The Influence of Bioactive Oxylipins from Marine Diatoms on Invertebrate Reproduction and Development

Abstract
:
1. Introduction to Diatom Chemical Defence
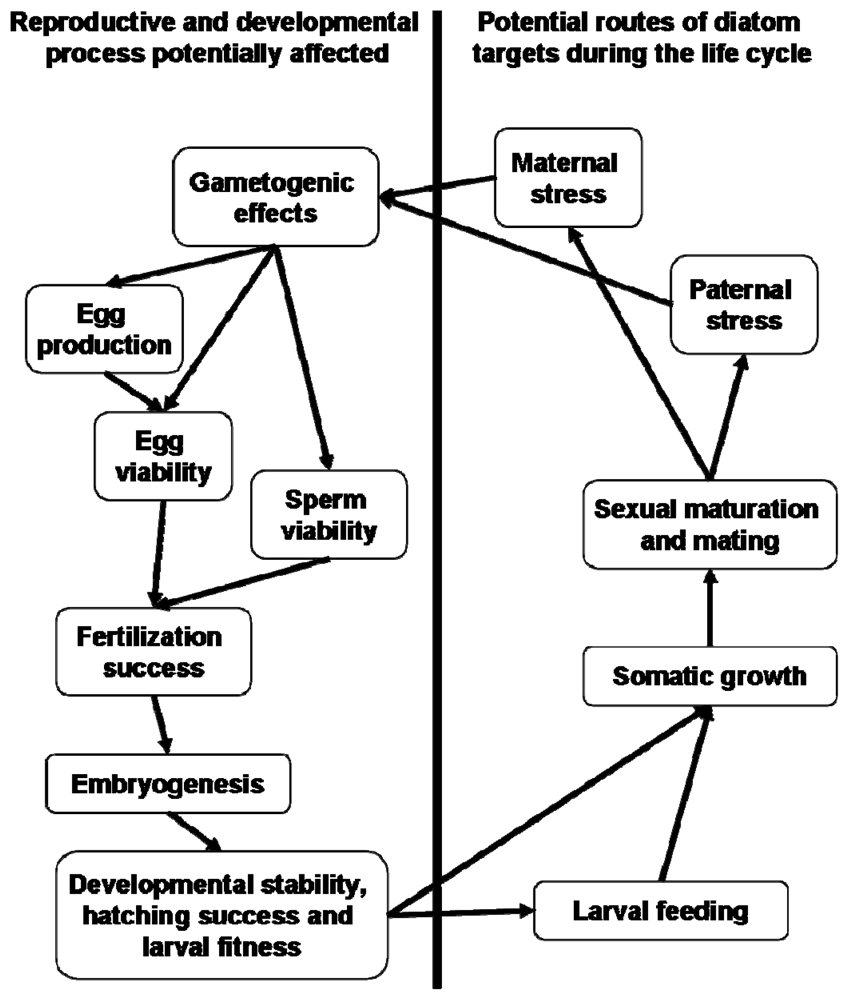
2. Gametogenic Effects
2.1. Oogenesis and Oocyte Maturation
2.2. Spermatogenesis and Sperm Motility
3. Fertilization
4. Embryogenesis
5. Larval Fitness
6. Concluding Remarks
- Samples Availability: Not available from the author.
References
- Legendre, L. The significance of microalgal blooms for fisheries and for the export of particulate organic carbon in oceans. J Plankton Res 1990, 12, 681–699. [Google Scholar]
- Mann, KH. Physical oceanography, food chains, and fish stocks: A review. ICES J Mar Sci 1993, 50, 105–119. [Google Scholar]
- Hamm, CE; Merkel, R; Springer, O; Jurkojc, P; Maier, C; Prechtel, K; Smetacek, V. Architecture and material properties of diatom shells provide effective mechanical protection. Nature 2003, 421, 841–843. [Google Scholar]
- Miralto, A; Barone, G; Romano, G; Poulet, SA; Ianora, A; Russo, GL; Buttino, I; Mazzarella, G; Laabir, M; Cabrini, M; Giacobbe, MG. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar]
- Ianora, A; Miralto, A; Poulet, SA; Carotenuto, Y; Buttino, I; Romano, G; Casotti, R; Pohnert, G; Wichard, T; Colucci-D’Amato, L; Terrazzano, G; Smetacek, V. Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 2004, 429, 103–407. [Google Scholar]
- Ianora, A; Boersma, M; Casotti, R; Fontana, A; Harder, J; Hoffmann, F; Pavia, H; Potin, P; Poulet, SA; Toth, G. New trends in marine chemical ecology. Estuaries Coasts 2006, 9, 531–555. [Google Scholar]
- Jónasdóttir, SH; Ki⊘rboe, T. Copepod recruitment and food composition: Do diatoms affect hatching success? Mar Biol 1996, 125, 743–750. [Google Scholar]
- Jónasdóttir, SH; Ki⊘rboe, T; Tang, KW; StJohn, M; Visser, AW; Saiz, E; Dam, HG. Role of diatoms in copepod production: Good, harmless or toxic? Mar Ecol Prog Ser 1998, 172, 305–308. [Google Scholar]
- Irigoien, X; Harris, RP; Verheye, HM; Joly, P; Runge, J; Starr, M; Pond, D; Campbell, R; Shreeve, R; Ward, P; Smith, AN; Dam, HG; Peterson, W; Tirelli, V; Koski, M; Smith, T; Harbour, D; Davidson, R. Copepod hatching success in marine ecosystems with high diatom concentrations. Nature 2002, 419, 387–389. [Google Scholar]
- Flynn, KJ; Irigoien, X. Aldehyde-induced insidious effects cannot be considered as a diatom defence mechanism against copepods. Mar Ecol Prog Ser 2009, 377, 79–89. [Google Scholar]
- Dutz, J; Koski, M; Jónasdóttir, SH. Copepod reproduction is unaffected by diatom aldehydes or lipid composition. Limnol Oceanogr 2008, 53, 225–235. [Google Scholar]
- Wichard, T; Poulet, SA; Bouleseix, AL; Ledoux, JB; Lebreton, B; Marchetti, J; Pohnert, G. Influence of diatoms on copepod reproduction. II. Uncorrelated effects of diatom-derived α,β,γ,δ-unsaturated aldehydes and polyunsaturated fatty acids on Calanus helgolandicus in the field. Prog Oceanogr 2008, 77, 30–44. [Google Scholar]
- Landsberg, JH. The effects of harmful algal blooms on aquatic organisms. Rev Fish Sci 2002, 10, 113–390. [Google Scholar]
- Nejstgaard, JC; Solberg, PT. Repression of copepod feeding and fecundity by the toxic haptophyte Prymnesium patelliferum. Sarsia 1996, 81, 339–344. [Google Scholar]
- Yan, T; Zhou, MJ; Fu, M; Wang, YF; Yu, RC; Li, J. Inhibition of egg hatching success and larvae survival of the scallop, Chlamys farreri, associated with exposure to cells and cell fragments of the dinoflagellate Alexandrium tamarense. Toxicon 2001, 39, 1239–1244. [Google Scholar]
- Colin, SP; Dam, HG. Testing for toxic effects of prey on zooplankton using sole versus mixed diets. Limnol Oceanogr 2002, 47, 1430–1437. [Google Scholar]
- Sellner, KG; Olson, MM; Olli, K. Copepod interactions with toxic and non-toxic cyanobacteria from the Gulf of Finland. Phycologia 1996, 35, 177–182. [Google Scholar]
- Turner, JT; Tester, PA; Hansen, PJ. Anderson, DM, Cembella, AD, Hallegraeff, GM, Eds.; Interactions between toxic marine phytoplankton and metazoan and protistan grazers. In Physiological Ecology of Harmful Algal Blooms; Springer-Verlag: Berlin, Germany, 1998; pp. 453–474. [Google Scholar]
- Shaw, BA; Andersen, RJ; Harrison, PJ. Feeding deterrent and toxicity effects of apo-fucoxanthinoids and phycotoxins on a marine copepod (Tigriopus californicus). Mar Biol 1997, 128, 273–280. [Google Scholar]
- Lincoln, JA; Turner, JT; Bates, SS; Léger, C; Gauthier, DA. Feeding, egg production, and egg hatching success of the copepods Acartia tonsa and Temora longicornis on diets of the toxic diatom Pseudonitzschia multiseries and the non-toxic diatom Pseudonitzschia pungens. Hydrobiologia 2001, 453, 107–120. [Google Scholar]
- Maneiro, I; Iglesias, P; Guisande, C; Riveiro, I; Barreiro, A; Zeroudaki, S; Granéli, E. Fate of domoic acid ingested by the copepod Acartia clausi. Mar Biol 2005, 148, 123–130. [Google Scholar]
- Pohnert, G. Wound-activated chemical defense in unicellular planktonic algae. Angew Chem, Int Ed 2000, 39, 4352–4354. [Google Scholar]
- Pohnert, G. Phospholipase A2 activity triggers the wound-activated chemical defense in the diatom Thalassiosira rotula. Plant Physiol 2002, 129, 103–111. [Google Scholar]
- d’Ippolito, G; Tucci, S; Cutignano, A; Romano, G; Cimino, G; Miralto, A; Fontana, A. The role of complex lipids in the synthesis of bioactive aldehydes of the marine diatom Skeletonema costatum. Biochim Biophys Acta 2004, 1686, 100–107. [Google Scholar]
- Fontana, A; d’Ippolito, G; Cutignano, A; Miralto, A; Ianora, A; Romano, G; Cimino, G. Chemistry of oxylipin pathways in marine diatoms. Pure Appl Chem 2007a, 79, 475–484. [Google Scholar]
- Ianora, A; Casotti, R; Bastianini, M; Brunet, C; d’Ippolito, G; Fontana, A; Cutignano, A; Turner, JT; Miralto, A. Low reproductive success in copepod communities during a bloom of the diatom Cerataulina pelagica in the North Adriatic Sea. Mar Ecol 2008, 29, 399–410. [Google Scholar]
- Andrianasolo, EH; Haramaty, L; Vardi, A; White, E; Lutz, R; Falkowski, P. Apoptosis-inducing galactolipids from a cultured marine diatom, Phaeodactylum tricornutum. J Nat Prod 2008, 71, 1197–1201. [Google Scholar]
- Fontana, A; d’Ippolito, G; Cutignano, A; Romano, G; Lamari, N; Massa Gallucci, A; Cimino, G; Miralto, A; Ianora, A. A metabolic mechanism for the detrimental effect of marine diatoms on zooplankton grazers. ChemBioChem 2007b, 8, 1810–1818. [Google Scholar]
- Wichard, T; Poulet, SA; Halsband-Lenk, C; Albaina, A; Harris, R; Liu, D; Pohnert, G. Survey of the chemical defence potential of diatoms: Screening of fifty one species for α,β,γ,δ-unsaturated aldehydes. J Chem Ecol 2005, 31, 949–958. [Google Scholar]
- Pohnert, G; Lumineau, O; Cueff, A; Adolph, S; Cordevant, C; Lange, M; Poulet, S. Are volatile unsaturated aldehydes from diatoms the main line of chemical defence against copepods? Mar Ecol Prog Ser 2002, 245, 33–45. [Google Scholar]
- Sarno, D; Kooistra, WH; Medlin, LK; Percopo, I; Zingone, A. Diversity of the genus Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of S. costatum-like species with the description of four new species. J Phycol 2005, 41, 151–176. [Google Scholar]
- d’Ippolito, G; Romano, G; Iadicicco, O; Miralto, A; Ianora, A; Cimino, G; Fontana, A. New birth-control aldehydes from the marine diatom Skeletonema costatum: Characterization and biogenesis. Tetrahedron Lett 2002a, 43, 6133–6136. [Google Scholar]
- d’Ippolito, G; Iadicicco, I; Romano, G; Fontana, A. Detection of short-chain aldehydes in marine organisms: The diatom Thalassiosira rotula. Tetrahedron Lett 2002b, 43, 6137–6140. [Google Scholar]
- d’Ippolito, G; Romano, G; Caruso, T; Spinella, A; Cimino, G; Fontana, A. Production of octadienal in the marine diatom Skeletonema costatum. Org Lett 2003, 5, 885–887. [Google Scholar]
- Collins, R; Bean, G. Volatile constituents obtained from Clamydomonas globosa, the carbonyl fraction. Phycologia 1963, 3, 55–59. [Google Scholar]
- Nuccio, J; Seaton, PJ; Kieber, RJ. Biological production of formaldehyde in the marine environment. Limnol Oceanogr 1995, 40, 521–527. [Google Scholar]
- Wendel, T; Jüttner, F. Lipoxygenase-mediated formation of hydrocarbons and unsaturated aldehydes in freshwater diatoms. Phytochemistry 1996, 41, 1445–1449. [Google Scholar]
- Jüttner, F; Dürst, U. High lipoxygenase activities in epilithic biofilms of diatoms. Arch für Hydrobiol 1997, 138, 451–463. [Google Scholar]
- Adolph, S; Poulet, SA; Pohnert, G. Synthesis and biological activity of α,β,γ,δ-unsaturated aldehydes from diatoms. Tetrahedron 2003, 59, 3003–3008. [Google Scholar]
- van Iersel, ML; Ploemen, JH; Lo Bello, M; Federici, G; van Bladeren, PJ. Interactions of α,β-unsaturated aldehydes and ketones with human glutathione S-transferase P1-1. Chem Biol Interact 1997, 108, 67–78. [Google Scholar]
- Refsgaard, HH; Tsai, L; Stadtman, ER. Modifications of proteins by polyunsaturated fatty acid peroxidation products. Proc Natl Acad Sci USA 2000, 97, 611–616. [Google Scholar]
- Zimmerman, BT; Crawford, GD; Dahl, R; Simon, FR; Mapoles, JE. Mechanisms of acetaldehyde-mediated growth inhibition - delayed cell-cycle progression and induction of apoptosis. Alcoholism-Clin Exp Res 1995, 19, 434–440. [Google Scholar]
- Nappez, C; Battu, S; Beneytout, JL. trans,trans-2,4-decadienal: Cytotoxicity and effect on glutathione level in human erythroleukemia (HEL) cells. Cancer Lett 1996, 99, 115–119. [Google Scholar]
- Bisignano, G; Lagana, MG; Trombetta, D; Arena, S; Nostro, A; Uccella, N; Mazzanti, G; Saija, A. In vitro antibacterial activity of some aliphatic aldehydes from Olea europaea L. FEMS Microbiol Lett 2001, 199, 9–13. [Google Scholar]
- Trombetta, D; Saija, A; Bisignano, G; Arena, S; Caruso, S; Mazzanti, G; Uccella, N; Castelli, F. Study on the mechanisms of the antibacterial action of some plant alpha, beta-unsaturated aldehydes. Lett Appl Microbiol 2002, 35, 285–290. [Google Scholar]
- Ribalet, F; Berges, JA; Ianora, A; Casotti, R. Growth inhibition of cultured marine phytoplankton by toxic algal-derived polyunsaturated aldehdyes. Aquat Toxicol 2007, 85, 219–227. [Google Scholar]
- Ribalet, F; Intertaglia, L; Lebaron, P; Casotti, R. Differential effect of three polyunsaturated aldehydes on marine bacterial isolates. Aquat Toxicol 2008, 86, 249–255. [Google Scholar]
- Krokan, H; Grafstrom, RC; Sundqvist, K; Esterbauer, H; Harris, CC. Cytotoxicity, thiol depletion and inhibition of O6-methylguanine-DNA methyltransferase by various aldehydes in cultured human bronchial fibroblasts. Carcinogenesis 1985, 6, 1755–1759. [Google Scholar]
- Wawra, E; Zollner, H; Schaur, RJ; Tillian, HM; Schauenstein, E. The inhibitory effects of 4-hydroxy-nonenal on DNA-polymerases alpha and beta from rat liver and rapidly dividing Yoshida ascites hepatoma. Cell Biochem Funct 1986, 4, 31–36. [Google Scholar]
- Uchida, K. Current status of acrolein as a lipid peroxidation product. Trends Cardiovasc Med 1999, 9, 109–113. [Google Scholar]
- Carvalho, VM; Di Mascio, P; Campos, IP; Douki, T; Cadet, J; Medeiros, MH. Formation of 1,N-6-etheno-2′-deoxyadenosine adducts by trans,trans-2,4-decadienal. Chem Res Toxicol 1998, 11, 1042–1047. [Google Scholar]
- Carvalho, VM; Asahara, F; Di Mascio, P; Campos, IP; Cadet, J; Medeiros, MH. Novel 1,N-6-etheno-2′-deoxyadenosine adducts from lipid peroxidation products. Chem Res Toxicol 2000, 13, 397–405. [Google Scholar]
- Ferre, N; Girona, J; Cabre, M; Joven, J; LaVille, A; Masana, L; Paternain, JL; Camps, J. Hepatic production of apolar aldehydes in rats with carbon tetrachloride-induced cirrhosis. Mol Cell Biochem 1999, 198, 57–60. [Google Scholar]
- Girona, J; LaVille, AE; Heras, M; Olive, S; Masana, L. Oxidized lipoproteins including HDL and their lipid peroxidation products inhibit TNF-alpha secretion by THP-1 human macrophages. Free Radic Biol Med 1997, 23, 658–667. [Google Scholar]
- Thomas, CE; Jackson, RL; Ohlweiler, DF; Ku, G. Multiple lipid oxidation products in low density lipoproteins induce interleukin-1-beta release from human blood mononuclear cells. J Lipid Res 1994, 35, 417–427. [Google Scholar]
- Girona, J; Vallve, JC; Ribalta, J; Heras, M; Olive, S; Masana, L. 2,4-Decadienal downregulates TNF-alpha gene expression in THP-1 human macrophages. Atherosclerosis 2001, 158, 95–101. [Google Scholar]
- Loureiro, AP; di Mascio, P; Gomes, OF; Medeiros, MH. trans,trans-2,4-Decadienal-induced 1,N-2-etheno-2′-deoxyguanosine adduct formation. Chem Res Toxicol 2000, 13, 601–609. [Google Scholar]
- Wu, SC; Yen, GC; Sheu, F. Mutagenicity and identification of mutagenic compounds of fumes obtained from heating peanut oil. J Food Prot 2001, 64, 240–245. [Google Scholar]
- Boevé, JL; Gfeller, H; Schlunegger, UP; Francke, W. The secretion of the ventral glands in Hoplocampa sawfly larvae. Biochem Syst Ecol 1997, 25, 195–201. [Google Scholar]
- Cheng, LY; Roussis, V. Sex attractant in the marine insect Trochopus plumbeus (Heteroptera: Veliidae): A preliminary report. Mar Ecol Prog Ser 1998, 170, 283–286. [Google Scholar]
- Himeno, K; Honda, H. (E,Z)-10,12-hexadecadienals and (E,E)-10,12-hexadecadienals, major components of female sex pheromone of the Cotton Leaf- Roller, Notarcha derogata (Fabricius) (Lepidoptera, Pyralidae). Appl Entomol Zoolog 1992, 27, 507–515. [Google Scholar]
- Hibbard, BE; Jewett, DK; Bjostad, LB. Pentanoic acid attracts Olcella parva (Adams) (Diptera: Chloropidae) in Colorado corn fields. J Kans Entomol Soc 1997, 70, 67–69. [Google Scholar]
- Tanaka, M; Tamura, K; Ide, H. Citral, an inhibitor of retinoic acid synthesis, modifies chick limb development. Dev Biol 1996, 175, 239–247. [Google Scholar]
- Akiyama, M; Sakata, T; Mori, N; Kato, T; Amano, H; Kuwahara, Y. Chemical ecology of astigmatid mites LXXIII. Neryl formate, the alarm pheromone of Rhizoglyphus setosus Manson (Acarina: Acaridae) and the common pheromone component among four Rhizoglyphus mites. Appl Entomol Zoolog 1997, 32, 75–79. [Google Scholar]
- Omura, H; Honda, K; Hayashi, N. Identification of odoriferous compounds from adults of a swallowtail butterfly, Papilio machaon (Lepidoptera: Papilionidae). Z Naturforsch(C) 2001, 56, 1126–1134. [Google Scholar]
- Kodama, K; Matsui, K; Hatanaka, A; Ishihara, M; Kajiwara, T. A female gamete-characteristic (3Z,6Z,9Z)-dodecatrienoic acid from Analipus japonicus. Phytochemistry 1993, 33, 1039–1042. [Google Scholar]
- Lorenzen, K; Anke, T; Sterner, O. 11-hydroxy-4-methyl-2,4,6-dodecatrienoic acid from fermentations of a Mucor species. Phytochemistry 1996, 43, 791–792. [Google Scholar]
- Naccache, PH; Leblanc, Y; Rokach, J; Patrignani, P; Delaclos, BF; Borgeat, P. Calcium mobilization and right-angle light scatter responses to 12-oxoderivatives of arachidonic acid in neutrophils-evidence for the involvement of the leukotriene B4 receptor. Biochim Biophys Acta 1991, 1133, 102–106. [Google Scholar]
- Romano, G; Russo, GL; Buttino, I; Ianora, A; Miralto, A. A marine diatom-derived aldehyde induces apoptosis in copepod and sea urchin embryos. J Exp Biol 2003, 206, 3487–3494. [Google Scholar]
- Tosti, E; Romano, G; Buttino, I; Cuomo, A; Ianora, A; Miralto, A. Bioactive aldehydes from diatoms block the fertilization current in ascidian oocytes. Mol Reprod Dev 2003, 66, 72–80. [Google Scholar]
- Caldwell, GS; Bentley, MG; Olive, PJ. Inhibition of embryonic development and fertilization in broadcast spawning marine invertebrates by water soluble diatom extracts and the diatom toxin 2-trans, 4-trans decadienal. Aquat Toxicol 2002, 60, 123–137. [Google Scholar]
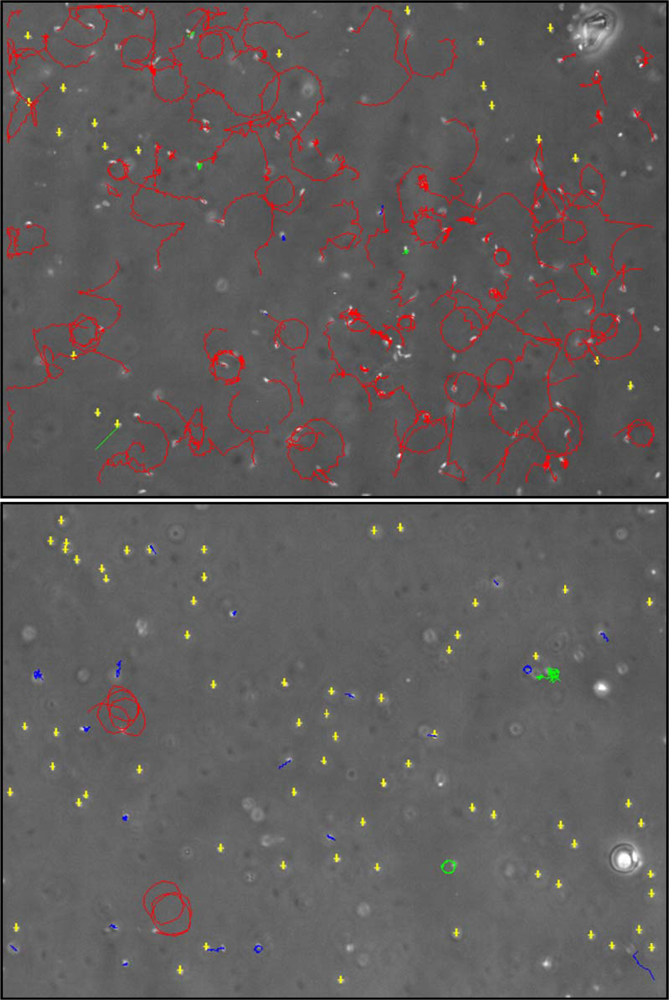
- Caldwell, GS; Bentley, MG; Olive, PJ. First evidence of sperm motility inhibition by the diatom aldehyde 2E,4E-decadienal. Mar Ecol Prog Ser 2004b, 273, 97–108. [Google Scholar]
- Caldwell, GS; Lewis, C; Olive, PJ; Bentley, MG. Exposure to 2,4-decadienal negatively impacts upon marine invertebrate larval fitness. Mar Env Res 2005, 59, 405–417. [Google Scholar]
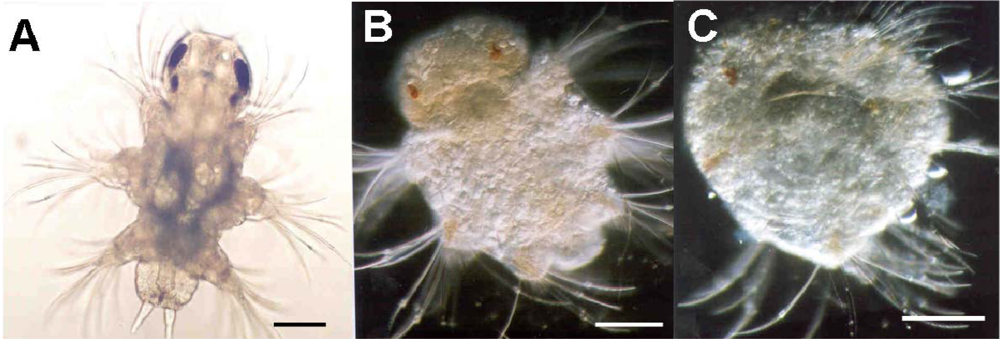
- Lewis, C; Caldwell, GS; Bentley, MG; Olive, PJ. Effects of a bioactive diatom derived aldehyde on developmental stability in Nereis virens (Sars) larvae: An analysis using fluctuating asymmetry. J Exp Mar Biol Ecol 2004, 304, 1–16. [Google Scholar]
- Buttino, I; De Rosa, G; Carotenuto, Y; Mazzela, M; Ianora, A; Esposito, F; Vitiello, V; Quaglia, F; La Rotonda, MI; Miralto, A. Aldehyde-encapsulating liposomes impair marine grazer survivorship. J Exp Biol 2008, 211, 1426–1433. [Google Scholar]
- Ianora, A; Poulet, SA; Miralto, A. The effects of diatoms on copepod reproduction: A review. Phycologia 2003, 42, 351–363. [Google Scholar]
- Cavill, GW; Hinterberger, H. The chemistry of ants IV. Terpenoid constituents of some Dolichoderus and Iridomyrmex species. Aust J Chem 1960, 13, 514–519. [Google Scholar]
- Kubo, I; Lee, Y-W; Pettei, M; Pilkiewicz, F; Nakanishi, K. Potent army worm antifeedants from the east African Warburgia plants. Chem Commun 1976, 1013–1014. [Google Scholar]
- Camazine, SM; Resch, JF; Eisner, T; Meinwald, J. Mushroom chemical defense: Pungent sesquiterpenoid dialdehyde antifeedant to opossum. J Chem Ecol 1983, 9, 1439–1447. [Google Scholar]
- Paul, VJ; Fenical, W. Novel bioactive diterpenoid metabolites from tropical marine algae of the genus Halimeda (Chlorophyta). Tetrahedron 1984, 40, 3053–3062. [Google Scholar]
- Paul, VJ; Fenical, W. Chemical defense in tropical green algae, order Caulerpales. Mar Ecol Prog Ser 1986, 34, 157–169. [Google Scholar]
- Morimoto, M; Tanimoto, K; Sakatani, A; Komai, K. Antifeedant activity of an anthraquinone aldehyde in Galium aparine L. against Spodoptera litura F. Phytochemistry 2002, 60, 163–166. [Google Scholar]
- Halsband-Lenk, C; Pierson, JJ; Leising, AW. Reproduction of Pseudocalanus newmani (Copepoda: Calanoida) is deleteriously affected by diatom blooms–a field study. Prog Oceanogr 2005, 67, 332–348. [Google Scholar]
- Carotenuto, Y; Ianora, A; di Pinto, M; Sarno, D; Miralto, A. Annual cycle of early developmental stage survival and recruitment in the copepods Temora stylifera and Centropages typicus. Mar Ecol Prog Ser 2006, 314, 227–238. [Google Scholar]
- Poulet, SA; Escribano, R; Hidalgo, P; Cueff, A; Wichard, T; Aguilera, V; Vargas, CA; Pohnert, G. Collapse of Calanus chilensis reproduction in a marine environment with high diatom concentration. J Exp Mar Biol Ecol 2007a, 352, 187–199. [Google Scholar]
- d’Ippolito, G; Cutignano, A; Tucci, S; Romano, G; Cimino, G; Fontana, A. Biosynthetic intermediates and stereochemical aspects of aldehyde biosynthesis in the marine diatom Thalassiosira rotula. Phytochemistry, 2006, 67, 314–322. [Google Scholar]
- Paffenhöfer, G-A. An assessment of the effects of diatoms on planktonic copepods. Mar Ecol Prog Ser 2002, 227, 305–310. [Google Scholar]
- Pohnert, G. Diatom-copepod interactions in plankton: The indirect chemical defense of unicellular algae. ChemBioChem 2005, 6, 946–959. [Google Scholar]
- Pohnert, G; Steinke, M; Tollrian, R. Chemical cues, defence metabolites and the shaping of pelagic interspecific interactions. Trends Ecol Evol 2007, 22, 198–204. [Google Scholar]
- Runge, JA. Egg production of the marine planktonic copepod, Calanus pacificus Brodsky: Laboratory observations. J Exp Mar Biol Ecol 1984, 74, 53–66. [Google Scholar]
- Rey-Rassat, C; Irigoien, X; Harris, R; Head, R; Carlotti, F. Egg production rates of Calanus helgolandicus females reared in the laboratory: Variability due to present and past feeding conditions. Mar Ecol Prog Ser 2002, 238, 139–151. [Google Scholar]
- Ban, S; Burns, C; Castel, J; Chaudron, Y; Christou, E; Escribano, R; Umani, SF; Gasparini, S; Ruiz, FG; Hoffmeyer, M; Ianora, A; Kang, HK; Laabir, M; Lacoste, A; Miralto, A; Ning, X; Poulet, S; Rodriguez, V; Runge, J; Shi, J; Starr, M; Uye, S; Wang, Y. The paradox of diatom-copepod interactions. Mar Ecol Prog Ser 1997, 157, 287–293. [Google Scholar]
- Buttino, I; Ianora, A; Carotenuto, Y; Miralto, A. Use of the confocal laser scanning microscope in studies on the developmental biology of marine crustaceans. Microsc Res Techniq 2003, 60, 458–464. [Google Scholar]
- Buttino, I; Do Spirito Santo, S; Ianora, A; Miralto, A. Rapid assessment of copepod embryo viability using fluorescent probes. Mar Biol 2004, 145, 393–399. [Google Scholar]
- Niehoff, B. The gonad morphology and maturation in Arctic Calanus species. J Mar Systems 1998, 15, 53–59. [Google Scholar]
- Niehoff, B. Gonad morphology and oocyte development in Pseudocalanus spp. In relation to spawning activity. Mar Biol 2003, 143, 759–768. [Google Scholar]
- Niehoff, B; Hirche, HJ. Oogenesis and gonad maturation in the copepod Calanus finmarchicus and the prediction of egg production from preserved samples. Polar Biol 1996, 16, 601–612. [Google Scholar]
- Poulet, SA; Cueff, A; Wichard, T; Marchetti, J; Dancie, C; Pohnert, G. Influence of diatoms on copepod reproduction. III. Consequences of abnormal oocyte maturation on reproductive factors in Calanus helgolandicus. Mar Biol 2007b, 152, 415–428. [Google Scholar]
- Laabir, M; Buttino, I; Ianora, A; Kattner, G; Poulet, SA; Romano, G; Carotenuto, Y; Miralto, A. Effect of specific dinoflagellate and diatom diets on gamete ultrastructure and fatty acid profiles of the copepod Temora stylifera. Mar Biol 2001, 138, 1241–1250. [Google Scholar]
- Wichard, T; Gerecht, A; Boersma, M; Poulet, SA; Wiltshire, K; Pohnert, G. Lipid and fatty acid composition of diatoms revisited: Rapid wound-activated change of food quality parameters influences herbivorous copepod reproductive success. ChemBioChem 2007, 8, 1146–1153. [Google Scholar]
- Eckelbarger, KJ; Blades-Eckelbarger, PI. Oogenesis in calanoid copepods. Invert Reprod Dev 2005, 47, 167–181. [Google Scholar]
- Niehoff, B. Life history strategies in zooplankton communities: The significance of female gonad morphology and maturation types for the reproductive biology of marine calanoid copepods. Prog Oceanogr 2007, 74, 1–47. [Google Scholar]
- Caldwell, GS. Diatom mediated disruption of invertebrate reproduction and development; PhD Thesis; University of Newcastle upon Tyne: UK, 2004. [Google Scholar]
- Kanatani, H. Barrington, EJ, Ed.; Hormones in echinoderms. In Hormones and Evolution; Academic Press: New York, NY, USA, 1979; pp. 273–307. [Google Scholar]
- Wylie, AH; Kerr, JF; Currie, AR. Cell death: The significance of apoptosis. Int Rev Cell Mol Biol 1980, 68, 251–306. [Google Scholar]
- Poulet, SA; de Forges, MR; Cueff, A; Lennon, JF. Double-labelling methods used to diagnose apoptotic and necrotic cell degradations in copepod nauplii. Mar Biol 2003, 143, 889–895. [Google Scholar]
- Hansen, E; Even, Y; Genevière, A-M. The α,β,γ,δ-unsaturated aldehyde 2-trans-4-trans-decadienal disturbs DNA replication and mitotic events in early sea urchin embryos. Toxicol Sci 2004a, 81, 190–197. [Google Scholar]
- Chausson, F; Paterson, LA; Betteley, KA; Hannah, L; Meijer, L; Bentley, MG. CDK1/cyclin B regulation during oocyte maturation in two closely related lugworm species, Arenicola marina and Arenicola defodiens. Dev Growth Diff 2004, 46, 71–82. [Google Scholar]
- Meijer, L; Wallace, RW. Hidaka, H, Hartshome, DJ, Eds.; The Role of Calmodulin in Oocyte Maturation. In Calmodulin Antagonists and Cellular Physiology; Academic Press: Orlando, FL, USA, 1985; pp. 129–145. [Google Scholar]
- Orrenius, S; Zhivotovsky, B; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat Rev Mol Cell Biol 2003, 4, 552–565. [Google Scholar]
- Poulet, SA; Ianora, A; Miralto, A; Meijer, L. Do diatoms arrest embryonic development in copepods? Mar Ecol Prog Ser 1994, 111, 79–86. [Google Scholar]
- White, AW. Marine zooplankton can accumulate and retain dinoflagellate toxins and cause fish kills. Limnol Oceanogr 1981, 26, 103–109. [Google Scholar]
- Laabir, M; Poulet, SA; Ianora, A; Miralto, A; Cueff, A. Reproductive response of Calanus helgolandicus. 2. In situ inhibition of embryonic development. Mar Ecol Prog Ser 1995, 129, 97–105. [Google Scholar]
- Boland, W; Jakoby, K; Jaenicke, L; Muller, DG; Folster, E. Receptor specificity and threshold concentration in chemotaxis of the Phaeophyte Cutleria multifida. Z Naturforsch(C) 1981, 36, 262–271. [Google Scholar]
- Boland, W; Marner, FJ; Jaenicke, L; Muller, DG; Folster, E. Comparative receptor study in gamete chemotaxis of the seaweeds Ectocarpus siliculosus and Cutleria multifida - an approach to interspecific communication of algal gametes. Eur J Biochem, 1983, 134, 97–103. [Google Scholar]
- Bender, ML; Brubacher, LJ. Catalysis and Enzyme Action; McGraw Hill Book Company: New York, NY, USA, 1973; p. 161. [Google Scholar]
- Abraham, RT; Acquarone, M; Andersen, A; Aseni, A; Bellé, R; Berger, F; Bergounioux, C; Brunn, G; Buquet-Fagot, C; Fagot, D; Glab, N; Goudeau, H; Goudeau, M; Guerrier, P; Houghton, P; Hendriks, H; Kloareg, B; Lippai, M; Marie, D; Maro, B; Meijer, L; Mester, J; Mulner-Lorillon, O; Poulet, SA; Schierenberg, E; Schutte, B; Vaulot, D; Verlhac, MH. Cellular effects of olomoucine, an inhibitor of cyclin-dependent kinases. Biol Cell 1995, 83, 105–120. [Google Scholar]
- Au, DW; Lee, CY; Chan, KL; Wu, RS. Reproductive impairment of sea urchins upon chronic exposure to cadmium. Part 1: Effects on gamete quality. Environ Pollut 2001, 111, 1–9. [Google Scholar]
- Jha, A; Cote, J; Hoeh, WR; Blier, PU; Stewart, DT. Sperm motility in Mytilus edulis in relation to mitochondrial DNA polymorphisms: Implications for the evolution of doubly uniparental inheritance in bivalves. Evolution 2008, 62, 99–106. [Google Scholar]
- Titelman, J; Varpe, Ø; Eliassen, S; Fiksen, Ø. Copepod mating: Chance or choice? J Plankton Res 2007, 29, 1023–1030. [Google Scholar]
- Ianora, A; Miralto, A; Buttino, I; Romano, G; Poulet, SA. First evidence of some dinoflagellates reducing male copepod fertilization capacity. Limnol Oceanogr 1999, 44, 147–153. [Google Scholar]
- Wicklund, E. The influence of some inhibiting substances on fertilization in the sea urchin egg. Arkiv Zool 1954, 6, 485–501. [Google Scholar]
- Hansen, E; Eilertsen, HC; Ernstsen, A; Genevière, AM. Anti-mitotic activity towards sea urchin embryos in extracts from the marine haptophycean Phaeocystis pouchetii (Hariot) Lagerheim collected along the coast of northern Norway. Toxicon 2003, 41, 803–812. [Google Scholar]
- Hansen, E; Ernstsen, A; Eilertsen, HC. Isolation and characterisation of a cytotoxic polyunsaturated aldehyde from the marine phytoplankter Phaeocystis pouchetii (Hariot) Lagerheim. Toxicology 2004b, 199, 207–217. [Google Scholar]
- Brokaw, CJ; Josslin, R; Bobrow, L. Calcium ion regulation of flagellar beat symmetry in reactivated sea urchin spermatozoa. Biochem Biophys Res Comm 1974, 58, 795–800. [Google Scholar]
- Su, YH; Vacquier, VD. A flagellar K+-dependent Na+/Ca2+ exchanger keeps Ca2+ low in sea urchin spermatozoa. Proc Natl Acad Sci USA 2002, 99, 6743–6748. [Google Scholar]
- Brokaw, CJ. Calcium-induced asymmetrical beating of triton-demembranated sea urchin spermatozoa. J Cell Biol 1979, 62, 401–411. [Google Scholar]
- Gibbons, BH; Gibbons, IR. Calcium-induced quiescence in reactivated sea urchin sperm. J Cell Biol 1980, 84, 13–27. [Google Scholar]
- Fechter, J; Schoneberg, A; Schatten, G. Excision and disassembly of sperm tail microtubules during sea urchin fertilization: Requirements for microtubule dynamics. Cell Motil Cytoskel 1996, 35, 281–288. [Google Scholar]
- Haimov-Kochman, R; Ben-Chetrit, E. The effect of colchicine treatment on sperm production and function: A review. Human Reprod 1998, 13, 360–362. [Google Scholar]
- Granmo, A; Havenhand, J; Magnusson, K; Svane, I. Effects of the planktonic flagellate Chrysochromulina polylepis Manton and Park on fertilization and early development of the ascidian Ciona intestinalis (L) and the blue mussel Mytilus edulis L. J Exp Mar Biol Ecol 1988, 124, 65–71. [Google Scholar]
- Pesando, D; Girard, JP; Durand-Clément, M; Payan, P; Puiseux-Dao, S. Effect of maitotoxin on sea urchin egg fertilisation and on Ca2+ permeabilities of eggs and intracellular stores. Biol Cell 1991, 72, 269–273. [Google Scholar]
- Nomura, K; Nakamura, H; Suzuki, N. False fertilization in sea urchin eggs induced by diabolin, a 120 K kelp protein. Biochem Biophys Res Comm 2000, 272, 691–693. [Google Scholar]
- Poulet, SA; Laabir, M; Ianora, A; Miralto, A. Reproductive response of Calanus helgolandicus. 1. Abnormal embryonic and naupliar development. Mar Ecol Prog Ser 1995, 129, 85–95. [Google Scholar]
- Buttino, I; Miralto, A; Ianora, A; Romano, G; Poulet, SA. Water-soluble extracts of the diatom Thalassiosira rotula induce aberrations in embryonic tubulin organisation of the sea urchin Paracentrotus lividus. Mar Biol 1999, 134, 147–154. [Google Scholar]
- Tilney, LG; Hatano, S; Ishikawa, H; Mooseker, M. The polymerisation of actin: Its role in the generation of the acrosomal process of certain echinoderm sperm. JCell Biol 1973, 59, 109–126. [Google Scholar]
- Schatten, G; Schatten, H. Effects of motility inhibitors during sea urchin fertilisation—microfilament inhibitors prevent sperm incorporation and restructuring of fertilised egg cortex, whereas microtubule inhibitors prevent pronuclear migrations. Exp Cell Res 1981, 135, 311–330. [Google Scholar]
- Bergé, JP; Bourgougnon, N; Carbonnelle, D; LeBert, V; Tomasoni, C; Durand, P; Roussakis, C. Antiproliferative effects of an organic extract from the marine diatom Skeletonema costatum (Grev) Cleve against a non-small-cell bronchopulmonary carcinoma line (NSCLC-N6). Anti-cancer Res 1997, 17, 2115–2120. [Google Scholar]
- Bai, RL; Pei, XF; Boye, O; Getahun, Z; Grover, S; Bekisz, J; Nguyen, NY; Brossi, A; Hamel, E. Identification of cysteine 354 of beta-tubulin as part of the binding site for the A ring of colchicine. J Biol Chem 1996, 271, 12639–12645. [Google Scholar]
- Wilson, JT; Dixon, DR; Dixon, LR. Numerical chromosomal aberrations in the early life-history stages of a marine tubeworm, Pomatoceros lamarckii (Polychaeta: Serpulidae). Aquat Toxicol 2002, 59, 163–175. [Google Scholar]
- Gore, AV; Sampath, K. Localization of transcripts of the zebrafish morphogen squint is dependent on egg activation and the microtubule cytoskeleton. Mech Dev 2002, 112, 153–156. [Google Scholar]
- Ikegami, R; Rivera-Bennetts, AK; Brooker, DL; Yager, TD. Effects of inhibitors of DNA replication on early zebrafish embryos: Evidence for coordinate activation of multiple intrinsic cell-cycle checkpoints at the mid-blastula transition. Zygote 1997, 5, 153–175. [Google Scholar]
- Gabriel, L; Bonelli, G; Dianzani, MU. Inhibition of colchicine binding to rat liver tubulin by aldehydes and by linoleic acid hydroperoxide. Chem Biol Interact 1977, 19, 101–109. [Google Scholar]
- Cetta, CM; Capuzzo, JM. Physiological and biochemical aspects of embryonic and larval development of the winter flounder Pseudopleuronectes americanus. Mar Biol 1982, 71, 327–337. [Google Scholar]
- Kjorsvik, E; Mangor-Jensen, A; Holmefjord, I. Egg quality in fishes. Adv Mar Biol 1990, 26, 71–113. [Google Scholar]
- Pepin, P; Myers, RA. Significance of egg and larval size to recruitment variability of temperate marine fish. Can J Fish Aquat Sci 1991, 48, 1820–1828. [Google Scholar]
- Frangoulos, M; Guisande, C; Maneiro, I; Riveiro, I; Franco, J. Short-term and long-term effects of the toxic dinoflagellate Alexandrium minutum on the copepod Acartia clausi. Mar Ecol Prog Ser 2000, 203, 161–169. [Google Scholar]
- Sellem, F; Pesando, D; Bodennec, G; El Abed, A; Girard, JP. Toxic effects of Gymnodinium cf. mikimotoi unsaturated fatty acids to gametes and embryos of the sea urchin Paracentrotus lividus. Water Res 2000, 34, 550–556. [Google Scholar]
- Gentien, P; Arzul, G; Toularastel, F. Fremy, JM, Ed.; Modes of action of the toxic principle of Gyrodinium cf. aureolum. In Marine Biotoxins; CNEVA Publishers: Paris, France, 1991; pp. 83–86. [Google Scholar]
- Casotti, R; Mazza, S; Brunet, C; Vantrepotte, V; Ianora, A; Miralto, A. Growth inhibition and toxicity of the algal aldehyde 2-trans-2-trans decadienal on Thalassiosira weissflogii (Bacillariophyceae). J Phycol 2005, 41, 7–20. [Google Scholar]
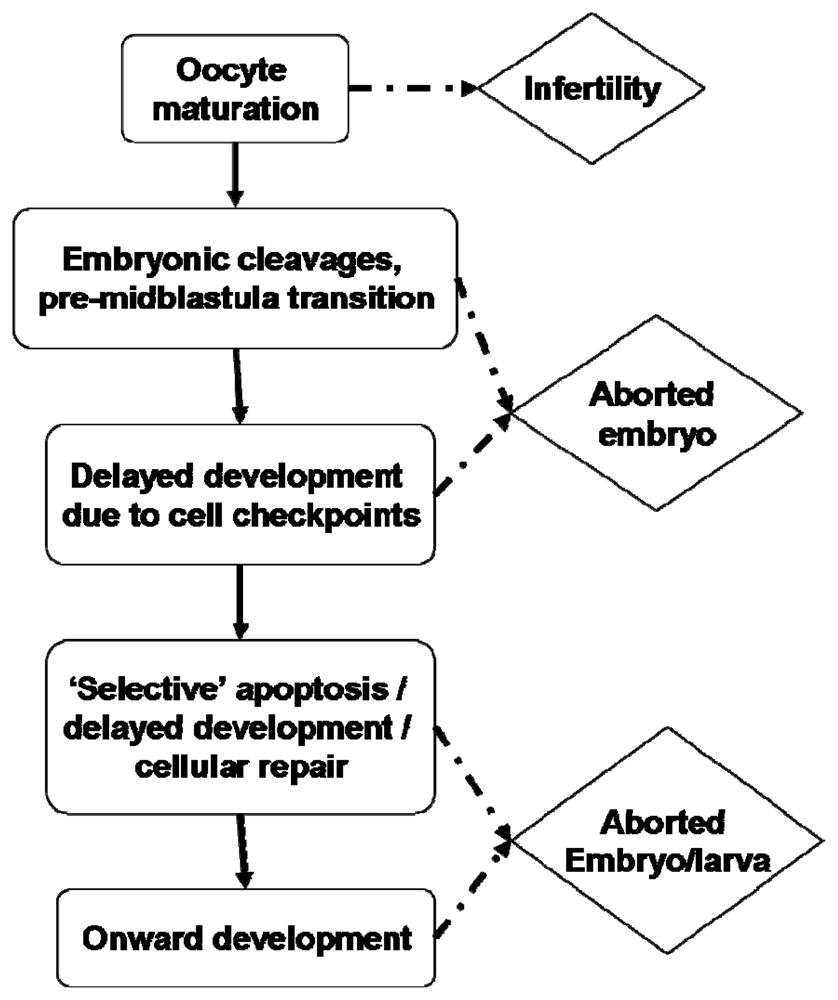
- Hartwell, LH; Weinert, TA. Checkpoints: Controls that ensure the order of cell cycle events. Science 1989, 246, 629–634. [Google Scholar]
- Zhou, BB; Elledge, SJ. The DNA damage response: Putting checkpoints in perspective. Nature 2000, 408, 433–439. [Google Scholar]
- Norbury, CJ; Zhivotovsky, B. DNA damage-indiced apoptosis. Oncogene 2004, 23, 2797–2808. [Google Scholar]
- Epel, D. Protection of DNA during early development: Adaptations and evolutionary consequences. Evol Dev 2003, 5, 83–88. [Google Scholar]
- Marc, J; Belle, R; Morales, J; Comier, P; Mulner-Lorillon, O. Formulated glyphosate activates the DNA-response checkpoint of the cell cycle leading to the prevention of G2/M transition. Toxicol Sci 2004, 82, 436–442. [Google Scholar]
- Le Bouffant, R; Cormier, P; Cueff, A; Bellé, R; Mulner-Lorillon, O. Sea urchin embryo as a model for analysis of the signaling pathways linking DNA damage checkpoint, DNA repair and apoptosis. Cell Mol Life Sci 2007, 64, 1723–1734. [Google Scholar]
- Kane, DA; Kimmel, CB. The zebrafish midblastula transition. Development 1993, 119, 447–456. [Google Scholar]
- Mendelsohn, BA; Gitlin, JD. Coordination of development and metabolism in the pre-midblastula transition zebrafish embryo. Dev Dynamics 2008, 237, 1789–1798. [Google Scholar]
- Vleck, CM; Vleck, D. Metabolism and energetics of avian embryos. J Exp Zool Suppl 1987, 1, 111–125. [Google Scholar]
- Uye, SI. Induction of reproductive failure in the planktonic copepod Calanus pacificus by diatoms. Mar Ecol Prog Ser 1996, 133, 89–97. [Google Scholar]
- Miralto, A; Ianora, A; Guglielmo, L; Zagami, G; Buttino, I. Egg production and hatching success in the peri-Antarctic copepod Calanus simillimus. J Plankton Res 1998, 20, 2369–2378. [Google Scholar]
- Ban, SH; Lee, HW; Shinada, A; Toda, T. In situ egg production and hatching success of the marine copepod Pseudocalanus newmani in Funka Bay and adjacent waters off southwestern Hokkaido, Japan: Associated to diatom bloom. J Plankton Res 2000, 22, 907–922. [Google Scholar]
- Carotenuto, Y; Ianora, A; Buttino, I; Romano, G; Miralto, A. Is postembryonic development in the copepod Temora stylifera negatively affected by diatom diets? J Exp Mar Biol Ecol 2002, 276, 49–66. [Google Scholar]
- Titelman, J; Ki⊘rboe, T. Motility of copepod naulpii and implications for food encounter. Mar Ecol Prog Ser 2003a, 247, 123–135. [Google Scholar]
- Titelman, J; Ki⊘rboe, T. Predator avoidance by naulpii. Mar Ecol Prog Ser 2003b, 247, 137–149. [Google Scholar]
- Williams, R; Wallace, MA. Continuous plankton records: A plankton atlas of the North Atlantic and North Sea: Supplement 1-the genus Clausocalanus (Crustacea: Copepoda, Calanoida) in 1965. Bull Mar Ecol 1975, 8, 167–184. [Google Scholar]
- McEdward, LR. Morphometric and metabolic analysis of the growth and form of an echinopluteus. J Exp Mar Biol Ecol 1984, 82, 259–287. [Google Scholar]
- Palmer, A. Markow, TA, Ed.; Fluctuating asymmetry analyses: A primer. In Developmental Instability: Its Origins and Evolutionary Implications; Kluwer: Dordrecht, Netherlands, 1994; pp. 335–364. [Google Scholar]
- M⊘ller, AP; Swaddle, JP. Asymmetry, Developmental Stability and Evolution; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Pélabon, C; Joly, P. What if anything, does visual asymmetry in fallow deer antlers reveal? Anim Behav 2000, 59, 193–199. [Google Scholar]
- Palmer, AR; Strobeck, C. Fluctuating asymmetry - measurement, analysis, patterns. Ann Rev Ecol Syst 1986, 17, 391–421. [Google Scholar]
- Andersen, HR; Wollenberger, L; Halling-Sorensen, B; Kusk, KO. Development of copepod nauplii to copepodites - a parameter for chronic toxicity including endocrine disruption. Environ Toxicol Chem 2001, 20, 2821–2829. [Google Scholar]
- Campero, M; Ollevier, F; Stoks, R. Ecological relevance and sensitivity depending on the exposure time for two biomarkers. Environ Toxicol 2007, 22, 572–581. [Google Scholar]
- Lewis, C; Pook, C; Galloway, T. Reproductive toxicity of the water accommodated fraction (WAF) of crude oil in the polychaetes Arenicola marina (L.) and Nereis virens (Sars). Aquat Toxicol 2008, 90, 73–81. [Google Scholar]
- Chang, XL; Zhai, BP; Wang, BX; Sun, CH. Effects of the mixture of avermectin and imidacloprid on mortality and developmental stability of Copera annulata (Odonata: Zygoptera) larvae. Biol J Linnean Soc 2009, 96, 44–50. [Google Scholar]
- James, LF. Teratological research at the USDA-ARS poisonous plant research laboratory. J Nat Toxins 1999, 8, 63–80. [Google Scholar]
- Webb, SE; Miller, AL. Calcium signalling during embryonic development. Nat Mol Cell Biol 2003, 4, 539–551. [Google Scholar]
- Wilding, M; Wright, EM; Patel, R; Ellis-Davies, G; Whitaker, MJ. Local perinuclear calcium signals associated with mitosis-entry in early sea urchin embryos. J Cell Biol 1996, 135, 191–200. [Google Scholar]
- Groigno, L; Whitaker, MJ. An anaphase calcium signal controls chromosome disjunction in sea urchin embryos. Cell 1998, 92, 193–204. [Google Scholar]
- Iwasaki, S. Antimitotic agents: Chemistry and recognition of tubulin molecule. Med Res Rev 1993, 13, 183–198. [Google Scholar]
- Borowitzka, MA. Microalgae as sources of pharmaceuticals and other biologically active compounds. J Appl Phycol 1995, 7, 3–15. [Google Scholar]
- Blagosklonny, MV. Teratogens as anti-cancer drugs. Cell Cycle 2005, 4, 1518–1521. [Google Scholar]
- Moore, MN. Biocomplexity: The post-genome challenge in ecotoxicology. Aquat Toxicol 2002, 59, 1–15. [Google Scholar]
- Caldwell, GS; Pickavance, G; Taylor, RL; Bentley, MG. Exposure to copper and a cytotoxic polyunsaturated aldehyde induces reproductive failure in the marine polychaete Nereis virens (Sars). To be submitted for publication.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Caldwell, G.S. The Influence of Bioactive Oxylipins from Marine Diatoms on Invertebrate Reproduction and Development. Mar. Drugs 2009, 7, 367-400. https://doi.org/10.3390/md7030367
Caldwell GS. The Influence of Bioactive Oxylipins from Marine Diatoms on Invertebrate Reproduction and Development. Marine Drugs. 2009; 7(3):367-400. https://doi.org/10.3390/md7030367
Chicago/Turabian StyleCaldwell, Gary S. 2009. "The Influence of Bioactive Oxylipins from Marine Diatoms on Invertebrate Reproduction and Development" Marine Drugs 7, no. 3: 367-400. https://doi.org/10.3390/md7030367