365, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, L.; Sass, A.; Baldwin, A.; Dowson, C.G.; Donoghue, D.; Mahenthiralingam, E. Key role for efflux in the preservative susceptibility and adaptive resistance of Burkholderia cepacia complex bacteria. Antimicrob. Agents Chemother. 2013, 57, 2972–2980. [Google Scholar] [CrossRef] [Green Version]
- Kutty, P.K.; Moody, B.; Gullion, J.S.; Zervos, M.; Ajluni, M.; Washburn, R.; Sanderson, R.; Kainer, M.A.; Powell, T.A.; Clarke, C.F.; et al. Multistate Outbreak of Burkholderia cenocepacia Colonization and Infection Associated with the Use of Intrinsically Contaminated Alcohol-Free Mouthwash. Chest 2007, 132, 1825–1831. [Google Scholar] [CrossRef] [PubMed]
- Molina-Cabrillana, J.; Bolaños-Rivero, M.; Alvarez-León, E.E.; Sánchez, A.M.M.; Sánchez-Palacios, M.; Alvarez, D.; Sáez-Nieto, J.A. Intrinsically Contaminated Alcohol-Free Mouthwash Implicated in a Nosocomial Outbreak of Burkholderia cepacia Colonization and Infection. Infect. Control Hosp. Epidemiol. 2006, 27, 1281–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson, J.R.; Heard, S.R.; Richards, M.A.; Tabaqchali, S. Outbreak of Septicaemia Due to Contaminated Mouthwash. Br. Med. J. 1984, 289, 1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matrician, L.; Ange, G.; Burns, S.; Fanning, W.L.; Kioski, C.; Cage, G.D.; Komatsu, K.K. Outbreak of Nosocomial Burkholderia cepacia Infection and Colonization Associated with Intrinsically Contaminated Mouthwash. Infect. Control Hosp. Epidemiol. 2000, 21, 739–741. [Google Scholar] [CrossRef] [PubMed]
- Neza, E.; Centini, M. Microbiologically Contaminated and Over-Preserved Cosmetic Products According Rapex 2008–2014. Cosmetics 2016, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Savli, H.; Karadenizli, A.; Kolayli, F.; Gundes, S.; Ozbek, U.; Vahaboglu, H. Expression stability of six housekeeping genes: A proposal for resistance gene quantification studies of Pseudomonas aeruginosa by real-time quantitative RT-PCR. J. Med. Microbiol. 2003, 52, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Vial, L.; Marie-Christine, G.; Dekimpe, V.; Deziel, E. Burkholderia Diversity and Versatility: An Inventory of the Extracellular Products. J. Microbiol. Biotechnol. 2007, 17, 1407–1429. [Google Scholar] [CrossRef]
- Breidenstein, E.B.M.; de la Fuente-Núñez, C.; Hancock, R.E.W. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- Nair, B.M.; Cheung, K.-J.; Griffith, A.; Burns, J.L. Salicylate induces an antibiotic efflux pump in Burkholderia cepacia complex genomovar III (B. cenocepacia). J. Clin. Invest. 2004, 113, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.; Lindow, S.E.; Wildermuth, M.C. Salicylic acid, Yersiniabactin, and Pyoverdin Production by the Model Phytopathogen Pseudomonas syringae pv. Tomato DC3000: Synthesis, Regulation, and Impact on Tomato and Arabidopsis Host Plants. J. Bacteriol. 2007, 189, 6773–6786. [Google Scholar] [CrossRef] [Green Version]
- Prithiviraj, B.; Bais, H.P.; Weir, T.; Suresh, B.; Najarro, E.H.; Dayakar, B.V.; Schweizer, H.P.; Vivanco, J.M. Down Regulation of Virulence Factors of Pseudomonas aeruginosa by Salicylic Acid Attenuates Its Virulence on Arabidopsis thaliana and Caenorhabditis elegans. Infect. Immun. 2005, 73, 5319–5328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattab, A.; Djibaoui, R.; Arabi, A.; Hichem, D. Effect of salicylic acid on biofilm formation and on some virulence factors in Pseudomonas aeruginosa. Int. J. Biosci. 2017, 10, 60–71. [Google Scholar] [CrossRef]
- Bandara, M.; Sankaridurg, P.; Zhu, H.; Hume, E.; Willcox, M. Effect of salicylic acid on the membrane proteome and virulence of Pseudomonas aeruginosa. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1213–1220. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.; Jones, J.W.; Ruge, M.A.; Kane, M.A.; Oglesby-Sherrouse, A.G. Iron depletion enhances production of antimicrobials by Pseudomonas aeruginosa. J. Bacteriol. 2015, 197, 2265–2275. [Google Scholar] [CrossRef] [Green Version]
- Leinweber, A.; Weigert, M.; Kümmerli, R. The bacterium Pseudomonas aeruginosa senses and gradually responds to interspecific competition for iron. Evolution (N. Y.) 2018, 72, 1515–1528. [Google Scholar] [CrossRef]
- Burns, J.L.; Clark, D.K. Salicylate-inducible antibiotic resistance in Pseudomonas cepacia associated with absence of a pore-forming outer membrane protein. Antimicrob. Agents Chemother. 1992, 36, 2280–2285. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SPF 30 (+OS) with Preservative AM-25 | ||
|---|---|---|
| Phase A | ||
| Trade Name | INCI Name | %w/w |
| Water | Aqua | Up to 100% |
| Xanthan gum | Xanthan gum | 0.5 |
| Phase B | ||
| Montanov 68 | Cetearyl Glucoside (and) Cetearyl Alcohol | 3 |
| Emulgade® 1000 NI | Cetearyl Alcohol (and) Ceteareth-20 | 3 |
| IPM | Isopropyl myristate | 5 |
| Octyl Salicylate | Octyl Salicylate | 5 |
| Octyl Methoxycinnamate | Ethylhexyl Methoxycinnamate | 7.5 |
| Benzophenone-3 | Benzophenone-3 | 4 |
| Avobenzone | Butyl Methoxydibenzoylmethane | 1.6 |
| Antaron™ V-220F | VP/Eicosene Copolymer | 3 |
| Phase C | ||
| Vitamin E | Tocopheryl acetate | 0.2 |
| Sharomix™ Amplify AM25 (Preservative) | Phenoxyethanol, Chlorphenesin, Caprylyl glycol, Didecyldimethylammonium chloride | 0.5 |
| Final pH 5.0–5.5 | ||
| SPF 45 (+OS) with Preservative AM-25 | ||
| Trade Name | INCI Name | %w/w |
| Phase A | ||
| Water | Aqua | Up to 100% |
| Xanthan gum | Xanthan gum | 0.5 |
| Phase B | ||
| Montanov 68 | Cetearyl Glucoside (and) Cetearyl Alcohol | 3 |
| Emulgade® 1000 NI | Cetearyl Alcohol (and) Ceteareth-20 | 3 |
| TCC | Caprylic/Capric Triglyceride | 5 |
| Octyl Salicylate | Octyl Salicylate | 5 |
| Homosalate | Homosalate | 10 |
| Octyl Methoxycinnamate | Ethylhexyl Methoxycinnamate | 7.5 |
| Benzophenone-3 | Benzophenone-3 | 6 |
| Avobenzone | Butyl Methoxydibenzoylmethane | 1.6 |
| Titanium Dioxide | Titanium Dioxide | 3 |
| Antaron™ V-220F | VP/Eicosene Copolymer | 3 |
| Phase C | ||
| Vitamin E | Tocopheryl acetate | 0.1 |
| Sharomix™ Amplify AM25 (Preservative) | Phenoxyethanol, Chlorphenesin, Caprylyl glycol, Didecyldimethylammonium chloride | 0.5 |
| Final pH 6.5–7.0 | ||
| Basic Cream (+/− OS/HS) with Preservative AM-25 | ||
| Trade Name | INCI Name | %w/w |
| Phase A | ||
| Water | Aqua | Up to 100% |
| Glycerin | Glycerin | 4 |
| Phase B | ||
| Montanov 68 | Cetearyl Glucoside (and) Cetearyl Alcohol | 5 |
| Emulgade® 1000 NI | Cetearyl Alcohol (and) Ceteareth-20 | 4.8 |
| TCC | Caprylic/Capric Triglyceride | 3 |
| Octyl Salicylate/Homosalate | Octyl Salicylate/Homosalate | 5 |
| Phase C | ||
| Vitamin E | Tocopheryl acetate | 0.1 |
| Sharomix™ Amplify AM25 (Preservative) | Phenoxyethanol, Chlorphenesin, Caprylyl glycol, Didecyldimethylammonium chloride | 0.5 |
| Final pH 5.5–6.0 | ||
| Material Name | Function | %w/w |
|---|---|---|
| Sample #1 (OS/HS) | ||
| Phase A | ||
| TSB media | Solvent | 92 |
| Tween 20 (INCI: Polysorbate 20) | Surfactant | 1.5 |
| Phase B | ||
| Span 20 (INCI: Sorbitan Laurate) | Surfactant | 1.5 |
| Octyl salicylate/Homosalate | UV-filter | 5 |
| Final pH 7 | ||
| Sample #2 (Surfactant) | ||
| Phase A | ||
| TSB media | Solvent | 97 |
| Tween 20 (INCI: Polysorbate 20) | Surfactant | 1.5 |
| Phase B | ||
| Span 20 (INCI: Sorbitan Laurate) | Surfactant | 1.5 |
| Final pH 7 | ||
| Primer | Sequence | Gene | Bacteria | Type | Amplicon |
|---|---|---|---|---|---|
| rpoSF [18] | CTCCCCGGGCAACTCCAAAAG | rpoS | P. aeruginosa | Reference | 198 bp |
| rpoSR [18] | CGATCATCCGCTTCCGACCAG | ||||
| pchDF_Ps | GAGAAGATCGCTGCGGAAGA | pchD | P. aeruginosa | Target | 170 bp |
| pchDR_Ps | GCGTGCAGGTATTGCTTCAG | ||||
| recAF | ATCGAGAAGCAGTTCGGCAA | recA | B. cepacia | Reference | 186 bp |
| recAR | CAGCGTGAGCGTGGTTTTAC | ||||
| pchD_BcF | GAAGTGGAGAACCTGCTGCT | pchD | B. cepacia | Target | 146 bp |
| pchD_BcR | CTGGTGCTGAAACGCTACCT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziklo, N.; Tzafrir, I.; Shulkin, R.; Salama, P. Salicylate UV-Filters in Sunscreen Formulations Compromise the Preservative System Efficacy against Pseudomonas aeruginosa and Burkholderia cepacia. Cosmetics 2020, 7, 63. https://doi.org/10.3390/cosmetics7030063
Ziklo N, Tzafrir I, Shulkin R, Salama P. Salicylate UV-Filters in Sunscreen Formulations Compromise the Preservative System Efficacy against Pseudomonas aeruginosa and Burkholderia cepacia. Cosmetics. 2020; 7(3):63. https://doi.org/10.3390/cosmetics7030063
Chicago/Turabian StyleZiklo, Noa, Inbal Tzafrir, Regina Shulkin, and Paul Salama. 2020. "Salicylate UV-Filters in Sunscreen Formulations Compromise the Preservative System Efficacy against Pseudomonas aeruginosa and Burkholderia cepacia" Cosmetics 7, no. 3: 63. https://doi.org/10.3390/cosmetics7030063