1. Introduction
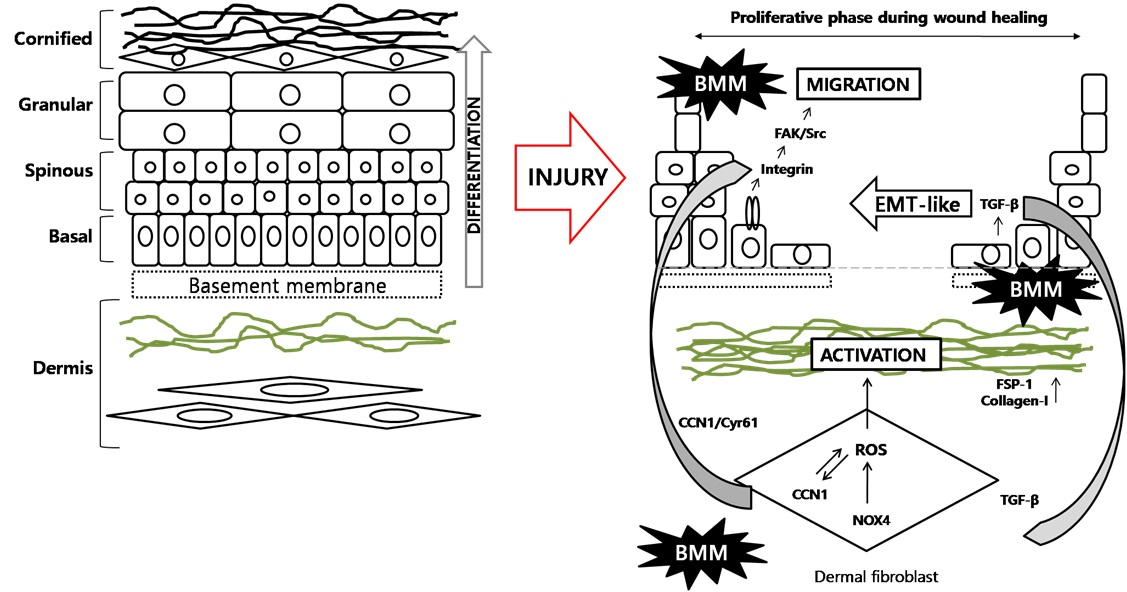
Wound healing is a complex biological process of restoring damaged skin after injury and is composed of three major phases: inflammation, proliferation, and maturation [
1], which are mediated by various cells derived from the epidermis and dermis [
2].
In the proliferation phase, basal layer human keratinocytes (HaCaT) move toward the wound site from the edge [
3]. HaCaT keratinocytes continue to divide and proliferate to cover the wound area and once complete, the cells cease division and begin to differentiate upward, forming spinous, granular, and cornified layers [
4]. This process is known as “re-epithelialization” and is crucial for rapid wound closure. In addition to differentiation, the migration of HaCaT cells also contributes to wound closure through basement layer formation.
Furthermore, dermal fibroblasts (Fbs) crowd around wound sites after injury and form granulation tissue. Myofibroblasts are an activated form of Fbs and are elevated during wound healing [
5]. These cells are able to contract by combining smooth muscle actin (SMA) and myosin, and this contraction contributes to wound repair. Extracellular matrix (ECM) components, such as collagens and gelatin, are synthesized from myofibroblasts. Initially, the ECM is composed of collagen-III, which is a weaker form of collagen that is later replaced by the longer formed-collagen-I, as evidenced in scarring [
6]. In the late phase of wound repair, the myofibroblasts are lost via the apoptosis pathway and expressed collagen-I is degraded by matrix metalloproteinases (MMPs) to balance the composition of the ECM [
7], which is known as the cause of several fibrotic diseases such as cirrhosis, organ fibrosis, or hypertrophic scarring of the skin [
8].
Previously, reactive oxygen species (ROS) have been examined for their involvement in dermal fibroblast activation. Sampson et al. demonstrated that dysregulated redox homeostasis underlies myofibroblast differentiation in the prostatic stroma via NOX4-derived signaling [
9]. NADPH oxidase is one of the ROS generating enzymes and has six types of subunits.
Several studies have reported a relationship between the epithelial–mesenchymal transition (EMT) and embryogenesis, wound healing, or metastasis. Epithelial cell adhesion molecules (ECAM) function to interlink epithelial cells and usually maintain a fixed form. However, when ECAM synthesis was blocked by Twist, Slug, or Snail transcription factors, cells lost their adhesive ability and transformed into spindle and motile mesenchymal cells types. These mechanisms are fundamentally relevant to the acquisition of cells’ migratory ability and are also related to wound healing [
10].
Focal adhesion kinase (FAK) and C-Src form a dual kinase complex and phosphorylate down-stream factors such as the paxillin and p130
Cas. Integrin-regulated activation of FAK and Src functions to increase cell migration, cycle, and survival in normal cells or leads to tumor growth and metastasis in tumor cells [
11].
In this study, we show that BMM, a novel synthetic material, has wound repair effects both in vitro and in vivo. BMM caused HaCaT keratinocytes to have an EMT-like phenotype and migratory effects via the FAK/Src pathway. Dermal fibroblasts were also activated by BMM through the generation of NOX2/4-ROS, and specific cytokines TGF-β or Cyr61 were related to these mechanisms. Specifically, we used a co-culture system that was recently developed in our previous studies to investigate whether BMM stimulates interaction between keratinocytes and dermal fibroblasts [
12]. Our results suggest that BMM accelerates cutaneous wound repair by stimulating re-epithelialization and fibroblast activation.
3. Discussion
Our study presents evidence that BMM treatment can accelerate cutaneous wound healing both in vitro and in vivo. BMM is a novel synthetic compound derived from DK223, which has been previously proven as a potential therapeutic agent for wound repair [
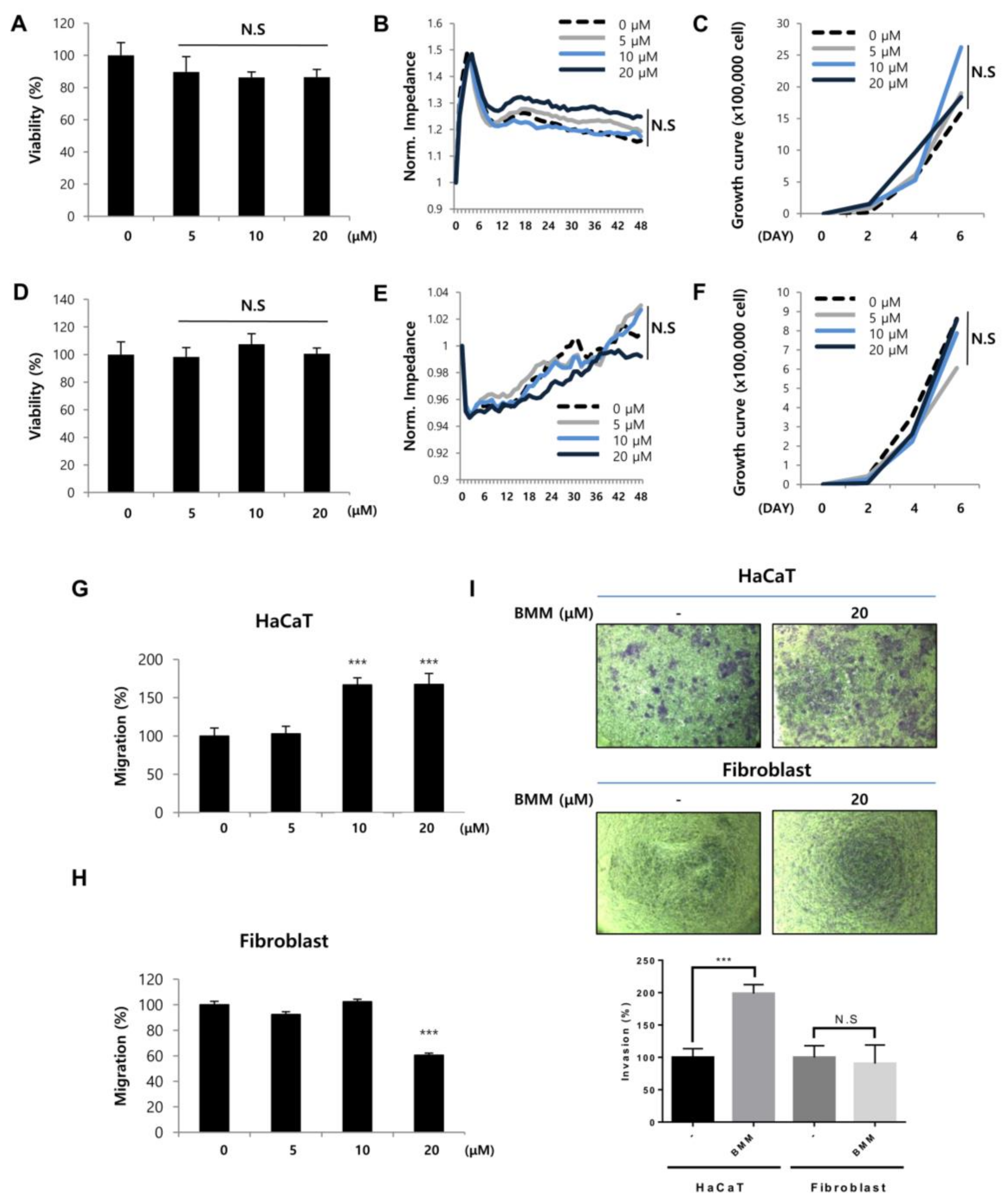
13]. BMM is derived from DK223 by the substitution of a methoxy group to a bromo group. Unlike DK223, BMM only has an effect on HaCaT migration, not proliferation, and Fb proliferation (
Figure 2).
The epidermal layer functions as a barrier of the outside skin and consists of several layers, such as cornified, granular, spinous, and basal layers that are respectively marked with loricrin, keratin 1/2/10, transglutaminase 1/5, and keratin 5/14/15 [
20]. It is the “re-epithelialization” that covers damaged skin with newly divided keratinocytes through migration along the dermis. After the cells meet beneath the scab, the scab is disconnected from the skin and epithelial cells continue to differentiate, approaching the normal thickness of the epidermis [
21]. We have found that this phenomenon occurred in HaCaT cells treated with BMM at an optimum concentration of 20 μM. While BMM has no effect on HaCaT proliferation, migratory ability was increased maximally two-fold. Our results showed that the underlying molecular mechanism to promote HaCaT cell migration results from multiple processes including the EMT, FAK/Src pathway, and MMP secretion. Samy Lamouille et al. have reported that the EMT, a process initiated by TGF-β, is related to wound healing, as well as development, fibrosis, and cancer progression by inducing cell motility [
22]. The epithelial marker, E-cadherin, was significantly decreased and the mesenchymal markers Snail and Slug were dose-dependently increased in HaCaT cells treated with BMM. The relationship between the FAK-Src pathway and wound healing was also identified by Zhao et al. [
15]. They explain that an active FAKSrc complex regulates membrane ruffling and cell migration via phosphorylation of downstream molecules, such as p130
Cas, cdc42, or PI3K, and also plays a critical role in angiogenesis [
15]. It is known that FAK, which is a focal adhesion-related protein kinase, is mediated by integrins. Autophosphorylation of FAK at tyrosine297 occurs by integrin activation and creates a binding site for the SH2 (Src-homology2) domain of Src, which in turn phosphorylates other tyrosine sites in FAK. The active FAK-Src complex promotes Rac1 and finally stimulates lamellipodia formation. As RhoA is reversibly suppressed by an active FAK-Src complex, the formation of stress fibers decreased. Integrin-mediated FAK-Src activation increases membrane protrusion and decreases contractility, thereby regulating cell migration. We confirmed that BMM stimulates the phosphorylation of FAK at Y397 and Src at Y416 (
Figure 4) and speculate that it is a key mechanism for cell mobility. The effect of BMM on MMP expression and secretion was different depending on subunits. The expression of MMP-1, well known as collagenase, was unchanged in spite of BMM treatment. However, the expression of MMP-2 and MMP-7, a gelatinase-A and matrilysin-1, was elevated and further secreted into the medium by BMM. It is known that MMP-2 digests collagen-I, II, and III and MMP-7 hydrolyzes cell surface molecules such as E-cadherin and a number of ECM components [
23]. These observations suggest that BMM modulates the secretion of MMP-2/7 but not MMP-1 to the basement membrane and promotes migration (
Figure 3).
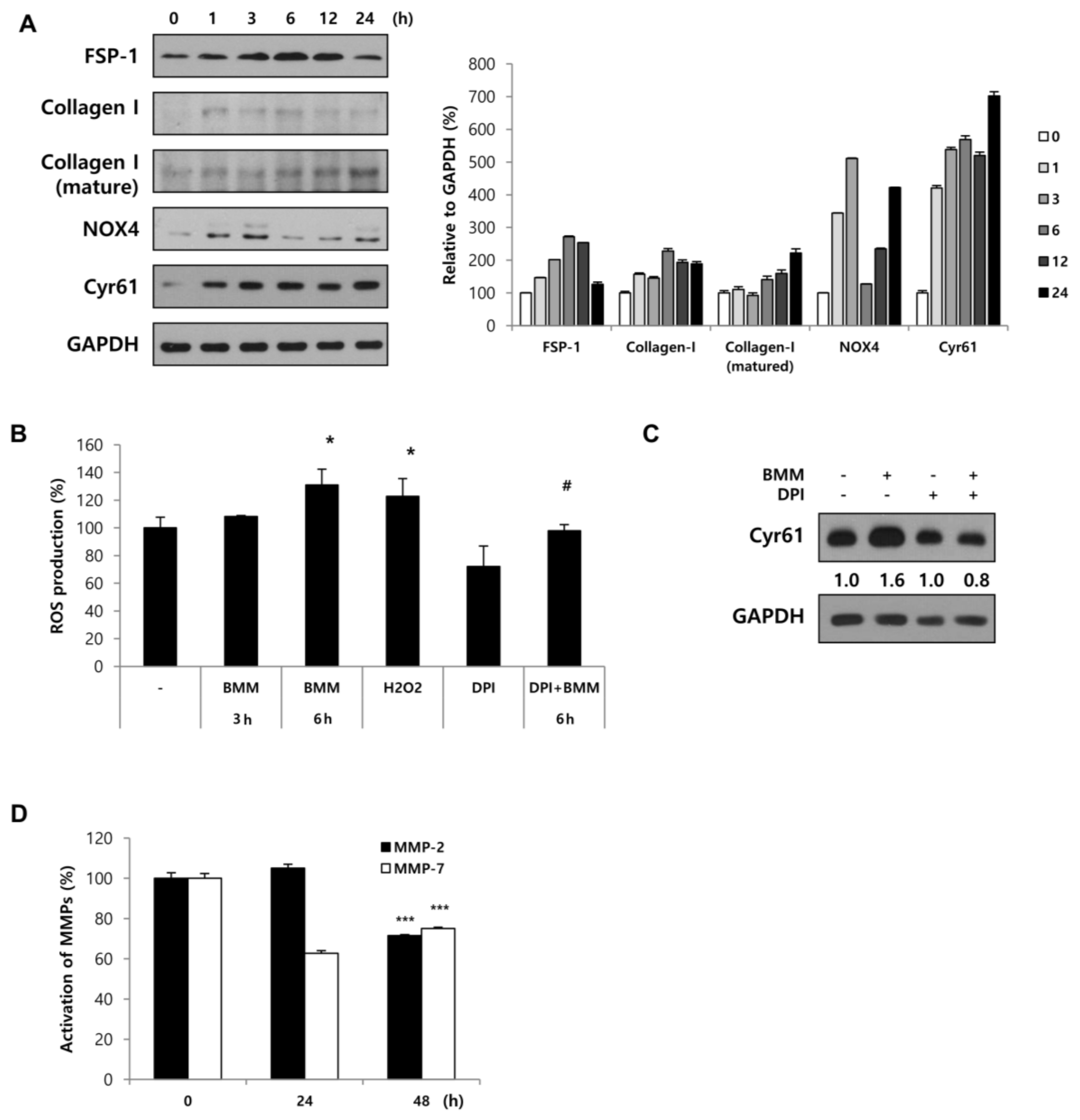
During re-epithelialization of the epidermis, ‘fibroplasia’ occurs in the dermis. Fbs increase proliferation around wound sites and form granulation tissue. Many types of growth factors or cytokines are secreted after injury, and these activate Fbs to differentiate into myofibroblasts, featured as contractile, differentiated, and capable of ECM production. In the last step of healing, the resolution phase, myofibroblasts are dramatically reduced by cell death, reducing ECM synthesis. However, it is not yet known whether myofibroblasts convert into the normal fibroblasts phenotype or a quiescent phenotype [
5].
As a result of our experiments, we observed the activation to myofibroblasts by BMM, and it was induced by ROS generation via NOX4 activation. DPI inhibited Cyr61 expression at early time points; therefore, we speculated that the increase in ROS-related stress due to NOX4 stimulates Cyr61 expression and that this expression forms a positive feedback loop between Cyr61 and ROS. In a study by Kim et al. [
24], Cyr61 (CCN1) functions to trigger cellular senescence by generating ROS through the induction of a RAC1-NOX1 complex in hepatic myofibroblasts [
24]. Recently, CCN1 was identified as a pro-inflammatory factor that binds integrin α6β1 in keratinocytes [
25]. Through this theory, we speculated that secreted Cyr61 from Fbs reaches keratinocytes and activates the integrin pathway, which is a specific receptor of FAK/Src. Further research is required to confirm these findings.
Angiogenesis is also an essential process for healing, as well as epithelialization and fibroplasia. Newly formed vessels offer micronutrients, amino acids, and oxygen to impaired skin [
26]. To detect the vascularization effect of BMM, we investigated the vessels under the skin after burn-injury. Compared to the control group, BMM had no effect on angiogenesis.
Epithelial recovery, indicated by the upregulation of keratin-10, was found in the excisional wound model, as well as in an in vitro experiment. In the case of dermal recovery, the appearance of myofibroblasts labeled with FSP-1 was shown to be inconsistent in vitro and in vivo. The role of myofibroblasts during wound healing varies with each step. Myofibroblasts form granulation tissue at the proliferative stage, induce a contractile effect, and increase to synthesize ECM. Since excessive ECM deposition results in fibrosis and scarring, this phenomenon is reduced by apoptosis. These results suggest that BMM can help repair wounds by affecting the epidermis and dermis, and has no side effect such as scar formation. Rather, it inhibited scar formation (
Figure 6 and
Figure 7).
Interestingly, when HaCaT and Fbs are cultured together, HaCaT motility was significantly increased and BMM treatment further promoted. As shown in
Figure 5C, this phenomenon was probably caused by synergistically increased TGF-β from HaCaT and Cyr61 from Fbs than from single cultured cells.
In recent research certifying the synergistic effect of a mixture of TMF and glycitin, we emphasized the interaction between HaCaTs and Fbs during wound repair [
12]. Cytokines or growth factors secreted from HaCaTs and Fbs affect other cells and promote keratinocyte regeneration and fibroblast activation.
Here, we demonstrate the wound healing effect of BMM by stimulating re-epithelialization and dermal differentiation. As a mechanism for HaCaT migration by BMM, TGF-β-EMT and FAK/Src signaling were revealed and the Cyr61-NOX4 pathway was a core mechanism for the differentiation of Fbs. Furthermore, skin cells stimulated by BMM beneficially interact with one another and showed a synergistic effect. These findings suggest that the novel synthetic material, BMM, is a potential drug candidate for the treatment of skin injury.
4. Materials and Methods
4.1. Generation and Verification of BMM
Compound
I (4-methoxybenzohydrazide) was purchased from Sigma-Aldrich (St. Louis, MO, USA) and used without purification. Compound
II was prepared according to previously described methods [
27]. Compound
I (166 mg, 1 mmol) and a catalytic amount of concentrated hydrochloric acid were added to a solution of compound
II (237 mg, 1 mmol) in ethanol. The reaction mixture was refluxed at 90 °C for 2 h. After completion of the reaction, the reaction mixture was cooled to room temperature to allow precipitation. The solid was filtered and washed with cold ethanol to yield a pure Compound
III (yield: 50%, melting point: 238–240 °C). The synthesis procedure for Compound
III, 1E,2E-1,2-bis((6-bromo-2H-chromen-3-yl)methylene)hydrazine, is shown in
Figure 1A and its structure and numbering is shown in
Figure 1B. The experimental methods for NMR spectroscopy and mass spectrometry to identify the compound followed a previously reported method [
28]. Spectroscopic data are summarized as follows: 1H NMR (500 MHz, pyridine-d5) δ ppm: 8.34 (s, 2H, H-3a), 7.37 (dd, 2H, H-7,
J = 2.4, 8.4 Hz), 7.35 (d, 2H, H-5,
J = 2.4 Hz), 6.94 (s, 2H, H-4), 6.85 (d, 2H, H-8,
J = 8.4), 5.31 (s, 4H, H-2); 13C NMR (125 MHz, pyridine-d5) δ ppm: 160.3 (C-3a), 155.1 (C-9), 133.8 (C-7), 131.6 (C-4), 130.9 (C-5), 130.6 (C-3), 124.8 (C-6), 118.3 (C-8), 113.8 (C-10), 65.0 (C-2); HRMS (
m/
z): calculated for [M + H]
+: 472.9500.
BMM ([1E,2E-1,2-bis(6-bromo-2H-chromen-3-yl)methylene]hydrazine) was dissolved in dimethyl sulfoxide (DMSO) and stored at −20 °C (Stock = 20 μM).
4.2. Cell Culture
HaCaTs and primary human FBs were cultured in DMEM (Gibco, Carlsbad, CA, USA) supplemented with 10% FBS (Omega, Biel/Bienne, Switzerland) and 1% penicillin/streptomycin. Cells were incubated at 37 °C in 5% CO2.
4.3. MTT Assay
Cells (2 × 104 cell/mL) were seeded in 96-well plates. The following day, cells were treated with DMSO (Sigma-Aldrich, St Louis, MO, USA) and BMM. After incubation for 24 or 48 h with HaCaT or Fbs, 10 μL of MTT solution (10 mg/mL) was added to each well, and cells were incubated for an additional 4 h at 37 °C. The medium was removed and replaced with 150 μL DMSO, and the plate was incubated for 30 min at room temperature with shaking. Absorbance was measured at 570 nm using a spectrophotometer.
4.4. ECIS Proliferation Assay
ECIS is an experimental machine to analyze the proliferation or migration of cells in real time and show resistance values as a graph. The ECIS cell culture plate consists of six wells coated with metal on the bottom. As cells grow, resistance detected by metal is increased and is converted to impedance through a self-program. Two hundred microliters of electrode-stabilizing solution, composed of 10 mM l-cysteine, were added to each well and incubated at room temperature. After 10 min, the wells were washed using medium and the cells were seeded at a density of 2 × 104 cell/mL. The following day, 0–20 μL of BMM was added to the wells and the cells were incubated for 48 h. Using the ECIS program, we transferred the Excel data sheet and calculated normalized impedance.
4.5. Cell Counting
The cell counting method used was one of the cell proliferation assays. A six-well plate was used for the cell counting assay (two wells per time point) and 5 × 103 cells were seeded in each well. Cells were treated with 0–20 μL of BMM and incubated at 37 °C. After incubation for two, four, or six days, cells were collected using trypsin and centrifuged. Collected cells were suspended and counted using a hemocytometer. Every two days, medium containing the diluted sample was carefully changed.
4.6. Scratch Wound Healing Assay
Cells were seeded in 48-well plates at a density of 1 × 105 cell/well for 24 h. A scratch was made on the cell monolayer by drawing a pipette tip across the well. The culture medium was supplemented with DMSO or BMM dose-dependently. At time 0 and 24 h post-treatment, wound closure was captured using an Olympus IX70 microscope (Tokyo, Japan) equipped with a digital camera at 40× magnification. The distance migrated was measured using the Image J software (Lviv, Ukraine), and calculated by comparing between the initial and final width of the scratch.
4.7. Matrigel Invasion Assay
We performed two types of Matrigel invasion assays: (1) single-culture or (2) co-culture. For single culture assays, 7 × 104 cell/well of HaCaT or Fbs were seeded in the insert of a 12-well invasion assay kit (SPL), and the bottom chamber was filled with media. On the other hand, in case of the co-culture assays, HaCaT cells were seeded in the insert, and the Fbs were seeded into the bottom, using equal numbers of each. The membrane allows the exchange of media during incubation. After 24 h, media containing DMSO or BMM was added to the insert and bottom for 24 h. Cells on the upper side of the insert were removed using a cotton swab, and cells on the lower side of the insert were fixed with 4% formaldehyde. After washing with PBS, invaded cells were stained with a 1% crystal violet solution and photographs were acquired at 4× magnification using an Olympus IX70 microscope. Invasive ability was measured using the ImageJ program.
4.8. Western Blot
HaCaT, Fbs, or co-cultured cells were seeded at a density of 1 × 105 each on 100 mm cell culture dishes and incubated for 24 h. These were treated with DMSO or BMM in a time or dose dependent manner. Total protein was extracted from treated cells using RIPA buffer, and the protein concentration was determined using the BCA Protein Assay Kit (Thermo Scientific, Walthm, MA, USA). To investigate the expression of secreted factors, conditioned medium was harvested and concentrated using an Amicon centrifugal filter (Merck Millipore, Burlington, MA, USA), and total protein concentration was measured using the Bradford assay. Equal amounts of protein (~30 μg/lane) were analyzed from each sample by resolving with 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Protein loaded gels were transferred to NC membranes, and these were blocked in 5% skim milk with Tris-buffered saline Tween 20 (TBST) buffer, followed by incubation with primary antibodies at 4 °C overnight. Cytokeratin 10 (sc-53252), Cytokeratin 14 (sc-17104), MMP-7 (sc-8832), Cyr61 (sc-271217), Collagen-I (sc-25974),gp91-phox (sc-130543), FAK (sc-1688), p-FAK (sc-11765), C-Src (sc-130124), and GAPDH (sc-25778) antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA); S100A4 (ab27957) antibody was purchased from ABcam (Bristol, UK); TGF-β (#3711), Slug (#9585), E-cadherin (#3195), Snail (#3879), p-Src (#2101), and MMP-2 (#4022) antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). NOX4 (NB110-58849) and MMP-1 (#444209) antibodies were purchased from Novus Biologicals (Littleton, CO, USA) and Calbiochem (San Diego, CA, USA), respectively. We used HRP-conjugated anti-mouse (K0211589, KOMABIOTECH, Seoul, Korea), anti-rabbit (K021178, KOMABIOTECH), and anti-goat (AP1079P, Millipore) as secondary antibodies. Proteins were detected using chemiluminescent reagent, ECL solution (W6002, Biosesang, Seongnam, Korea).
4.9. 2′,7′-Dichlorofluorescein Diacetate (DCF-DA) Assay
DCF-DA was used to detect intracellular ROS production. Dermal fibroblast cells (2 × 104 cell/mL) were seeded in 96-well plates. To confirm the relationship between NOX and Cyr61, the cells were treated with BMM (20 μM) for 3 or 6 h or pre-treated with DPI (5 μM) for 1 h on the following day. Cells treated with H2O2 (100 μM) for 1 h before detection were used as a positive control, following which they were washed using 1× PBS. DCF-DA solution, which is a 1× PBS solution containing 1% FBS and 20 μM DCF-DA, was added to each well. Fluorescence was measured using a fluorescence plate reader (TECAN GENios, Grodig, Austria) with the following settings: excitation, 485 nm; emission, 535 nm.
4.10. In Vivo Experiments
Six-week old male ICR mice (n = 6 for each group) were chosen for the experiment. All procedures were approved by the Animal Care and Use Committee at Jeju National University (permission number 2015-0033, 29 December 2015). First, the fur was removed with an electronic hair clipper and removal cream. Then, wounds were made on the middle of the back using a 6-mm biopsy punch. Wounds of mice in the experimental group were treated daily with 200 μL of BMM (200 μM) dissolved in butylene glycol for two weeks. Madecassol was used as a positive control and applied to wound sites in the same manner as BMM. Images were captured every two days to evaluate wound closure and the rate was calculated as a relative percentage of the original wound area, using the ImageJ program. At seven, nine, and 14 days post-wound, skin tissue was isolated and fixed using a 4% formaldehyde solution. Paraffin-embedded tissues were cut into 3-μm sections and stained using Hematoxylin-Eosin and Immunohistochemistry of Keratin 10 and FSP-1.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






