
Detection of SARS-CoV-2 Derived Small RNAs and Changes in Circulating Small RNAs Associated with COVID-19

 , and
, and
Abstract
:
1. Introduction
- Do we see circulating small RNAs corresponding to parts of viral genome, and can they be used as marker for ongoing or cleared infections?
- Do we see circulating small RNAs corresponding to parts of viral genome with distinct features?
2. Methods
2.1. Patients and Samples
2.2. Detection of SARS-CoV-2 Antibody in Human Plasma
2.3. RNA Isolation, Library Prep and Sequencing
3. Results
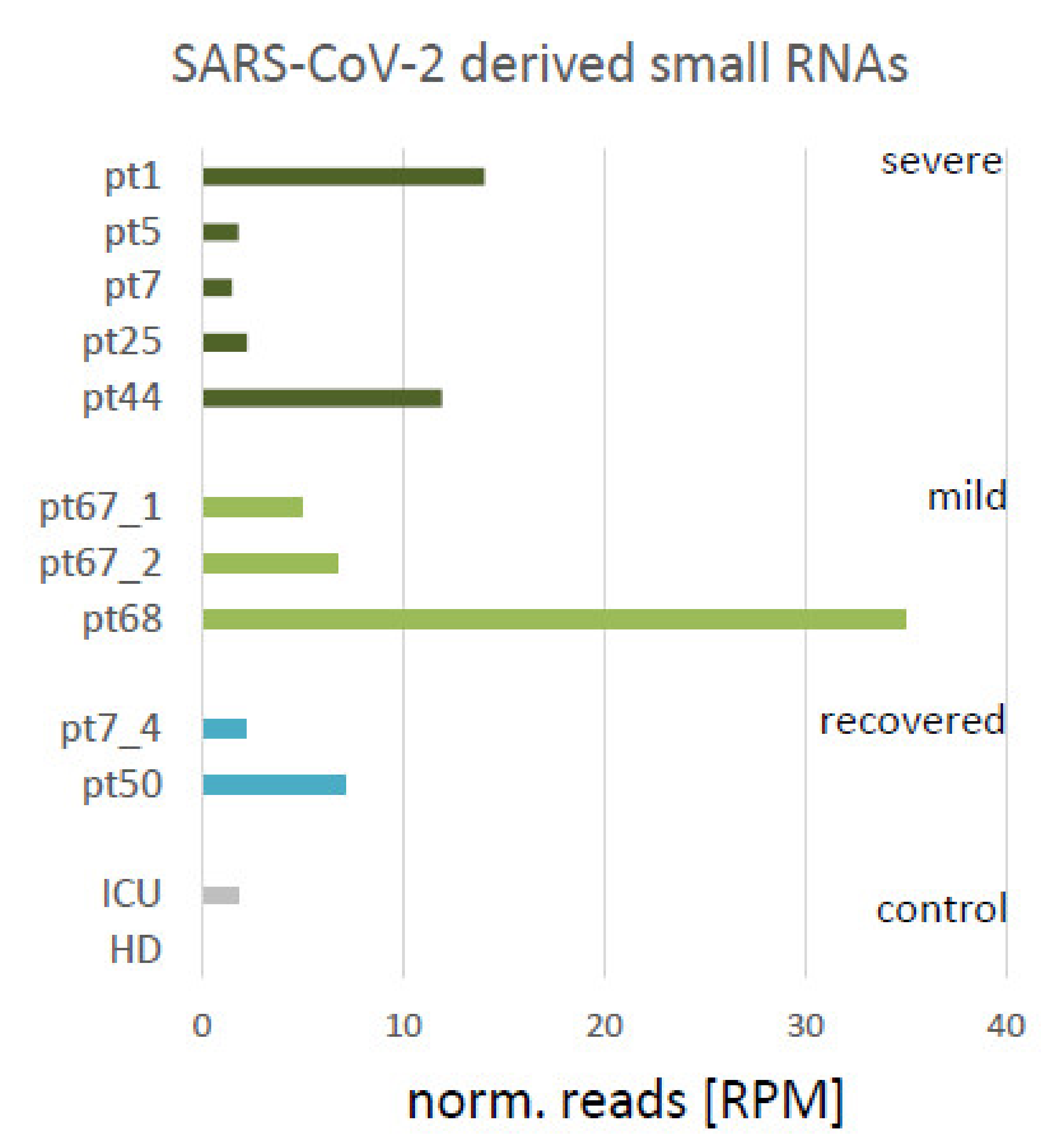
3.1. Small SARS-CoV-2 Derived RNAs Are Detectable in Plasma of COVID-19 Patients
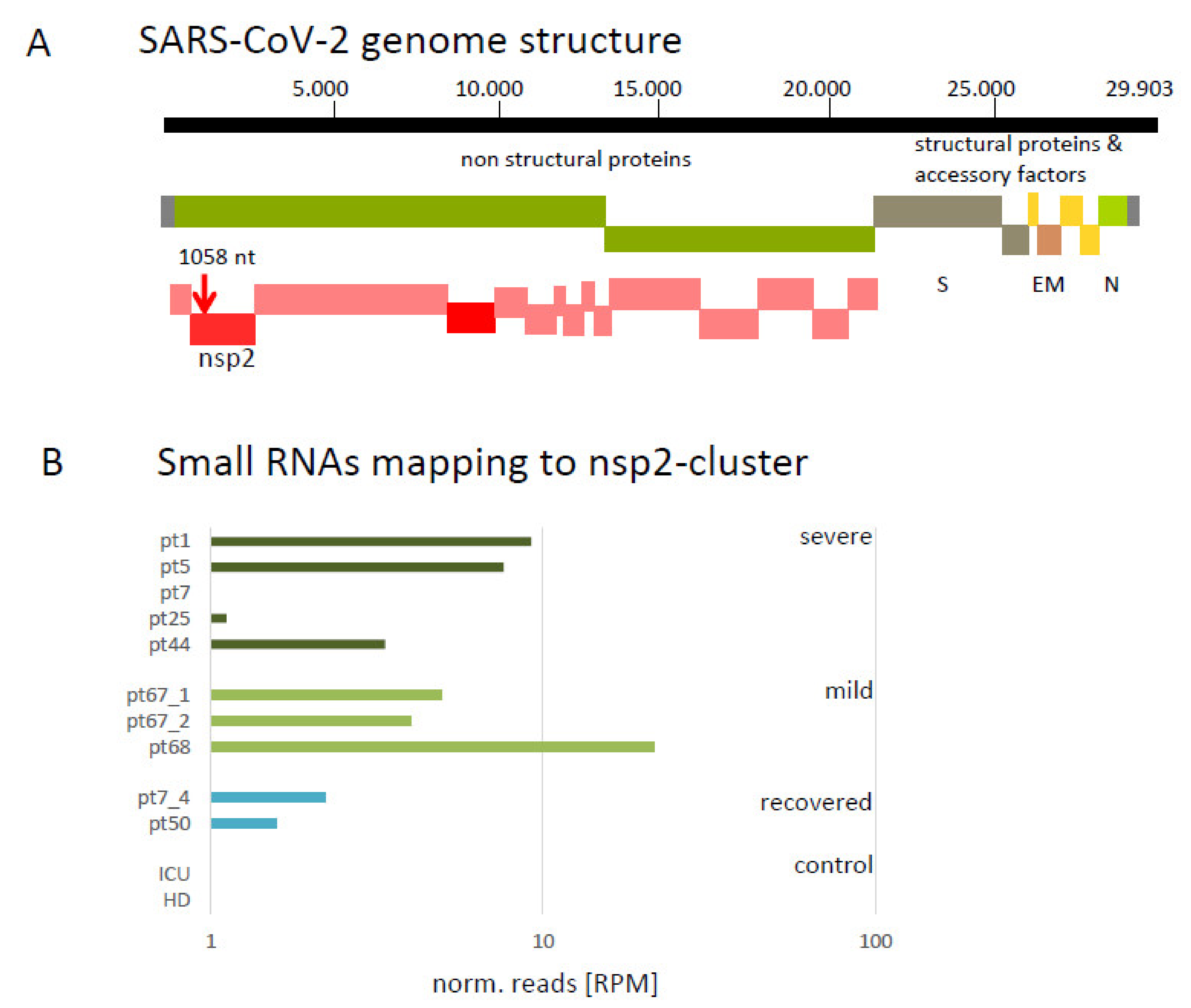
3.2. Mapping of Small Virus-Derived RNAs Reveal a Cluster in the 3′-Region Coding for nsp2 and Four 19 nt Long Small RNA Sequences
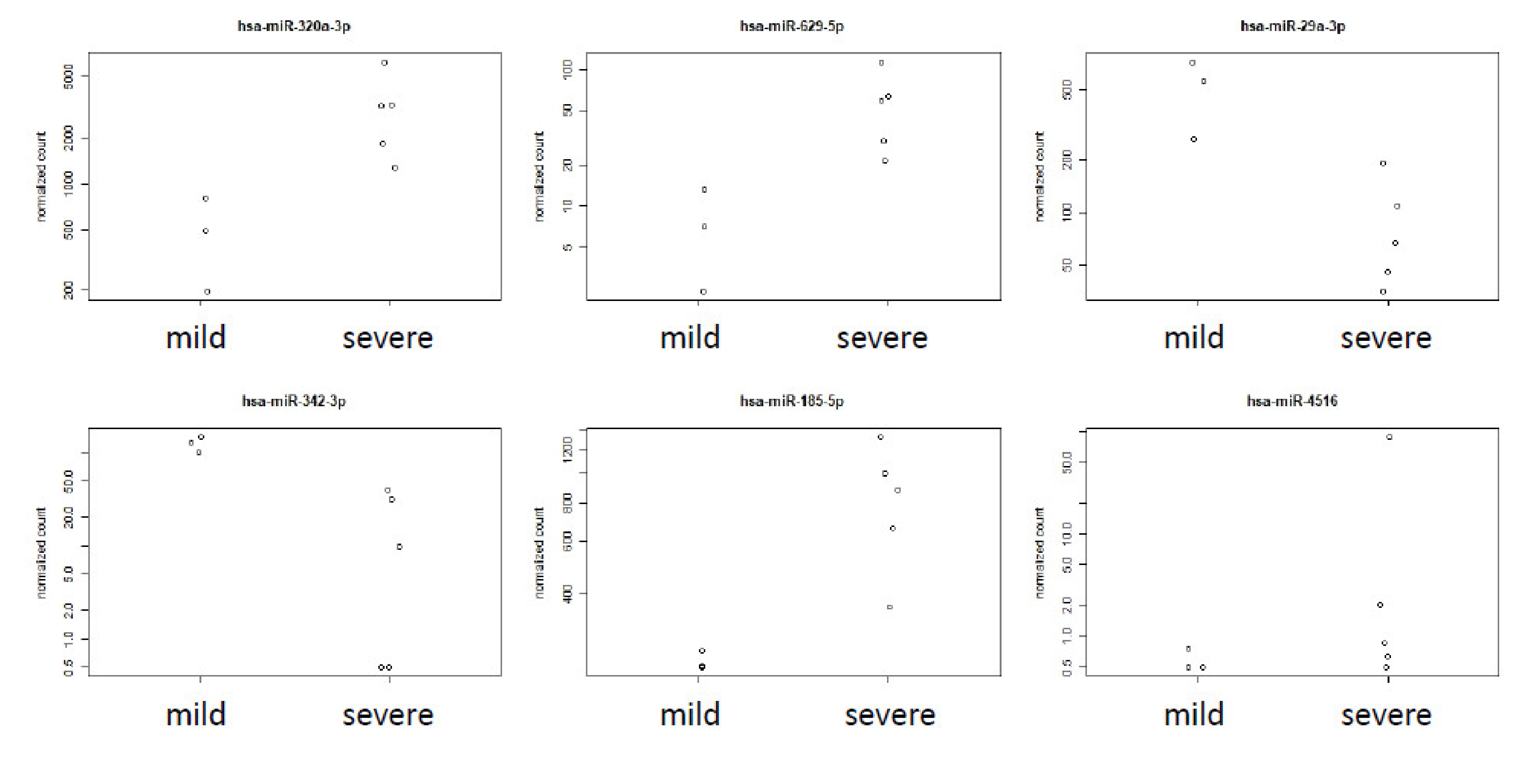
3.3. Analysis of Circulating Human miRNA Showed Differences in Samples of Mild and Severe Cases
4. Discussion
4.1. SARS-CoV-2 Derived Small RNAs Are Detectable
4.2. Small Virus-Derived RNAs from the Cluster Might Be Usable as Biomarkers
4.3. Severe Cases of COVID-19 Are Associated with Differential Abundance of Specific Circulating Human miRNAs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Felsenstein, S.; Herbert, J.; McNamara, P.S.; Hedrich, C.M. COVID-19: Immunology and treatment options. Clin. Immunol. 2020, 215, 108448. [Google Scholar] [CrossRef] [PubMed]
- Rosano, A.; Bella, A.; Gesualdo, F.; Acampora, A.; Pezzotti, P.; Marchetti, S.; Ricciardi, W.; Rizzo, C. Investigating the impact of influenza on excess mortality in all ages in Italy during recent seasons (2013/14–2016/17 seasons). Int. J. Infect. Dis. 2019, 88, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Munster, V.J.; Koopmans, M.; van Doremalen, N.; van Riel, D.; de Wit, E. A novel coronavirus emerging in China—Key questions for impact assessment. N. Engl. J. Med. 2020, 382, 692–694. [Google Scholar] [CrossRef]
- Clift, M.A.K.; Coupland, C.A.; Keogh, R.H.; Hemingway, H.; Hippisley-Cox, J. COVID-19 mortality risk in Down syndrome: Results from a cohort study of 8 million adults. Ann. Intern. Med. 2021, 174, 572–576. [Google Scholar] [CrossRef]
- Cullen, B.R. RNA interference: Antiviral defense and genetic tool. Nat. Immunol. 2002, 3, 597–599. [Google Scholar] [CrossRef]
- Shabalina, S.A.; Koonin, E.V. Origins and evolution of eukaryotic RNA interference. Trends Ecol. Evol. 2008, 23, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Grundhoff, A.; Sullivan, C.S. Virus-encoded microRNAs. Virology 2011, 411, 325–343. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Hackbart, M.; Mettelman, R.; O’Brien, A.; Mielech, A.M.; Yi, G.; Kao, C.C.; Baker, S.C. Coronavirus nonstructural protein 15 mediates evasion of dsRNA sensors and limits apoptosis in macrophages. Proc. Natl. Acad. Sci. USA 2017, 114, E4251–E4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeang, K.-T. RNAi in the regulation of mammalian viral infections. BMC Biol. 2012, 10, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.-J.; Tang, Q.; Cheng, D.; Qin, C.; Xie, F.Y.; Wei, Q.; Xu, J.; Liu, Y.; Zheng, B.-J.; Woodle, M.C.; et al. Using siRNA in prophylactic and therapeutic regimens against SARS coronavirus in Rhesus macaque. Nat. Med. 2005, 11, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, Y.-L.; Yang, F.; Zhang, Y.; Wang, S.-H.; Liu, L. Small interfering RNA effectively inhibits the expression of SARS coronavirus membrane gene at two novel targeting sites. Molecules 2010, 15, 7197–7207. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Yuan, Y.; Cho, J.-H.; McClarty, S.; Baxter, D.; Galas, D.J. Comparing the microRNA spectrum between serum and plasma. PLoS ONE 2012, 7, e41561. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Wang, J.; Wang, Z.; Sun, Y.; Wu, J.; Zhang, Y.; Liu, X.; Zhou, Z.; Zhou, L.; Zhang, C.Y.; et al. A virus-derived microRNA-like small RNA serves as a serum biomarker to prioritize the COVID-19 patients at high risk of developing severe disease. Cell Discov. 2021, 7, 48. [Google Scholar] [CrossRef]
- Rong, X.; Jia, L.; Hong, L.; Pan, L.; Xue, X.; Zhang, C.; Lu, J.; Jin, Z.; Qiu, H.; Wu, R.; et al. Serum miR-92a-3p as a new potential biomarker for diagnosis of Kawasaki disease with coronary artery lesions. J. Cardiovasc. Transl. Res. 2016, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tijsen, A.J.; Pinto, Y.M.; Creemers, E.E. Circulating microRNAs as diagnostic biomarkers for cardiovascular diseases. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1085–H1095. [Google Scholar] [CrossRef] [Green Version]
- Geekiyanage, H.; Rayatpisheh, S.; Wohlschlegel, J.A.; Brown, R.; Ambros, V. Extracellular microRNAs in human circulation are associated with miRISC complexes that are accessible to anti-AGO2 antibody and can bind target mimic oligonucleotides. Proc. Natl. Acad. Sci. USA 2020, 117, 24213–24223. [Google Scholar] [CrossRef]
- Godoy, P.M.; Bhakta, N.R.; Barczak, A.J.; Cakmak, H.; Fisher, S.; MacKenzie, T.C.; Patel, T.; Price, R.W.; Smith, J.F.; Woodruff, P.G.; et al. Large differences in small RNA composition between human biofluids. Cell Rep. 2018, 25, 1346–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothan, H.A.; Byrareddy, S.N. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef]
- Patterson, E.I.; Prince, T.; Anderson, E.R.; Casas-Sanchez, A.; Smith, S.L.; Cansado-Utrilla, C.; Solomon, T.; Griffiths, M.J.; Acosta-Serrano, A.; Turtle, L.; et al. Methods of inactivation of SARS-CoV-2 for downstream biological assays. J. Infect. Dis. 2020, 222, 1462–1467. [Google Scholar] [CrossRef]
- Lipps, C.; Northe, P.; Figueiredo, R.; Rohde, M.; Brahmer, A.; Krämer-Albers, E.-M.; Liebetrau, C.; Wiedenroth, C.B.; Mayer, E.; Kriechbaum, S.D.; et al. Non-invasive approach for evaluation of pulmonary hypertension using extracellular vesicle-associated small non-coding RNA. Biomolecules 2019, 9, 666. [Google Scholar] [CrossRef] [Green Version]
- Schultheiß, C.; Paschold, L.; Simnica, D.; Mohme, M.; Willscher, E.; von Wenserski, L.; Scholz, R.; Wieters, I.; Dahlke, C.; Tolosa, E.; et al. Next-generation sequencing of T and B cell receptor repertoires from COVID-19 patients showed signatures associated with severity of disease. Immunity 2020, 53, 442–455.e4. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Zhu, H.; Gao, B.; Weng, H.; Ding, Z.; Li, M.; Weng, X.; He, G. Diagnosis of severe community-acquired pneumonia caused by Acinetobacter baumannii through next-generation sequencing: A case report. BMC Infect. Dis. 2020, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Du, R.; Edwards, A.; Flemington, E.K.; Zhang, K. The sequence structures of human microRNA molecules and their implications. PLoS ONE 2013, 8, e54215. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [Green Version]
- Li, F. Taste perception: From the tongue to the testis. Mol. Hum. Reprod. 2013, 19, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Marks, C.; Belluscio, L.; Ibáñez, C.F. Critical role of GFR1 in the development and function of the main olfactory system. J. Neurosci. 2012, 32, 17306–17320. [Google Scholar] [CrossRef]
- Cheng, Y.C.; O’Connell, J.R.; Cole, J.W.; Stine, O.C.; Dueker, N.; McArdle, P.F.; Sparks, M.J.; Shen, J.; Laurie, C.C.; Nelson, S.; et al. Genome-wide association analysis of ischemic stroke in young adults. G3 Genes Genomes Genet. 2011, 1, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Piatto, V.B.; De Carvalho, T.B.O.; De Marchi, N.S.A.; Molina, F.D.; Maniglia, J.V. Polymorphisms in the 5-HTR2A gene related to obstructive sleep apnea syndrome. Braz. J. Otorhinolaryngol. 2011, 77, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.; Staines, D.; Nilius, B.; Smith, P.; Marshall-Gradisnik, S. Novel identification and characterisation of transient receptor potential melastatin 3 ion channels on natural killer cells and B lymphocytes: Effects on cell signalling in chronic fatigue syndrome/myalgic encephalomyelitis patients. Biol. Res. 2016, 49, 1–8. [Google Scholar] [CrossRef]
- Meini, S.; Giani, T.; Tascini, C. Intussusceptive angiogenesis in Covid-19: Hypothesis on the significance and focus on the possible role of FGF2. Mol. Biol. Rep. 2020, 47, 8301–8304. [Google Scholar] [CrossRef]
- Smadja, D.M.; Philippe, A.; Bory, O.; Gendron, N.; Beauvais, A.; Gruest, M.; Peron, N.; Khider, L.; Guerin, C.L.; Goudot, G.; et al. Placental growth factor level in plasma predicts COVID-19 severity and in-hospital mortality. J. Thromb. Haemost. 2021, 19, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Seong, R.-K.; Seo, S.-W.; Kim, J.-A.; Fletcher, S.J.; Morgan, N.; Kumar, M.; Choi, Y.-K.; Shin, O.S. Schlafen 14 (SLFN14) is a novel antiviral factor involved in the control of viral replication. Immunobiology 2017, 222, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2018, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Fan, J.; Zhao, Y.; Zhang, X.; Dai, B.; Zhan, J.; Yin, Z.; Nie, X.; Fu, X.-D.; Chen, C.; et al. Nuclear miR-320 mediates diabetes-induced cardiac dysfunction by activating transcription of fatty acid metabolic genes to cause lipotoxicity in the heart. Circ. Res. 2019, 125, 1106–1120. [Google Scholar] [CrossRef]
- Liu, L.; Li, X. Downregulation of miR-320 alleviates endoplasmic reticulum stress and inflammatory response in 3T3-L1 adipocytes. Exp. Clin. Endocrinol. Diabetes 2019, 129, 131–137. [Google Scholar] [CrossRef]
- Tian, Z.-Q.; Jiang, H.; Lu, Z.-B. MiR-320 regulates cardiomyocyte apoptosis induced by ischemia–reperfusion injury by targeting AKIP1. Cell. Mol. Biol. Lett. 2018, 23, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndzi, E.N.; Nkenfou, C.N.; Mekue, L.M.; Zentilin, L.; Tamgue, O.; Pefura, E.W.Y.; Kuiate, J.R.; Giacca, M.; Ndjolo, A. MicroRNA hsa-miR-29a-3p is a plasma biomarker for the differential diagnosis and monitoring of tuberculosis. Tuberculosis 2019, 114, 69–76. [Google Scholar] [CrossRef]
- Kirschner, M.B.; Cheng, Y.Y.; Badrian, B.; Kao, S.C.; Creaney, J.; Edelman, J.J.B.; Armstrong, N.J.; Vallely, M.P.; Musk, A.W.; Robinson, B.W.; et al. Increased Circulating miR-625-3p: A potential biomarker for patients with malignant pleural mesothelioma. J. Thorac. Oncol. 2012, 7, 1184–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.A.; Jhun, B.W.; Kim, S.-Y.; Moon, S.M.; Yang, B.; Kwon, O.J.; Daley, C.L.; Shin, S.J.; Koh, W.-J. miRNA expression profiles and potential as biomarkers in nontuberculous mycobacterial pulmonary disease. Sci. Rep. 2020, 10, 3178. [Google Scholar] [CrossRef] [Green Version]
- Qian, F.-H.; Deng, X.; Zhuang, Q.-X.; Wei, B.; Zheng, D.-D. miR-625-5p suppresses inflammatory responses by targeting AKT2 in human bronchial epithelial cells. Mol. Med. Rep. 2019, 19, 1951–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graier, T.; Golob-Schwarzl, N.; Weger, W.; Benezeder, T.; Painsi, C.; Salmhofer, W.; Wolf, P. Furin expression in patients with psoriasis—A patient cohort endangered to SARS-COV2? Front. Med. 2021, 8, 624462. [Google Scholar] [CrossRef]
- Nejad, C.; Stunden, H.J.; Gantier, M.P. A guide to miRNAs in inflammation and innate immune responses. FEBS J. 2018, 285, 3695–3716. [Google Scholar] [CrossRef] [PubMed]
- Alam, T.; Lipovich, L. miRCOVID-19: Potential targets of human miRNAs in SARS-CoV-2 for RNA-based drug discovery. Noncoding RNA 2021, 7, 18. [Google Scholar] [PubMed]
- Lee, K.-M.; Gong, Y.-N.; Shih, S.-R. Methods for detection and study of virus-derived small RNAs produced from the intramolecular base-pairing region of the picornavirus genome. Methods 2019, 183, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.; Courtney, D.G.; Kennedy, E.M.; Cullen, B.R. Influenza A virus-derived siRNAs increase in the absence of NS1 yet fail to inhibit virus replication. RNA 2018, 24, 1172–1182. [Google Scholar] [CrossRef]
- Schuster, S.; Overheul, G.; Bauer, L.; Van Kuppeveld, F.J.M.; Van Rij, R.P. No evidence for viral small RNA production and antiviral function of Argonaute 2 in human cells. Sci. Rep. 2019, 9, 13752. [Google Scholar] [CrossRef] [Green Version]
- Merino, G.A.; Raad, J.; Bugnon, L.A.; Yones, C.; Kamenetzky, L.; Claus, J.; Ariel, F.; Milone, D.H.; Stegmayer, G. Novel SARS-CoV-2 encoded small RNAs in the passage to humans. Bioinformatics 2021, 36, 5571–5581. [Google Scholar] [CrossRef]
- Kincaid, R.P.; Burke, J.M.; Sullivan, C.S. RNA virus microRNA that mimics a B-cell oncomiR. Proc. Natl. Acad. Sci. USA 2012, 109, 3077–3082. [Google Scholar] [CrossRef] [Green Version]
- Duy, J.; Honko, A.N.; Altamura, L.A.; Bixler, S.L.; Wollen-Roberts, S.; Wauquier, N.; O’Hearn, A.; Mucker, E.M.; Johnson, J.C.; Shamblin, J.D.; et al. Virus-encoded miRNAs in Ebola virus disease. Sci. Rep. 2018, 8, 6480. [Google Scholar] [CrossRef]
- Dani, M.; Dirksen, A.; Taraborrelli, P.; Torocastro, M.; Panagopoulos, D.; Sutton, R.; Lim, P.B. Autonomic dysfunction in “long COVID”: Rationale, physiology and management strategies. Clin. Med. 2021, 21, e63–e67. [Google Scholar] [CrossRef]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-time tracking of pathogen evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef] [PubMed]
- Varkonyi-Gasic, E.; Wu, R.; Wood, M.; Walton, E.F.; Hellens, R.P. Protocol: A highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods 2007, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dao, T.L.; Hoang, V.T.; Gautret, P. Recurrence of SARS-CoV-2 viral RNA in recovered COVID-19 patients: A narrative review. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 13–25. [Google Scholar] [CrossRef]
- Roltgen, K.; Wirz, O.F.; Stevens, B.A.; Powell, A.E.; Hogan, C.A.; Najeeb, J.; Hunter, M.; Sahoo, M.K.; Huang, C.; Yamamoto, F.; et al. SARS-CoV-2 antibody responses correlate with resolution of RNAemia but are short-lived in patients with mild illness. medRxiv 2020. [Google Scholar] [CrossRef]
- Yang, C.; Jiang, M.; Wang, X.; Tang, X.; Fang, S.; Li, H.; Zuo, L.; Jiang, Y.; Zhong, Y.; Chen, Q.; et al. Viral RNA level, serum antibody responses, and transmission risk in recovered COVID-19 patients with recurrent positive SARS-CoV-2 RNA test results: A population-based observational cohort study. Emerg. Microbes Infect. 2020, 9, 2368–2378. [Google Scholar] [CrossRef] [PubMed]
- Azghandi, M.; Kerachian, M.A. Detection of novel coronavirus (SARS-CoV-2) RNA in peripheral blood specimens. J. Transl. Med. 2020, 18, 412. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Lu, R.-B.; Wang, L.-J.; Chang, C.-H.; Lu, T.; Wang, T.-Y.; Tsai, K.-W. Serum miRNA as a possible biomarker in the diagnosis of bipolar II disorder. Sci. Rep. 2020, 10, 1131. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Zhao, K.; Qi, Y.; Min, X.; Shi, Z.; Qi, X.; Shan, Y.; Cui, L.; Zhou, M.; Wang, Y.; et al. Serum microRNA expression profile as a biomarker for the diagnosis of pertussis. Mol. Biol. Rep. 2012, 40, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Leidinger, P.; Vogel, B.; Backes, C.; El Sharawy, A.; Galata, V.; Mueller, S.C.; Marquart, S.; Schrauder, M.G.; Strick, R.; et al. miRNAs can be generally associated with human pathologies as exemplified for miR-144. BMC Med. 2014, 12, 224. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Noroozi, R.; Branicki, W.; Pyrc, K.; Labaj, P.P.; Pospiech, E.; Taheri, M.; Ghafouri-Fard, S. Altered cytokine levels and immune responses in patients with SARS-CoV-2 infection and related conditions. Cytokine 2020, 133, 155143. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, D.; Mussa, B.M. Identification of novel hypothalamic microRNAs as promising therapeutics for SARS-CoV-2 by regulating ACE2 and TMPRSS2 expression: An in silico analysis. Brain Sci. 2020, 10, 666. [Google Scholar] [CrossRef]
- Tang, H.; Gao, Y.; Li, Z.; Miao, Y.; Huang, Z.; Liu, X.; Xie, L.; Li, H.; Wen, W.; Zheng, Y.; et al. The noncoding and coding transcriptional landscape of the peripheral immune response in patients with COVID-19. Clin. Transl. Med. 2020, 10, e200. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.X.; Yu, R.; Wu, X.; Wu, S.Y.; Pi, C.; Chen, Z.H.; Zhang, X.C.; Gao, C.Y.; Shao, Y.W.; Liu, L.; et al. Correlation of plasma exosomal microRNAs with the efficacy of immunotherapy in EGFR/ALK wild-type advanced non-small cell lung cancer. J. Immunother. Cancer 2020, 8, e000376. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Chen, M.; Tellgren-Roth, C.; Pettersson, U. Fluctuating expression of microRNAs in adenovirus infected cells. Virology 2015, 478, 99–111. [Google Scholar] [CrossRef]
- Keller, A.; Ludwig, N.; Fehlmann, T.; Kahraman, M.; Backes, C.; Kern, F.; Vogelmeier, C.F.; Diener, C.; Fischer, U.; Biertz, F.; et al. Low miR-150-5p and miR-320b expression predicts reduced survival of COPD patients. Cells 2019, 8, 1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Gong, Y.; Ma, J.; Gong, X. Overexpressed circ-RPL15 predicts poor survival and promotes the progression of gastric cancer via regulating miR-502-3p/OLFM4/STAT3 pathway. Biomed. Pharmacother. 2020, 127, 110219. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Yang, Q.; Xi, R.; Li, L.; Shi, D.; Chen, K. miR-941 as a promising biomarker for acute coronary syndrome. BMC Cardiovasc. Disord. 2017, 17, 227. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.; Seeliger, B.; Derda, A.A.; Xiao, K.; Gietz, A.; Scherf, K.; Sonnenschein, K.; Pink, I.; Hoeper, M.M.; Welte, T.; et al. Circulating cardiovascular microRNAs in critically ill COVID-19 patients. Eur. J. Heart Fail. 2021, 23, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Paschalaki, K.E.; Zampetaki, A.; Baker, J.R.; Birrell, M.A.; Starke, R.D.; Belvisi, M.G.; Donnelly, L.E.; Mayr, M.; Randi, A.M.; Barnes, P.J. Downregulation of microRNA-126 augments DNA damage response in cigarette smokers and patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2018, 197, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Arend, D.; Lange, M.; Chen, J.; Colmsee, C.; Flemming, S.; Hecht, D.; Scholz, U. e! DAL—A framework to store, share and publish research data. BMC Bioinform. 2014, 15, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Gender | COVID-19 | Sampling (dai) | SARS-CoV2 PCR | COVID-19 Symptoms | Color-Code | Reads |
|---|---|---|---|---|---|---|---|
| pt1 | male | pos. | 5 | pos. | severe, dead | 1849274 | |
| pt5 | male | pos. | 20 | pos. | severe | 2230128 | |
| pt7 | female | pos. | 15 | pos. | severe | 1360989 | |
| pt25 | male | pos. | 16 | pos. | severe, dead | 2694258 | |
| pt44 | male | pos. | 74 | pos. | severe, dead | 6872831 | |
| pt67_1 | female | pos. | 3 | pos. | mild | 3442766 | |
| pt67_2 | female | pos. | 17 | pos. | mild 2nd sample | 1498621 | |
| pt68 | female | pos. | 10 | pos. | mild | 829230 | |
| pt7_4 | female | pos. | 27 | neg. | recovered 2nd sample | 1364983 | |
| pt50 | female | pos. | 41 | neg. | recovered after mild | 1273895 | |
| ICU | female | neg. | 0 | neg. | none | 1132230 | |
| HD | female | neg. | 0 | neg. | none | 1318929 |
| NGS | SARS-CoV-2 Genome | ||||
|---|---|---|---|---|---|
| Sense | Antisense | ||||
| Category | Reads | Reads | Sequences | Reads | Sequences |
| severe | 13.65 × 107 | 118 | 40 | 53 | 23 |
| mild | 8.41 × 107 | 68 | 16 | 17 | 8 |
| control | 2.45 × 107 | 2 | 2 | 1 | 1 |
| log2FoldChange | p-Value | |
|---|---|---|
| hsa-miR-4516 | 5.338 | 0.00493 |
| hsa-miR-362-5p | 4.099 | 0.00912 |
| hsa-miR-548k | 3.726 | 0.02429 |
| hsa-miR-320a-3p | 2.672 | 0.00033 |
| hsa-miR-320b | 2.484 | 0.00517 |
| hsa-miR-320c | 2.339 | 0.01413 |
| hsa-miR-320d | 2.167 | 0.04336 |
| hsa-miR-185-5p | 1.812 | 0.00484 |
| hsa-miR-629-5p | 3.072 | 0.00067 |
| hsa-miR-1180-3p | 2.521 | 0.02330 |
| hsa-miR-502-3p | 1.710 | 0.03886 |
| hsa-miR-454-3p | −3.235 | 0.01749 |
| hsa-miR-625-3p | −1.879 | 0.04092 |
| hsa-miR-30b-5p | −1.893 | 0.03071 |
| hsa-miR-192-5p | −1.934 | 0.03201 |
| hsa-miR-451a | −1.945 | 0.01957 |
| hsa-miR-197-3p | −1.955 | 0.01970 |
| hsa-miR-29b-3p | −2.080 | 0.04121 |
| hsa-miR-126-3p | −2.114 | 0.01316 |
| hsa-miR-146b-5p | −2.118 | 0.01188 |
| hsa-miR-30c-5p | −2.226 | 0.01768 |
| hsa-miR-144-5p | −2.444 | 0.00713 |
| hsa-miR-29a-3p | −2.481 | 0.00100 |
| hsa-miR-363-3p | −2.508 | 0.01054 |
| hsa-miR-99a-5p | −2.523 | 0.00570 |
| hsa-miR-342-3p | −2.753 | 0.00373 |
| hsa-miR-193b-3p | −2.828 | 0.03472 |
| hsa-miR-190a-5p | −3.118 | 0.03146 |
| hsa-miR-365b-3p | −3.523 | 0.02085 |
| hsa-miR-122b-5p | −3.960 | 0.02326 |
| hsa-miR-122-3p | −4.128 | 0.02050 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grehl, C.; Schultheiß, C.; Hoffmann, K.; Binder, M.; Altmann, T.; Grosse, I.; Kuhlmann, M. Detection of SARS-CoV-2 Derived Small RNAs and Changes in Circulating Small RNAs Associated with COVID-19. Viruses 2021, 13, 1593. https://doi.org/10.3390/v13081593
Grehl C, Schultheiß C, Hoffmann K, Binder M, Altmann T, Grosse I, Kuhlmann M. Detection of SARS-CoV-2 Derived Small RNAs and Changes in Circulating Small RNAs Associated with COVID-19. Viruses. 2021; 13(8):1593. https://doi.org/10.3390/v13081593
Chicago/Turabian StyleGrehl, Claudius, Christoph Schultheiß, Katrin Hoffmann, Mascha Binder, Thomas Altmann, Ivo Grosse, and Markus Kuhlmann. 2021. "Detection of SARS-CoV-2 Derived Small RNAs and Changes in Circulating Small RNAs Associated with COVID-19" Viruses 13, no. 8: 1593. https://doi.org/10.3390/v13081593