PF2401-SF, Standardized Fraction of Salvia miltiorrhiza, Induces Apoptosis of Activated Hepatic Stellate Cells in Vitro and in Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
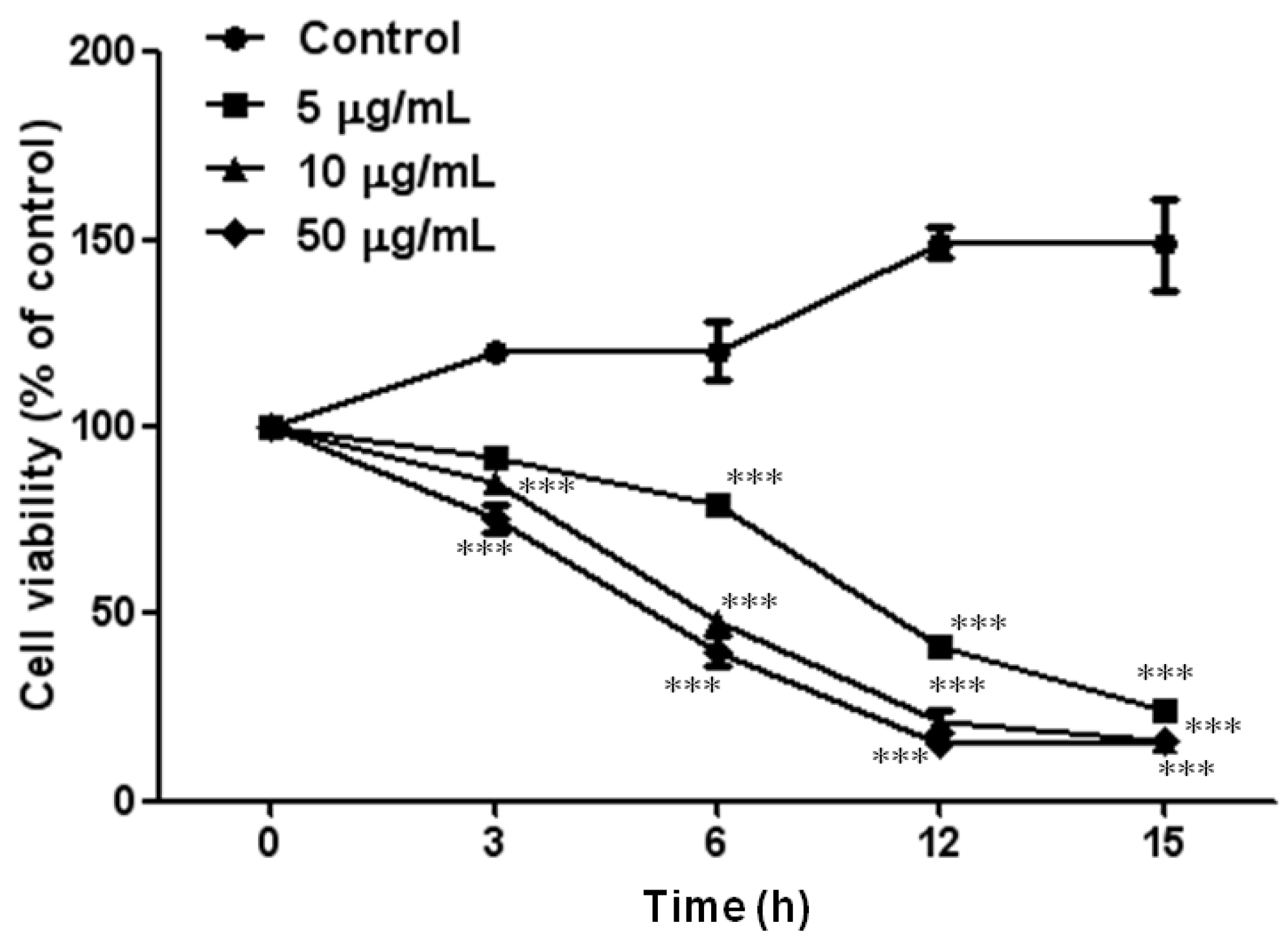
2.1. Effect of PF2401-SF on Cell Viability
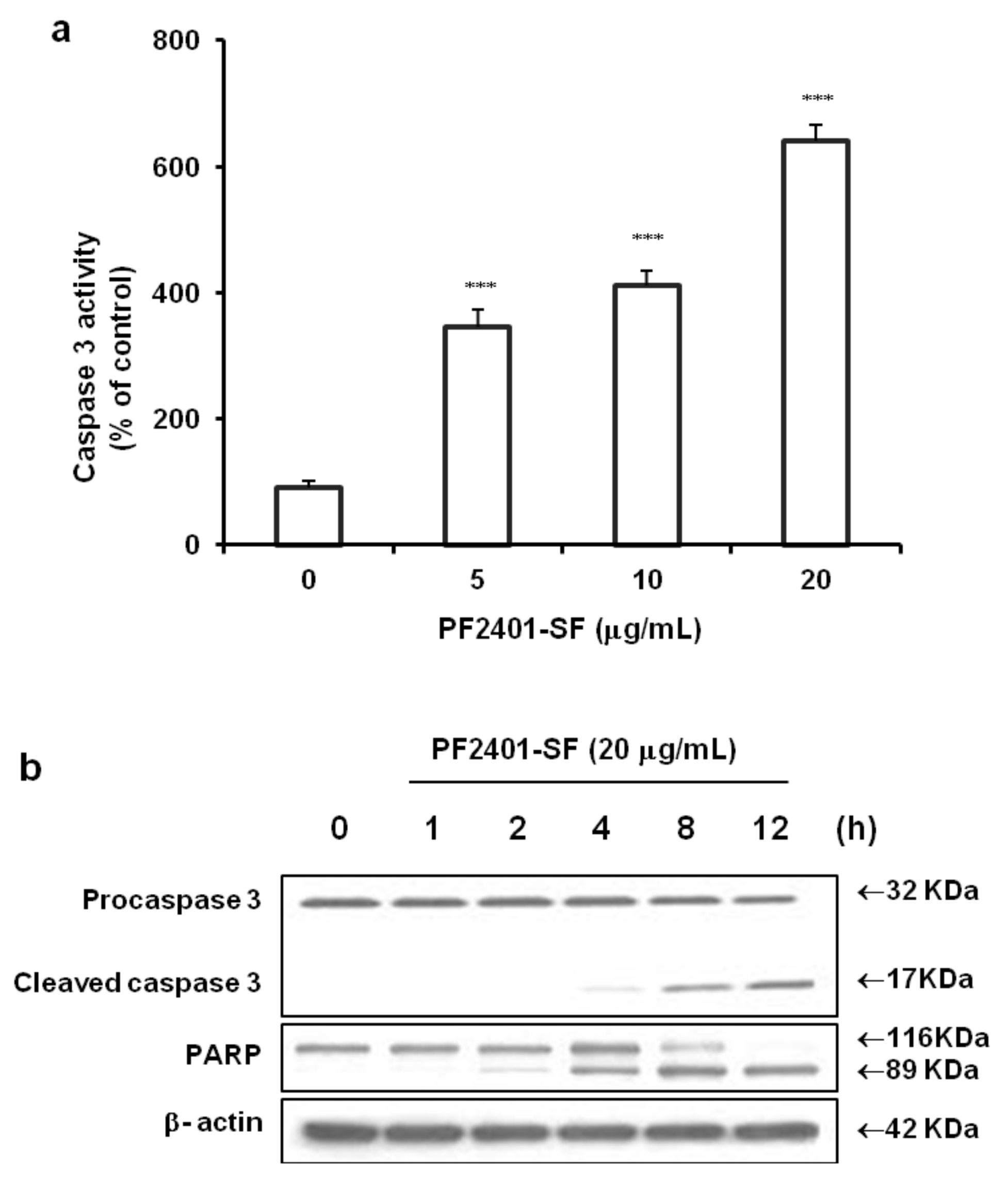
2.2. Effect of PF2401-SF on Apoptosis Induction in Vitro
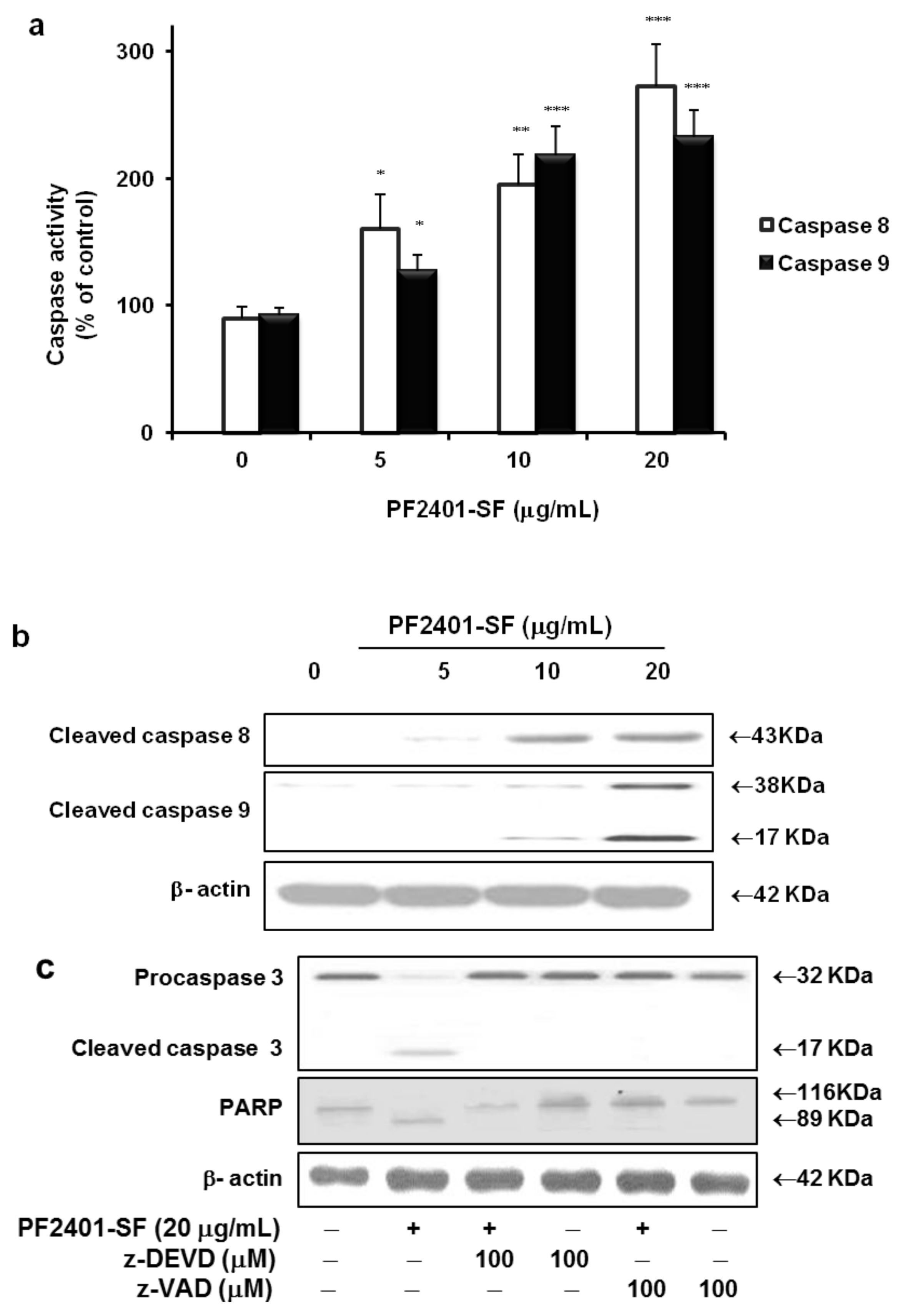
2.3. Effect of PF2401-SF on Pro-Apoptotic and Anti-Apoptotic Proteins
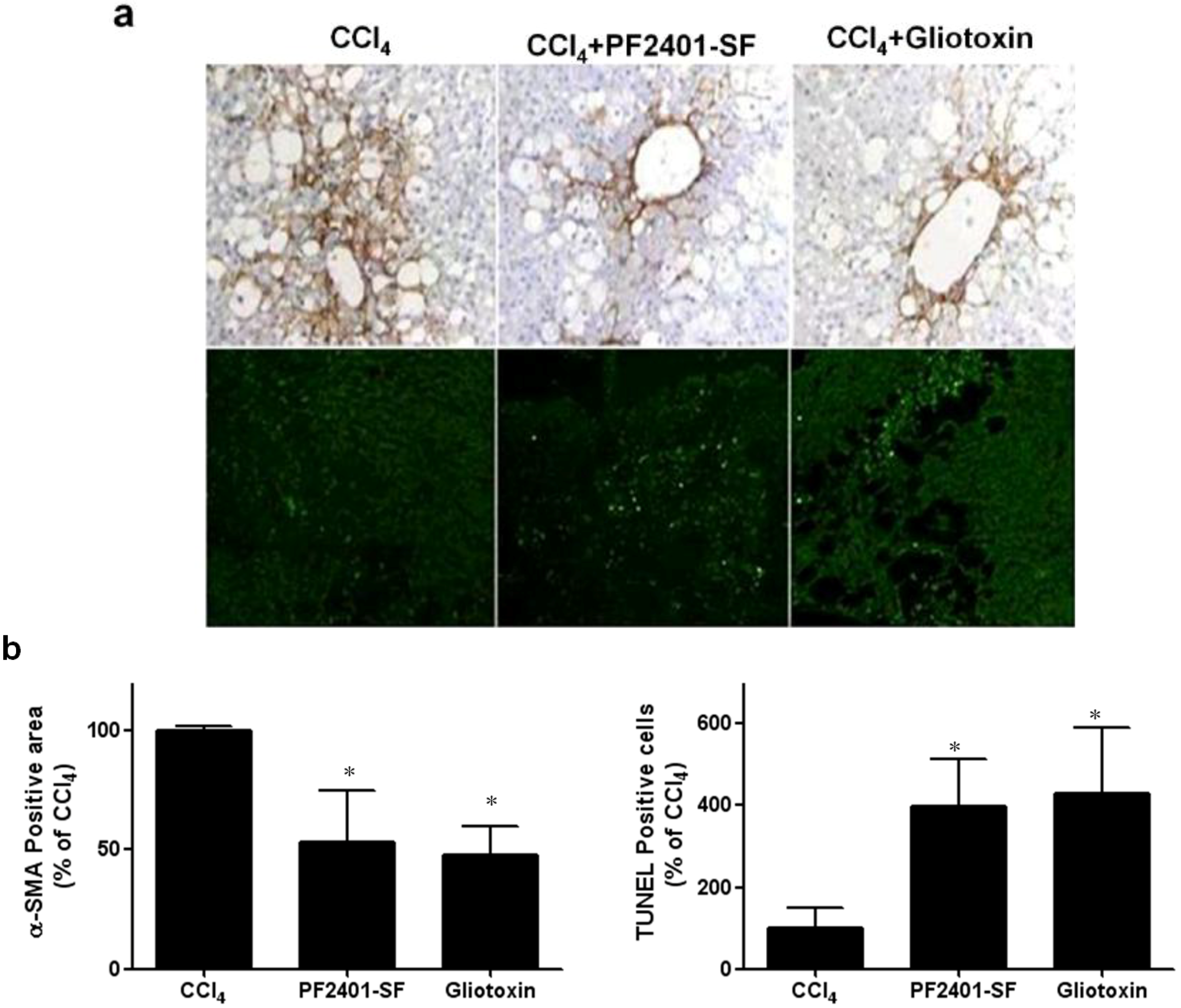
2.4. Effect of PF2401-SF on Apoptosis Induction in Vivo
3. Discussion
4. Experimental
4.1. Preparation of PF2401-SF
4.2. Chemical Reagents
4.3. Cell Culture
4.4. Cell Viability Measurement
4.5. Caspase 3, 8 and 9 Colorimetric Assay
4.6. Western Blot Analysis
4.7. Animals
4.8. Animal Experiments Design
4.9. Immunohistochemical Staining for α-SMA in Rat Liver
4.10. TUNEL Assay
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Friedman, S.L. Liver fibrosis-from bench to bedside. J. Hepatol. 2003, 1, 38–53. [Google Scholar] [CrossRef]
- Iredale, J.P. Models of liver fibrosis: Exploring the dynamic nature of inflammation and repair in a solid organ. J. Clin. Invest. 2007, 117, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Rosenbloom, J.; Castro, S.V.; Jimenez, S.A. Narrative review: Fibrotic diseases: Cellular and molecular mechanisms and novel therapies. Ann. Intern. Med. 2010, 152, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Trebicka, J.; Hennenberg, M.; Odenthal, M.; Shir, K.; Klein, S.; Granzow, M.; Vogt, A.; Dienes, H.P.; Lammert, F.; Reichen, J.; et al. Atorvastatin attenuates hepatic fibrosis in rats after bile duct ligation via decreased turnover of hepatic stellate cells. J. Hepatol. 2010, 53, 702–712. [Google Scholar] [CrossRef] [PubMed]
- Fallofield, J.A. Therapeutic targets in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Bansel, M.B. Reversal of hepatic fibrosis-fact or fantassy? Hepatology 2006, 43, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Issa, R.; Williams, E.; Trim, N.; Kendall, T.; Arthur, M.J.; Reichen, J.; Benyon, R.C.; Iredale, J.P. Apoptosis of hepatic stellate cells: Involvement in resolution of biliary fibrosis and regulation by soluble growth factors. Gut 2001, 48, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ikejima, K.; Kon, K.; Arai, K.; Aoyama, T.; Okumura, K.; Abe, W.; Sato, N.; Watanabe, S. Ursolic acid ameliorates hepatic fibrosis in the rat by specific induction of apoptosis in hepatic stellate cells. J. Hepatol. 2011, 55, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, G.; Serra, S.; Vacca, G.; Lobina, C.; Morazzoni, P.; Bombardelli, E.; Colombo, G.; Gessa, G.L.; Carai, M.A. IDN 5082, a standardized extract of salvia miltiorrhiza, delays acquisition of alcohol drinking behavior in rats. J. Ethnopharmacol. 2003, 85, 93–97. [Google Scholar] [CrossRef]
- Zhou, L.; Zhu, Z.; Chow, M.S. Danshen: An overview of its chemistry, pharmacology, pharmacokineics and clinical use. J. Clin. Pharmacol. 2005, 45, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.; Ho, J.M.; Ang, H.K.; Tan, C.E. Salvia miltiorrhiza reduces experimental-induced hepatic fibrosis in rats. J. Hepatol. 1998, 29, 760–771. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Lin, Y.L.; Chiu, Y.T.; Shiao, M.S.; Lee, C.Y.; Huang, Y.T. Antifibrotic effects of Salvia miltiorrhiza on dimethylnitorsamine-intoxicated rats. J. Biomed. Sci. 2005, 12, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.Y.; Chang, H.H.; Wang, G.J.; Chiu, J.H.; Yang, Y.Y.; Lin, H.C. Water-soluble extract of Salvia miltiorrhiza ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. J. Pharm. Pharmacol. 2006, 58, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Hsu, Y.T; Chiu, Y.T.; Huang, Y.T. Antifibrotic effects of herbal combination regimen on hepatic fibrotic rats. Phytother. Res. 2008, 22, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Zhao, Y.Z.; Kim, Y.C.; Sohn, D.H. PF2401-SF, standardized fraction of Salvia miltiorrhiza and its constituents, tanshinon I, tanshinone IIA, and cryptotansinone, protect primary cultured rat hepatocytes from bile acid-induced apoptosis by inhibiting JNK phosphorylation. Food Chem. Toxicol. 2007, 45, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Zhao, Y.Z.; Kim, Y.C.; Sohn, D.H. Preventive effects of apurified extract isolated from Salvia miltiorrhiza enriched with tanshinone I, tanshinone IIA and cryptotanshinone on hepatocyte injury in vitro and in vivo. Food Chem. Toxicol. 2009, 47, 2742–2748. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.L.; Cheng, Y.Y.; Ding, D.; Mei, S.; Xu, J.W.; Yu, J.; Ou-yang, Q.; Deng, L.; Chen, Q.; Li, Q.Q.; et al. C/EBP-a ameliorates CCl4-induced liver fibrosis in mice through promoting apoptosis of hepatic stellate cells with little apoptosis effect on hepatocytes in vitro and in vivo. Apoptosis 2012, 17, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Ikejima, K.; Honda, H.; Hirose, M.; Kitamura, T.; Takei, Y.; Sato, N. Leptin augments inflammatory and profibrogenic response in the murine liver induced by hepatotoxic chemicals. Hepatology 2001, 34, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.C.; Issa, R.; Smart, D.E.; Trim, N.; Murray, G.I.; Primrose, J.N.; Arthur, M.J.; Iredale, J.P. Gliotoxin stimulates the apoptosis of human and rat hepatic stellate cells and enhances the resolution of liver fibrosis in rats. Gastroenterology 2001, 121, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Alshawsh, M.A.; Abdulla, M.A.; Ismail, S.; Amin, Z.A. Hepatoprotective effects of Orthoisphon stamineus extract on thioacetamide-induced liver cirrhosis in rats. J. Evid. Based Complementary Altern. Med. 2011, 2011, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Iredale, J.P. Cirrhosis: New research provides a basis for rational and targeted treatments. BMJ 2003, 327, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Balnus, M.J.; Kinghorn, A.D. Drug discovery from medicinal plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.J.; Chen, J.Y.; Lin, C.F.; Kako, S.T.; Cheng, J.C.; Chen, H.L.; Chen, C.M. Hepatoprotective effects of Yi Guan Jian, an herbal medicine, in rats with dimethylnitrosamine-induced liver fibrosis. J. Ethnopharmacol. 2011, 134, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Henderson, N.C.; Iredale, J.P. Liver fibrosis: Cellular mechanisms of progression and resolution. Clin. Sci. 2007, 112, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Zhao, Y.Z.; Park, E.J.; Che, X.C.; Seo, G.S.; Sohn, D.H. 2'4'6'-Tris(methoxymethoxy) chalcone induces apoptosis by enhancing Fas-ligand in activated hepatic stellate cells. Eur. J. Pharmacol. 2011, 658, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Budihardjo, I.; Oliver, H.; Lutter, M.; Luo, X.; Wang, X.D. Biochemical pathways of caspase activation during apoptosis. Annu. Rev. Cell Dev. Biol. 1999, 15, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Duriez, P.J.; Sha, G.M. Cleavage of poly polymerase (ADP-robose). Biochem. Cell Biol. 1997, 74, 337–349. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Z.; Kwong, S.Q.; Lui, E.L.; Friedman, S.L.; Li, F.R.; Lam, R.W.; Zhang, G.C.; Zhang, H.; Ye, T. Inhibition of PDGF, TGF-β, and Abl signaling and reduction of liver fibrosis by the small molecule Bcr-Abl tyrosine kinase antagonist Nilotinib. J. Hepatol. 2011, 55, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R. Apoptotic pathways: Paper wraps stone blunts scissors. Cell 2000, 102, 1–4. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell death: Critical control points. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef]
- Elsharkawy, A.M.; Oakley, F.; Lin, F.; Packham, G.; Mann, D.A.; Mann, J. The NF-kappa B p50:p50:HDAC-a repressor complex orchestrates transcriptional inhibition of multiple pro-inflammatory genes. J. Hepatol. 2010, 53, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Invest. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Kim, J, Y.; Kim, K.M.; Nan, J.X.; Zhao, Y.Z.; Park, P.H.; Lee, S.J.; Sohn, D.H. Induction of apoptosis by tanshinone I via cytochrome c release in activated hepatic stellae cells. Pharmacol. Toxicol. 2003, 92, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.C.; Nan, J.X.; Park, P.H.; Kim, J.Y.; Lee, S.H.; Woo, S.W.; Zhao, Y.Z.; Park, E.J.; Sohn, D.H. Curcumin inhibits collagen synthesis and hepatic stellate cell activation in vivo and in vitro. J. Pharm. Pharmacol. 2002, 54, 119–126. [Google Scholar] [CrossRef] [PubMed]
Sample availability: Samples of the PF2401-SF are available from the authors. |





© 2013 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Parajuli, D.R.; Park, E.-J.; Che, X.-H.; Jiang, W.-Y.; Kim, Y.-C.; Sohn, D.H.; Lee, S.H. PF2401-SF, Standardized Fraction of Salvia miltiorrhiza, Induces Apoptosis of Activated Hepatic Stellate Cells in Vitro and in Vivo. Molecules 2013, 18, 2122-2134. https://doi.org/10.3390/molecules18022122
Parajuli DR, Park E-J, Che X-H, Jiang W-Y, Kim Y-C, Sohn DH, Lee SH. PF2401-SF, Standardized Fraction of Salvia miltiorrhiza, Induces Apoptosis of Activated Hepatic Stellate Cells in Vitro and in Vivo. Molecules. 2013; 18(2):2122-2134. https://doi.org/10.3390/molecules18022122
Chicago/Turabian StyleParajuli, Daya Ram, Eun-Jeon Park, Xian-Hua Che, Wen-Yi Jiang, Youn-Chul Kim, Dong Hwan Sohn, and Sung Hee Lee. 2013. "PF2401-SF, Standardized Fraction of Salvia miltiorrhiza, Induces Apoptosis of Activated Hepatic Stellate Cells in Vitro and in Vivo" Molecules 18, no. 2: 2122-2134. https://doi.org/10.3390/molecules18022122