From Orphan Phage to a Proposed New Family–The Diversity of N4-Like Viruses

 , and
, and
Abstract
:1. Introduction
2. Results
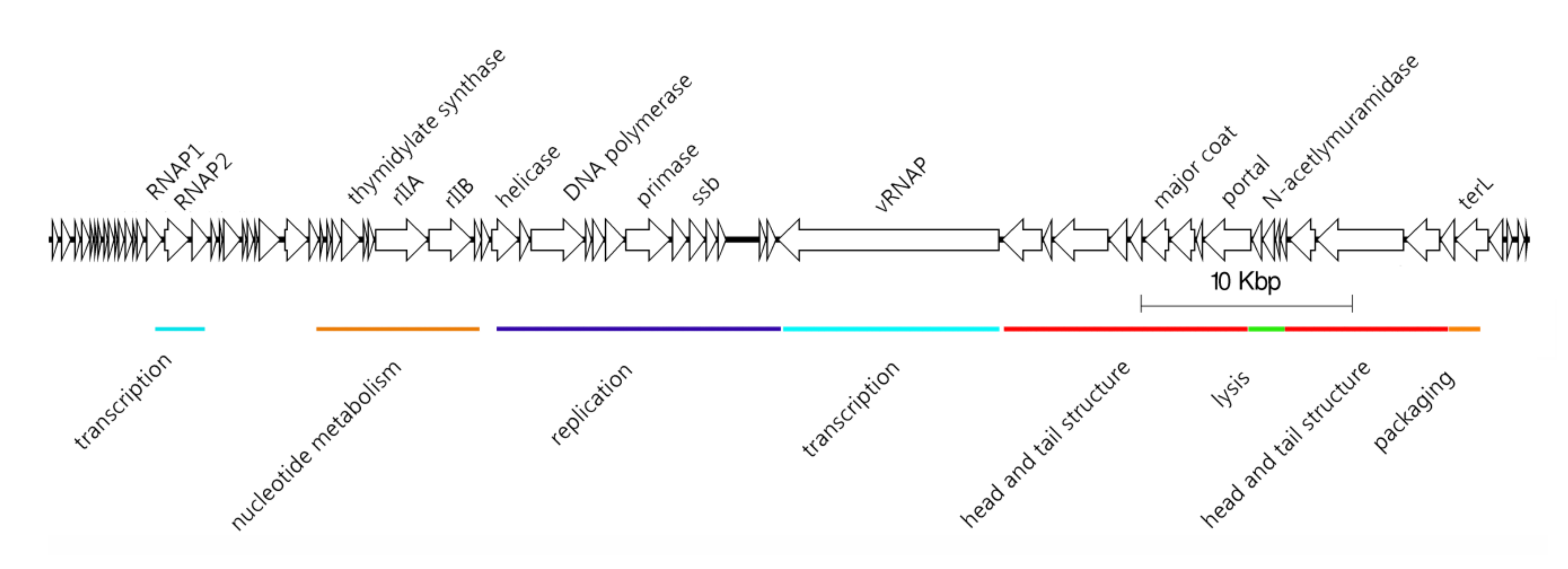
2.1. Description of N4-Like Viruses
- Podovirus morphology
- Genome size of 59–80 kb
- Linear genome with defined ends (terminal repeats expected)
- Presence of three RNA polymerase genes, including a large (~3500 aa) virion-associated RNA polymerase (vRNAP)
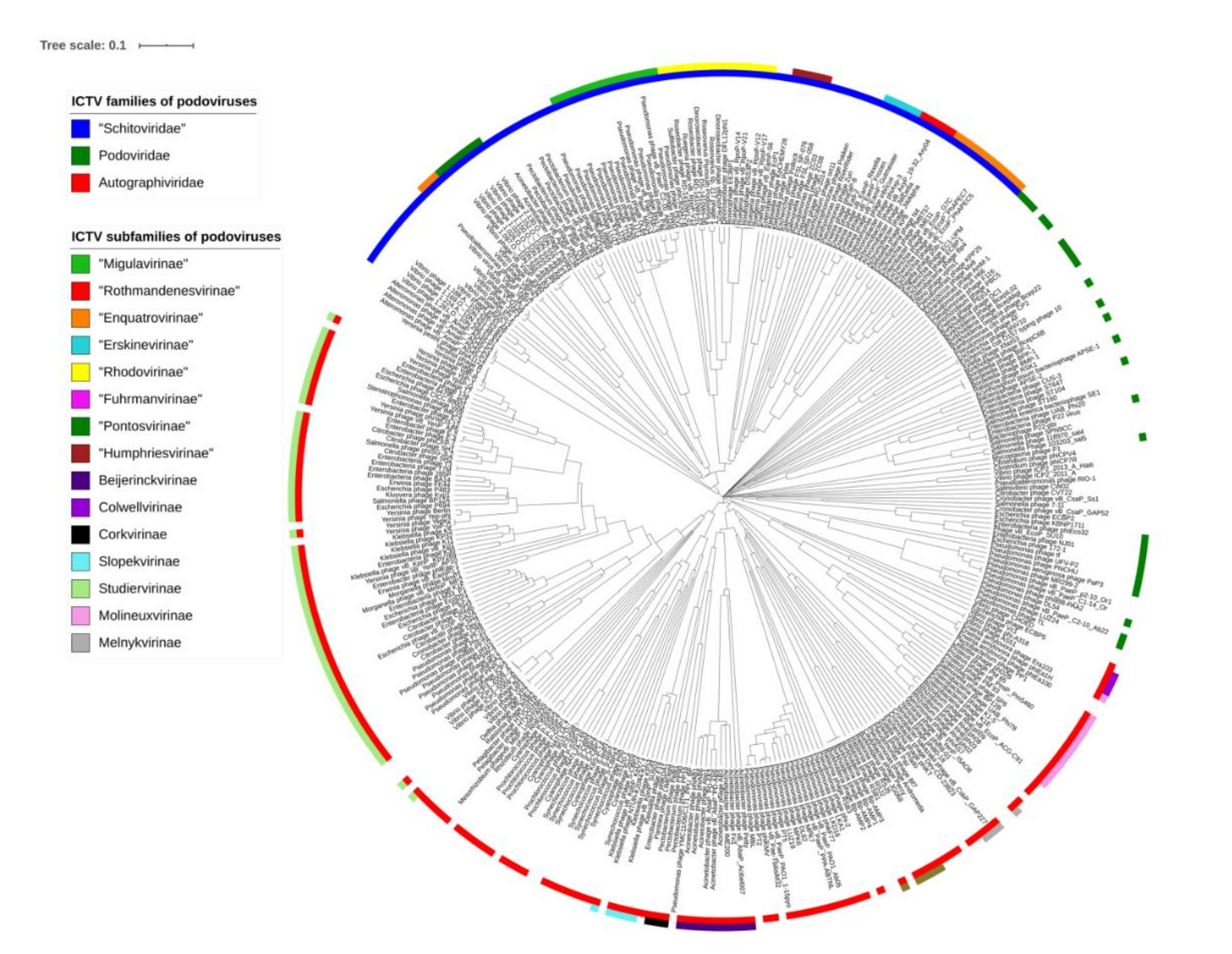
2.2. Proposal of a New Family
2.3. Proposal of New Subfamilies and Genera
3. Discussion
4. Materials and Methods
4.1. vConTACT2 Analysis
4.2. Panproteome Analysis
4.3. VIRIDIC Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schito, G.C.; Rialdi, G.; Pesce, A. Biophysical properties of N4 coliphage. Biochim. Biophys. Acta 1966, 129, 482–490. [Google Scholar] [CrossRef]
- Ohmori, H.; Haynes, L.L.; Rothman-Denes, L.B. Structure of the ends of the coliphage N4 genome. J. Mol. Biol. 1988, 202, 1–10. [Google Scholar] [CrossRef]
- Paddison, P.; Abedon, S.T.; Dressman, H.K.; Gailbreath, K.; Tracy, J.; Mosser, E.; Neitzel, J.; Guttman, B.; Kutter, E. The roles of the bacteriophage T4 r genes in lysis inhibition and fine-structure genetics: A new perspective. Genetics 1998, 148, 1539–1550. [Google Scholar] [PubMed]
- Kazmierczak, K.M.; Rothman-Denes, L.B. Bacteriophage N4. In The Bacteriophages, 2nd ed.; Calendar, R., Ed.; Oxford University Press: New York, NY, USA, 2006; pp. 302–314. [Google Scholar]
- Falco, S.C.; Laan, K.V.; Rothman-Denes, L.B. Virion-associated RNA polymerase required for bacteriophage N4 development. Proc. Natl. Acad. Sci. USA 1977, 74, 520–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zivin, R.; Zehring, W.; Rothman-Denes, L.B. Transcriptional map of bacteriophage N4: Location and polarity of N4 RNAs. J. Mol. Biol. 1988, 152, 335–356. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.W.; DuBow, M. Natural groups of bacteriophages. In Viruses of Prokaryotes; CRC Press: Boca Raton, FL, USA, 1987; pp. 85–100. [Google Scholar]
- Wittmann, J.; Klumpp, J.; Moreno Switt, A.I.; Yagubi, A.; Ackermann, H.W.; Wiedemann, M.; Svircev, A.; Nash, H.E.; Kropinski, A.M. Taxonomic reassessment of N4-like viruses using comparative genomics and proteomics suggests a new subfamily-“Enquartavirinae”. Arch. Virol. 2015, 160, 3053–3062. [Google Scholar] [CrossRef]
- Kropinski, A.M. Bacteriophage research–what we have learnt and what still needs to be addressed. Res. Microbiol. 2018, 169, 481–487. [Google Scholar] [CrossRef]
- Adriaenssens, E.M.; Sullivan, M.B.; Knezevic, P.; van Zyl, L.J.; Sarkar, B.L.; Dutilh, B.E.; Alfenas-Zerbini, P.; Łobocka, M.; Tong, Y.; Brister, J.R.; et al. Taxonomy of prokaryotic viruses: 2018–2019 update from the ICTV Bacterial and Archaeal Viruses Subcommittee. Arch. Virol. 2020, 165, 1253–1260. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.; Brister, J.R. How to name and classify your phage: An informal guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Karsch-Mizrachi, I.; Nakamura, Y.; Cochrane, G. The international nucleotide sequence database collaboration. Nucleic Acids Res. 2012, 40, 33–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brister, J.R.; Ako-adjei, D.; Bao, Y.; Blinkova, O. NCBI viral genomes resource. Nucleic. Acids. Res. 2015, 43, 571–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittmann, J.; Dreiseikelmann, B.; Rohde, M.; Meier-Kolthoff, J.P.; Bunk, B.; Rohde, C. First genome sequences of Achromobacter phages reveal new members of the N4 family. Virol. J. 2014, 27, 11–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Li, E.; Qi, Z.; Li, H.; Wei, X.; Lin, W.; Zhao, R.; Jiang, A.; Yang, H.; Yin, Z.; et al. Isolation and molecular characterisation of Achromobacter phage phiAxp-3, an N4-like bacteriophage. Sci. Rep. 2016, 6, 24776. [Google Scholar] [CrossRef] [Green Version]
- Essoh, C.; Vernadet, J.P.; Vergnaud, G.; Coulibaly, A.; Kakou-N’Douba, A.; N’Guetta, A.S.P.; Ouassa, T.; Pourcel, C. Characterization of sixteen Achromobacter xylosoxidans phages from Abidjan, Côte d’Ivoire, isolated on a single clinical strain. Arch. Virol. 2020, 165, 725–730. [Google Scholar] [CrossRef]
- Farmer, N.G.; Wood, T.L.; Chamakura, K.R.; Kuty Everett, G.F. Complete genome of Acinetobacter baumannii N4-Like podophage Presley. Genome Announc. 2013, 1, e00852-13. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Luo, M.; Xi, H.; Zhao, Y.; Le, S.; Chen, L.K.; Tan, D.; Guan, Y.; Wang, T.; Han, W.; et al. The characteristics and genome analysis of vB_ApiP_XC38, a novel phage infecting Acinetobacter pittii. Virus Genes 2020, 56, 498–507. [Google Scholar] [CrossRef]
- Bhattacharjee, A.S.; Motlagh, A.M.; Gilcrease, E.B.; Imdadul Islam, M.I.; Casjens, S.R.; Goel, R. Complete genome sequence of lytic bacteriophage RG-2014 that infects the multidrug resistant bacterium Delftia tsuruhatensis ARB-1. Stand. Genom. Sci. 2017, 12, 82. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhang, S.; Long, L.; Huang, S. Characterization and complete genome sequences of three N4-like roseobacter phages isolated from the South China sea. Curr. Microbiol. 2016, 73, 409–418. [Google Scholar] [CrossRef]
- Cai, L.; Yang, Y.; Jiao, N.; Zhang, R. Complete genome sequence of vB_DshP-R2C, a N4-like lytic roseophage. Mar. Genom. 2015, 22, 15–17. [Google Scholar] [CrossRef]
- Park, J.; Lee, G.M.; Kim, D.; Park, D.H.; Oh, C.S. Characterization of the lytic bacteriophage phiEaP-8 effective against both Erwinia amylovora and Erwinia pyrifoliae causing severe diseases in apple and pear. plant. Pathol. J. 2018, 34, 445–450. [Google Scholar]
- Esplin, I.N.D.; Berg, J.A.; Sharma, R.; Allen, R.C.; Arens, D.K.; Ashcroft, C.R.; Bairett, S.R.; Beatty, N.J.; Bickmore, M.; Bloomfield, T.J.; et al. Genome sequences of 19 novel Erwinia amylovora bacteriophages. Genome Announc. 2017, 5, e00931-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Born, Y.; Fieseler, L.; Marazzi, J.; Lurz, R.; Duffy, B.; Loessner, M.J. Novel virulent and broad-host-range Erwinia amylovora bacteriophages reveal a high degree of mosaicism and a relationship to Enterobacteriaceae phages. Appl. Environ. Microbiol. 2011, 77, 5945–5954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Ma, Y.; Wang, T.; Sun, H.; Lu, G.; Ren, H. Characterization and complete genome sequence of vB_EcoP-Bp4, a novel polyvalent N4-like bacteriophage that infects chicken pathogenic Escherichia coli. Virol. Sin. 2016, 31, 353–356. [Google Scholar] [CrossRef]
- Gan, H.M.; Sieo, C.C.; Tang, S.G.H.; Omar, A.R.; Ho, Y.W. The complete genome sequence of EC1-UPM, a novel N4-like bacteriophage that infects Escherichia coli O78:K80. Virol. J. 2013, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- Nho, S.W.; Ha, M.A.; Kim, K.S.; Kim, T.H.; Jang, H.B.; Cha, I.S.; Park, S.B.; Kim, Y.K.; Jung, T.S. Complete genome sequence of the bacteriophages ECBP1 and ECBP2 isolated from two different Escherichia coli strains. J. Virol. 2012, 86, 12439–12440. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Fan, H.; An, X.; Huang, Y.; Zhang, Z.; Mi, Z.; Tong, Y. Complete genome sequence of IME11, a new N4-like bacteriophage. J. Virol. 2012, 86, 13861. [Google Scholar] [CrossRef] [Green Version]
- Akindolire, M.A.; Aremu, B.R.; Ateba, C.N. Complete genome sequence of Escherichia coli O157:H7 phage PhiG17. Microbiol. Resour. Announc. 2019, 8, e01296-18. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.S.; Lessor, L.E.; Hernandez, A.C.; Kuty Everett, G.F. Complete genome sequence of enterotoxigenic Escherichia coli N4-like podophage pollock. Genome Announc. 2015, 3, e01431-14. [Google Scholar] [CrossRef] [Green Version]
- Golomidova, A.K.; Kulikov, E.E.; Babenko, V.V.; Kostryukova, E.S.; Letarov, A.V. Complete genome sequence of bacteriophage St11Ph5, which infects uropathogenic Escherichia coli strain up11. Genome Announc. 2018, 6, e01371-17. [Google Scholar] [CrossRef] [Green Version]
- Kulikov, E.; Kropinski, A.M.; Golomidova, A.; Lingohr, E.; Govorun, V.; Serebryakova, M.; Prokhorov, N.; Letarova, M.; Manykin, A.; Strotskaya, A.; et al. Isolation and characterization of a novel indigenous intestinal N4-related coliphage vB_EcoP_G7C. Virology 2012, 426, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsonos, J.; Oosterik, L.H.; Tuntufye, H.N.; Klumpp, J.; Butaye, P.; De Greve, H.; Hernalsteens, J.P.; Lavigne, R.; Goddeeris, B.M. A cocktail of in vitro efficient phages is not a guarantee for in vivo therapeutic results against avian colibacillosis. Vet. Microbiol. 2014, 171, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Morozova, V.; Babkin, I.; Kozlova, Y.; Baykov, I.; Bokovaya, O.; Tikunov, A.; Ushakova, T.; Bardasheva, A.; Ryabchikova, E.; Zelentsova, E.; et al. Isolation and characterization of a novel Klebsiella pneumoniae N4-like bacteriophage KP8. Viruses 2019, 11, 1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerushalmy, O.; Coppenhagen-Glazer, S.; Nir-Paz, R.; Tuomala, H.; Skurnik, M.; Kiljunen, S.; Hazan, R. Complete genome sequences of two Klebsiella pneumoniae phages isolated as part of an international effort. Microbiol. Resour. Announc. 2019, 8, e00843-19. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.E.; Lessor, L.; O’Leary, C.; Gill, J.; Liu, M. Complete genome sequence of Klebsiella pneumoniae podophage pylas. Microbiol. Resour. Announc. 2019, 8, e01287-19. [Google Scholar] [CrossRef] [Green Version]
- Smolarska, A.; Rabalski, L.; Narajczyk, M.; Czajkowski, R. Isolation and phenotypic and morphological characterization of the first Podoviridae lytic bacteriophages ϕA38 and ϕA41 infecting Pectobacterium parmentieri (former Pectobacterium wasabiae). Eur. J. Plant. Pathol. 2018, 150, 413–425. [Google Scholar] [CrossRef]
- Buttimer, C.; Hendrix, H.; Lucid, A.; Neve, H.; Noben, J.P.; Franz, C.; O’Mahony, J.; Lavigne, R.; Coffey, A. Novel N4-Like Bacteriophages of Pectobacterium atrosepticum. Pharmaceuticals (Basel) 2018, 11, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazimierczak, J.; Wójcik, E.A.; Witaszewska, J.; Guziński, A.; Górecka, E.; Stańczyk, M.; Kaczorek, E.; Siwicki, A.K.; Dastych, J. Complete genome sequences of Aeromonas and Pseudomonas phages as a supportive tool for development of antibacterial treatment in aquaculture. Virol. J. 2019, 16, 4. [Google Scholar] [CrossRef]
- Alves, D.R.; Perez-Esteban, P.; Kot, W.; Bean, J.E.; Arnot, T.; Hansen, L.H.; Enright, M.C.; Jenkins, A.T. A novel bacteriophage cocktail reduces and disperses Pseudomonas aeruginosa biofilms under static and flow conditions. Microb. Biotechnol. 2016, 9, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Shigehisa, R.; Uchiyama, J.; Kato, S.I.; Takemura-Uchiyama, I.; Yamaguchi, K.; Miyata, R.; Ujihara, T.; Sakaguchi, Y.; Okamoto, N.; Shimakura, H.; et al. Characterization of Pseudomonas aeruginosa phage KPP21 belonging to family Podoviridae genus N4-like viruses isolated in Japan. Microbiol. Immunol. 2016, 60, 64–67. [Google Scholar] [CrossRef] [Green Version]
- Ceyssens, P.J.; Brabban, A.; Rogge, L.; Lewis, M.S.; Pickard, D.; Goulding, D.; Dougan, G.; Noben, J.P.; Kropinski, A.; Kutter, E.; et al. Molecular and physiological analysis of three Pseudomonas aeruginosa phages belonging to the “N4-like viruses”. Virology 2010, 405, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Zhao, F.; Sun, H.; Yu, X.; Zhang, C.; Liu, W.; Pan, Q.; Ren, H. Characterization and complete genome analysis of Pseudomonas aeruginosa bacteriophage vB_PaeP_LP14 belonging to genus Litunavirus. Curr. Microbiol. 2020, 77, 2465–2474. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Cha, K.E.; Myung, H. Complete genome of Pseudomonas aeruginosa phage PA26. J. Virol. 2012, 86, 10244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrowes, B.H.; Molineux, I.J.; Fralick, J.A. Directed in vitro evolution of therapeutic bacteriophages: The appelmans protocol. Viruses 2019, 11, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essoh, C.; Latino, L.; Midoux, C.; Blouin, Y.; Loukou, G.; Nguetta, S.P.A.; Lathro, S.; Cablanmian, A.; Kouassi, A.K.; Vergnaud, G.; et al. Investigation of a large collection of Pseudomonas aeruginosa bacteriophages collected from a single environmental source in Abidjan, Côte d’Ivoire. PLoS ONE 2015, 10, e0130548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forti, F.; Roach, D.R.; Cafora, M.; Pasini, M.E.; Horner, D.S.; Fiscarelli, E.V.; Rossitto, M.; Cariani, L.; Briani, F.; Debarbieux, L.; et al. Design of a broad-range bacteriophage cocktail that reduces pseudomonas aeruginosa biofilms and treats acute infections in two animal models. Antimicrob. Agents Chemother. 2018, 62, e02573-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatek, M.; Parasion, S.; Rutyna, P.; Mizak, L.; Gryko, R.; Niemcewicz, M.; Olender, A.; Łobocka, M. Isolation of bacteriophages and their application to control Pseudomonas aeruginosa in planktonic and biofilm models. Res. Microbiol. 2017, 168, 194–207. [Google Scholar] [CrossRef]
- Gu, J.; Li, X.; Yang, M.; Du, C.; Cui, Z.; Gong, P.; Xia, F.; Song, J.; Zhang, L.; Li, J.; et al. Therapeutic effect of Pseudomonas aeruginosa phage YH30 on mink hemorrhagic pneumonia. Vet. Microbiol. 2016, 190, 5–11. [Google Scholar] [CrossRef]
- Yang, M.; Du, C.; Gong, P.; Xia, F.; Sun, C.; Feng, X.; Lei, L.; Song, J.; Zhang, L.; Wang, B.; et al. Therapeutic effect of the YH6 phage in a murine hemorrhagic pneumonia model. Res. Microbiol. 2015, 166, 633–643. [Google Scholar] [CrossRef]
- Amgarten, D.; Martins, L.F.; Lombardi, K.C.; Principal Antunes, L.; Silva de Souza, A.P.; Gonçalves Nicastro, G.; Kitajima, E.W.; Quaggio, R.B.; Upton, C.; Setubal, J.C.; et al. Three novel Pseudomonas phages isolated from composting provide insights into the evolution and diversity of tailed phages. BMC Genom. 2017, 18, 346. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.Z.M.; Millard, A.D.; Mann, N.H.; Schäfer, H. Comparative genomics defines the core genome of the growing N4-like phage genus and identifies N4-like roseophage specific genes. Front. Microbiol. 2014, 5, 506. [Google Scholar] [CrossRef] [PubMed]
- Moreno Switt, A.I.; Orsi, R.H.; den Bakker, H.C.; Vongkamjan, K.; Altier, C.; Wiedmann, M. Genomic characterization provides new insight into Salmonella phage diversity. BMC Genom. 2013, 14, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.W.; Yun, S.K.; Kim, H.J.; Chai, J.Y.; Park, S.C. Characterization and complete genome sequence of a novel N4-like bacteriophage, pSb-1 infecting Shigella boydii. Res. Microbiol. 2014, 165, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, K.; Jiao, N.; Chen, F. Genome sequences of two novel phages infecting marine roseobacters. Environ. Microbiol. 2009, 11, 2055–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubo, M.T.; Alías-Villegas, C.; Balsanelli, E.; Mesa, D.; de Souza, E.; Espuny, M.R. Diversity of Sinorhizobium (Ensifer) meliloti bacteriophages in the rhizosphere of Medicago marina: Myoviruses, filamentous and N4-like podovirus. Front. Microbiol. 2020, 11, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, A.; Martinez, N.; Moreland, R.; Liu, M.; Gonzalez, C.F.; Gill, J.J.; Ramsey, J. Complete genome sequence of Stenotrophomonas phage pokken. Microbiol. Resour. Announc. 2019, 8, e01095-19. [Google Scholar] [CrossRef] [Green Version]
- Ankrah, N.Y.D.; Budinoff, C.R.; Wilson, W.H.; Wilhelm, S.W.; Buchan, A. Genome sequence of the sulfitobacter sp. strain 2047–infecting lytic phage Φ CB2047–B. Genome Announc. 2014, 2, e00945-13. [Google Scholar] [CrossRef] [Green Version]
- Fouts, D.E.; Klumpp, J.; Bishop-Lilly, K.A.; Rajavel, M.; Willner, K.M.; Butani, A.; Henry, M.; Biswas, B.; Li, M.; Albert, M.J.; et al. Whole genome sequencing and comparative genomic analyses of two Vibrio cholerae O139 Bengal-specific podoviruses to other N4-like phages reveal extensive genetic diversity. Virol. J. 2013, 10, 165. [Google Scholar] [CrossRef] [Green Version]
- Naser, I.B.; Hoque, M.M.; Abdullah, A.; Bari, N.S.M.; Ghosh, A.N.; Faruque, S.M. Environmental bacteriophages active on biofilms and planktonic forms of toxigenic Vibrio cholerae: Potential relevance in cholera epidemiology. PLoS ONE 2017, 12, e0180838. [Google Scholar] [CrossRef] [Green Version]
- Bhandare, S.; Colom, J.; Baig, A.; Ritchie, J.M.; Bukhari, H.; Shah, M.A.; Sarkar, B.L.; Su, J.; Wren, B.; Barrow, P.; et al. Reviving phage therapy for the treatment of cholera. J. Infect. Dis. 2019, 219, 786–794. [Google Scholar] [CrossRef]
- Katharios, P.; Kalatzis, P.G.; Kokkari, C.; Sarropoulou, E.; Middelboe, M. Isolation and characterization of a N4-like lytic bacteriophage infecting Vibrio splendidus, a pathogen of fish and bivalves. PLoS ONE 2017, 12, e0190083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Park, S.C. Complete Genome Sequence of a Bacteriophage, pVco-5, That infects Vibrio coralliilyticus, which causes bacillary necrosis in Pacific Oyster (Crassostrea gigas) Larvae. Genome Announc. 2018, 6, e01143-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.; Deiulio, A.; Holland, C.; Douthitt, C.; McMahon, J.; Wiersma-Koch, H.; Turechek, W.W.; D’Elia, T. Complete genome sequence of Xanthomonas phage RiverRider, a novel N4-like bacteriophage that infects the strawberry pathogen Xanthomonas fragariae. Arch. Virol. 2020, 165, 1481–1484. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The viral proteomic tree server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Rohwer, F.; Edwards, R. The phage proteomic tree: A genome-based taxonomy for phage. J. Bacteriol. 2002, 184, 4529–4535. [Google Scholar] [CrossRef] [Green Version]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2020. [Google Scholar] [CrossRef]
- Barylski, J.; Enault, F.; Dutilh, B.E.; Schuller, M.B.P.; Edwards, R.A.; Gillis, A.; Klumpp, J.; Knezevic, P.; Krupovic, M.; Kuhn, J.H.; et al. Analysis of spounaviruses as a case study for the overdue reclassification of tailed phages. Syst. Biol. 2020, 69, 110–123. [Google Scholar] [CrossRef] [Green Version]
- Preprint: Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC–a novel tool to calculate the intergenomic similarities of prokaryote–infecting viruses. bioRxiv 2020. [Google Scholar] [CrossRef]
- Michniewski, S.; Redgwell, T.; Grigonyte, A.; Rihtman, B.; Aguilo-Ferretjans, M.; Christie-Oleza, J.; Jameson, E.; Scanlan, D.J.; Millard, A.D. Riding the wave of genomics to investigate aquatic coliphage diversity and activity. Environ. Microbiol. 2019, 21, 2112–2128. [Google Scholar] [CrossRef] [Green Version]
- Bin Jang, H.; Bolduc, B.; Zablocki, O.; Kuhn, J.H.; Roux, S.; Adriaenssens, E.M.; Brister, R.; Kropinski, A.M.; Krupovic, M.; Lavigne, R.; et al. Taxonomic assignment of uncultivated prokaryotic virus genomes is enabled by gene-sharing networks. Nat. Biotechnol. 2019, 37, 632–639. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Lechner, M.; Findeisz, S.; Steiner, L.; Marz, M.; Stadler, P.F.; Prohaska, S.J. Proteinortho: Detection of (co-) orthologs in large-scale analysis. BMC Bioinform. 2011, 12, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayliss, S.C.; Thorpe, H.A.; Coyle, N.M.; Sheppard, S.K.; Feil, E.J. PIRATE: A fast and scalable pangenomics toolbox for clustering diverged orthologues in bacteria. Gigascience 2019, 8, giz119. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a versatile software package for scalable and robust microbial pangenome analysis. Appl. Env. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Phage | Subfamily | Genus | Reference |
|---|---|---|---|
| Achromobacter phage JWAlpha | “Rothmandenesvirinae” | Jwalphavirus | [15] |
| Achromobacter phage JWDelta | “Rothmandenesvirinae” | Jwalphavirus | [15] |
| Achromobacter phage phiAxp–3 | “Rothmandenesvirinae” | “Dongdastvirus” | [16] |
| Achromobacter phage vB_AxyP_19–32_Axy04 | “Rothmandenesvirinae” | “Dongdastvirus” | [17] |
| Achromobacter phage vB_AxyP_19–32_Axy10 | “Rothmandenesvirinae” | “Pourcelvirus” | [17] |
| Achromobacter phage vB_AxyP_19–32_Axy11 | “Rothmandenesvirinae” | “Pourcelvirus” | [17] |
| Achromobacter phage vB_AxyP_19–32_Axy12 | “Rothmandenesvirinae” | “Dongdastvirus” | [17] |
| Achromobacter phage vB_AxyP_19–32_Axy13 | “Rothmandenesvirinae” | “Inbricusvirus” | [17] |
| Achromobacter phage vB_AxyP_19–32_Axy22 | “Rothmandenesvirinae” | “Pourcelvirus” | [17] |
| Achromobacter phage vB_AxyP_19–32_Axy24 | “Rothmandenesvirinae” | “Dongdastvirus” | [17] |
| Acinetobacter phage Presley | “Presleyvirus” | [18] | |
| Acinetobacter phage VB_ApiP_XC38 | [19] | ||
| Delftia phage RG–2014 | “Dendoorenvirus” | [20] | |
| Dinoroseobacter phage DFL12phi1 | “Rhodovirinae” | Baltimorevirus | |
| Dinoroseobacter phage DS–1410Ws–06 | “Rhodovirinae” | “Sanyabayvirus” | [21] |
| Dinoroseobacter phage vBDshPR2C | “Rhodovirinae” | Baltimorevirus | [22] |
| Enterobacter phage EcP1 | “Eceepunavirus” | ||
| Enterobacteria phage N4 | “Enquatrovirinae” | Enquatrovirus | [3] |
| Erwinia phage Ea9–2 | “Erskinevirinae” | Johnsonvirus | |
| Erwinia phage phiEaP–8 | “Erskinevirinae” | Yonginvirus | [23] |
| Erwinia phage vB_EamP_Frozen | “Erskinevirinae” | Johnsonvirus | [24] |
| Erwinia phage vB_EamP_Gutmeister | “Erskinevirinae” | Johnsonvirus | [24] |
| Erwinia phage vB_EamP_Rexella | “Erskinevirinae” | Johnsonvirus | [24] |
| Erwinia phage vB_EamP–S6 | “Waedenswilvirus” | [25] | |
| Escherichia phage Bp4 | “Enquatrovirinae” | Gamaleyavirus | [26] |
| Escherichia phage EC1–UPM | “Enquatrovirinae” | Gamaleyavirus | [27] |
| Escherichia phage ECBP1 | “Enquatrovirinae” | Gamaleyavirus | [28] |
| Escherichia phage IME11 | “Enquatrovirinae” | Gamaleyavirus | [29] |
| Escherichia phage OLB145 | “Enquatrovirinae” | Enquatrovirus | |
| Escherichia phage PD38 | “Enquatrovirinae” | Gamaleyavirus | |
| Escherichia phage PGN829.1 | “Enquatrovirinae” | Gamaleyavirus | |
| Escherichia phage phi G17 | “Enquatrovirinae” | Gamaleyavirus | [30] |
| Escherichia phage PMBT57 | “Enquatrovirinae” | Enquatrovirus | |
| Escherichia phage Pollock | “Humphriesvirinae” | “Pollockvirus” | [31] |
| Escherichia phage St11Ph5 | “Enquatrovirinae” | Gamaleyavirus | [32] |
| Escherichia phage vB_EcoP_3HA13 | “Enquatrovirinae” | Enquatrovirus | |
| Escherichia phage vB_EcoP_G7C | “Enquatrovirinae” | Gamaleyavirus | [33] |
| Escherichia phage vB_EcoP_PhAPEC5 | “Enquatrovirinae” | Gamaleyavirus | [34] |
| Escherichia phage vB_EcoP_PhAPEC7 | “Enquatrovirinae” | Gamaleyavirus | [34] |
| Klebsiella phage KP8 | “Enquatrovirinae” | “Kaypoctavirus” | [35] |
| Klebsiella phage KpCHEMY26 | “Humphriesvirinae” | “Pylasvirus” | [36] |
| Klebsiella phage Pylas | “Humphriesvirinae” | “Pylasvirus” | [37] |
| Pectobacterium phage Nepra | “Cbunavirus” | ||
| Pectobacterium phage phiA41 | “Cbunavirus” | [38] | |
| Pectobacterium phage vB_PatP_CB1 | “Cbunavirus” | [39] | |
| Pectobacterium phage vB_PatP_CB3 | “Cbunavirus” | [39] | |
| Pectobacterium phage vB_PatP_CB4 | “Cbunavirus” | [39] | |
| Pseudoalteromonas phage pYD6-A | “Fuhrmanvirinae” | “Mazuvirus” | |
| Pseudomonas phage 98PfluR60PP | “Littlefixvirus” | [40] | |
| Pseudomonas phage DL64 | “Migulavirinae” | Litunavirus | [41] |
| Pseudomonas phage inbricus | “Rothmandenesvirinae” | “Inbricusvirus” | |
| Pseudomonas phage KPP21 | “Migulavirinae” | Luzseptimavirus | [42] |
| Pseudomonas phage LIT1 | “Migulavirinae” | Litunavirus | [43] |
| Pseudomonas phage Littlefix | “Littlefixvirus” | ||
| Pseudomonas phage LP14 | “Migulavirinae” | Litunavirus | [44] |
| Pseudomonas phage LUZ7 | “Migulavirinae” | Luzseptimavirus | [43] |
| Pseudomonas phage LY218 | “Migulavirinae” | Litunavirus | |
| Pseudomonas phage Pa2 | “Migulavirinae” | Litunavirus | |
| Pseudomonas phage PA26 | “Migulavirinae” | Litunavirus | [45] |
| Pseudomonas phage PEV2 | “Migulavirinae” | Litunavirus | [43] |
| Pseudomonas phage phCDa | Shizishanvirus | ||
| Pseudomonas phage phi176 | “Migulavirinae” | Litunavirus | [46] |
| Pseudomonas phage RWG | “Migulavirinae” | Litunavirus | [46] |
| Pseudomonas phage vB_Pae1396P-5 | “Migulavirinae” | Litunavirus | |
| Pseudomonas phage vB_Pae575P-3 | “Migulavirinae” | Litunavirus | |
| Pseudomonas phage vB_PaeP_C2–10_Ab09 | “Migulavirinae” | Litunavirus | [47] |
| Pseudomonas phage vB_PaeP_DEV | “Migulavirinae” | Litunavirus | [48] |
| Pseudomonas phage vB_PaeP_MAG4 | “Migulavirinae” | Litunavirus | [49] |
| Pseudomonas phage vB_PaeP_PYO2 | “Migulavirinae” | Litunavirus | [48] |
| Pseudomonas phage YH30 | “Migulavirinae” | Litunavirus | [50] |
| Pseudomonas phage YH6 | “Migulavirinae” | Litunavirus | [51] |
| Pseudomonas phage ZC03 | “Zicotriavirus” | [52] | |
| Pseudomonas phage ZC08 | “Zicotriavirus” | [52] | |
| Pseudomonas phage Zuri | “Zurivirus” | ||
| Roseobacter phage RD–1410W1–01 | “Rhodovirinae” | “Aoquinvirus” | [21] |
| Roseobacter phage RD–1410Ws–07 | “Rhodovirinae” | “Sanyabayvirus” | [21] |
| Roseovarius Plymouth podovirus 1 | “Rhodovirinae” | “Plymouthvirus” | [53] |
| Roseovarius sp. 217 phage 1 | “Rhodovirinae” | “Plymouthvirus” | [53] |
| Ruegeria phage vB_RpoP–V12 | “Rhodovirinae” | “Aorunvirus” | |
| Ruegeria phage vB_RpoP–V13 | “Rhodovirinae” | “Pomeroyivirus” | |
| Ruegeria phage vB_RpoP–V14 | “Rhodovirinae” | “Aorunvirus” | |
| Ruegeria phage vB_RpoP–V17 | “Rhodovirinae” | “Aorunvirus” | |
| Ruegeria phage vB_RpoP–V21 | “Rhodovirinae” | “Aorunvirus” | |
| Salmonella phage FSL SP–058 | “Humphriesvirinae” | “Ithacavirus” | [54] |
| Salmonella phage FSL SP–076 | “Humphriesvirinae” | “Ithacavirus” | [54] |
| Shigella phage pSb–1 | “Enquatrovirinae” | Gamaleyavirus | [55] |
| Silicibacter phage DSS3phi2 | “Rhodovirinae” | “Aorunvirus” | [56] |
| Sinorhizobium phage ort11 | “Huelvavirus” | [57] | |
| Stenotrophomonas phage Pokken | “Pokkenvirus” | [58] | |
| Sulfitobacter phage EE36phi1 | “Rhodovirinae” | “Aorunvirus” | [56] |
| Sulfitobacter phage phiCB2047-B | “Rhodovirinae” | “Raunefjordvirus” | [59] |
| Vibrio phage 1.025.O._10N.222.46.B6 | “Pontosvirinae” | “Nahantvirus” | |
| Vibrio phage 1.026.O._10N.222.49.C7 | “Pontosvirinae” | “Nahantvirus” | |
| Vibrio phage 1.097.O._10N.286.49.B3 | “Pontosvirinae” | “Dorisvirus” | |
| Vibrio phage 1.150.O._10N.222.46.A6 | “Pontosvirinae” | “Nahantvirus” | |
| Vibrio phage 1.152.O._10N.222.46.E1 | “Pontosvirinae” | “Nahantvirus” | |
| Vibrio phage 1.169.O._10N.261.52.B1 | Mukerjeevirus | ||
| Vibrio phage 1.188.A._10N.286.51.A6 | Mukerjeevirus | ||
| Vibrio phage 1.224.A._10N.261.48.B1 | Mukerjeevirus | ||
| Vibrio phage 1.261.O._10N.286.51.A7 | Mukerjeevirus | ||
| Vibrio phage 2.130.O._10N.222.46.C2 | “Pontosvirinae” | “Nahantvirus” | |
| Vibrio phage JA–1 | “Pacinivirus” | [60] | |
| Vibrio phage JSF3 | “Pacinivirus” | [61] | |
| Vibrio phage phi 1 | “Pacinivirus” | [62] | |
| Vibrio phage phi50–12 | |||
| Vibrio phage pVa5 | “Pontosvirinae” | “Galateavirus” | [63] |
| Vibrio phage pVco–5 | [64] | ||
| Vibrio phage VBP32 | “Fuhrmanvirinae” | “Stoningtonvirus” | |
| Vibrio phage VBP47 | “Fuhrmanvirinae” | “Stoningtonvirus” | |
| Vibrio phage VCO139 | “Pacinivirus” | [60] | |
| Vibrio virus vB_VspP_SBP1 | |||
| Xanthomonas phage RiverRider | “Riverridervirus” | [65] | |
| From metagenomes | |||
| Podoviridae sp. isolate ctda_1 | |||
| Podoviridae sp. ctLUJ1 | |||
| Siphoviridae sp. isolate 355 | “Enquatrovirinae” | Gamaleyavirus |
| # | N4_Product | N4 Locus Tag | N4 Protein Accession | PIRATE | Proteinortho_30 | CoreGenes 5.0 |
|---|---|---|---|---|---|---|
| 1 | RNAP 1 | EPNV4_gp15 | YP_950493.1 | Y | Y | Y |
| 2 | RNAP 2 | EPNV4_gp16 | YP_950494.1 | N | Y | Y |
| 3 | AAA+ ATPase | EPNV4_gp24 | YP_950502.1 | * | Y | Y |
| 4 | gp25 | EPNV4_gp25 | YP_950503.1 | N | N | Y |
| 5 | DNA polymerase | EPNV4_gp39 | YP_950517.1 | Y | Y | Y |
| 6 | gp42 | EPNV4_gp42 | YP_950520.1 | Y | Y | Y |
| 7 | DNA primase | EPNV4_gp43 | YP_950521.1 | Y | Y | Y |
| 8 | gp44 | EPNV4_gp44 | YP_950522.1 | Y | Y | Y |
| 9 | vRNAP | EPNV4_gp50 | YP_950528.1 | N | N | Y |
| 10 | 16.5 kDa protein | EPNV4_gp52 | YP_950530.1 | Y | Y | Y |
| 11 | gp54 | EPNV4_gp54 | YP_950532.1 | N | N | Y |
| 12 | Major capsid protein | EPNV4_gp56 | YP_950534.1 | Y | Y | Y |
| 13 | gp57 (tape measure) | EPNV4_gp57 | YP_950535.1 | N | * | Y |
| 14 | 94 kDa protein (portal vertex protein) | EPNV4_gp59 | YP_950537.1 | Y | Y | Y |
| 15 | 30 kDa protein | EPNV4_gp67 | YP_950545.1 | N | N | Y |
| 16 | Terminase, large subunit | EPNV4_gp68 | YP_950546.1 | Y | Y | Y |
| 17 | gp69 | EPNV4_gp69 | YP_950547.1 | Y | N | Y |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wittmann, J.; Turner, D.; Millard, A.D.; Mahadevan, P.; Kropinski, A.M.; Adriaenssens, E.M. From Orphan Phage to a Proposed New Family–The Diversity of N4-Like Viruses. Antibiotics 2020, 9, 663. https://doi.org/10.3390/antibiotics9100663
Wittmann J, Turner D, Millard AD, Mahadevan P, Kropinski AM, Adriaenssens EM. From Orphan Phage to a Proposed New Family–The Diversity of N4-Like Viruses. Antibiotics. 2020; 9(10):663. https://doi.org/10.3390/antibiotics9100663
Chicago/Turabian StyleWittmann, Johannes, Dann Turner, Andrew D. Millard, Padmanabhan Mahadevan, Andrew M. Kropinski, and Evelien M. Adriaenssens. 2020. "From Orphan Phage to a Proposed New Family–The Diversity of N4-Like Viruses" Antibiotics 9, no. 10: 663. https://doi.org/10.3390/antibiotics9100663