Apple Pomace Extract as a Sustainable Food Ingredient

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
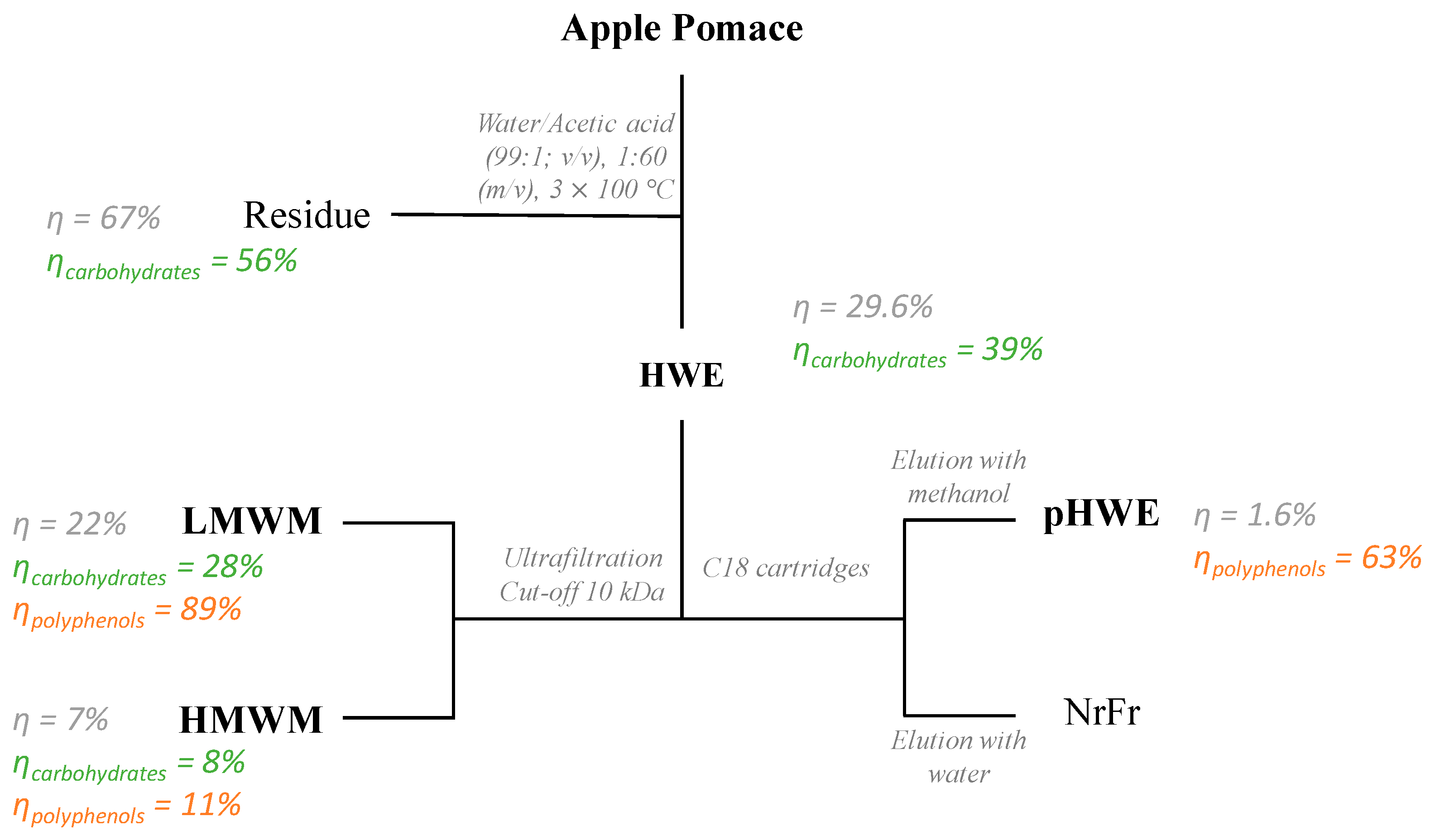
2.2. Preparation of Extracts
2.3. General Chemical Characterization
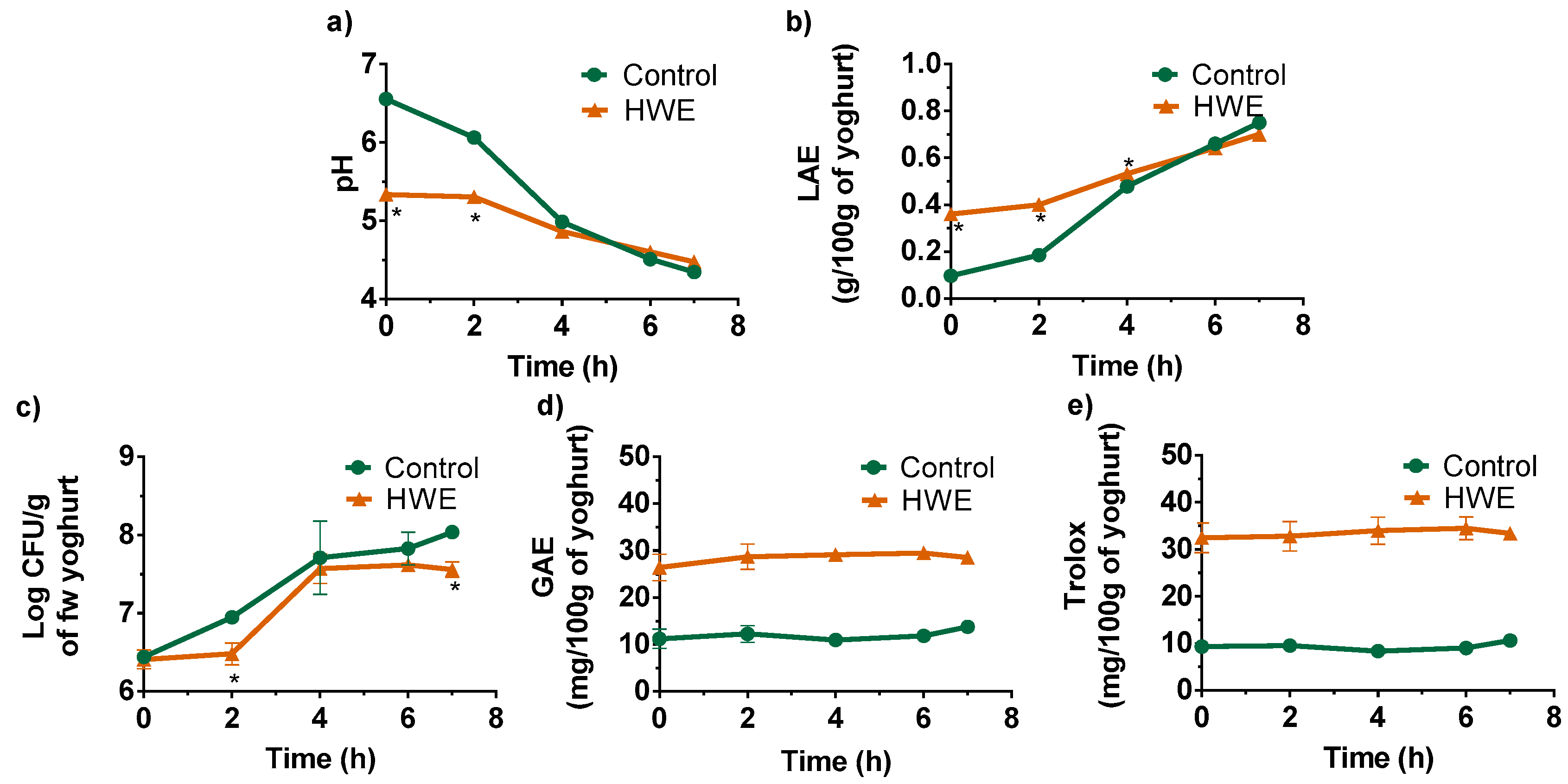
2.4. Formulation of a Hot Water Extract (HWE)-Fortified Yogurt
2.5. Antioxidant Activity
2.6. Anti-Inflammatory Potential
2.6.1. Inhibition of Chemically-Induced NO• Production
2.6.2. Inhibition of NO• Production in LPS-Stimulated Macrophages Cell Line Raw 264.7
2.7. Nutritional Properties of the Yogurt
2.8. Statistical Analysis
3. Results
3.1. Apple Pomace Extracts
3.2. Antioxidant and Anti-Inflammatory Potential
3.3. Application as a Food Ingredient in Yogurt Formulation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- O’Shea, N.; Arendt, E.K.; Gallagher, E. Dietary fibre and phytochemical characteristics of fruit and vegetable by-products and their recent applications as novel ingredients in food products. Innov. Food Sci. Emerg. Technol. 2012, 16, 1–10. [Google Scholar] [CrossRef]
- Rabetafika, H.N.; Bchir, B.; Blecker, C.; Richel, A. Fractionation of apple by-products as source of new ingredients: Current situation and perspectives. Trends Food Sci. Technol. 2014, 40, 99–114. [Google Scholar] [CrossRef]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Mirabella, N.; Castellani, V.; Sala, S. Current options for the valorization of food manufacturing waste: A review. J. Clean. Prod. 2014, 65, 28–41. [Google Scholar] [CrossRef]
- Çam, M.; Aaby, K. Optimization of Extraction of Apple Pomace Phenolics with Water by Response Surface Methodology. J. Agric. Food Chem. 2010, 58, 9103–9111. [Google Scholar] [CrossRef] [PubMed]
- Kschonsek, J.; Wolfram, T.; Stöckl, A.; Böhm, V. Polyphenolic Compounds Analysis of Old and New Apple Cultivars and Contribution of Polyphenolic Profile to the In Vitro Antioxidant Capacity. Antioxidants 2018, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Le Bourvellec, C.; Guyot, S.; Renard, C.M.G.C. Interactions between apple (Malus x domestica Borkh.) polyphenols and cell walls modulate the extractability of polysaccharides. Carbohydr. Polym. 2009, 75, 251–261. [Google Scholar] [CrossRef]
- Fernandes, P.A.R.; Le Bourvellec, C.; Renard, C.M.G.C.; Nunes, F.M.; Bastos, R.; Coelho, E.; Wessel, D.F.; Coimbra, M.A.; Cardoso, S.M. Revisiting the chemistry of apple pomace polyphenols. Food Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Suárez, B.; Álvarez, Á.L.; García, Y.D.; del Barrio, G.; Lobo, A.P.; Parra, F. Phenolic profiles, antioxidant activity and in vitro antiviral properties of apple pomace. Food Chem. 2010, 120, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, M.; List, D.; Lu, Y.; Foo, L.Y.; Newman, R.H.; Sims, I.M.; Bain, P.J.S.; Hamilton, B.; Fenton, G. Apple Pomace and Products Derived from Apple Pomace: Uses, Composition and Analysis. In Analysis of Plant Waste Materials; Linskens, H.F., Jackson, J.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 75–119. [Google Scholar]
- Rohn, S.; Buchner, N.; Driemel, G.; Rauser, M.; Kroh, L.W. Thermal Degradation of Onion Quercetin Glucosides under Roasting Conditions. J. Agric. Food Chem. 2007, 55, 1568–1573. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Gouble, B.; Bureau, S.; Loonis, M.; Plé, Y.; Renard, C.M.G.C. Pink Discoloration of Canned Pears: Role of Procyanidin Chemical Depolymerization and Procyanidin/Cell Wall Interactions. J. Agric. Food Chem. 2013, 61, 6679–6692. [Google Scholar] [CrossRef] [PubMed]
- Passos, C.P.; Ferreira, S.S.; Serôdio, A.; Basil, E.; Marková, L.; Kukurová, K.; Ciesarová, Z.; Coimbra, M.A. Pectic polysaccharides as an acrylamide mitigation strategy—Competition between reducing sugars and sugar acids. Food Hydrocoll. 2018, 81, 113–119. [Google Scholar] [CrossRef]
- Sosulski, F.W.; Imafidon, G.I. Amino acid composition and nitrogen-to-protein conversion factors for animal and plant foods. J. Agric. Food Chem. 1990, 38, 1351–1356. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Brunton, N.P.; Gormley, T.R.; Murray, B. Use of the alditol acetate derivatisation for the analysis of reducing sugars in potato tubers. Food Chem. 2007, 104, 398–402. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Viticult. 1965, 16, 144. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Kunchandy, E.; Rao, M.N.A. Oxygen radical scavenging activity of curcumin. Int. J. Pharm. 1990, 58, 237–240. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Antioxidant and anti-inflammatory activities of Geranium robertianum L. decoctions. Food Funct. 2017, 8, 3355–3365. [Google Scholar] [CrossRef]
- Búfalo, M.C.; Ferreira, I.; Costa, G.; Francisco, V.; Liberal, J.; Cruz, M.T.; Lopes, M.C.; Batista, M.T.; Sforcin, J.M. Propolis and its constituent caffeic acid suppress LPS-stimulated pro-inflammatory response by blocking NF-κB and MAPK activation in macrophages. J. Ethnopharmacol. 2013, 149, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Renard, C.M.G.C.; Lemeunier, C.; Thibault, J.F. Alkaline extraction of xyloglucan from depectinised apple pomace: Optimisation and characterisation. Carbohydr. Polym. 1995, 28, 209–216. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.t.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- European Parliament. The Council of the European Union. Regulation (EU) No 1169/2011. Off. J. Eur. Union 2011, L 304, 18–63. [Google Scholar]
- Percy, A.E.; Melton, L.D.; Jameson, P.E. Xyloglucan and hemicelluloses in the cell wall during apple fruit development and ripening. Plant Sci. 1997, 125, 31–39. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Voragen, A.G.J.; Thibault, J.F.; Pilnik, W. Studies on apple protopectin V: Structural studies on enzymatically extracted pectins. Carbohydr. Polym. 1991, 16, 137–154. [Google Scholar] [CrossRef]
- Ferreira, D.; Guyot, S.; Marnet, N.; Delgadillo, I.; Renard, C.M.G.C.; Coimbra, M.A. Composition of Phenolic Compounds in a Portuguese Pear (Pyrus communis L. Var. S. Bartolomeu) and Changes after Sun-Drying. J. Agric. Food Chem. 2002, 50, 4537–4544. [Google Scholar] [CrossRef] [PubMed]
- Reis, S.F.; Rai, D.K.; Abu-Ghannam, N. Water at room temperature as a solvent for the extraction of apple pomace phenolic compounds. Food Chem. 2012, 135, 1991–1998. [Google Scholar] [CrossRef] [Green Version]
- Guyot, S.; Marnet, N.; Sanoner, P.; Drilleau, J.-F. Variability of the Polyphenolic Composition of Cider Apple (Malus domestica) Fruits and Juices. J. Agric. Food Chem. 2003, 51, 6240–6247. [Google Scholar] [CrossRef]
- Renard, C.M.G.C.; Baron, A.; Guyot, S.; Drilleau, J.F. Interactions between apple cell walls and native apple polyphenols: Quantification and some consequences. Int. J. Biol. Macromol. 2001, 29, 115–125. [Google Scholar] [CrossRef]
- Guyot, S.; Le Bourvellec, C.; Marnet, N.; Drilleau, J.F. Procyanidins are the most Abundant Polyphenols in Dessert Apples at Maturity. LWT Food Sci. Technol. 2002, 35, 289–291. [Google Scholar] [CrossRef]
- Gonçalves, F.J.; Rocha, S.M.; Coimbra, M.A. Study of the retention capacity of anthocyanins by wine polymeric material. Food Chem. 2012, 134, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.J.; Fernandes, P.A.R.; Wessel, D.F.; Cardoso, S.M.; Rocha, S.M.; Coimbra, M.A. Interaction of wine mannoproteins and arabinogalactans with anthocyanins. Food Chem. 2018, 243, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, M.G.; Bastos, R.; Pinto, M.; Ferreira, J.M.; Santos, J.F.; Wessel, D.F.; Coelho, E.; Coimbra, M.A. Waste mitigation: From an effluent of apple juice concentrate industry to a valuable ingredient for food and feed applications. J. Clean. Prod. 2018. [Google Scholar] [CrossRef]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Lamuela-Raventós, R.M.; Viladomat, F.; Bastida, J.; Codina, C. Qualitative analysis of phenolic compounds in apple pomace using liquid chromatography coupled to mass spectrometry in tandem mode. Rapid Commun. Mass Spectrom. 2004, 18, 553–563. [Google Scholar] [CrossRef]
- Millet, M.; Poupard, P.; Le Quéré, J.-M.; Bauduin, R.; Guyot, S. Haze in Apple-Based Beverages: Detailed Polyphenol, Polysaccharide, Protein, and Mineral Compositions. J. Agric. Food Chem. 2017, 65, 6404–6414. [Google Scholar] [CrossRef]
- Nicolas, J.J.; Richard-Forget, F.C.; Goupy, P.M.; Amiot, M.J.; Aubert, S.Y. Enzymatic browning reactions in apple and apple products. Crit. Rev. Food Sci. Nutr. 1994, 34, 109–157. [Google Scholar] [CrossRef]
- Bernillon, S.; Guyot, S.; Renard, C.M.G.C. Detection of phenolic oxidation products in cider apple juice by high-performance liquid chromatography electrospray ionisation ion trap mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 939–943. [Google Scholar] [CrossRef]
- Mouls, L.; Fulcrand, H. Identification of new oxidation markers of grape-condensed tannins by UPLC–MS analysis after chemical depolymerization. Tetrahedron 2015, 71, 3012–3019. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Panchon, M.S.; Villano, D.; Troncoso, A.M.; Garcia-Parrilla, M.C. Antioxidant Activity of Phenolic Compounds: From In Vitro Results to In Vivo Evidence. Crit. Rev. Food Sci. Nutr. 2008, 48, 649–671. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-C.; Shen, S.-C.; Lee, W.-R.; Hou, W.-C.; Yang, L.-L.; Lee, T.J.F. Inhibition of nitric oxide synthase inhibitors and lipopolysaccharide induced inducible NOS and cyclooxygenase-2 gene expressions by rutin, quercetin, and quercetin pentaacetate in RAW 264.7 macrophages. J. Cell. Biochem. 2001, 82, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-C.; Chang, W.-T.; Wu, S.-J.; Xu, P.-Y.; Ting, N.-C.; Liou, C.-J. Phloretin and phlorizin promote lipolysis and inhibit inflammation in mouse 3T3-L1 cells and in macrophage-adipocyte co-cultures. Mol. Nutr. Food Res. 2013, 57, 1803–1813. [Google Scholar] [CrossRef] [PubMed]
- Tseng, A.; Zhao, Y. Wine grape pomace as antioxidant dietary fibre for enhancing nutritional value and improving storability of yogurt and salad dressing. Food Chem. 2013, 138, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.P. Effects of milk-derived bioactives: An overview. Br. J. Nutr. 2000, 84, 3–10. [Google Scholar] [CrossRef]
- Bertolino, M.; Belviso, S.; Dal Bello, B.; Ghirardello, D.; Giordano, M.; Rolle, L.; Gerbi, V.; Zeppa, G. Influence of the addition of different hazelnut skins on the physicochemical, antioxidant, polyphenol and sensory properties of yogurt. LWT Food Sci. Technol. 2015, 63, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.B.; Huo, J.L.; Yang, F.; Chen, X.Q. Noncovalent Interaction of Dietary Polyphenols with Bovine Hemoglobin in Vitro: Molecular Structure/Property–Affinity Relationship Aspects. J. Agric. Food Chem. 2011, 59, 8484–8490. [Google Scholar] [CrossRef]
- Hutkins, R.W.; Nannen, N.L. pH Homeostasis in Lactic Acid Bacteria1. J. Dairy Sci. 1993, 76, 2354–2365. [Google Scholar] [CrossRef]
- Massa, S.; Altieri, C.; Quaranta, V.; De Pace, R. Survival of Escherichia coli O157:H7 in yogurt during preparation and storage at 4 °C. Lett. Appl. Microbiol. 1997, 24, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Massa, S.; Trovatelli, L.D.; Canganella, F. Survival of Listeria monocytogenes in yogurt during storage at 4 °C. Lett. Appl. Microbiol. 1991, 13, 112–114. [Google Scholar] [CrossRef]
- Bruckner, R.; Titgemeyer, F. Carbon catabolite repression in bacteria: Choice of the carbon source and autoregulatory limitation of sugar utilization. FEMS Microbiol. Lett. 2002, 209, 141–148. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Díaz-Rubio, M.E.; Saura-Calixto, F. Non-extractable polyphenols, a major dietary antioxidant: Occurrence, metabolic fate and health effects. Nutr. Res. Rev. 2013, 26, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Lazarte, A.; Kachrimanidou, V.; Villamiel, M.; Rastall, R.A.; Moreno, F.J. In vitro fermentation properties of pectins and enzymatic-modified pectins obtained from different renewable bioresources. Carbohydr. Polym. 2018, 199, 482–491. [Google Scholar] [CrossRef]
- Wang, X.; Kristo, E.; LaPointe, G. The effect of apple pomace on the texture, rheology and microstructure of set type yogurt. Food Hydrocoll. 2019, 91, 83–91. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fractions | Yield (%) | Yield of Carbohydrate (%) | Carbohydrate (mol%) | Total Carbohydrate (%) | Total Protein (%) | Total PC (g GAE/kg) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rha | Fuc | Ara | Xyl | Man | Gal | Glc | Fru | GalA | ||||||||
| Apple pomace | Polysaccharides | 1 | 1 | 12 | 10 | 5 | 9 | 41 | 19 | 53.4 | 71.7 | 5.2 | ND | |||
| Free Sugars | 23 | 77 | 18.3 | |||||||||||||
| HWE | 29.6 | 39.2 | Polysaccharides | 3 | 1 | 25 | 3 | 10 | 9 | 50 | 42.9 | 94.9 | 1.3 | 11 | ||
| Free Sugars | 18 | 82 | 52.0 | |||||||||||||
| HMWM | 6.9 | 7.5 | Polysaccharides | 1 | t | 35 | 6 | 1 | 10 | 9 | 36 | 77.7 | 77.7 | ND | 5 | |
| LMWM | 22.3 | 27.7 | Polysaccharides | 1 | t | 15 | 2 | 6 | 34 | 42 | 33.5 | 89.0 | ND | 9 | ||
| Free Sugars | 6 | 94 | 55.5 | |||||||||||||
| Residue | 67.4 | 55.9 | Polysaccharides | 1 | 1 | 8 | 14 | 4 | 8 | 51 | 13 | 59.5 | 59.5 | 7.1 | ||
| N° | RT | Compound | λmax | MS (m/z) | MS2 (m/z) | Extract |
|---|---|---|---|---|---|---|
| pHWE | ||||||
| 1 | 12.3 | Quercetin-3-O-rutinoside a | 254, 353 | 609 | 463, 301 | 3.27 ± 0.06 |
| 2 | 12.6 | Quercetin-3-O-galactoside b | 256, 354 | 463 | 301 | 31.37 ± 0.32 |
| 3 | 12.7 | Quercetin-3-O-glucoside a | 256, 353 | 463 | 301 | 8.45 ± 0.10 |
| 4 | 13.2 | Quercetin-3-O-xylanoside b | 256, 354 | 433 | 301 | 8.88 ± 0.10 |
| 5 | 13.4 | Quercetin 3-O-arabinopyranoside b | 243, 352 | 433 | 301 | 2.31 ± 0.04 |
| 6 | 13.5 | Quercetin 3-O-arabinofuranoside b | 256, 352 | 433 | 301 | 15.09 ± 0.16 |
| 7 | 13.7 | Quercetin-O-pentoside b | 256, 351 | 433 | 301 | 3.32 ± 0.05 |
| 8 | 13.8 | Quercetin-3-O-rhamnoside b | 256, 350 | 447 | 301 | 26.05 ± 0.27 |
| 9 | 14.9 | Phloretin-2-O-glucoside a | 227, 284 | 435 | 273 | 15.96 ± 0.20 |
| Total | 114.75 ± 1.25 |
| Extract | TPC | DPPH• | ABTS•+ | OH• | NO• |
|---|---|---|---|---|---|
| HWE | 10.7 ± 0.2 | 1339 ± 211 (14.2 ± 1.7) | 532 ± 11.5 (5.69 ± 0.12) | - | - |
| pHWE | 149 ± 1.87 | 82.4 ± 11.2 (12.3 ± 1.7) | 35.2 ± 5.9 (5.23 ± 0.44) | 6.75 ± 0.45 | 35.2 ± 5.9 |
| AA | - | 2.70 ± 0.30 | 2.68 ± 0.03 | - | 57.3 ± 2.3 |
| Components | Control | HWE |
|---|---|---|
| Moisture | 88.8 ± 0.0 | 87.2 ± 0.1 |
| Total Sugars | 5.45 ± 0.12 | 7.30 ± 0.18 |
| Reducing Sugars | 4.49 ± 0.10 | 5.05 ± 0.04 |
| HWE polysaccharides * | 1.43 ± 0.01 | |
| Protein | 3.32 ± 0.04 | 3.21 ± 0.08 |
| Fat | 1.68 ± 0.15 | 1.46 ± 0.07 |
| Ash | 0.75 ± 0.01 | 0.84 ± 0.01 |
| Energetic (kcal) | 46.5 ± 1.1 | 49.1 ± 0.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, P.A.R.; Ferreira, S.S.; Bastos, R.; Ferreira, I.; Cruz, M.T.; Pinto, A.; Coelho, E.; Passos, C.P.; Coimbra, M.A.; Cardoso, S.M.; et al. Apple Pomace Extract as a Sustainable Food Ingredient. Antioxidants 2019, 8, 189. https://doi.org/10.3390/antiox8060189
Fernandes PAR, Ferreira SS, Bastos R, Ferreira I, Cruz MT, Pinto A, Coelho E, Passos CP, Coimbra MA, Cardoso SM, et al. Apple Pomace Extract as a Sustainable Food Ingredient. Antioxidants. 2019; 8(6):189. https://doi.org/10.3390/antiox8060189
Chicago/Turabian StyleFernandes, Pedro A. R., Sónia S. Ferreira, Rita Bastos, Isabel Ferreira, Maria T. Cruz, António Pinto, Elisabete Coelho, Cláudia P. Passos, Manuel A. Coimbra, Susana M. Cardoso, and et al. 2019. "Apple Pomace Extract as a Sustainable Food Ingredient" Antioxidants 8, no. 6: 189. https://doi.org/10.3390/antiox8060189