Re-Introduction of Ancient Wheat Cultivars into Organic Agriculture—Emmer and Einkorn Cultivation Experiences under Marginal Conditions


 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Designs
2.2. Grain Quality Determination
2.2.1. Macronutrient Content
2.2.2. Dietary Fiber Components
2.2.3. Phenolic Compounds
2.2.4. Antioxidant Activity Assays
2.2.5. Statistical Analyses
3. Results
3.1. Yield Potential and Stability
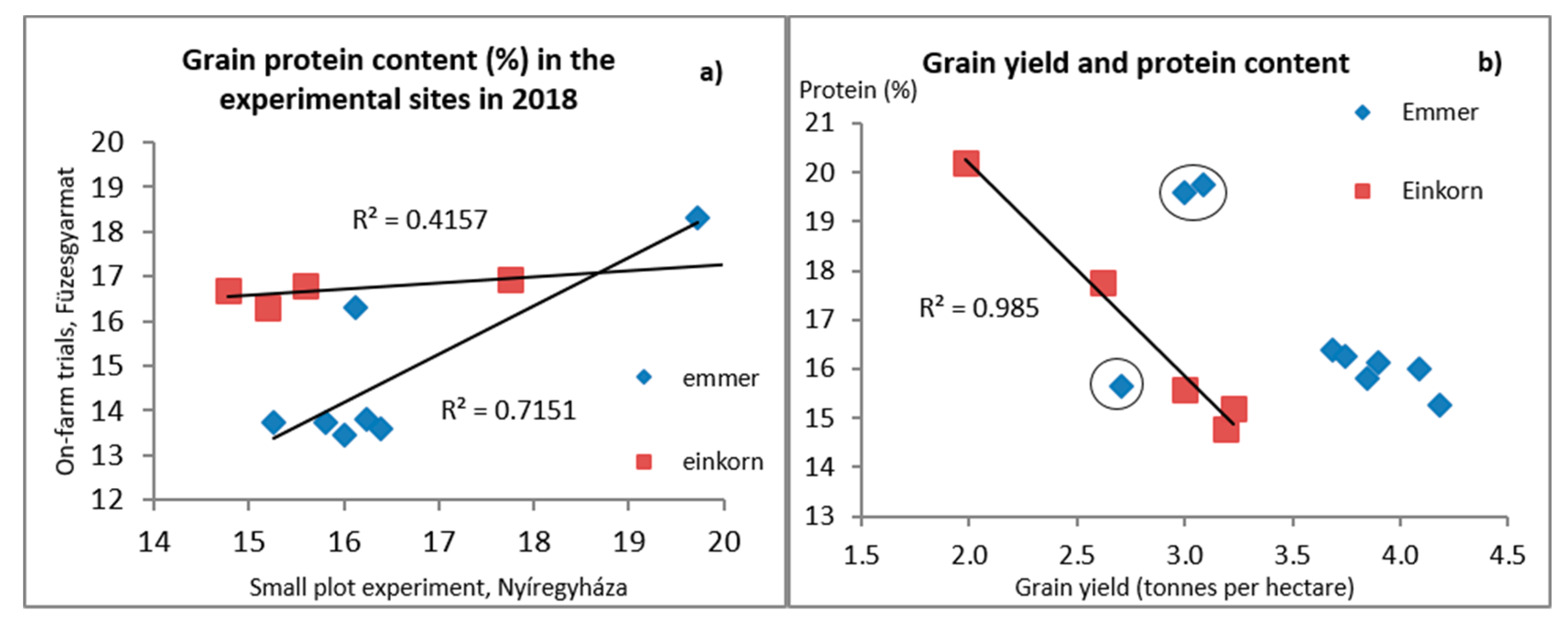
3.2. Factors Affecting Grain Yield
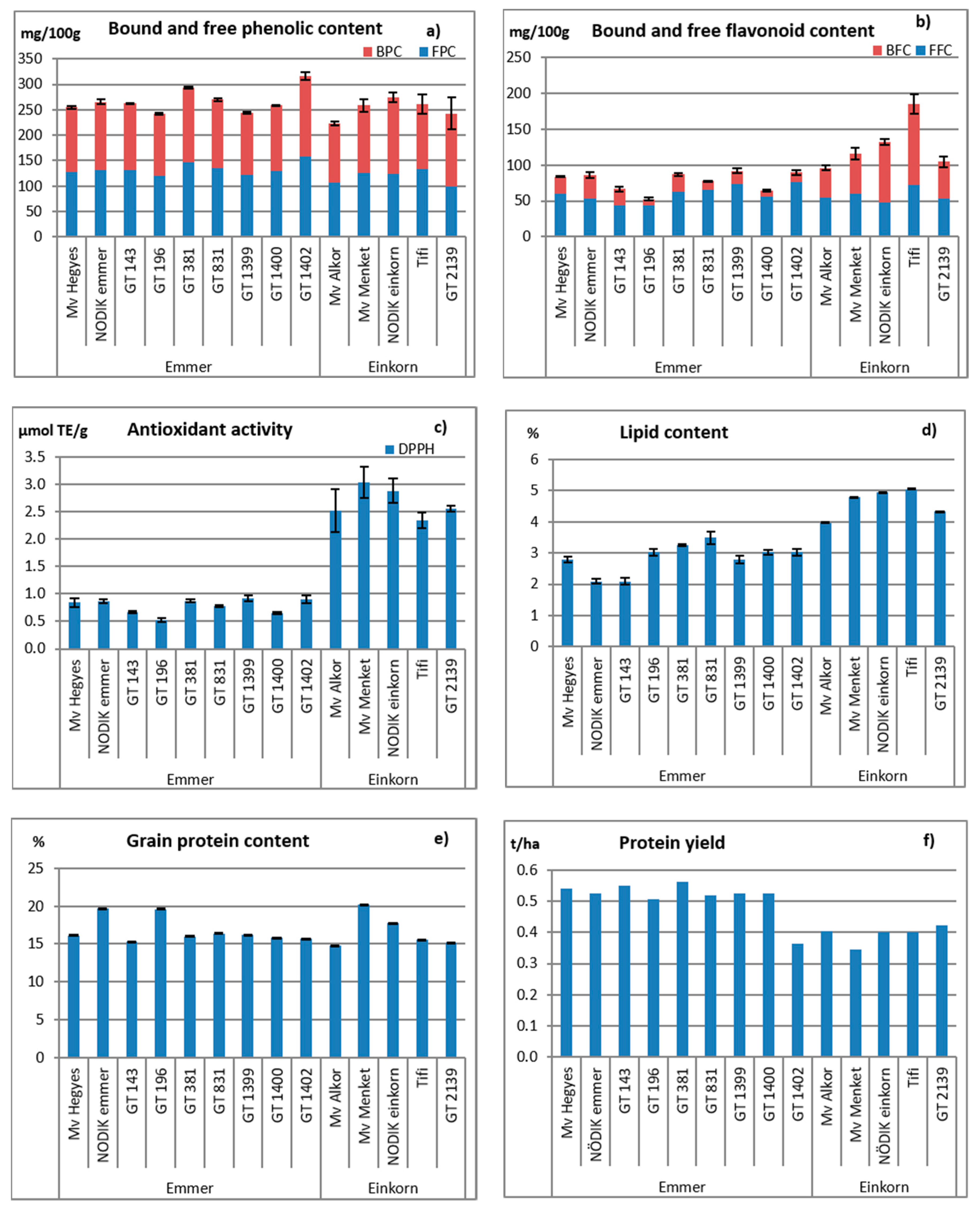
3.3. Grain Quality Attributes
3.4. Disease Resistance of Emmer and Einkorn
4. Discussion
4.1. Grain Yield and Yield Stability
4.2. Grain Quality
4.3. Disease Resistance of Emmer and Einkorn
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Füleky, G. Cultivated plants, primarily as food sources. In Knowledge for Sustainable Development; UNESCO: Paris, France, 2004; pp. 1035–1057. [Google Scholar]
- Garvin, D.F.; Welch, R.M.; Finley, J.W. Historical shifts in the seed mineral micronutrient concentration of US hard red winter wheat germplasm. J. Sci. Food Agric. 2006, 86, 2213–2220. [Google Scholar] [CrossRef]
- Murphy, K.M.; Reeves, P.G.; Jones, S.S. Relationship between yield and mineral nutrient concentrations in historical and modern spring wheat cultivars. Euphytica 2008, 163, 381–390. [Google Scholar] [CrossRef]
- Mayer, A.M. Historical changes in the mineral content of fruits and vegetables. Br. Food J. 1997, 99, 207–211. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Historical variation in the mineral composition of edible horticultural products. J. Hortic. Sci. Biotech. 2005, 80, 660–667. [Google Scholar] [CrossRef]
- Davis, D.R. Declining fruit and vegetable nutrient composition: What is the evidence? J. Am. Soc. Hortic. Sci. 2009, 44, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Marles, R.J. Mineral nutrient composition of vegetables, fruits and grains: The context of reports of apparent historical declines. J. Food Compos. Anal. 2017, 56, 93–103. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Bueren, E.T.; Ceccarelli, S.; Grando, S.; Upadhyaya, H.D.; Ortiz, R. Diversifying Food Systems in the Pursuit of Sustainable Food Production and Healthy Diets. Trends Plant Sci. 2017, 22, 842–856. [Google Scholar] [CrossRef] [Green Version]
- Mie, A.; Andersen, H.R.; Gunnarsson, S.; Kahl, J.; Kesse-Guyot, E.; Rembiałkowska, E.; Quaglio, G.; Grandjean, P. Human health implications of organic food and organic agriculture: A comprehensive review. Environ. Health 2017, 16, 111. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5658984/ (accessed on 19 February 2020). [CrossRef] [Green Version]
- European Soil Data Centre (ESDAC) Database. Available online: https://esdac.jrc.ec.europa.eu/content/soil-map-hungary-clay-mineral-association-soils-main-substance-regime-type-soils-main (accessed on 19 February 2020).
- Yao, G.; Zhang, J.; Yang, L.; Xu, H.; Jiang, Y.; Xiong, L.; Zhang, C.; Zhang, Z.; Ma, Z.; Sorrells, M.E. Genetic mapping of two powdery mildew resistance genes in einkorn (Triticum monococcum L.) accessions. Theor. Appl. Genet. 2007, 114, 351–358. [Google Scholar] [CrossRef]
- Zaharieva, M.; Ayana, N.G.; Al Hakimi, A.; Misra, S.C.; Monneveux, P. Cultivated emmer wheat (Triticum dicoccon Schrank), an old crop with promising future: A review. Genet. Resour. Crop. Evol. 2010, 57, 937–962. [Google Scholar] [CrossRef]
- Góral, T.; Ochodzki, P. Fusarium head blight resistance and mycotoxin profiles of four Triticum species genotypes. Phytopathol. Mediterr. 2017, 56, 175–186. [Google Scholar]
- Buvaneshwari, G.; Yenagi, N.B.; Hanchinal, R.R.; Naik, R.K. Glycaemic responses to dicoccum products in the dietary management of diabetes. Ind. J. Nutr. Dietet. 2003, 40, 363–368. [Google Scholar]
- Čurná, V.; Lacko-Bartošová, M. Chemical composition and nutritional value of emmer wheat (Triticum dicoccon Schrank): A Review. J. Cent. Eur. Agric. 2017, 18, 117–134. [Google Scholar]
- Dinu, M.; Whittaker, A.; Pagliai, G.; Benedettelli, S.; Sofi, F. Ancient wheat species and human health: Biochemical and clinical implications. J. Nutr. Biochem. 2018, 52, 1–9. [Google Scholar] [CrossRef]
- Loje, H.; Moller, B.; Laustsen, A.M.; Hansen, A. Chemical composition, functional properties and sensory profiling of einkorn (Triticum monococcum L.). J. Cereal. Sci. 2003, 37, 231–240. [Google Scholar] [CrossRef]
- Nakov, G.; Stamatovska, V.; Vasileva, N.; Damyanova, S.; Necinova, L. Nutritional properties of einkorn wheat (Triticum monococcum L.)—Review. Reports Awarded with “Best Paper” Crystal Prize. In 55th Science Conference of Ruse University, Razgrad, Bulgaria, 4 November 2016; “Angel Kanchev” University of Ruse: Ruse, Bulgaria, 2016; pp. 381–384. [Google Scholar]
- Piergiovanni, A.R.; Laghetti, G.; Perrino, P. Characteristics of meal from hulled wheats (Triticum dicoccon Schrank and T. spelta L.): An evaluation of selected accessions. Cereal. Chem. 1996, 73, 732–735. [Google Scholar]
- Arzani, A. Emmer (Triticum tugidum spp. Dicoccum) flour and breads. In Flour and Breads and Their Fortification in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2011; pp. 69–78. ISBN 978-0-12-380886-8. [Google Scholar]
- Akar, T.; Cengiz, M.F.; Tekin, M. A comparative study of protein and free amino acid contents in some important ancient wheat lines. Qual. Assur. Saf. Crop. 2019, 11, 191–200. [Google Scholar] [CrossRef]
- Lachman, J.; Miholová, D.; Pivec, V.; Jírů, K.; Janovská, D. Content of phenolic antioxidants and selenium in grain of einkorn (Triticum monococcum), emmer (Triticum dicoccum) and spring wheat (Triticum aestivum) varieties. Plant Soil Environ. 2011, 57, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Şahin, Y.; Yıldırım, A.; Yücesan, B.; Zencirci, N.; Erbayram, Ş.; Gürel, E. Phytochemical content and antioxidant activity of einkorn (Triticum monococcum ssp. monococcum), bread (Triticum aestivum L.), and durum (Triticum durum Desf.) wheat. Progr. Nutr. 2018, 19, 450–459. Available online: https://mattioli1885journals.com/index.php/progressinnutrition/article/view/5847 (accessed on 10 January 2020).
- Zrcková, M.; Capouchová, I.; Paznocht, L.; Eliášová, M.; Dvořák, P.; Konvalina, P.; Janovská, D.; Orsák, M.; Bečková, L. Variation of the total content of polyphenols and phenolic acids in einkorn, emmer, spelt and common wheat grain as a function of genotype, wheat species and crop year. Plant Soil Environ. 2019, 65, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.J.; Su, Y.H.; Dunham, S.J.; Rakszegi, M.; Bedő, Z.; McGrath, S.P.; Shewry, P.R. Variation in mineral micronutrient concentrations in grain of wheat lines of diverse origin. J. Cereal. Sci. 2009, 49, 290–295. [Google Scholar] [CrossRef]
- Piergiovanni, A.R.; Rizzi, R.; Pannacciulli, E.; Della Gatta, C. Mineral composition in hulled wheat grains: A comparison between emmer (Triticum dicoccon Schrank) and spelt (T. spelta L.) accessions. Int. J. Food Sci. Nutr. 1997, 48, 381–386. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Shewry, R.P.; Hey, S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J. Cereal. Sci. 2015, 65, 236–243. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Approved Methods of Association of Official Analytical Chemists, 5th ed.; Association of Official Analytical Chemists, Inc.: Arlington, VA, USA, 1990. [Google Scholar]
- Prosky, L.; Asp, N.G.; Scheweizer, T.F.; de Vries, J.W.; Furda, I. Determination of insoluble, soluble, and total dietary fiber in foods and food products: Interlaboratory study. J. Assoc. Off. Anal. Chem. 1988, 71, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Dinelli, G.; Segura-Carretero, A.; Di Silvestro, R.; Marotti, I.; Fu, S.; Benedettelli, S.; Ghiselli, L.; Fernandez-Gutierrez, A. Profiles of phenolic compounds in modern and old common wheat varieties determined by liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 7670–7681. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela–Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin–Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Brand-Williams, W.; Cuvelier, H.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Castagna, R.; Borghi, B.; Di Fonzo, N.; Heun, M.; Salamini, F. Yield and related traits of einkorn (T. monococcum ssp. monococcum) in different environments. Eur. J. Agron. 1995, 4, 371–378. [Google Scholar] [CrossRef]
- Konvalina, P.; Capouchová, I.; Stehno, Z.; Moudrý, J. Differences in yield parameters of emmer in comparison with old and new varieties of bread wheat. Afr. J. Agric. Res. 2012, 7, 986–992. [Google Scholar]
- Marino, S.; Tognetti, R.; Alvino, A. Crop yield and grain quality of emmer populations grown in central Italy, as affected by nitrogen fertilization. Eur. J. Agron. 2009, 31, 233–240. [Google Scholar] [CrossRef]
- Troccoli, A.; Codianni, P. Appropriate seeding rate for einkorn, emmer, and spelt grown under rainfed condition in southern Italy. Eur. J. Agron. 2005, 22, 293–300. [Google Scholar] [CrossRef]
- Dhanavath, S.; Rao, U.J. Nutritional and nutraceutical properties of Triticum dicoccum wheat and its health benefits: An overview. J. Food Sci. 2017, 82, 2243–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachman, J.; Orsák, M.; Pivec, V.; Jírů, K. Antioxidant activity of grain of einkorn (Triticum monococcum L.), emmer (Triticum dicoccum Schuebl [Schrank]) and spring wheat (Triticum aestivum L.) varieties. Plant Soil Environ. 2012, 58, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Serpen, A.; Gökmen, V.; Karagöz, A.; Scanlon, M.G. Phytochemical quantification and total antioxidant capacities of emmer (Triticum dicoccon Schrank) and einkorn (Triticum monococcum L.) wheat landraces. J. Agric. Food Chem. 2008, 56, 7285–7292. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A.; Ratti, S. Influence of genetic and environmental factors on selected nutritional traits of Triticum monococcum. J. Agric. Food Chem. 2009, 57, 6342–6348. [Google Scholar] [CrossRef]
- Konvalina, P.; Capouchová, I.; Stehno, Z.; Moudrý, J. Agronomic characteristics of the spring forms of the wheat landraces (einkorn, emmer, spelt, intermediate bread wheat) grown in organic farming. J. Agrobiol. 2010, 27, 9–17. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name/Code of Accession | Common Name/Origin | Number of Replicates | ||
|---|---|---|---|---|
| 2015 | 2016– | |||
| Winter emmer | Mv Hegyes | registered variety—HU | 4 | 4 |
| NÖDIK emmer (RCAT 004664) | Emmer roter (German landrace) | 4 | 4 | |
| GT 143 | Schwarzwerdender—CH | 3 | 4 | |
| GT 196 | Zweikorn—CH | 1 | 4 | |
| GT 381 | Schwarzer Samtemmer—CH | 1 | 4 | |
| GT 831 | Blauemmer—CH | 3 | 4 | |
| GT 1399 | Grauer—CH | 2 | 4 | |
| GT 1400 | Schwarzbehaarter—CH | 3 | 4 | |
| GT 1402 | Weisser behaarter—CH | 1 | 4 | |
| GT 2140 | "Züblin" WS—CH | 3 | 0 | |
| Winter einkorn | Mv Alkor | registered variety—HU | 4 | 4 |
| Mv Menket | registered variety—HU | 4 | 4 | |
| NÖDIK einkorn (RCAT 074129) | Morocco (COLL. SCHIEMANN) | 4 | 4 | |
| Tifi | registered variety—NL | 4 | 4 | |
| GT 2139 | unknown—CH | 1 | 4 | |
| Nyíregyháza, 2016 | Nyíregyháza, 2017 | Nyíregyháza, 2018 | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Name of Accession | Drechs. gram. (%) | YR (%) | LR (%) | FHB/10 spikes | S.tritici (%) | YR (%) | LR (%) | FHB (%) | S.tritici (%) | YR (%) | LR (%) | FHB (%) | |||||||||
| freq/sev | freq/sev | freq. | sev. | freq. | sev. | freq. | sev. | freq. | sev. | freq. | sev. | freq. | sev. | freq. | sev. | freq. | sev. | freq. | sev. | ||
| Emmer | |||||||||||||||||||||
| Mv Hegyes | 2/2 | 35/100 | 0 | 2.5 | 2.5 | 0 | 0 | 17.5 | 3.8 | 72.5 | 10.0 | 31.3 | 8.5 | 1.25 | 1.25 | 22.5 | 3.5 | 2.5 | 1.25 | 2.5 | 0.75 |
| NÖDIK emmer | 100/65 | 60/100 | 0 | 17.1 | 24.4 | 91.3 | 26.3 | 11.3 | 2.0 | 63.8 | 6.8 | 22.5 | 7.5 | 5 | 3.75 | 7.5 | 2.5 | 1.25 | 1.25 | 0 | 0 |
| GT-143 | 0 | 35/100 | 0 | 3.3 | 3.3 | 0 | 0 | 0.5 | 0.3 | 70.0 | 13.8 | 21.3 | 6.3 | 0 | 0 | 11.25 | 2.25 | 0 | 0 | 0 | 0 |
| GT-196 | 100/80 | 50/100 | 0 | 20 | 20 | 40.0 | 8.3 | 1.7 | 0.7 | 46.7 | 5.0 | 16.7 | 6.7 | 3.75 | 2.5 | 8.75 | 2 | 1.25 | 1.25 | 5 | 1.25 |
| GT-381 | 5/5 | 30/100 | 0 | 30 | 70 | 0.3 | 0.3 | 2.5 | 1.3 | 67.5 | 13.8 | 21.7 | 7.0 | 0 | 0 | 2.5 | 1.5 | 5 | 2.5 | 0 | 0 |
| GT-831 | 10/15 | 45/100 | 0 | 3.3 | 23.3 | 0 | 0 | 8.8 | 1.5 | 62.5 | 10.0 | 22.5 | 6.5 | 1.25 | 1.25 | 13.75 | 5.5 | 0 | 0 | 0 | 0 |
| GT-1399 | 8/10 | 40/100 | 0 | 0 | 0 | 0 | 0 | 1.3 | 0.8 | 76.3 | 14.8 | 12.0 | 6.0 | 3.75 | 1.25 | 10 | 2.5 | 7.5 | 3.75 | 0 | 0 |
| GT-1400 | 15/15 | 70/100 | 0 | 13.3 | 12.3 | 0 | 0 | 13.8 | 4.8 | 50.0 | 13.3 | 27.5 | 7.5 | 1.25 | 1.25 | 17.5 | 5 | 6.25 | 2 | 0 | 0 |
| GT-1402 | 90/10 | 20/100 | 0 | 10 | 30 | 0 | 0 | 0 | 0 | 60.0 | 13.0 | 21.3 | 7.0 | 1.25 | 1.25 | 16.25 | 4.5 | 6.25 | 5 | 3.75 | 0.75 |
| Einkorn | |||||||||||||||||||||
| Mv Alkor | 0 | 0 | 5/15 | 40 | 25.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0.25 | 0.75 | 10 | 3.75 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mv Menket | 0 | 0 | 0 | 100 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0.75 | 1.25 | 20 | 10 | 0 | 0 | 2.5 | 11.25 | 0 | 0 |
| NÖDIK einkorn | 0 | 0 | 5/15 | 22.5 | 22.0 | 5 | 1.25 | 0 | 0 | 0 | 0 | 0 | 0 | 7.5 | 3.25 | 0 | 0 | 0 | 0 | 0 | 0 |
| Tifi | 0 | 0 | 10/5 | 25.0 | 32.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 3.75 | 0 | 0 | 0 | 0 | 2.5 | 1.75 |
| GT-2139 | 0 | 0 | 80/100 | 20 | 15 | 0 | 0 | 0 | 0 | 0 | 0 | 1.5 | 3.25 | 3.75 | 2.5 | 0 | 0 | 0 | 0 | 2.5 | 1.25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bencze, S.; Makádi, M.; Aranyos, T.J.; Földi, M.; Hertelendy, P.; Mikó, P.; Bosi, S.; Negri, L.; Drexler, D. Re-Introduction of Ancient Wheat Cultivars into Organic Agriculture—Emmer and Einkorn Cultivation Experiences under Marginal Conditions. Sustainability 2020, 12, 1584. https://doi.org/10.3390/su12041584
Bencze S, Makádi M, Aranyos TJ, Földi M, Hertelendy P, Mikó P, Bosi S, Negri L, Drexler D. Re-Introduction of Ancient Wheat Cultivars into Organic Agriculture—Emmer and Einkorn Cultivation Experiences under Marginal Conditions. Sustainability. 2020; 12(4):1584. https://doi.org/10.3390/su12041584
Chicago/Turabian StyleBencze, Szilvia, Marianna Makádi, Tibor J. Aranyos, Mihály Földi, Péter Hertelendy, Péter Mikó, Sara Bosi, Lorenzo Negri, and Dóra Drexler. 2020. "Re-Introduction of Ancient Wheat Cultivars into Organic Agriculture—Emmer and Einkorn Cultivation Experiences under Marginal Conditions" Sustainability 12, no. 4: 1584. https://doi.org/10.3390/su12041584