
Biochar Volatile Matter and Feedstock Effects on Soil Nitrogen Mineralization and Soil Fungal Colonization


1
Department of Tropical Plant and Soil Sciences, College of Tropical Agriculture and Human Resources (CTAHR), University of Hawaii, Honolulu, HI 96822, USA
2
Department of Geosciences, Baylor University, Waco, TX 76706, USA
*
Author to whom correspondence should be addressed.
Sustainability 2021, 13(4), 2018; https://doi.org/10.3390/su13042018
Submission received: 8 December 2020
/
Revised: 28 January 2021
/
Accepted: 3 February 2021
/
Published: 13 February 2021
(This article belongs to the Collection Feature Papers in Sustainable Use of the Environment and Resources)
Abstract
:Biochar has important biogeochemical functions in soil—first as a means to sequester carbon, and second as a soil conditioner to potentially enhance soil quality and fertility. Volatile matter (VM) content is a property of biochar that describes its degree of thermal alteration, which can have a direct influence on carbon and nitrogen dynamics in soil. In this study, we characterized the VM in biochars derived from two locally sourced feedstocks (corncob and kiawe wood) and evaluated the relationship of VM content to nitrogen transformations and culturable fungal biomass. Using 13C nuclear magnetic resonance (NMR) spectroscopy, we found that the VM content of biochar primarily consisted of alkyl (5.1–10.1%), oxygen-substituted alkyl (2.2–6.7%), and phenolic carbon (9.4–11.6%). In a series of laboratory incubations, we demonstrated that corncob biochars with high VM (23%) content provide a source of bioavailable carbon that appeared to support enhanced viable, culturable fungi (up to 8 fold increase) and cause nitrogen immobilization in the short-term. Corncob biochar with bioavailable VM was nitrogen-limited, and the addition of nitrogen fertilizer resulted in a four-fold increase in total hydrolytic enzyme activity and the abundance of culturable fungal colonies. In contrast, kiawe biochar with an equivalent VM content differed substantially in its composition and effect on these same biological parameters. Therefore, the rapid measurement of VM content is too coarse to differentiate chemical composition and to predict the behavior of biochars across feedstocks and production methods.
1. Introduction
Biochar is widely considered to be a source of stable carbon (C) that can be applied as a soil amendment, although a broad range of turnover times have been reported, depending upon biochar properties and emergent behavior of the soil to which it is applied [1,2,3,4,5,6,7,8]. As a stable compound, biochar has the potential to sequester carbon in soil, but newly produced biochar can initially undergo decomposition upon its addition to soil [9,10] resulting in the immobilization of nutrients and the decline of plant growth [11,12,13]. The bioavailability of biochar C is not only a function of feedstock and pyrolysis conditions to the extent that they control chemical structure and composition [9,14,15,16,17,18,19], but also energy return-on-investment to the microbial community [20], which depends upon soil nutrient availability, structure, and redox conditions.
The defining characteristic of biochar is the presence of condensed polyaromatic structures [21,22] with chemical composition, structure, and functionality largely influenced by pyrolysis conditions. As biomass is continually heated from 200 to 800 °C, the carbon concentration in the biochar fraction can increase from less than 50% wt. to almost 90% wt. [23]. As pyrolysis temperatures increase beyond 450 °C, aliphatic C is lost or converted to aromatic C [24] and as temperatures approach 600 °C, the oxygen-containing functional groups and cross-linking of polyaromatic stacks volatilize [25]. By 650 °C, stronger carbonization results in the loss of hydroxyl, carbonyl, and aliphatic structures [23,26]. Keiluweit et al. [15] presented a conceptual model describing the effects of increasing temperature on biochar structure and related structural changes to biochar stability and persistence in the environment.
Volatile matter (VM) content is an easily measured property of biochar [27] that is associated with biochar elemental composition [28], and inversely related to peak temperature [15,23,29]. Volatile matter content, therefore, characterizes biochar’s degree of thermal alteration, which in turn, is related to its chemical composition [30]. Meszaros et al. [29] found that high VM biochars evolved aliphatic and aromatic fragments and carbohydrate decomposition products. Keiluweit et al. [15] observed a substantial loss of VM between 400 °C and 600 °C, which consisted of labile components. Further work demonstrated a strong relationship between biochar VM content and C mineralization [4,29]. Deenik et al. [13] found that high VM biochar contained a range of phenolic and other aromatic compounds soluble in acetone that were not present in more fully carbonized biochars with low VM content. The VM-rich biochar negatively affected plant growth through enhanced C mineralization leading to nitrogen (N) immobilization. On the other hand, Rajkovich et al. [31] found that biochar VM content, ranging from 23% to 61% in eight feedstocks, did not correlate with maize growth and N uptake, concluding that VM did not represent a microbial mineralizable fraction. In a review of the literature, Ameloot et al. [32] concluded that pyrolysis severity and feedstock had inconsistent effect on N immobilization.
The purpose of the present study was to further investigate the effects of biochar with varying VM content and C structural chemistry on microbial activity and nitrogen availability. We hypothesized that both feedstock and thermal alteration would affect the structural and chemical properties of the VM and that biochar VM content can be used as an indicator of bioavailable C. Furthermore, we posited that high VM content biochars would enhance microbial hydrolytic activity and viable, culturable fungi leading to short-term N immobilization. Our work adds to an improved understanding of the important effects of biochar VM effects on microbially driven soil N transformations.
2. Materials and Methods
We characterized biochars from corncob (Zea mays), collected from a commercial seed corn operation, and kiawe wood (Prosopis pallida), a leguminous tree found throughout the dry tropics. We obtained three corncob biochar batches produced using the flash carbonization process [33] distinct in their thermal alteration with a wide range of VM contents (34%, 23%, and 7%). The kiawe biochar, produced using traditional kiln methods by a commercial charcoal company, contained 23% VM content. Volatile matter and ash contents were determined according to the procedures outlined in ASTM D1762-84 [34]. Biochar fixed C (fC) content was derived by subtraction (fC = 100% − %VM − %ash) [28]. In addition to the corncob and kiawe biochars, we also obtained activated charcoal (CAS # 7440-440) from Fisher Scientific. Finally, we obtained samples of the raw feedstock materials for the corncob husks and kiawe wood prior to carbonization. All materials were ground and passed through a 2 mm sieve.
13C NMR spectra of the 23% and 7% VM corncob and 23% VM kiawe biochars and their corresponding water extracts was obtained using a 200 MHz Bruker spectrometer (13C frequency 50 MHz) equipped with a 4mm magic angle spinning (MAS) probe. All analyses were conducted at a rotor spinning rate of 7 kHz.
Cross polarization (CPMAS) was acquired by applying a 90 degree 1H excitation pulse, 1 ms contact pulse, two-pulse phase-modulated (TPPM) 1H decoupling, and a 3 s recycle delay. Direct polarization (DPMAS) was acquired with a 20 degree 13C excitation pulse and 5 s recycle delay. The DPMAS spectra with 1H-13C dipolar-dephasing were acquired with a 50 μs dephasing delay for quantifying non-protonated carbons. Signal relaxation during the dephasing delay was corrected using the approach of Smernik and Oades [35]. Background signals in DPMAS arising from carbon-containing probe and rotor components from each of the biochar spectra were subtracted prior to phase correction and integration. The percentage of observable biochar carbon (Cobs) was estimated for all nuclear magnetic resonance NMR spectra using Equation (1) [36].
2.1. Biochar Structural Dimensions
A 13C-1H dipolar dephasing technique based upon the DPMAS sequence (described above) was used to quantitatively measure the protonated versus the bridgehead aromatic carbons within the biochar backbone structure. The application of algorithms derived by Solum et al. [36,37] enabled an estimate of the average number of aromatic carbon atoms fused in a cluster, average number of oxygen atoms per cluster, and the average number and length of the alkyl side chains per cluster of aromatic carbons.
2.2. Incubations
We performed three independent incubation experiments with biochar-amended agricultural soil. The aim of the first incubation was to determine the interactive effect of biochar, VM content, and feedstock on fluorescein diacetate hydrolysis (FDA)—derived hydrolytic enzyme activity over a period of 60 days. The aim of the second incubation was to determine the effect of VM content on FDA hydrolytic enzyme activity and N and C dynamics in N-rich and N-limited systems over a period of 30 days. The third incubation was intended to determine the extent to which changes in soil fungal abundance and FDA hydrolytic enzyme activity could be attributed to the acetone- and water-extractable constituents of the biochar.
For laboratory incubations 1 and 2, the 23% VM corncob biochar was extracted with acetone by shaking 1 g of biochar with 10 mL of 90% acetone for 30 min, followed by filtration through 0.45 micron cellulose filter (Millipore, Billerica, MA) under vacuum. The filtrate was diluted five-fold with deionized water, placed under a nitrogen gas flow for 2 h to volatilize the acetone, and stored in a freezer. The remaining extracted biochar material was washed with 100 mL of deionized H2O in 10 mL increments to remove any remaining acetone, dried at 105 °C for 24 h, and stored for use in subsequent incubation experiments.
Incubation 1—Effects of biochar VM and feedstock on microbial abundance and activity: The 23% and 7% VM content corncob biochars, acetone-extracted 23% VM corncob biochar, 23% VM kiawe biochar, activated charcoal, and the raw corncob and kiawe materials were thoroughly mixed with an Ultisol (Leilehua series, Very fine, Ferruginous, Isothermic, Ustic, Kanhaplohumult) collected from 30–80 cm depth (see Table 1 for soil properties). All amendments were added at a 2.0% total carbon basis (weight/weight) by weighing 0.720 g of 23% VM corncob biochar (including acetone-extracted biochar), 0.608 g of 7% VM corncob, 0.784 g of kiawe biochar, 1.120 g of raw corncob, and 1.188 g raw kiawe into 25 g (oven dry equivalent) of soil. The mixtures and an unamended control soil were placed in 250-mL plastic containers and sealed with paraffin, which received six punctures to facilitate air circulation. All treatments were prepared in triplicate. Over the course of the experiment, the moisture was maintained at 50% gravimetric water content in laboratory maintained at approximately 25 °C. Incubations were sampled destructively at weeks 0, 1, 2, 4, 6, and 8.
Incubation 2—Effects of biochar VM on N dynamics: The aim of the second incubation was to determine the effect of VM content on microbial activity and N and C dynamics in N-rich and N-limited systems. The Leilehua soil was passed through a 2 mm sieve and amended with Ca(OH)2 (3 g kg−1) and Ca(H2PO4)2·H20 (320 mg P kg−1) to eliminate negative effects of acidity and phosphorus deficiency. Incubations were conducted in triplicate for soil mixtures containing corncob biochars with either 34% or 7% VM content added to achieve a rate of 2.5% (weight/weight), with the unamended soil as the control. All treatments were carried out with and without N amendment applied as NH4NO3 at a rate of 50 mg N kg−1. Incubation units were prepared and maintained as described in Incubation Study 1. Incubations were sampled destructively at days 0, 1, 3, 7, 10, 14, 21, and 28.
Incubation 3—Effects of extractable biochar constituents on fungal growth: The effect of 23% VM corncob biochar on the number of retrievable fungal colonies was investigated to determine the bioavailability of the VM fraction. Treatments included the 23% VM corncob biochar, its acetone-extracted biochar (i.e., residual biochar remaining after extraction), and the acetone extract (i.e., the biochar constituents removed by the acetone solvent). One gram of the 23% VM corncob biochar, its acetone-extracted counterpart, and all of the extracted solution were transferred to 250 mL flasks, in triplicate. A 1 mL aliquot of the inoculant solution was prepared by collecting fungal colonies isolated from cultured 23% VM corncob biochar after an incubation with soil microbes. For culture preparation, subsamples of 1 g (<60 mesh) were taken from each biochar type and placed in 25 mL plastic containers, in triplicate. Each sample received 0.1 mL of microbial inoculant and 1 mL of a modified Hoagland’s solution. The inoculant solution was prepared by using 10 mL of deionized water to 1 g of soil. Both solutions were shaken vigorously by hand, and then set in a water bath for 24 h at 35 °C. The suspensions were shaken and allowed to settle. A final solution was obtained by combining 30 mL of each microbial solution with a modified Hoagland stock solution containing 2.5 mL of 1 M KNO3, 2.5 mL of 1 M Ca(NO3)2 * 4 H2O, 1 mL of 1 M MgSO4 * 7 H2O, 1 mL of 1 M NH4NO3, and 1 mL of 1 M KH2PO4.
2.3. Microbial Assays
Fungal colonies were suspended in 100 mL of sterilized water containing added mineral nutrients. After the addition of the inoculant solution an additional 100 mL of sterilized water was added. Approximately 90 mL of sterilized water was added to bring the final volume of the biochar extract to 100 mL. The rates of mineral additions were made to achieve a final concentration of 0.25 g K2HPO4 L−1, 0.1 MgSO4 × 7H2O L−1, 0.025 NaCl L−1, and 1.44 g KNO3 L−1 for each sampling unit. Flasks were stoppered with foam and sealed with aluminum foil. Following sample preparation, the samples were incubated at 28 °C for two weeks. Samples were mixed by hand daily and placed on a horizontal shaker for 10 min on alternate days. After two weeks, we transferred 1 g of biochar into a dilution bottle containing 99 mL of sterile water and mixed vigorously for 1 min. A series of 10-fold dilutions were obtained by aseptically transferring 1 mL of the suspension into 9 mL of sterile water in 30 mL test tubes. Approximately 30 mL of the media solution, specific to fungi, was added to the petri dishes. Fungal colony counts were determined after incubating for 72 h at 28 °C with the aid of a colony counter.
We used the fluorescein diacetate hydrolysis (FDA) method as described by Green et al. [38], which is a general estimate of viable cell hydrolytic enzyme activity associated with biochemical reactions required for the mineralization of nutrients [39], and it is regularly used to measure total microbial activity in soils [40,41]. Water extractable organic C was determined in a < 0.45 µm soluble organic matter soil filtrate using a total organic carbon Analyzer Model 5000A (Shimadzu, Kyoto, Japan) according to procedures outlined in Ghani et al. [42]. For inorganic N analysis, soils were extracted with 2M KCl and filtered with Whatman 42 filtration paper. Ammonium-N and nitrate-N were determined colorimetrically using an EasyChem Discrete Analyzer (Systea Scientific, Oak Brook, IL, USA), following methods by Mulvaney [43]. The pH of the soil was measured at a 1:1 soil to deionized water ratio.
2.4. Statistical Analysis
The effects of time, biochar type, nitrogen, and biochar type by time and nitrogen by time interactions were analyzed using Proc Mixed repeated measures with unstructured model (SAS 9.1). Both overall effects and effects by time were analyzed. A nonlinear regression was used to model the release of fluorescein, with and without N, using the Gompertz–Lay equation [44]. In the third experiment, the data for fungal counts and FDA activity met the assumptions of normality and homoscedasticity and were analyzed with analysis of variance (ANOVA). In case of significant effects, multiple mean comparisons were done using Fisher’s least significant difference (LSD). All analyses were performed using SAS 9.2 software (raw data used in all analyses are available online in the Supplementary Materials file). Regression analysis was performed on the fluorescein production and the fungal colony forming units using Sigma Plot 10.0.
3. Results
3.1. Biochar Composition Differed by VM Content and Feedstock
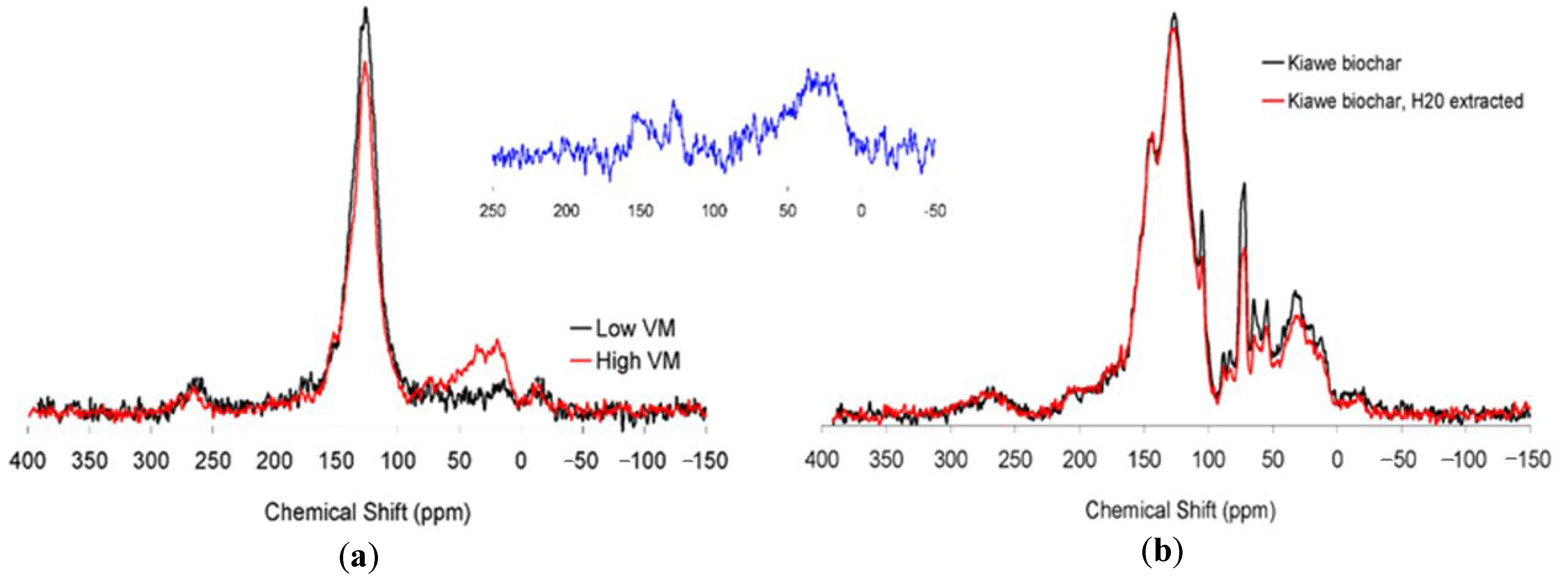
Our 13C CPMAS NMR data indicate that biochar chemical and structural composition varies depending upon both VM content and feedstock. Figure 1a shows significant structural differences between the 23% and 7% VM corncob biochar. We performed a mathematical subtraction of the 7% VM corncob biochar spectrum from the 23% VM corncob biochar (inset in Figure 1a), showing that the difference largely consists of alkyl carbon (0–45 ppm), oxygen-substituted alkyl carbon (45–95 ppm), and phenolic compounds (145–165 ppm). In comparison to the higher VM biochar, the low VM corncob biochar contained more condensed aromatic carbon (120–135 ppm) and relatively less alkyl C (0–45 ppm). Similar shifts in structure between high and low temperature maize charcoal were reported by Wang et al. (2015), which we attribute to VM content. The structure of the water-extractable matter of the 23% VM corncob biochar was similar to the non-extracted biochar (Table 2). In contrast, the water-extractable matter from the 7% corncob biochar was dominated by alkyl, amide, and carboxyl C.
Biochar chemistry also differed by feedstock. Corncob biochar had substantially higher aromatic and carboxyl C than Kiawe biochar of equivalent VM content (Table 2). A portion of the VM content is water extractable, as demonstrated by the reduction in alkyl carbon when the kiawe biochar was extracted with water.
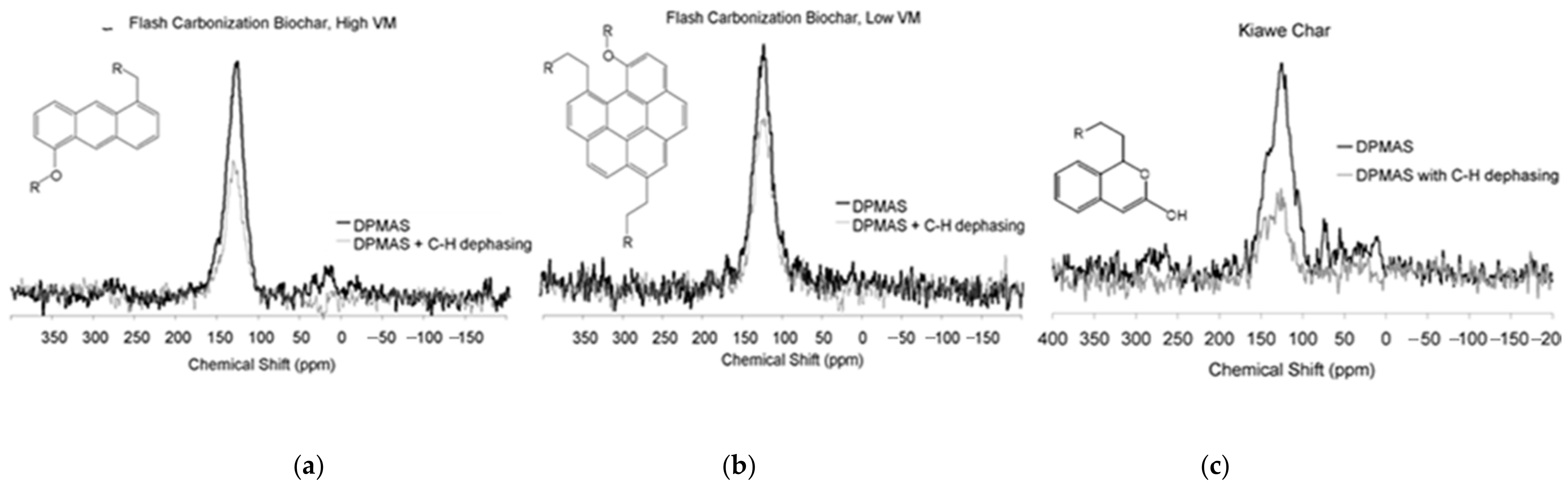
Structural differences can also be inferred from the DPMAS NMR measurements. Using the method of Solum et al. [36,37], we estimate that the average number of carbon atoms comprising an aromatic domain in the biochar was approximately 14 for the 23% VM corncob biochar (Figure 2a), approximately 22 for the 7% VM corncob biochar (Figure 2b), and approximately 8 for the kiawe biochar (Figure 2c). Another structural difference was the alkyl sidechains that serve as cross-linkages between neighboring aromatic clusters (R). Corncob biochars had sidechains of two alkyl carbons, while kiawe biochar sidechains were four carbons in length, on average. The proposed hypothetical chemical structures for each of the biochars are shown in Figure 2, which synthesize all structural information from NMR data. The 7% VM corncob biochar structure showed the most extensive pyrolytic loss of alkyl C suggesting that it experienced greater pyrolysis severity than the higher VM biochars.
3.2. Biochar Effects on Microbial N and C Dynamics
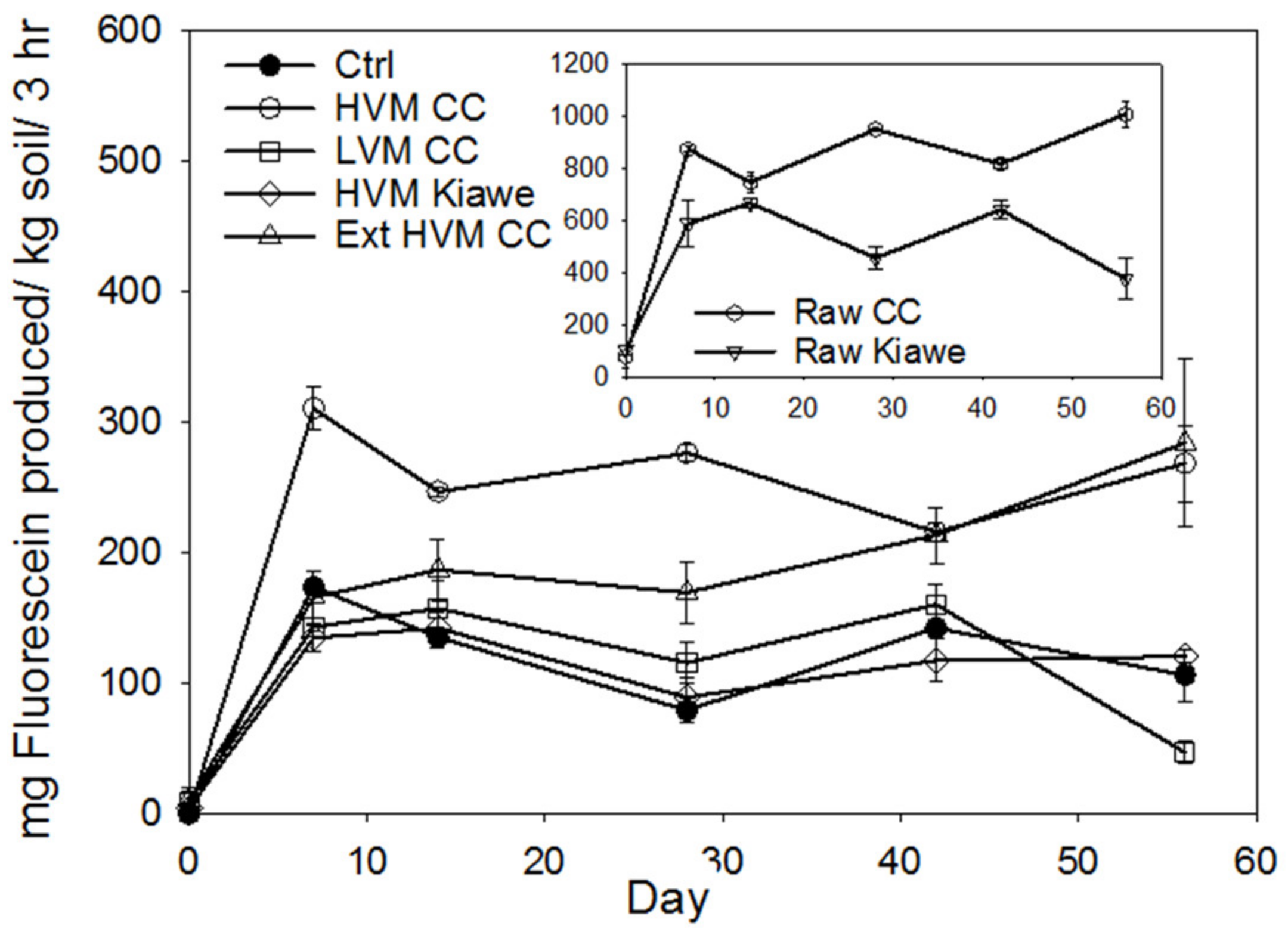
The FDA-derived hydrolytic activity was inversely proportional to VM content of the corncob biochar materials (Figure 3). The addition of raw corncob husk materials enhanced FDA activity almost 7-fold and the 34% VM corncob biochar resulted in a 2.5-fold increase in activity. This was followed by 23% VM corncob biochar, which also doubled the FDA activity. In contrast, 7% VM corncob biochar did not significantly affect the FDA activity relative to the no-biochar control. The hydrolytic enzyme activity in the soils with acetone-extracted 23% VM corncob biochar was significantly less than the non-extracted 23% VM corncob biochar during the first month of incubation, but the activity increased to that of the non-extracted biochar during the second month of incubation. We also observed a feedstock effect, where contrary to our hypothesis, the 23% VM content in kiawe biochar did not stimulate FDA-derived hydrolytic enzyme activity despite the kiawe biochar’s lower aromaticity.
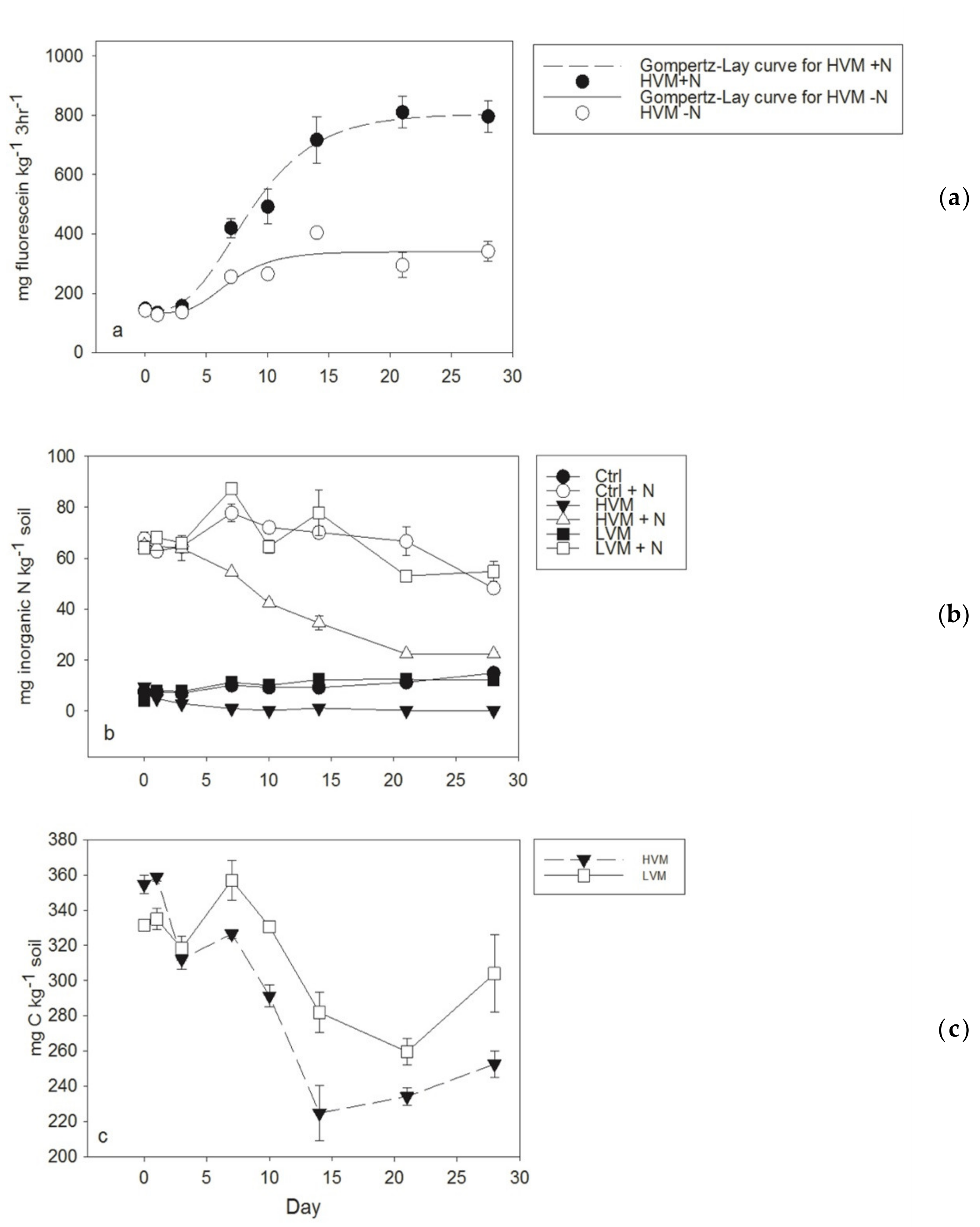
In the second incubation, we found four indirect lines of evidence showing the bioavailability of the high VM corncob biochar. First, the enhancement of FDA activity by the high VM corncob biochar appeared to be limited by N, rather than C, as evidenced by the 4-fold increase in FDA activity when N was applied in combination with the 34% VM corncob (Figure 4a). Secondly, we observed a significant decline in inorganic N (NH4+, NO3-) in soils receiving 34% VM corncob indicating N immobilization, unlike the low VM biochar where soil inorganic N did not change significantly during the incubation period in comparison to the control (Figure 4b). This observation was combined with a decline in water extractable C, despite the enhancement of this C pool with the addition of high VM biochar (Figure 4c). Thirdly, the stimulation of FDA activity, used as a measure of total microbial activity, follows a typical microbial growth pattern upon the addition of 34% VM corncob (Figure 4a) wherein exponential growth gave way to steady state growth after two weeks. Finally, the acetone-extracted biochar did not enhance FDA activity during the first month of incubation in major contrast to the non-extracted biochar, which stimulated activity within the first week (Figure 3). However, by the sixth week of incubation, their effect on FDA activities was the same, which suggests that acetone extracts can contain highly labile compounds from the high VM biochar while the insoluble fraction is somewhat more recalcitrant and not readily available within the first month.
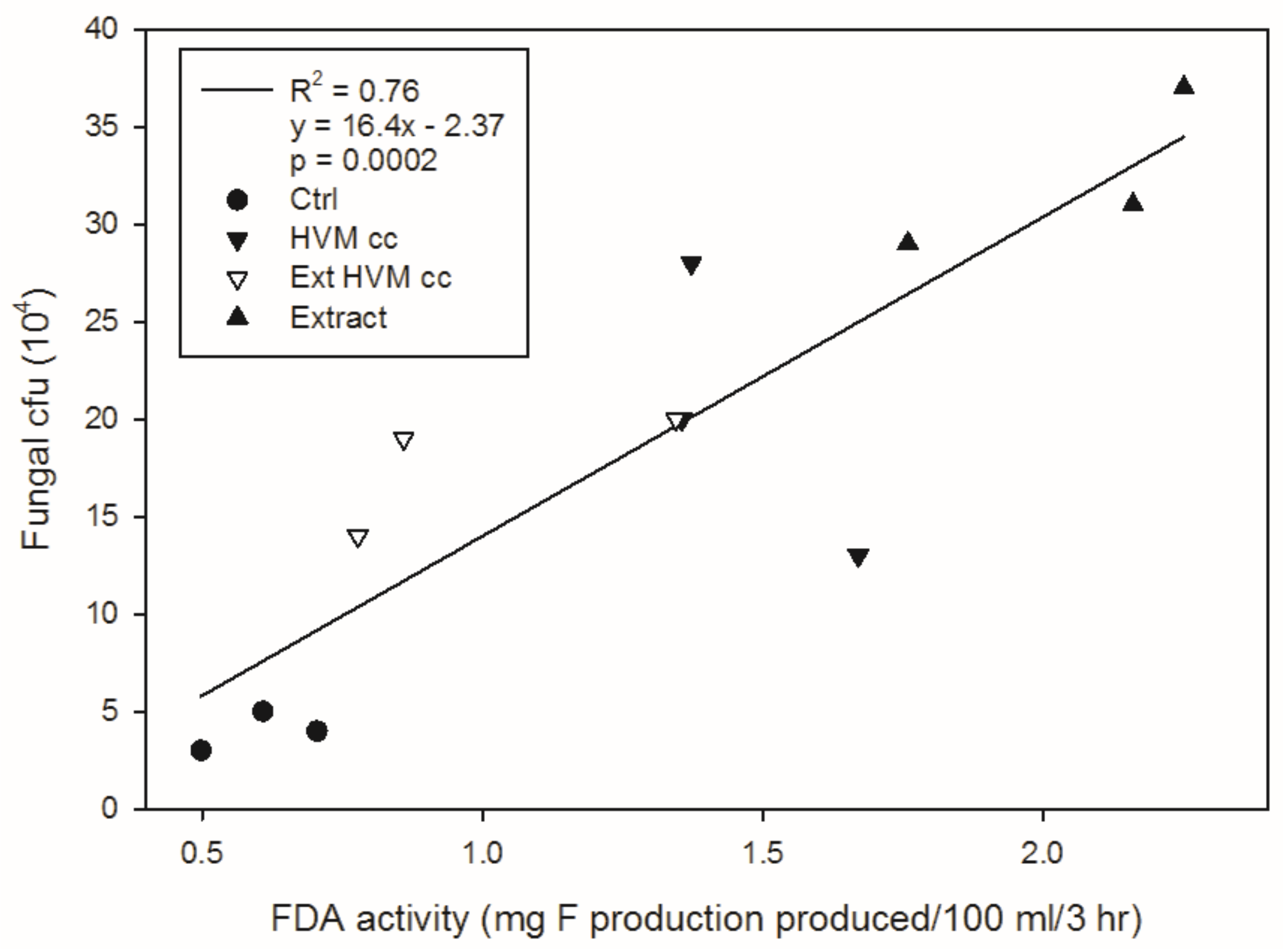
In the third incubation, we measured the number of fungal colonies and FDA activity for acetone extract and the acetone-extractable fractions of the 23% VM corncob biochar. Fungal counts and FDA activity were greatest in the acetone extracts taken from the 23% VM corncob biochar (Figure 5). This was followed by the non-extracted 23% VM corncob biochar and the acetone-extracted biochar, whereas the no-biochar control had a significantly lower abundance of fungal colonies and FDA activity than all the biochar treatments.
4. Discussion
Our findings demonstrated notable differences between the chemical structures of high and low VM corncob biochars with the high VM biochars containing greater amounts of alkyl, oxygen-containing compounds, and phenolic carbons than low VM biochars. These results support previous observations reported by Meszaros et al. [29] who showed that high VM biochars contained carbohydrate, aliphatic, oxygen-containing compounds, and aromatic (e.g., phenolic) compounds, and Bourke et al. [45] observed a loss of oxygen functionality of heat-treated flash carbonized low VM corncob biochar as the result of dehydration, decarbonylation, decarboxylation, and increased condensation and aromatic growth. Keiluweit et al. [15] also reported a relationship between decreasing VM content, pyrolysis severity, and changes in the chemical composition of biochar. By referencing the regression analysis between carbon content and production temperature provided in Figure 3b of Antal and Gronli [23], we estimate that the 7% VM corncob biochar was produced around 550°C; 23% VM corncob at 375°C; and 23% VM kiawe, 325°C. In agreement with Brewer et al. [46], our DPMAS NMR analysis indicates a relationship between temperature and the cluster size of the condensed aromatic ring structures.
The degree of pyrolysis has a significant effect on biochar stability [4,7]. Less carbonized biochars with a high VM content contain a relatively higher alkyl C: aromatic C ratio than more carbonized biochars with low VM. Indeed, previous studies demonstrated that the highly aromatic biochars with lower alkyl carbon content were less susceptible to microbial degradation [9,14]. In two studies, Bruun et al. [47,48] found that rapid pyrolysis can result in incomplete carbonization leaving labile C sources that are subject to C mineralization and greater levels of microbial biomass and immobilization of N. Biochar with high VM content (40%) has also been linked to increases in short-term C mineralization, which subsides with time [49]. Our direct polarization NMR data support the hypothesis that VM content is related to the presence or absence of bioavailable C.
Our results show that biochars for a given feedstock differed in their labile carbon pools according to their VM content or degree of thermal alteration, in agreement with previous studies [9,14,18,19,48,50,51,52]. Our incubations provide evidence showing elevated hydrolytic enzyme activity in a soil amended with high VM corncob biochar, which appeared to be N-limited rather than C-limited. Wu et al. [53] also reported that microorganisms were N limited when biochar was added to soil, and N amendments increased soil phospholipid fatty acid concentrations. Other researchers found that additions of biochar enhanced the abundance of bacterial families involved in N cycling [52,53,54], and increased activities of dehydrogenase [51,53,55,56,57], and other C-cycling enzymes [58,59]. Similar to our findings, Gundale and De Luca [11] also showed that the addition of a labile N substrate relieved N limitations and enhanced carbon dioxide respiration in soils amended with biochar produced at 350 °C (low temperature) where we expect VM content to be high.
Without N additions, we observed a depression in inorganic N levels in soils amended with high VM corncob biochar. Our previous work has shown that high VM content in macadamia nutshell biochar strongly reduced nitrogen availability in soil [13]. Other researchers have also observed the immobilization of N in soils receiving low-temperature [11,60,61,62] and high VM biochars [12], in addition to increases in nitrous oxide and carbon dioxide emissions [63] and reductions in plant N concentrations [64]. Nitrogen immobilization can also increase in soils amended with both crop residues and biochar [65], with biochar inducing the preferential utilization of litter by microbes [66]. Mukherjee and Zimmerman [67] reported that high VM charcoal initially released more dissolved organic C, N, and P than their low VM counterparts, which serves as a potential labile C source to stimulate microbial growth and the promotion of N immobilization under N limiting conditions. On the other hand, low VM biochar can enhance N mineralization [68]. Additionally, the effects of VM are not limited to N transformations. Guerena et al. [69] found that the removal of VM with an acetone extraction in low-temperature biochar resulted in an increase in plant growth, which was attributed to phytotoxicity effects of the chemical constituents in the VM rather than bioavailable C effects on N availability. There are other examples of chemicals in VM that can have toxic effects on soil microbes [70] and plant growth [71].
We used the enumeration of fungal colonies and FDA activity as indicators of microbial response to bioavailable C in VM, and we found that biological activity decreased upon a series of acetone extractions aimed to isolate the VM fraction. Previous gas chromatography mass spectroscopy (GC-MS) analyses demonstrated that high VM corncob biochar contained a range of phenolic compounds [72]. High VM biochars are known to contain recondensed tarry vapors on their surfaces, in addition to partially carbonized compounds [29,70,73], and the condensed tarry vapors, or pyroligneous acid, have been demonstrated to strongly enhance the microbial growth [74]. In a recent review of the literature, there is ample evidence showing chemical constituents in VM stimulate microbial activity [70]. However, as observed in this study and others, the biological effects of biochar are temporal and appear to not persist [61,70,72,75,76,77].
Volatile matter content explained pyrolysis severity, chemical diversity, and their relationship to bioavailable C and N dynamics in biochars made from a single feedstock (corncob in this case), but not when comparing two different feedstocks of similar VM contents. The 13C NMR analysis of the kiawe biochar showed less thermal alteration and higher alkyl C suggesting more potentially bioavailable C. Rittl et al. [78] observed differences in the aliphatic compounds among biochar feedstocks produced at the same temperature (380 °C) using 13C NMR spectroscopy, and related a higher proportion of aliphatic compounds to enhanced C mineralization. Therefore, we might expect that kiawe charcoal would have the highest bioavailability. However, our previous research demonstrated that the high VM content kiawe biochar did not contain any detectable acetone-extractable chemical compounds with GC-MS nor did it affect corn growth by reducing plant available N, in contrast to the high VM corncob biochar [72]. In agreement with our results, Hamer et al. [9] observed that wood biochar derived at 250 °C had approximately three times less carbon mineralization during a two-month incubation than corncob biochar produced at the same temperature. On the other hand, Anders et al. [79] reported that biochar derived from wood-chip feedstocks did not affect soil microbial biomass, while Dempster et al. [75] found that eucalyptus biochar decreased microbial biomass and net N mineralization. In another study, the microbial effects of wood-based biochars were delayed in comparison to biochars derived from crop residue feedstocks [80]. In support of our observations, Rajkovich et al. [31] reported that VM content did not correlate with N availability across a range of feedstocks, but that within a given feedstock, VM better matched N uptake. In a more recent meta-analysis, neither pyrolysis conditions nor feedstock correlated well with soil N in biochar amended soils [81]. Therefore, while VM content may be indicative of biochar stability and bioavailable C4, the chemical heterogeneity of the VM component across feedstocks suggest it is less reliable when predicting the behavior of biochars across feedstock types [7]. The work of Spokas [5] and Dutta et al. [70] demonstrates that VM is characterized by high chemical heterogeneity, and that this heterogeneity impacts microbially driven biochemical reactions in the soil.
As other studies show, our results confirm that there is a need for a more complete characterization of VM as part of a quality assessment of the biochar. Ultimately, an analysis is needed to identify the properties of the extractable component controlling microbial activity in relation to N transformations, and an assessment of the toxicity or enzyme activity of extractable material before applying biochar to soil. However, our findings only highlight one aspect of the complexities that control the stability of biochar C and other soil organic carbon, where the new paradigm is the notion that stability is as much a function of the soil’s physical, chemical, and biological properties (and dynamics therein) as it is about the biochar’s inherent properties. [8,15].
5. Conclusions
We have shown that VM content is related to changes in chemical structure and composition of biochar due to increasing pyrolysis severity for a corncob biochar produced by flash carbonization. Structurally, the VM content of biochars appears to be comprised of alkyl carbons, oxygen-substituted alkyl carbons, and phenolics. Despite VM content being an easily measurable property that denotes major structural and molecular differences within one feedstock, its interpretation is limited when making comparisons among different feedstocks. Two biochars may have the same VM content but differ substantially in chemical composition with differential effects on biological and biochemical properties of soils. These results reinforce the notion that biochars are complex materials, and that a rapid measurement such as VM content is too coarse to differentiate chemical structure and composition across feedstocks and production methods.
Supplementary Materials
Author Contributions
Conceptualization, T.M.M. and J.L.D.; methodology, T.M.M., J.L.D., and W.C.H.; software, T.M.M. and W.C.H.; validation, T.M.M., J.L.D., and W.C.H.; formal analysis, T.M.M. and W.C.H.; investigation, T.M.M.; resources, J.L.D., and W.C.H..; data curation, T.M.M. and W.C.H.; writing—original draft preparation, T.M.M.; writing—review and editing, J.L.D. and W.C.H.; visualization, T.M.M.; supervision, J.L.D.; project administration, J.L.D.; funding acquisition, J.L.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by United States Department of Agriculture, Tropical and Sub-tropical Agriculture Research program (USDA-TSTAR) grant number 2008-34135-19412.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Materials.
Acknowledgments
We would like to honor the lives and careers of Goro Uehara and Michael J. Antal Jr., who helped design our study, but passed away before the publication of this manuscript. We would also like to acknowledge technical feedback provided by Hue Nguyen, Russell Yost, Qing Li, and Sonia Campbell.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shindo, H. Elementary composition, humus composition, and decomposition in soil of charred grassland plants. Soil Sci. Plant Nutr. 1991, 37, 651–657. [Google Scholar] [CrossRef]
- Bird, M.I.; Moyo, C.; Veenendaal, E.M.; Lloyd, J.; Frost, P. Stability of elemental carbon in a savanna soil. Glob. Biogeochem. Cycles 1999, 13, 923–932. [Google Scholar] [CrossRef]
- Lehmann, J.; Solomon, D.; Kinyangi, J.; Dathe, L.; Wirick, S.; Jacobsen, C. Spatial Complexity of Soil Organic Matter Forms at Nanometer Scales. Nat. Geosci. 2008, 1, 238–242. [Google Scholar] [CrossRef]
- Zimmerman, A.R. Abiotic and Microbial Oxidation of Laboratory-Produced Black Carbon (Biochar). Environ. Sci. Technol. 2010, 44, 1295–1301. [Google Scholar] [CrossRef]
- Spokas, K.A. Review of the stability of biochar in soils: Predictability of O:C molar ratios. Carbon Manag. 2010, 1, 289–303. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Xu, Y.; Liu, G.; Liu, Q.; Zhu, J.; Tu, C.; Amonette, J.E.; Cadisch, G.; Yong, J.W.H.; Hu, S. Impact of biochar application on nitrogen nutrition of rice, greenhouse-gas emissions and soil organic carbon dynamics in two paddy soils of China. Plant Soil 2013, 370, 527–540. [Google Scholar] [CrossRef]
- Leng, L.; Huang, H.; Li, H.; Li, J.; Zhou, W. Biochar stability assessment methods: A review. Sci. Total Environ. 2019, 647, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Hansel, C.M.; Kaiser, C.; Kleber, M.; Maher, K.; Manzoni, S.; Nunan, N.; Reichstein, M.; Schimel, J.P.; Torn, M.S.; et al. Persistence of soil organic carbon caused by functional complexity. Nat. Geosci. 2020, 13, 529–534. [Google Scholar] [CrossRef]
- Hamer, U.; Marschner, B.; Brodowski, S.; Amelung, W. Interactive priming of black carbon and glucose mineralisation. Org. Geochem. 2004, 35, 823–830. [Google Scholar] [CrossRef]
- Wang, J.; Xiong, Z.; Kuzyakov, Y. Biochar stability in soil: Meta-analysis of decomposition and priming effects. GCB Bioenergy 2016, 8, 512–523. [Google Scholar] [CrossRef] [Green Version]
- Gundale, M.J.; DeLuca, T.H. Charcoal effects on soil solution chemistry and growth of Koeleria macrantha in the ponderosa pine/Douglas-fir ecosystem. Biol. Fertil. Soils 2006, 43, 303–311. [Google Scholar] [CrossRef]
- Rondon, M.A.; Lehmann, J.; Ramirez, J.; Hurtado, M. Biological Nitrogen Fixation by Common Beans (Phaseolus Vulgaris L.) Increases with Bio-Char Additions. Biol. Fertil. Soils 2007, 43, 699–708. [Google Scholar] [CrossRef]
- Deenik, J.L.; McClellan, T.; Uehara, G.; Antal, M.J.; Campbell, S. Charcoal Volatile Matter Content Influences Plant Growth and Soil Nitrogen Transformations. Soil Sci. Soc. Am. J. 2010, 74, 1259–1270. [Google Scholar] [CrossRef]
- Baldock, J.A.; Smernik, R.J. Chemical composition and bioavailability of thermally altered Pinus resinosa (Red pine) wood. Org. Geochem. 2002, 33, 1093–1109. [Google Scholar] [CrossRef]
- Keiluweit, M.; Nico, P.S.; Johnson, M.G.; Kleber, M. Dynamic Molecular Structure of Plant Biomass-Derived Black Carbon (Biochar). Environ. Sci. Technol. 2010, 44, 1247–1253. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Kim, J.-Y.; Cho, T.-S.; Choi, J.W. Influence of pyrolysis temperature on physicochemical properties of biochar obtained from the fast pyrolysis of pitch pine (Pinus rigida). Bioresour. Technol. 2012, 118, 158–162. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, W.; Liang, G.; Song, D.; Zhang, X. Characteristics of maize biochar with different pyrolysis temperatures and its effects on organic carbon, nitrogen and enzymatic activities after addition to fluvo-aquic soil. Sci. Total. Environ. 2015, 538, 137–144. [Google Scholar] [CrossRef]
- Chen, D.; Yu, X.; Song, C.; Pang, X.; Huang, J.; Li, Y. Effect of pyrolysis temperature on the chemical oxidation stability of bamboo biochar. Bioresour. Technol. 2016, 218, 1303–1306. [Google Scholar] [CrossRef]
- Jindo, K.; Sonoki, T. Comparative Assessment of Biochar Stability Using Multiple Indicators. Agronomy 2019, 9, 254. [Google Scholar] [CrossRef] [Green Version]
- Harvey, O.R.; Myers-Pigg, A.N.; Kuo, L.-J.; Singh, B.; Kuehn, K.A.; Louchouarn, P. Discrimination in Degradability of Soil Pyrogenic Organic Matter Follows a Return-On-Energy-Investment Principle. Environ. Sci. Technol. 2016, 50, 8578–8585. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.W.I.; Noack, A.G. Black carbon in soils and sediments: Analysis, distribution, implications, and current challenges. Glob. Biogeochem. Cycles 2000, 14, 777–793. [Google Scholar] [CrossRef]
- Uchimiya, M.; Hiradate, S.; Antal, J.M.J. Influence of Carbonization Methods on the Aromaticity of Pyrogenic Dissolved Organic Carbon. Energy Fuels 2015, 29, 2503–2513. [Google Scholar] [CrossRef]
- Antal, M.J.; Grønli, M. The Art, Science, and Technology of Charcoal Production†. Ind. Eng. Chem. Res. 2003, 42, 1619–1640. [Google Scholar] [CrossRef]
- Yua, F.; Steele, P.H.; Ruan, R. Microwave Pyrolysis of Corn Cob and Characteristics of the Pyrolytic Chars. Energy Sources Part A Recover. Util. Environ. Eff. 2010, 32, 475–484. [Google Scholar] [CrossRef]
- Ishimaru, K.; Hata, T.; Bronsveld, P.; Meier, D.; Imamura, Y. Spectroscopic analysis of carbonization behavior of wood, cellulose and lignin. J. Mater. Sci. 2007, 42, 122–129. [Google Scholar] [CrossRef]
- Várhegyi, G.; Szabo, P.; Till, F.; Zelei, B.; Antal, M.J.; Dai, X. TG, TG-MS, and FTIR Characterization of High-Yield Biomass Charcoals. Energy Fuels 1998, 12, 969–974. [Google Scholar] [CrossRef] [Green Version]
- Antal, M.J.; Allen, S.G.; Dai, X.; Shimizu, B.; Tam, M.S.; Grønli, M. Attainment of the Theoretical Yield of Carbon from Biomass. Ind. Eng. Chem. Res. 2000, 39, 4024–4031. [Google Scholar] [CrossRef]
- Parikh, J.; Channiwala, S.; Ghosal, G. A correlation for calculating elemental composition from proximate analysis of biomass materials. Fuel 2007, 86, 1710–1719. [Google Scholar] [CrossRef]
- Mészáros, E.; Jakab, E.; Várhegyi, G.; Bourke, J.; Manley-Harris, M.; Nunoura, T.; Antal, M.J. Do All Carbonized Charcoals Have the Same Chemical Structure? 1. Implications of Thermogravimetry−Mass Spectrometry Measurements. Ind. Eng. Chem. Res. 2007, 46, 5943–5953. [Google Scholar] [CrossRef]
- Enders, A.; Hanley, K.; Whitman, T.; Joseph, S.; Lehmann, J. Characterization of biochars to evaluate recalcitrance and agronomic performance. Bioresour. Technol. 2012, 114, 644–653. [Google Scholar] [CrossRef]
- Rajkovich, S.; Enders, A.; Hanley, K.; Hyland, C.; Zimmerman, A.R.; Lehmann, J. Corn growth and nitrogen nutrition after additions of biochars with varying properties to a temperate soil. Biol. Fertil. Soils 2012, 48, 271–284. [Google Scholar] [CrossRef]
- Ameloot, N.; Graber, E.R.; Verheijen, F.G.A.; De Neve, S. Interactions between biochar stability and soil organisms: Review and research needs. Eur. J. Soil Sci. 2013, 64, 379–390. [Google Scholar] [CrossRef]
- Antal, J.M.J.; Mochidzuki, A.K.; Paredes, L.S. Flash Carbonization of Biomass. Ind. Eng. Chem. Res. 2003, 42, 3690–3699. [Google Scholar] [CrossRef]
- ASTM International. ASTM D1762-84. In Standard Test Method for Chemical Analysis of Wood Charcoal; ASTM International: West Conshohocken, PA, USA, 2013. [Google Scholar]
- Smernik, R.J.; Oades, J.M. Solid-state 13C-NMR dipolar dephasing experiments for quantifying protonated and non-protonated carbon in soil organic matter and model systems. Eur. J. Soil Sci. 2001, 52, 103–120. [Google Scholar] [CrossRef]
- Solum, M.S.; Pugmire, R.J.; Grant, D.M. Carbon-13 solid-state NMR of Argonne-premium coals. Energy Fuels 1989, 3, 187–193. [Google Scholar] [CrossRef]
- Solum, M.S.; Sarofim, A.F.; Pugmire, R.J.; Fletcher, T.H.; Zhang†, H. 13C NMR Analysis of Soot Produced from Model Compounds and a Coal. Energy Fuels 2001, 15, 961–971. [Google Scholar] [CrossRef]
- Green, V.; Stott, D.E.; Diack, M. Assay for fluorescein diacetate hydrolytic activity: Optimization for soil samples. Soil Biol. Biochem. 2006, 38, 693–701. [Google Scholar] [CrossRef]
- Heal, O.W.; MacLean, S.F. Comparative Productivity in Ecosystems—Secondary Productivity. In Unifying Concepts in Ecology; van Dobben, W.H., Lowe-McConnell, R.H., Eds.; Springer: Dordrecht, The Netherlands, 1975; pp. 89–108. [Google Scholar]
- Chintala, R.; Schumacher, T.E.; Kumar, S.; Malo, D.D.; Rice, J.A.; Bleakley, B.; Chilom, G.; Clay, D.E.; Julson, J.L.; Papiernik, S.K.; et al. Molecular characterization of biochars and their influence on microbiological properties of soil. J. Hazard. Mater. 2014, 279, 244–256. [Google Scholar] [CrossRef]
- Schumacher, T.E.; Eynard, A.; Chintala, R. Rapid cost-effective analysis of microbial activity in soils using modified fluorescein diacetate method. Environ. Sci. Pollut. Res. 2014, 22, 4759–4762. [Google Scholar] [CrossRef] [PubMed]
- Ghani, A.; Dexter, M.; Perrott, K. Hot-water extractable carbon in soils: A sensitive measurement for determining impacts of fertilisation, grazing and cultivation. Soil Biol. Biochem. 2003, 35, 1231–1243. [Google Scholar] [CrossRef]
- Mulvaney, R.L. Nitrogen-Inorganic Forms. In Methods of Soil Analysis: Part 3 Chemical Methods, 5.3; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpoor, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA: Madison, WI, USA, 1996; pp. 1123–1184. [Google Scholar]
- Lay, J.J.; Li, Y.Y.; Noike, T.; Endo, J.; Ishimoto, S. Analysis of Environmental Factors Affecting Methane Production from High-Solids Organic Wastes. Water Sci. Technol. 1997, 36, 493–500. [Google Scholar] [CrossRef]
- Bourke, J.; Manley-Harris, M.; Fushimi, C.; Dowaki, K.; Nunoura, T.; Antal, M.J. Do All Carbonized Charcoals Have the Same Chemical Structure? 2. A Model of the Chemical Structure of Carbonized Charcoal†. Ind. Eng. Chem. Res. 2007, 46, 5954–5967. [Google Scholar] [CrossRef]
- Brewer, C.E.; Schmidt-Rohr, K.; Satrio, J.A.; Brown, R.C. Characterization of biochar from fast pyrolysis and gasification systems. Environ. Prog. Sustain. Energy 2009, 28, 386–396. [Google Scholar] [CrossRef]
- Bruun, E.W.; Hauggaard-Nielsen, H.; Ibrahim, N.; Egsgaard, H.; Ambus, P.; Jensen, P.A.; Dam-Johansen, K. Influence of fast pyrolysis temperature on biochar labile fraction and short-term carbon loss in a loamy soil. Biomass Bioenergy 2011, 35, 1182–1189. [Google Scholar] [CrossRef]
- Bruun, E.W.; Ambus, P.; Egsgaard, H.; Hauggaard-Nielsen, H. Effects of slow and fast pyrolysis biochar on soil C and N turnover dynamics. Soil Biol. Biochem. 2012, 46, 73–79. [Google Scholar] [CrossRef]
- Smith, J.L.; Collins, H.P.; Bailey, V.L. The effect of young biochar on soil respiration. Soil Biol. Biochem. 2010, 42, 2345–2347. [Google Scholar] [CrossRef]
- Czimczik, C.I.; Masiello, C.A. Controls on black carbon storage in soils. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Ameloot, N.; De Neve, S.; Jegajeevagan, K.; Yildiz, G.; Buchan, D.; Funkuin, Y.N.; Prins, W.; Bouckaert, L.; Sleutel, S. Short-term CO2 and N2O emissions and microbial properties of biochar amended sandy loam soils. Soil Biol. Biochem. 2013, 57, 401–410. [Google Scholar] [CrossRef]
- Anderson, C.R.; Condron, L.M.; Clough, T.J.; Fiers, M.; Stewart, A.; Hill, R.A.; Sherlock, R.R. Biochar induced soil microbial community change: Implications for biogeochemical cycling of carbon, nitrogen and phosphorus. Pedobiology 2011, 54, 309–320. [Google Scholar] [CrossRef]
- Wu, H.; Zeng, G.; Liang, J.; Chen, J.; Xu, J.; Dai, J.; Li, X.; Chen, M.; Xu, P.; Zhou, Y.; et al. Responses of bacterial community and functional marker genes of nitrogen cycling to biochar, compost and combined amendments in soil. Appl. Microbiol. Biotechnol. 2016, 100, 8583–8591. [Google Scholar] [CrossRef]
- Ducey, T.F.; Ippolito, J.A.; Cantrell, K.B.; Novak, J.M.; Lentz, R.D. Addition of activated switchgrass biochar to an aridic subsoil increases microbial nitrogen cycling gene abundances. Appl. Soil Ecol. 2013, 65, 65–72. [Google Scholar] [CrossRef]
- Mierzwa-Hersztek, M.; Wolny-Koładka, K.; Gondek, K.; Gałązka, A.; Gawryjołek, K. Effect of Coapplication of Biochar and Nutrients on Microbiocenotic Composition, Dehydrogenase Activity Index and Chemical Properties of Sandy Soil. Waste Biomass Valorization 2019, 11, 3911–3923. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Liu, X.; Zheng, J.; Zhang, B.; Lu, H.; Chi, Z.; Pan, G.; Li, L.; Zheng, J.; Zhang, X.; et al. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl. Soil Ecol. 2013, 71, 33–44. [Google Scholar] [CrossRef]
- Gao, S.; Hoffman-Krull, K.; DeLuca, T.H. Soil biochemical properties and crop productivity following application of locally produced biochar at organic farms on Waldron Island, WA. Biogeochemestry 2017, 136, 31–46. [Google Scholar] [CrossRef]
- Liao, N.; Li, Q.; Zhang, W.; Zhou, G.; Ma, L.; Min, W.; Ye, J.; Hou, Z. Effects of biochar on soil microbial community composition and activity in drip-irrigated desert soil. Eur. J. Soil Biol. 2016, 72, 27–34. [Google Scholar] [CrossRef]
- Wu, D.; Senbayram, M.; Zang, H.; Ugurlar, F.; Aydemir, S.; Brüggemann, N.; Kuzyakov, Y.; Bol, R.; Blagodatskaya, E. Effect of biochar origin and soil pH on greenhouse gas emissions from sandy and clay soils. Appl. Soil Ecol. 2018, 129, 121–127. [Google Scholar] [CrossRef]
- Nelissen, V.; Rütting, T.; Huygens, D.; Ruysschaert, G.; Boeckx, P. Temporal evolution of biochar’s impact on soil nitrogen processes—a15N tracing study. GCB Bioenergy 2015, 7, 635–645. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Xu, C.-Y.; Tahmasbian, I.; Che, R.; Xu, Z.; Zhou, X.; Wallace, H.M.; Bai, S.H. Effects of biochar on soil available inorganic nitrogen: A review and meta-analysis. Geoderma 2017, 288, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Y.; Chen, L.; Sun, F.L.; Luo, X.X.; Wang, H.F.; Liu, G.; Xu, Z.H.; Jiang, Z.X.; Pan, B.; Zheng, H. Effects of adding biochar on the properties and nitrogen bioavailability of an acidic soil. Eur. J. Soil Sci. 2017, 68, 559–572. [Google Scholar] [CrossRef]
- Subedi, R.; Taupe, N.; Pelissetti, S.; Petruzzelli, L.; Bertora, C.; Leahy, J.J.; Grignani, C. Greenhouse gas emissions and soil properties following amendment with manure-derived biochars: Influence of pyrolysis temperature and feedstock type. J. Environ. Manag. 2016, 166, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Hagner, M.; Kemppainen, R.; Jauhiainen, L.; Tiilikkala, K.; Setälä, H. The effects of birch (Betula spp.) biochar and pyrolysis temperature on soil properties and plant growth. Soil Tillage Res. 2016, 163, 224–234. [Google Scholar] [CrossRef]
- Prayogo, C.; Jones, J.E.; Baeyens, J.; Bending, G.D. Impact of biochar on mineralisation of C and N from soil and willow litter and its relationship with microbial community biomass and structure. Biol. Fertil. Soils 2013, 50, 695–702. [Google Scholar] [CrossRef]
- Cui, J.; Ge, T.; Kuzyakov, Y.; Nie, M.; Fang, C.; Tang, B.; Zhou, C. Interactions between biochar and litter priming: A three-source 14C and δ13C partitioning study. Soil Biol. Biochem. 2017, 104, 49–58. [Google Scholar] [CrossRef]
- Mukherjee, A.; Zimmerman, A.R. Organic carbon and nutrient release from a range of laboratory-produced biochars and biochar–soil mixtures. Geoderma 2013, 193, 122–130. [Google Scholar] [CrossRef]
- Zhao, R.; Wu, J.; Jiang, C.; Liu, F. Effects of biochar particle size and concomitant nitrogen fertilization on soil microbial community structure during the maize seedling stage. Environ. Sci. Pollut. Res. 2020, 27, 13095–13104. [Google Scholar] [CrossRef]
- Güereña, D.T.; Lehmann, J.; Thies, J.E.; Enders, A.; Karanja, N.; Neufeldt, H. Partitioning the contributions of biochar properties to enhanced biological nitrogen fixation in common bean (Phaseolus vulgaris). Biol. Fertil. Soils 2015, 51, 479–491. [Google Scholar] [CrossRef]
- Dutta, T.; Kwon, E.E.; Bhattacharya, S.S.; Jeon, B.-H.; Deep, A.; Uchimiya, M.; Kim, K.-H. Polycyclic aromatic hydrocarbons and volatile organic compounds in biochar and biochar-amended soil: A review. GCB Bioenergy 2016, 9, 990–1004. [Google Scholar] [CrossRef]
- Backer, R.; Ghidotti, M.; Schwinghamer, T.D.; Saeed, W.; Grenier, C.; Dion-Laplante, C.; Fabbri, D.; Dutilleul, P.; Seguin, P.; Smith, D.L. Getting to the root of the matter: Water-soluble and volatile components in thermally-treated biosolids and biochar differentially regulate maize (Zea mays) seedling growth. PLoS ONE 2018, 13, e0206924. [Google Scholar] [CrossRef]
- Deenik, J.L.; Diarra, A.; Uehara, G.; Campbell, S.B.; Sumiyoshi, Y.; Antal, M.J. Charcoal Ash and Volatile Matter Effects on Soil Properties and Plant Growth in an Acid Ultisol. Soil Sci. 2011, 176, 336–345. [Google Scholar] [CrossRef]
- Spokas, K.A.; Novak, J.M.; Stewart, C.E.; Cantrell, K.B.; Uchimiya, M.; DuSaire, M.G.; Ro, K.S. Qualitative analysis of volatile organic compounds on biochar. Chemosphere 2011, 85, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Steiner, C.; Das, K.C.; Garcia, M.; Förster, B.; Zech, W. Charcoal and smoke extract stimulate the soil microbial community in a highly weathered xanthic Ferralsol. Pedobiology 2008, 51, 359–366. [Google Scholar] [CrossRef]
- Dempster, D.N.; Gleeson, D.B.; Solaiman, Z.; Jones, D.L.; Murphy, D.V. Decreased soil microbial biomass and nitrogen mineralisation with Eucalyptus biochar addition to a coarse textured soil. Plant Soil 2012, 354, 311–324. [Google Scholar] [CrossRef]
- Jones, D.L.; Rousk, J.; Edwardsjones, G.; DeLuca, T.H.; Murphy, D.V. Biochar-mediated changes in soil quality and plant growth in a three year field trial. Soil Biol. Biochem. 2012, 45, 113–124. [Google Scholar] [CrossRef]
- Rousk, J.; Dempster, D.N.; Jones, D.L. Transient biochar effects on decomposer microbial growth rates: Evidence from two agricultural case-studies. Eur. J. Soil Sci. 2013, 64, 770–776. [Google Scholar] [CrossRef]
- Rittl, T.F.; Novotny, E.H.; Balieiro, F.C.; Hoffland, E.; Alves, B.J.R.; Kuyper, T.W. Negative priming of native soil organic carbon mineralization by oilseed biochars of contrasting quality. Eur. J. Soil Sci. 2015, 66, 714–721. [Google Scholar] [CrossRef]
- Anders, E.; Watzinger, A.; Rempt, F.; Kitzler, B.; Wimmer, B.; Zehetner, F.; Stahr, K.; Zechmeister-Boltenstern, S.; Soja, G. Biochar affects the structure rather than the total biomass of microbial communities in temperate soils. Agric. Food Sci. 2013, 22, 404–423. [Google Scholar] [CrossRef] [Green Version]
- Gul, S.; Whalen, J.K.; Thomas, B.W.; Sachdeva, V.; Deng, H. Physico-chemical properties and microbial responses in biochar-amended soils: Mechanisms and future directions. Agric. Ecosyst. Environ. 2015, 206, 46–59. [Google Scholar] [CrossRef]
- Gao, S.; DeLuca, T.H.; Cleveland, C.C. Biochar additions alter phosphorus and nitrogen availability in agricultural ecosystems: A meta-analysis. Sci. Total Environ. 2019, 654, 463–472. [Google Scholar] [CrossRef]
Figure 1.
13C cross polarization magic angle spinning (CPMAS) NMR spectra of (a) high volatile matter (VM) (23%) and low VM (7%) corncob biochar and (b) 13C CPMAS NMR spectra of high VM (23%) kiawe charcoal before and after a water-extraction. Spectra are scaled to equal quantities of carbon so that peak intensities may be compared directly. The blue spectrum (inset) represents the VM, obtained by mathematical subtraction.
Figure 1.
13C cross polarization magic angle spinning (CPMAS) NMR spectra of (a) high volatile matter (VM) (23%) and low VM (7%) corncob biochar and (b) 13C CPMAS NMR spectra of high VM (23%) kiawe charcoal before and after a water-extraction. Spectra are scaled to equal quantities of carbon so that peak intensities may be compared directly. The blue spectrum (inset) represents the VM, obtained by mathematical subtraction.

Figure 2.
Quantitative 13C NMR (nuclear magnetic resonance) spectra. Proposed structure of average molecular repeating units for the (a) high VM corncob, (b) low VM corncob biochar, and (c) high VM kiawe biochar.
Figure 2.
Quantitative 13C NMR (nuclear magnetic resonance) spectra. Proposed structure of average molecular repeating units for the (a) high VM corncob, (b) low VM corncob biochar, and (c) high VM kiawe biochar.

Figure 3.
Effects of raw feedstocks, biochar, extracted biochar on fluorescein diacetate hydrolysis (FDA) enzyme activity in first incubation and Ctrl = no biochar control; HVM cc = 23% volatile matter corncob (CC) biochar; LVM cc = 7% volatile matter corncob (CC) biochar, HVM kiawe = 23% volatile matter kiawe biochar; cc-r = raw corncob husks; K-r = raw kiawe wood; Ext HVM cc = extracted 23% VM corncob (CC) biochar.
Figure 3.
Effects of raw feedstocks, biochar, extracted biochar on fluorescein diacetate hydrolysis (FDA) enzyme activity in first incubation and Ctrl = no biochar control; HVM cc = 23% volatile matter corncob (CC) biochar; LVM cc = 7% volatile matter corncob (CC) biochar, HVM kiawe = 23% volatile matter kiawe biochar; cc-r = raw corncob husks; K-r = raw kiawe wood; Ext HVM cc = extracted 23% VM corncob (CC) biochar.

Figure 4.
(a) Fluorescein diacetate (FDA) hydrolytic enzyme activity during a second one-month incubation of Leilehua soils with additions of different corncob biochar, with and without nitrogen (N) fertilization, fitted Gompertz–Lay equation to FDA hydrolytic enzyme activities (b) fluctuations in soil nitrate upon the additions of biochar, with and without nitrogen (N), in time and (c) water extractable organic carbon upon the additions of high volatile matter (HVM) vs. low volatile matter (LVM)corncob biochar volatile matter. Ctrl = no biochar control; HVM = 34% volatile matter corncob biochar; and LVM = 7% volatile matter corncob biochar.
Figure 4.
(a) Fluorescein diacetate (FDA) hydrolytic enzyme activity during a second one-month incubation of Leilehua soils with additions of different corncob biochar, with and without nitrogen (N) fertilization, fitted Gompertz–Lay equation to FDA hydrolytic enzyme activities (b) fluctuations in soil nitrate upon the additions of biochar, with and without nitrogen (N), in time and (c) water extractable organic carbon upon the additions of high volatile matter (HVM) vs. low volatile matter (LVM)corncob biochar volatile matter. Ctrl = no biochar control; HVM = 34% volatile matter corncob biochar; and LVM = 7% volatile matter corncob biochar.

Figure 5.
The relationship between fungal counts and FDA (fluorescein diacetate hydrolysis) hydrolytic enzyme activity in third incubation. Ctrl = no biochar control; HVM cc = non-extracted 23% volatile matter corncob biochar; Ext = acetone extracted biochar; Extract = acetone extract.
Figure 5.
The relationship between fungal counts and FDA (fluorescein diacetate hydrolysis) hydrolytic enzyme activity in third incubation. Ctrl = no biochar control; HVM cc = non-extracted 23% volatile matter corncob biochar; Ext = acetone extracted biochar; Extract = acetone extract.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Selected physical and chemical properties of the Leilehua soil and the biochars used as soil amendments in the incubation experiments.
Table 1.
Selected physical and chemical properties of the Leilehua soil and the biochars used as soil amendments in the incubation experiments.
| Ash | VMa | TCb | TNc | pH | Pd | K | Ca | Mg | Na | Al | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ---------------g kg−1-------------- | mg kg−1 | -------------cmolc kg−1------------ | |||||||||
| Unamended Leilehua soil | 42.8 | 1.2 | 4.7 | 2.22 | 0.09 | 0.72 | 0.52 | 0.29 | 1.61 | ||
| Incubation 1: soil amendments | |||||||||||
| Biomass | |||||||||||
| Raw corncob | 417 | ||||||||||
| Raw kiawe | 421 | ||||||||||
| Biochar | |||||||||||
| 23% VM corncob | 31 | 230 | 694 | 7.3 | |||||||
| 23% VM corncob e | ndf | nd | nd | nd | |||||||
| 7.6% VM corncob | 76 | 76 | 823 | 7.5 | |||||||
| 23% VM kiawe | nd | 230 | 784 | 6.2 | |||||||
| Incubation 2: soil amendments | |||||||||||
| Biochar | |||||||||||
| 34% VM corncob | 38 | 340 | 624 | ||||||||
| 7.6% VM corncob | 76 | 76 | 823 | 7.5 | |||||||
| Incubation 3: soil amendments | |||||||||||
| Biochar | |||||||||||
| 23% VM corncob | 31 | 230 | 694 | 7.3 | |||||||
| 23% VM corncob e | nd | nd | nd | nd | 23% VM corncob e | nd | nd | nd | nd | ||
| 23% VM corncob extract g | nd | nd | nd | nd | 23% VM corncob extract g | nd | nd | nd | nd | ||
VMa —volatile matter, TCb —total carbon, TNc— total nitrogen, Pd —extractable phosphorus (modified Truog), e acetone-extracted biochar = residual biochar remaining after extraction with acetone, ndf —not determined, g solution containing acetone extractable biochar constituents.
Table 2.
Direct polarization 13C NMR peak areas † and chemical assignments for corncob and kiawe charcoal.
Table 2.
Direct polarization 13C NMR peak areas † and chemical assignments for corncob and kiawe charcoal.
| Region | Alkyl | Amine + Methoxyl | O-Alkyl | O2- Alkyl + Alkene | Aromatic | Phenolic | Amide + Carboxyl | Ketone + Aldehyde |
|---|---|---|---|---|---|---|---|---|
| (ppm) | 0–45 | 45–60 | 60–95 | 95–110 | 110–145 | 145–165 | 165–190 | 190–215 |
| Integral areas (percentage of total spectral area) | ||||||||
| 23% VM corncob | 8.8 | 0.9 | 2.2 | 3.2 | 65.9 | 10.6 | 7.1 | 1.3 |
| H2O extract | 9.6 | 1.0 | 2.9 | 2.9 | 63.9 | 15.2 | 2.7 | 1.8 |
| 7% VM corncob | 5.8 | 1.6 | 3.2 | 3.2 | 72.3 | 9.4 | 3.9 | 0.6 |
| H2O extract | 61.4 | 38.6 | ||||||
| 23% VM kiawe | 10.1 | 3.5 | 6.7 | 7.0 | 56.7 | 11.6 | 3.0 | 1.5 |
| H2O extract | 9.8 | 2.6 | 3.7 | 5.2 | 51.7 | 18.9 | 6.1 | 1.9 |
† The standard deviation on all direct polarization magic angle spinning (DPMAS) values are 0.015.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maaz, T.M.; Hockaday, W.C.; Deenik, J.L. Biochar Volatile Matter and Feedstock Effects on Soil Nitrogen Mineralization and Soil Fungal Colonization. Sustainability 2021, 13, 2018. https://doi.org/10.3390/su13042018
AMA Style
Maaz TM, Hockaday WC, Deenik JL. Biochar Volatile Matter and Feedstock Effects on Soil Nitrogen Mineralization and Soil Fungal Colonization. Sustainability. 2021; 13(4):2018. https://doi.org/10.3390/su13042018
Chicago/Turabian StyleMaaz, Tai McClellan, William C. Hockaday, and Jonathan L. Deenik. 2021. "Biochar Volatile Matter and Feedstock Effects on Soil Nitrogen Mineralization and Soil Fungal Colonization" Sustainability 13, no. 4: 2018. https://doi.org/10.3390/su13042018
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.