Development of a Sphingosylphosphorylcholine Detection System Using RNA Aptamers

Abstract
:1. Introduction

2. Results and Discussion
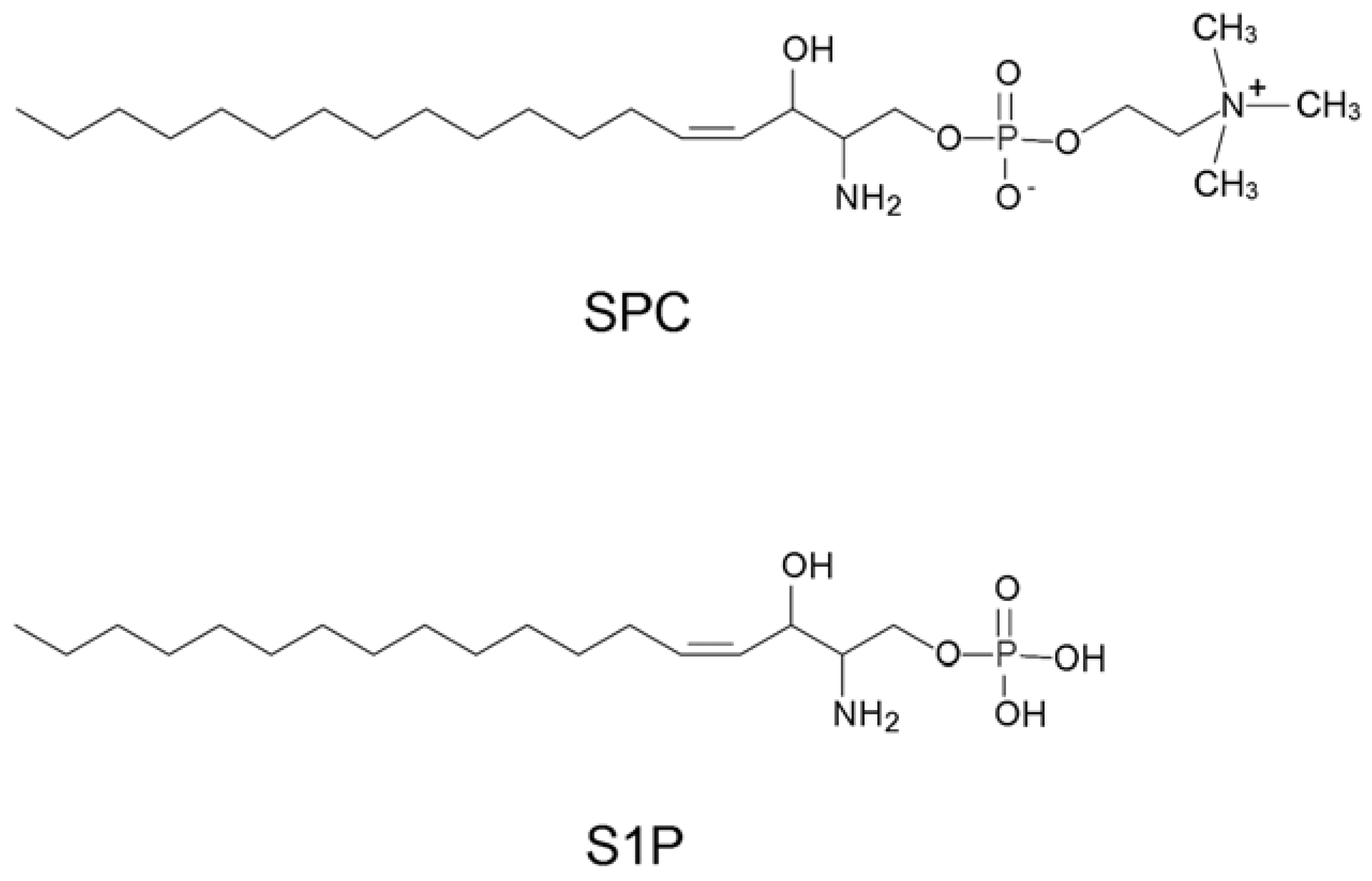
2.1. Selection of SPC binding aptamers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone ID | Frequency | Sequence |
|---|---|---|
| C1 | 12/85 | gggaauggauccacaucuacgaauucUACCGUUAUUGGUGUCACCGAAGAUGUUAuucacugcagacuugacgaagcuu (79 mer) |
| C2 | 7/85 | gggaauggauccacaucuacgaauucUUCCGUUAUUGGAGCCAAGUCGUAUCCCGAuucacugcagacuugacgaagcuu (80 mer) |
| C3 | 4/85 | gggaauggauccacaucuacgaauucUACCGUUAUUGGAGCACGCGUAGUAUGGUUuucacugcagacuugacgaagcuu (80 mer) |
| C4 | 3/85 | gggaauggauccacaucuacgaauucUACCGUUAUUGGUGUAACGUAAUUGUGAuucacugcagacuugacgaagcuu (78 mer) |
| C5 | 2/85 | gggaauggauccacaucuacgaauucUACCAUUAUUGGAGCUGUCGAUUUGCUGGAuucacugcagacuugacgaagcuu (80 mer) |
| C6 | 2/85 | gggaauggauccacaucuacgaauucUCCCGUUAUUGGAGUCACGCGUAGUCCUCCuucacugcagacuugacgaagcuu (80 mer) |
| C7 | 2/85 | gggaauggauccacaucuacgaauucUACCGUUGUUGGAGACGCUUAGAUGUCCGAuucacugcagacuugacgaagcuu (80 mer) |
| C8 | 2/85 | gggaauggauccacaucuacgaauucGCAUUGUCCGCACGCAAAGCAUUAUUGUGAuucacugcagacuugacgaagcuu (80 mer) |
| C9 | 2/85 | gggaauggauccacaucuacgaauucGUCCGUUAUUGGCGCCAGCGUACAUGCGGGuucacugcagacuugacgaagcuu (80 mer) |
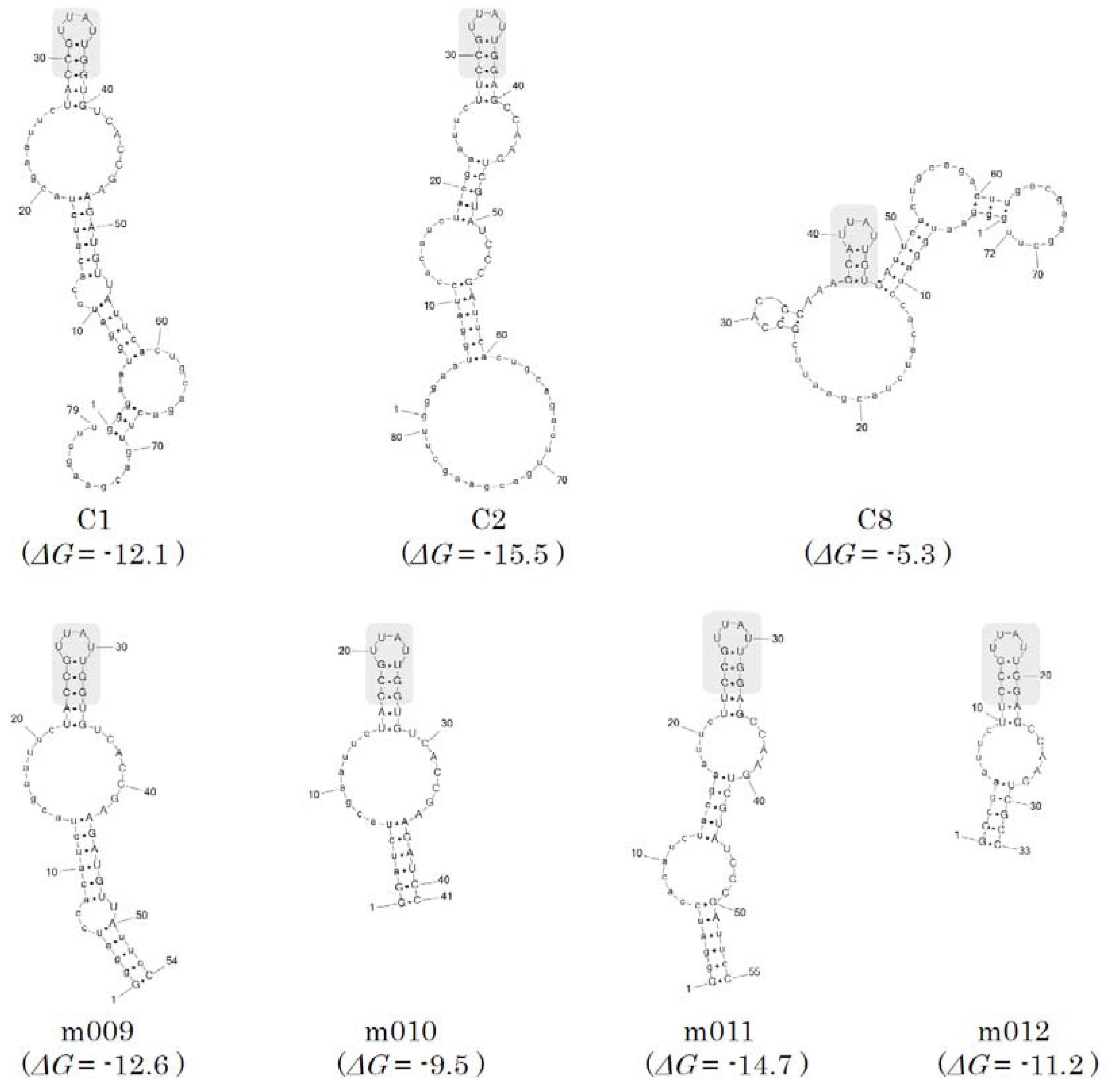
| m009 | GggauccacaucuacgaauucUACCGUUAUUGGUGUCACCGAAGAUGUUAuucC (54 mer) | |
| m010 | GGaucuacgaauucUACCGUUAUUGGUGUCACCGAAGAUCC (41 mer) | |
| m011 | GggauccacaucuacgaauucUUCCGUUAUUGGAGCCAAGUCGUAUCCCGAuucC (55 mer) | |
| m012 | GGcgaauucUUCCGUUAUUGGAGCCAAGUCGCC (33 mer) |
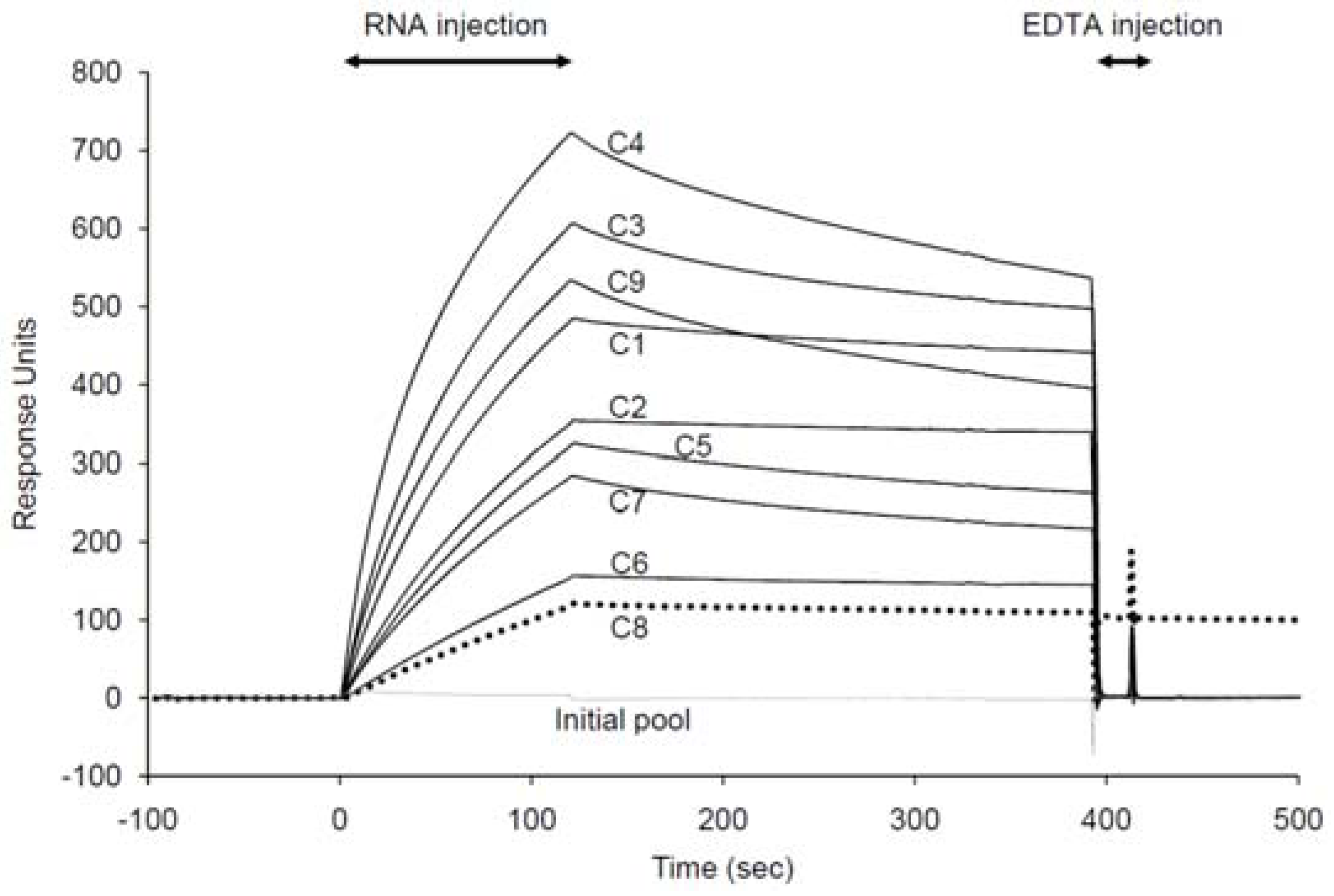
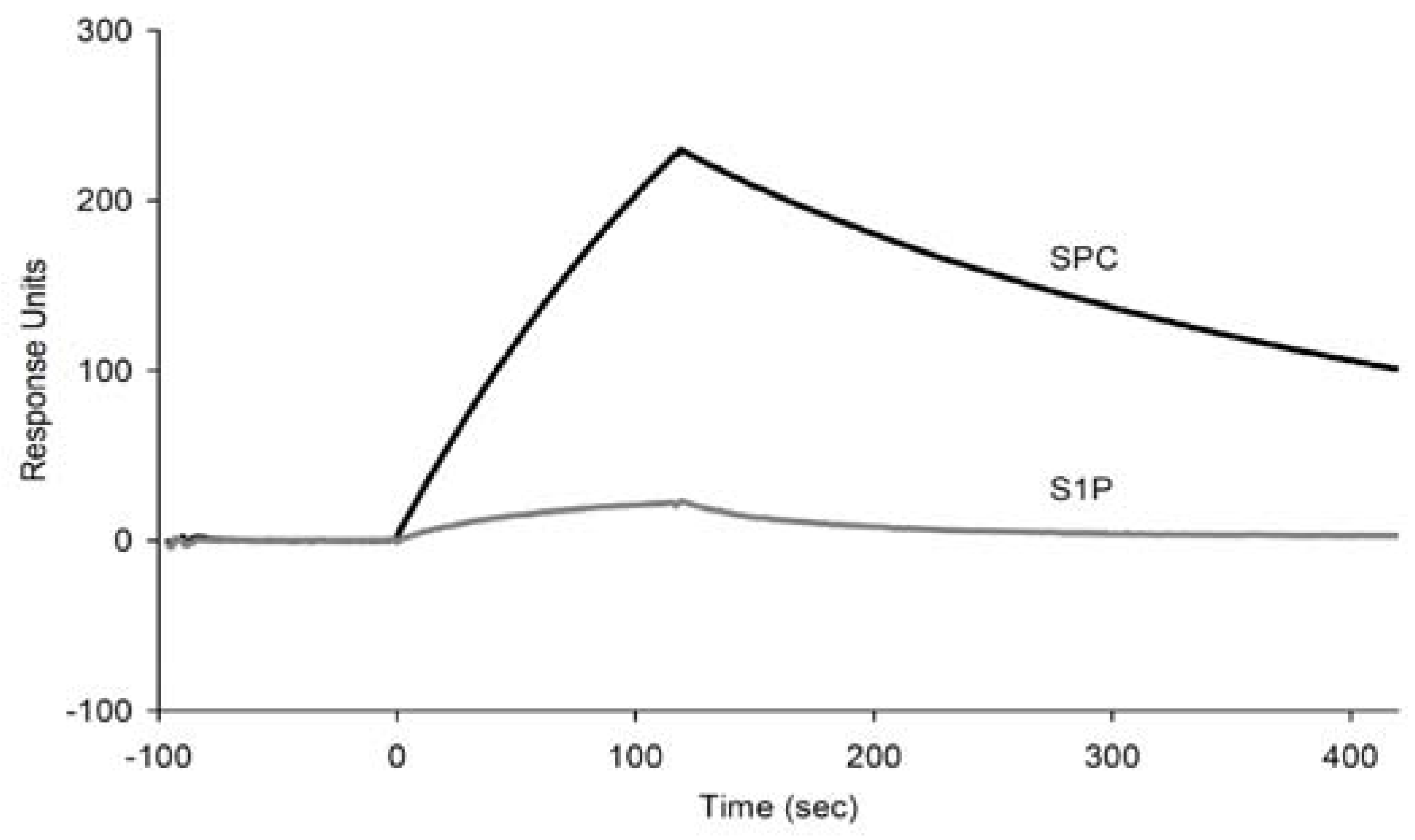
2.2. Binding assay of the selected aptamer candidates using SPR

| Aptamer | ka (104/Ms) | kd (10-4/s) | KD (nM) |
|---|---|---|---|
| C1 | 2.04 ± 0.01 | 4.27 ± 0.03 | 20.9 ± 0.19 |
| C2 | 1.41 ± 0.01 | 4.67 ± 0.01 | 33.2 ± 0.19 |
| C8 | 0.59 ± 0.02 | 5.36 ± 0.02 | 90.4 ± 2.35 |
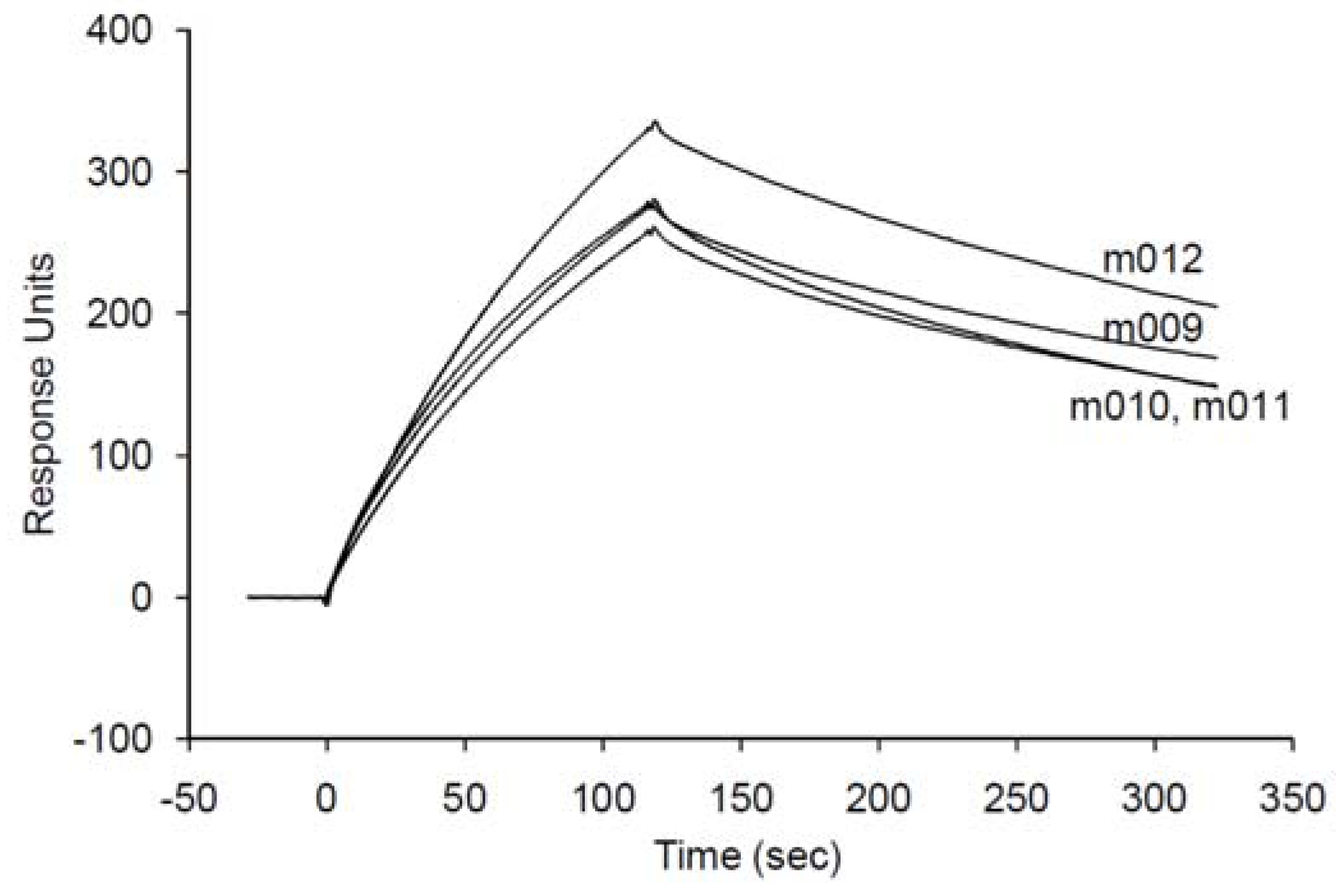
| m009 | 1.24 ± 0.33 | 23.1 ± 0.08 | 186 ± 5.05 |
| m010 | 1.17 ± 0.33 | 29.5 ± 0.09 | 252 ± 7.04 |
| m011 | 1.13 ± 0.25 | 24.9 ± 0.06 | 221 ± 4.84 |
| m012 | 1.11 ± 0.17 | 21.8 ± 0.04 | 196 ± 2.98 |
2.3. Secondary structure prediction and minimization of the SPC aptamers


2.4. Validation of an SPC assay system based on the minimized aptamer


3. Experimental
3.1. Reagents
3.2. SELEX
| Round | RNA (pmol) | Resins (µL) | Incubation time (min) | Total wash volume (mL) | Pre-selection resins (uL) |
|---|---|---|---|---|---|
| 1 | 750 | SA 150 | 180 | 0.9 | 0 |
| 2 | 225 | NA 150 | 30 | 1.5 | 0 |
| 3 | 112.5 | SA 150 | 30 | 1.5 | 150 |
| 4 | 45 | NA 150 | 30 | 1.5 | 150 |
| 5 | 15 | SA 75 | 30 | 1.125 | 112.5 |
| 6 | 7.5 | NA 50 | 30 | 1 | 100 |
| 7 | 1.25 | SA 25 | 30 | 0.625 | 50 |
| 8 | 0.625 | NA 25 | 30 | 0.625 | 50 |
| 9 | 0.625 | SA 25 | 30 | 0.625 | 100 |
| 10 | 0.5 | NA 25 | 30 | 0.75 | 100 |
| 11 | 0.375 | SA 25 | 30 | 0.9 | 100 |
| 12 | 0.25 | NA 25 | 30 | 0.9 | 100 |
3.3. SPR spectroscopy
3.4. ELAA
4. Conclusions
Acknowledgements
- Sample Availability: Samples of the compounds are available from the authors.
References
- Higuchi, K.; Hara, J.; Okamoto, R.; Kawashima, M.; Imokawa, G. The skin of atopic dermatitis patients contains a novel enzyme, glucosylceramide sphingomyelin deacylase, which cleaves the N-acyl linkage of sphingomyelin and glucosylceramide. Biochem. J. 2000, 350, 747–756. [Google Scholar] [CrossRef]
- Spiegel, S.; Milstien, S. Functions of the multifaceted family of sphingosine kinases and some close relatives. J. Biol. Chem. 2007, 282, 2125–2129. [Google Scholar] [CrossRef]
- Nixon, G.F.; Mathieson, F.A.; Hunter, I. The multi-functional role of sphingosylphosphorylcholine. Prog. Lipid Res. 2008, 47, 62–75. [Google Scholar] [CrossRef]
- Meyer zu Heringdorf, D.; Himmel, H.M.; Jakobs, K.H. Sphingosylphosphorylcholine-biological functions and mechanisms of action. Biochim. Biophys. Acta 2002, 1582, 178–189. [Google Scholar] [CrossRef]
- Boguslawski, G.; Lyons, D.; Harvey, K.A.; Kovala, A.T.; English, D. Sphingosylphosphorylcholine induces endothelial cell migration and morphogenesis. Biochem. Biophys. Res. Commun. 2000, 272, 603–609. [Google Scholar] [CrossRef]
- Desai, N.N.; Spiegel, S. Sphingosylphosphorylcholine is a remarkably potent mitogen for a variety of cell lines. Biochem. Biophys. Res. Commun. 1991, 181, 361–366. [Google Scholar] [CrossRef]
- Wakita, H.; Matsushita, K.; Nishimura, K.; Tokura, Y.; Furukawa, F.; Takigawa, M. Sphingosylphosphorylcholine stimulates proliferation and upregulates cell surface-associated plasminogen activator activity in cultured human keratinocytes. J. Invest. Dermatol. 1998, 110, 253–258. [Google Scholar] [CrossRef]
- Imokawa, G.; Takagi, Y.; Higuchi, K.; Kondo, H.; Yada, Y. Sphingosylphosphorylcholine is a potent inducer of intercellular adhesion molecule-1 expression in human keratinocytes. J. Invest. Dermatol. 1999, 112, 91–96. [Google Scholar] [CrossRef]
- Mathieson, F.A.; Nixon, G.F. Sphingolipids differentially regulate mitogen-activated protein kinases and intracellular Ca2+ in vascular smooth muscle: effects on CREB activation. Br. J. Pharmacol. 2006, 147, 351–359. [Google Scholar] [CrossRef]
- Okamoto, R.; Arikawa, J.; Ishibashi, M.; Kawashima, M.; Takagi, Y.; Imokawa, G. Sphingosylphosphorylcholine is upregulated in the stratum corneum of patients with atopic dermatitis. J. Lipid Res. 2003, 44, 93–102. [Google Scholar] [CrossRef]
- Kurokawa, T.; Yumiya, Y.; Fujisawa, H.; Shirao, S.; Kashiwagi, S.; Sato, M.; Kishi, H.; Miwa, S.; Mogami, K.; Kato, S.; et al. Elevated concentrations of sphingosylphosphorylcholine in cerebrospinal fluid after subarachnoid hemorrhage: a possible role as a spasmogen. J. Clin. Neurosci. 2009, 16, 1064–1068. [Google Scholar] [CrossRef]
- Liliom, K.; Sun, G.; Bunemann, M.; Virag, T.; Nusser, N.; Baker, D.L.; Wang, D.A.; Fabian, M.J.; Brandts, B.; Bender, K.; et al. Sphingosylphosphocholine is a naturally occurring lipid mediator in blood plasma: a possible role in regulating cardiac function via sphingolipid receptors. Biochem. J. 2001, 355, 189–197. [Google Scholar] [CrossRef]
- Shirao, S.; Kashiwagi, S.; Sato, M.; Miwa, S.; Nakao, F.; Kurokawa, T.; Todoroki-Ikeda, N.; Mogami, K.; Mizukami, Y.; Kuriyama, S.; et al. Sphingosylphosphorylcholine is a novel messenger for Rho-kinase-mediated Ca2+ sensitization in the bovine cerebral artery: unimportant role for protein kinase C. Circ. Res. 2002, 91, 112–119. [Google Scholar] [CrossRef]
- Pluta, R.M. Delayed cerebral vasospasm and nitric oxide: review, new hypothesis, and proposed treatment. Pharmacol. Ther. 2005, 105, 23–56. [Google Scholar] [CrossRef]
- Merrill, A.H. Jr.; Sullards, M.C.; Allegood, J.C.; Kelly, S.; Wang, E. Sphingolipidomics: high-throughput, structure-specific, and quantitative analysis of sphingolipids by liquid chromatography tandem mass spectrometry. Methods 2005, 36, 207–224. [Google Scholar] [CrossRef]
- Scherer, M.; Leuthaeuser-Jaschinski, K.; Ecker, J.; Schmitz, G.; Liebisch, G. A rapid and quantitative LC-MS/MS method to profile sphingolipids. J. Lipid Res. 2010.
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar]
- Mathews, D.H.; Disney, M.D.; Childs, J.L.; Schroeder, S.J.; Zuker, M.; Turner, D.H. Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure. Proc. Natl. Acad. Sci. USA 2004, 101, 7287–7292. [Google Scholar]
- Byun, Y.; Han, K. PseudoViewer3: generating planar drawings of large-scale RNA structures with pseudoknots. Bioinformatics 2009, 25, 1435–1437. [Google Scholar] [CrossRef]
- Khvorova, A.; Kwak, Y.G.; Tamkun, M.; Majerfeld, I.; Yarus, M. RNAs that bind and change the permeability of phospholipid membranes. Proc. Natl. Acad. Sci. USA 1999, 96, 10649–10654. [Google Scholar] [CrossRef]
- Vlassov, A.; Khvorova, A.; Yarus, M. Binding and disruption of phospholipid bilayers by supramolecular RNA complexes. Proc. Natl. Acad. Sci. USA 2001, 98, 7706–7711. [Google Scholar] [CrossRef]
- Betat, H.; Vogel, S.; Struhalla, M.; Forster, H.H.; Famulok, M.; Welzel, P.; Hahn, U. Aptamers that recognize the lipid moiety of the antibiotic moenomycin A. Biol. Chem. 2003, 384, 1497–1500. [Google Scholar]
- Yarus, M. Amino acids as RNA ligands: a direct-RNA-template theory for the code’s origin. J. Mol. Evol. 1998, 47, 109–117. [Google Scholar] [CrossRef]
- Yatomi, Y.; Igarashi, Y.; Yang, L.; Hisano, N.; Qi, R.; Asazuma, N.; Satoh, K.; Ozaki, Y.; Kume, S. Sphingosine 1-phosphate, a bioactive sphingolipid abundantly stored in platelets, is a normal constituent of human plasma and serum. J. Biochem. 1997, 121, 969–973. [Google Scholar] [CrossRef]
- Lato, S.M.; Boles, A.R.; Ellington, A.D. In vitro selection of RNA lectins: using combinatorial chemistry to interpret ribozyme evolution. Chem. Biol. 1995, 2, 291–303. [Google Scholar] [CrossRef]
- Misono, T.S.; Kumar, P.K. Selection of RNA aptamers against human influenza virus hemagglutinin using surface plasmon resonance. Anal. Biochem. 2005, 342, 312–317. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Horii, K.; Omi, K.; Yoshida, Y.; Imai, Y.; Sakai, N.; Oka, A.; Masuda, H.; Furuichi, M.; Tanimoto, T.; Waga, I. Development of a Sphingosylphosphorylcholine Detection System Using RNA Aptamers. Molecules 2010, 15, 5742-5755. https://doi.org/10.3390/molecules15085742
Horii K, Omi K, Yoshida Y, Imai Y, Sakai N, Oka A, Masuda H, Furuichi M, Tanimoto T, Waga I. Development of a Sphingosylphosphorylcholine Detection System Using RNA Aptamers. Molecules. 2010; 15(8):5742-5755. https://doi.org/10.3390/molecules15085742
Chicago/Turabian StyleHorii, Katsunori, Kazuya Omi, Yoshihito Yoshida, Yuka Imai, Nobuya Sakai, Asako Oka, Hiromi Masuda, Makio Furuichi, Tetsuji Tanimoto, and Iwao Waga. 2010. "Development of a Sphingosylphosphorylcholine Detection System Using RNA Aptamers" Molecules 15, no. 8: 5742-5755. https://doi.org/10.3390/molecules15085742