
Markers for Ongoing or Previous Hepatitis E Virus Infection Are as Common in Wild Ungulates as in Humans in Sweden

Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Total HEV Antibodies (Anti-HEV) Detection
2.3. HEV RNA Detection
2.3.1. Nucleic Acid Extraction
2.3.2. Reverse Transcription Real-Time PCR (RT-qPCR) Assay
2.3.3. cDNA Synthesis
2.3.4. PCR Amplification for Sequencing
2.3.5. Controls Used in the Procedures
2.3.6. Sequencing
2.3.7. Phylogenetic Analysis
2.3.8. Statistical Analysis
3. Results
3.1. Anti-HEV Detection
3.2. HEV RNA Detection
3.3. HEV Markers (Anti-HEV and/or HEV RNA) and Geographical Origin of the Animals
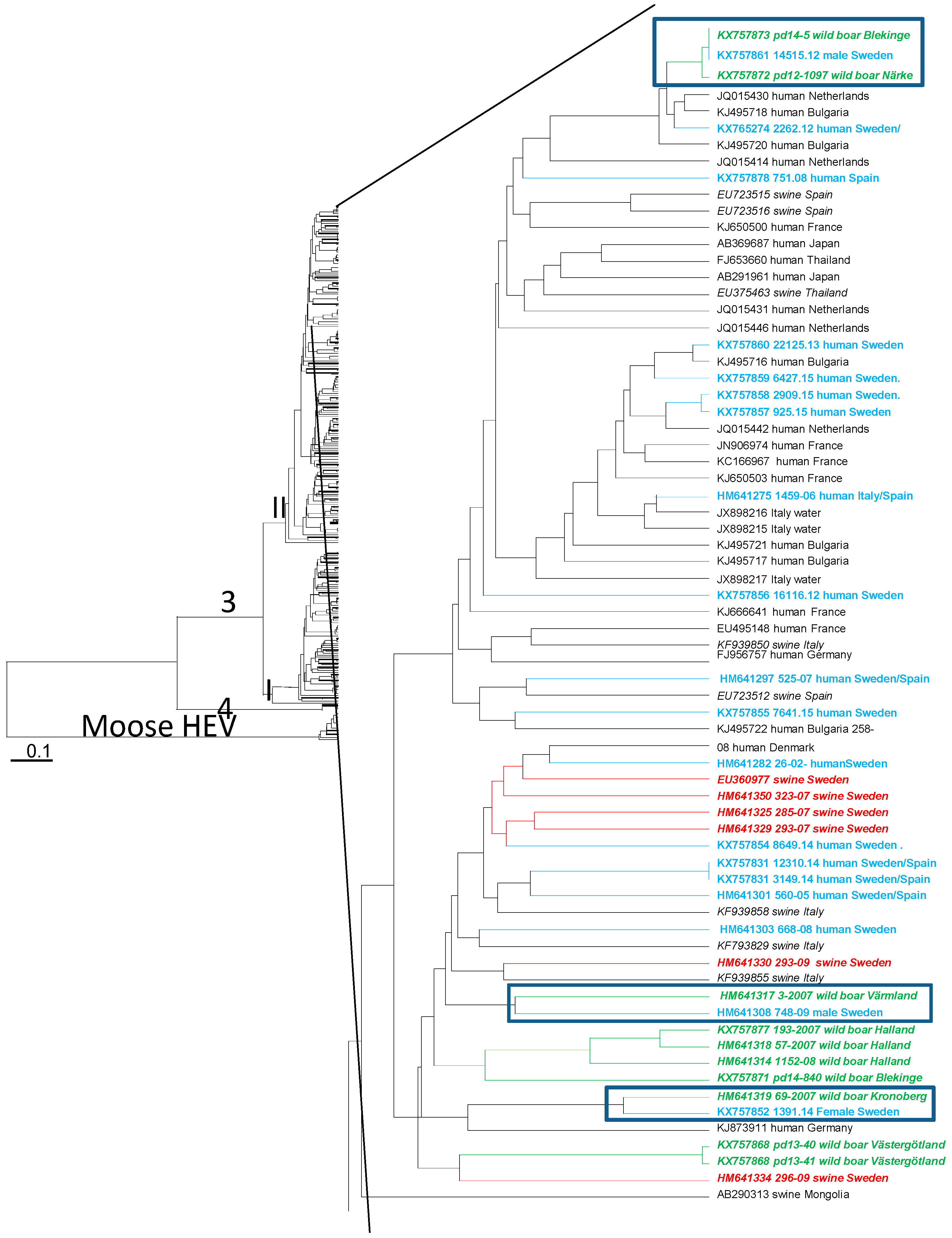
3.4. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bradley, D.W.; Krawczynski, K.; Cook, E.H., Jr.; McCaustland, K.A.; Humphrey, C.D.; Spelbring, J.E.; Myint, H.; Maynard, J.E. Enterically transmitted non-A, non-B hepatitis: Serial passage of disease in cynomolgus macaques and tamarins and recovery of disease-associated 27- to 34-nm viruslike particles. Proc. Natl. Acad. Sci. USA 1987, 84, 6277–6281. [Google Scholar] [CrossRef] [PubMed]
- Holla, R.P.; Ahmad, I.; Ahmad, Z.; Jameel, S. Molecular virology of hepatitis E virus. Semin. Liver Dis. 2013, 33, 3–14. [Google Scholar] [PubMed]
- Smith, D.B.; Simmonds, P.; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; van der Poel, W.H.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2015, 96, 1191–1192. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.J. Zoonotic and foodborne transmission of hepatitis E virus. Semin. Liver Dis. 2013, 33, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Owolodun, O.A.; Gerber, P.F.; Gimenez-Lirola, L.G.; Kwaga, J.K.; Opriessnig, T. First report of hepatitis E virus circulation in domestic pigs in Nigeria. Am. J. Trop. Med. Hyg. 2014, 91, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Kaba, M.; Moal, V.; Gerolami, R.; Colson, P. Epidemiology of mammalian hepatitis E virus infection. Intervirology 2013, 56, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Caruso, C.; Modesto, P.; Prato, R.; Scaglione, F.E.; de Marco, L.; Bollo, E.; Acutis, P.L.; Masoero, L.; Peletto, S. Hepatitis E virus: First description in a pet house rabbit. A new transmission route for human? Transbound. Emerg. Dis. 2015, 62, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Yugo, D.M.; Meng, X.J. Hepatitis E virus: Foodborne, waterborne and zoonotic transmission. Int. J. Environ. Res. Public Health 2013, 10, 4507–4533. [Google Scholar] [CrossRef] [PubMed]
- Garbuglia, A.R.; Scognamiglio, P.; Petrosillo, N.; Mastroianni, C.M.; Sordillo, P.; Gentile, D.; la Scala, P.; Girardi, E.; Capobianchi, M.R. Hepatitis E virus genotype 4 outbreak, Italy, 2011. Emerg. Infect. Dis. 2013, 19, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Romanet, P.; Moal, V.; Borentain, P.; Purgus, R.; Benezech, A.; Motte, A.; Gerolami, R. Autochthonous infections with hepatitis E virus genotype 4, France. Emerg. Infect. Dis. 2012, 18, 1361–1364. [Google Scholar] [CrossRef] [PubMed]
- Midgley, S.; Vestergaard, H.T.; Dalgaard, C.; Enggaard, L.; Fischer, T.K. Hepatitis E virus genotype 4, Denmark, 2012. Emerg. Infect. Dis. 2014, 20, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Mansuy, J.M. Hepatitis E in high-income countries. Curr. Opin. Infect. Dis. 2010, 23, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Meng, X.J.; Renou, C. Zoonotic hepatitis E: Animal reservoirs and emerging risks. Vet. Res. 2010, 41, 46. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Nishizawa, T.; Nagashima, S.; Jirintai, S.; Kawakami, M.; Sonoda, Y.; Suzuki, T.; Yamamoto, S.; Shigemoto, K.; Ashida, K.; et al. Molecular characterization of a novel hepatitis E virus (HEV) strain obtained from a wild boar in Japan that is highly divergent from the previously recognized HEV strains. Virus Res. 2014, 180, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Terada, S.; Kokuryu, H.; Arai, M.; Mishiro, S. A wild boar-derived hepatitis E virus isolate presumably representing so far unidentified “genotype 5”. Kanzo 2010, 51, 536–538. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New hepatitis E virus genotype in camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Norder, H.; Uhlhorn, H.; Belak, S.; Widen, F. Novel hepatitis E like virus found in Swedish moose. J. Gen. Virol. 2014, 95, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Labrique, A.B.; Zaman, K.; Hossain, Z.; Saha, P.; Yunus, M.; Hossain, A.; Ticehurst, J.; Kmush, B.; Nelson, K.E. An exploratory case control study of risk factors for hepatitis E in rural Bangladesh. PLoS ONE 2013, 8, e61351. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S.; Kamili, S. Aetiology, clinical course and outcome of sporadic acute viral hepatitis in pregnancy. J. Viral Hepat. 2003, 10, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolo, I.; Angeloni, G.; Ponterio, E.; Ostanello, F.; Ruggeri, F.M. Detection of hepatitis E virus in pork liver sausages. Int. J. Food Microbiol. 2015, 193, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Abravanel, F.; Lhomme, S.; Dubois, M.; Peron, J.M.; Alric, L.; Kamar, N.; Izopet, J. Hepatitis E virus. Med. Mal. Infect. 2013, 43, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Emerson, S.U.; Purcell, R.H. Hepatitis E. Pediatr. Infect. Dis. J. 2007, 26, 1147–1148. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Selves, J.; Mansuy, J.M.; Ouezzani, L.; Peron, J.M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E virus and chronic hepatitis in organ-transplant recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Mansuy, J.M.; Bendall, R.; Legrand-Abravanel, F.; Saune, K.; Miedouge, M.; Ellis, V.; Rech, H.; Destruel, F.; Kamar, N.; Dalton, H.R.; et al. Hepatitis E virus antibodies in blood donors, France. Emerg. Infect. Dis. 2011, 17, 2309–2312. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.R.; Kamar, N.; Izopet, J. Hepatitis E in developed countries: Current status and future perspectives. Future Microbiol. 2014, 9, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.R.; Kamar, N.; van Eijk, J.J.; McLean, B.N.; Cintas, P.; Bendall, R.P.; Jacobs, B.C. Hepatitis E virus and neurological injury. Nat. Rev. Neurol 2016, 12, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Olsen, B.; Axelsson-Olsson, D.; Thelin, A.; Weiland, O. Unexpected high prevalence of IgG-antibodies to hepatitis E virus in Swedish pig farmers and controls. Scand. J. Infect. Dis. 2006, 38, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Sylvan, S.P. The high rate of antibodies to hepatitis E virus in young, intravenous drug-abusers with acute hepatitis B-virus infection in a Swedish community: A study of hepatitis markers in individuals with intravenously or sexually acquired hepatitis B-virus infection. Scand. J. Infect. Dis. 1998, 30, 429–430. [Google Scholar] [PubMed]
- Norder, H.K.M.; Mellgren, Å.; Konar, J.; Sandberg, E.; Lasson, A.; Castedal, M.; Magnius, L.; Lagging, M. Diagnostic performance of five assays for anti-HEV IgG and IgM in a large cohort study. J. Clin. Microbiol. 2016, 54, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Caruso, C.; Peletto, S.; Rosamilia, A.; Modesto, P.; Chiavacci, L.; Sona, B.; Balsamelli, F.; Ghisetti, V.; Acutis, P.L.; Pezzoni, G.; et al. Hepatitis E virus: A cross-sectional serological and virological study in pigs and humans at zoonotic risk within a high-density pig farming area. Transbound. Emerg. Dis. 2016. [Google Scholar] [CrossRef] [PubMed]
- Public Health Agency of Sweden. Numer of Reported Hepatitis E Cases in Sweden. Available online: https://www.folkhalsomyndigheten.se/about-folkhalsomyndigheten-the-public-health-agency-of-sweden/ (accessed on 20 June 2016).
- Jansson, G.M.J.; Magnusson, M. How many wild boars are there? Svensk Jakt 2010, 4, 86–87. (In Swedish) [Google Scholar]
- Jägarförbundet, Wild Animals. Available online: http://jagareforbundet.se/vilt/vilt-vetande/artpresentationer-ny/ (accessed on 20 June 2016). (In Swedish)
- Lin, J.; Karlsson, M.; Olofson, A.S.; Belak, S.; Malmsten, J.; Dalin, A.M.; Widen, F.; Norder, H. High prevalence of hepatitis E virus in Swedish moose—A phylogenetic characterization and comparison of the virus from different regions. PLoS ONE 2015, 10, e0122102. [Google Scholar] [CrossRef] [PubMed]
- Grierson, S.; Heaney, J.; Cheney, T.; Morgan, D.; Wyllie, S.; Powell, L.; Smith, D.; Ijaz, S.; Steinbach, F.; Choudhury, B.; et al. Prevalence of hepatitis E virus infection in pigs at the time of slaughter, United Kingdom, 2013. Emerg. Infect. Dis. 2015, 21, 1396–1401. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Inferring phylogeneies from protein sequences by parsimony, distance and likelyhood methods. Methods Enzymol. 1996, 266, 418–427. [Google Scholar] [PubMed]
- Norder, H.; Sundqvist, L.; Magnusson, L.; Ostergaard Breum, S.; Lofdahl, M.; Larsen, L.E.; Hjulsager, C.K.; Magnius, L.; Bottiger, B.E.; Widen, F. Endemic hepatitis E in two Nordic countries. Eurosurveillance 2009, 14, 20–28. [Google Scholar]
- Widen, F.; Sundqvist, L.; Matyi-Toth, A.; Metreveli, G.; Belak, S.; Hallgren, G.; Norder, H. Molecular epidemiology of hepatitis E virus in humans, pigs and wild boars in Sweden. Epidemiol. Infect. 2011, 139, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Larska, M.; Krzysiak, M.K.; Jablonski, A.; Kesik, J.; Bednarski, M.; Rola, J. Hepatitis E virus antibody prevalence in wildlife in Poland. Zoonoses Public Health 2015, 62, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Martelli, F.; Caprioli, A.; Zengarini, M.; Marata, A.; Fiegna, C.; di Bartolo, I.; Ruggeri, F.M.; Delogu, M.; Ostanello, F. Detection of hepatitis E virus (HEV) in a demographic managed wild boar (Sus scrofa scrofa) population in Italy. Vet. Microbiol. 2008, 126, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Montagnaro, S.; de Martinis, C.; Sasso, S.; Ciarcia, R.; Damiano, S.; Auletta, L.; Iovane, V.; Zottola, T.; Pagnini, U. Viral and antibody prevalence of hepatitis E in European wild boars (Sus scrofa) and hunters at zoonotic risk in the Latium region. J. Comp. Pathol. 2015, 153, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, N.; Pavoni, E.; Filogari, D.; Ferrari, N.; Chiari, M.; Canelli, E.; Lombardi, G. Hepatitis E virus in wild boar in the central northern part of Italy. Transbound. Emerg. Dis. 2015, 62, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C.; Wolf, A.; Meisel, H.; Kaiser, M.; Ellerbrok, H.; Pauli, G. High HEV presence in four different wild boar populations in East and West Germany. Vet. Microbiol. 2009, 139, 270–278. [Google Scholar] [CrossRef] [PubMed]
- De Deus, N.; Peralta, B.; Pina, S.; Allepuz, A.; Mateu, E.; Vidal, D.; Ruiz-Fons, F.; Martin, M.; Gortazar, C.; Segales, J. Epidemiological study of hepatitis E virus infection in European wild boars (Sus scrofa) in Spain. Vet. Microbiol. 2008, 129, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Rutjes, S.A.; Lodder-Verschoor, F.; Lodder, W.J.; van der Giessen, J.; Reesink, H.; Bouwknegt, M.; de Roda Husman, A.M. Seroprevalence and molecular detection of hepatitis E virus in wild boar and red deer in The Netherlands. J. Virol. Methods 2010, 168, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Weger, S.; Elkin, B.; Lindsay, R.; Bollinger, T.; Crichton, V.; Andonov, A. Hepatitis E virus seroprevalence in free-ranging deer in Canada. Transbound. Emerg. Dis. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kubankova, M.; Kralik, P.; Lamka, J.; Zakovcik, V.; Dolansky, M.; Vasickova, P. Prevalence of hepatitis E virus in populations of wild animals in comparison with animals bred in game enclosures. Food Environ. Virol. 2015, 7, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Serracca, L.; Battistini, R.; Rossini, I.; Mignone, W.; Peletto, S.; Boin, C.; Pistone, G.; Ercolini, R.; Ercolini, C. Molecular investigation on the presence of hepatitis E virus (HEV) in wild game in north-western Italy. Food Environ. Virol. 2015, 7, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Kukielka, D.; Rodriguez-Prieto, V.; Vicente, J.; Sanchez-Vizcaino, J.M. Constant hepatitis E virus (HEV) circulation in wild boar and red deer in Spain: An increasing concern source of HEV zoonotic transmission. Transbound. Emerg. Dis. 2016, 63, e360–e368. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolo, I.; Ponterio, E.; Angeloni, G.; Morandi, F.; Ostanello, F.; Nicoloso, S.; Ruggeri, F.M. Presence of hepatitis E virus in a RED Deer (Cervus elaphus) population in central Italy. Transbound. Emerg. Dis. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tei, S.; Kitajima, N.; Takahashi, K.; Mishiro, S. Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet 2003, 362, 371–373. [Google Scholar] [CrossRef]
- Tamada, Y.; Yano, K.; Yatsuhashi, H.; Inoue, O.; Mawatari, F.; Ishibashi, H. Consumption of wild boar linked to cases of hepatitis E. J. Hepatol. 2004, 40, 869–870. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Lee, J.M.; Jo, Y.W.; Min, H.J.; Kim, H.J.; Jung, W.T.; Lee, O.J.; Yun, H.; Yoon, Y.S. Genotype-4 hepatitis E in a human after ingesting roe deer meat in South Korea. Clin. Mol. Hepatol. 2013, 19, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Brunet, P.; Lano, G.; Moal, V. Hepatitis E virus genotype 4 in Southeastern France: Still around. Liver Int. 2016, 36, 765–767. [Google Scholar] [CrossRef] [PubMed]
- Bouamra, Y.; Gerolami, R.; Arzouni, J.P.; Grimaud, J.C.; Lafforgue, P.; Nelli, M.; Tivoli, N.; Ferretti, A.; Motte, A.; Colson, P. Emergence of autochthonous infections with hepatitis E virus of genotype 4 in Europe. Intervirology 2014, 57, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Meng, X.J.; Doceul, V. Zoonotic origin of hepatitis E. Curr. Opin. Virol. 2015, 10, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Baylis, S.A.; Gartner, T.; Nick, S.; Ovemyr, J.; Blumel, J. Occurrence of hepatitis E virus RNA in plasma donations from Sweden, Germany and the United States. Vox Sang. 2012, 103, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Maunula, L.; Kaupke, A.; Vasickova, P.; Soderberg, K.; Kozyra, I.; Lazic, S.; van der Poel, W.H.; Bouwknegt, M.; Rutjes, S.; Willems, K.A.; et al. Tracing enteric viruses in the European berry fruit supply chain. Int. J. Food Microbiol. 2013, 167, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Bouwknegt, M.; Rutjes, S.A.; Reusken, C.B.; Stockhofe-Zurwieden, N.; Frankena, K.; de Jong, M.C.; de Roda Husman, A.M.; Poel, W.H. The course of hepatitis E virus infection in pigs after contact-infection and intravenous inoculation. BMC Vet. Res. 2009, 5, 7. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Primer/Probe Designation | Primer Sequence from 5′ to 3′ | Position | Reference |
|---|---|---|---|
| HEV genotype 1–4 | |||
| HEVF | CGGTGGTTTCTGGGGTGAC | 5292–5310 1 | This study |
| HEVR | GAAGGGGTTGGTTGGATGAA | 5364–5345 1 | This study |
| HEVP | CGGGTTGATTCTCAGCCCTTCGC FAM | 5311–5333 1 | This study |
| Moose-HEV | |||
| HEVF8 | AGGTGGTGGTTGGGGCCCT | 5037–5055 2 | [34] |
| HEVR8 | TGGCGAATGGGTTTGAGGGG | 5113–5094 2 | [34] |
| HEVP8 | CGCCTCGACTCGCAGCCATTTGC FAM | 5056–5078 2 | [34] |
| Animal Species | No. Animals | No. of Anti-HEV Positive Sera/Number Tested (%) | No. of HEV RNA Positive Serum Samples/Number Tested (%) | No. of HEV RNA Positive Stool Samples/Number Tested (%) | No. of Animals with Any HEV Marker (%) |
|---|---|---|---|---|---|
| Wild boar Sus scrofa | 139 | 11/134 (8.2) | 7/134 (5.2) | 5/130 (3.8) | 21 1 (16) |
| Moose Alces alces | 69 | 9/66 (14) | 10 2/66 (15) | 7 2/63 (11) | 19 3 (28) |
| Roe deer Capreolus capreolus | 30 | 2/29 (7) | 0/29 | 0/27 | 2 |
| Red deer Cervus elaphus | 15 | 1/14 | 0/14 | 0/13 | 1 |
| Fallow deer Dama dama | 7 | 0/7 | 0/7 | 0/7 | 0 |
| TOTAL | 260 | 23/250 (9.2) | 17/250 (6.8) | 12/240 (5) | 43/260 (17) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roth, A.; Lin, J.; Magnius, L.; Karlsson, M.; Belák, S.; Widén, F.; Norder, H. Markers for Ongoing or Previous Hepatitis E Virus Infection Are as Common in Wild Ungulates as in Humans in Sweden. Viruses 2016, 8, 259. https://doi.org/10.3390/v8090259
Roth A, Lin J, Magnius L, Karlsson M, Belák S, Widén F, Norder H. Markers for Ongoing or Previous Hepatitis E Virus Infection Are as Common in Wild Ungulates as in Humans in Sweden. Viruses. 2016; 8(9):259. https://doi.org/10.3390/v8090259
Chicago/Turabian StyleRoth, Anette, Jay Lin, Lars Magnius, Marie Karlsson, Sándór Belák, Frederik Widén, and Heléne Norder. 2016. "Markers for Ongoing or Previous Hepatitis E Virus Infection Are as Common in Wild Ungulates as in Humans in Sweden" Viruses 8, no. 9: 259. https://doi.org/10.3390/v8090259