Hot Spots and Hot Moments of Soil Moisture Explain Fluctuations in Iron and Carbon Cycling in a Humid Tropical Forest Soil

 ,
,  , , and
, , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Site Description, Experimental Design, and Soil Sampling Procedure
2.2. Soil Biogeochemical Characterization and Extractions (Measurements within Days)
Rapidly-Reducible Fe Oxides (FeIIIRR) Microbial Bioreduction Assay
2.3. Soil Sensors and Data Collection (Measurements within Hours)
2.4. Statistical Analyses
3. Results
3.1. Soil Characteristics across Sites
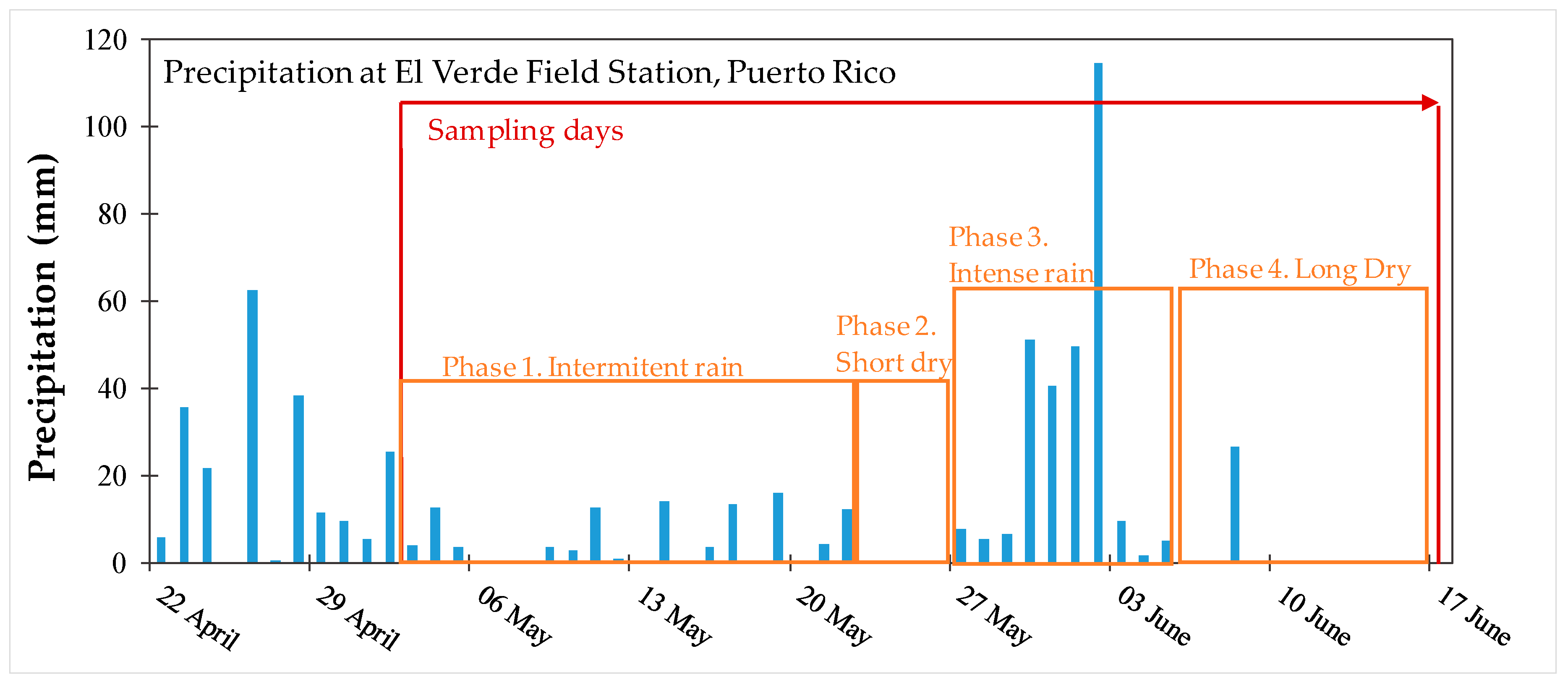
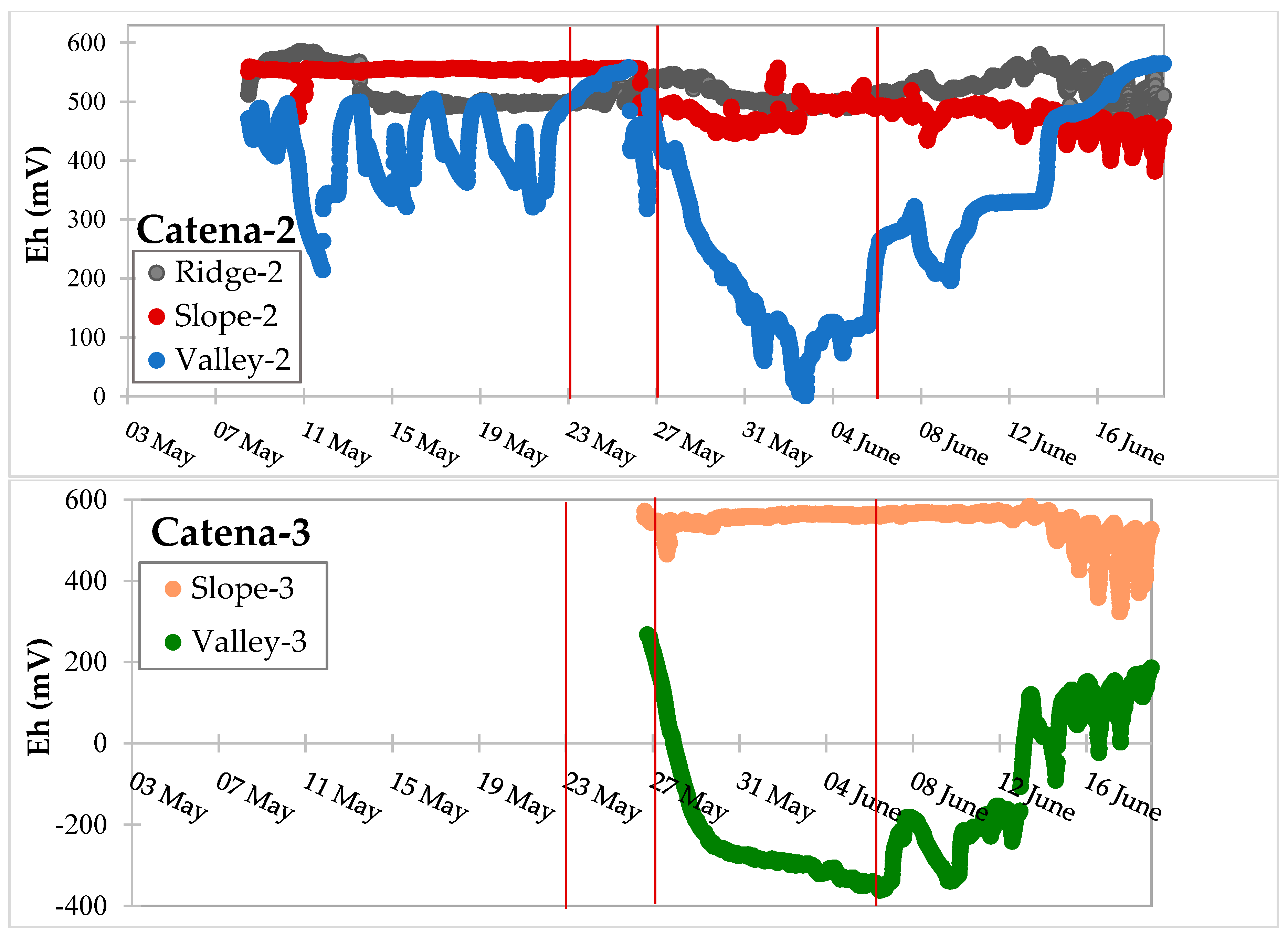
3.2. Precipitation and Environmental Factors Involved in Redox Processes
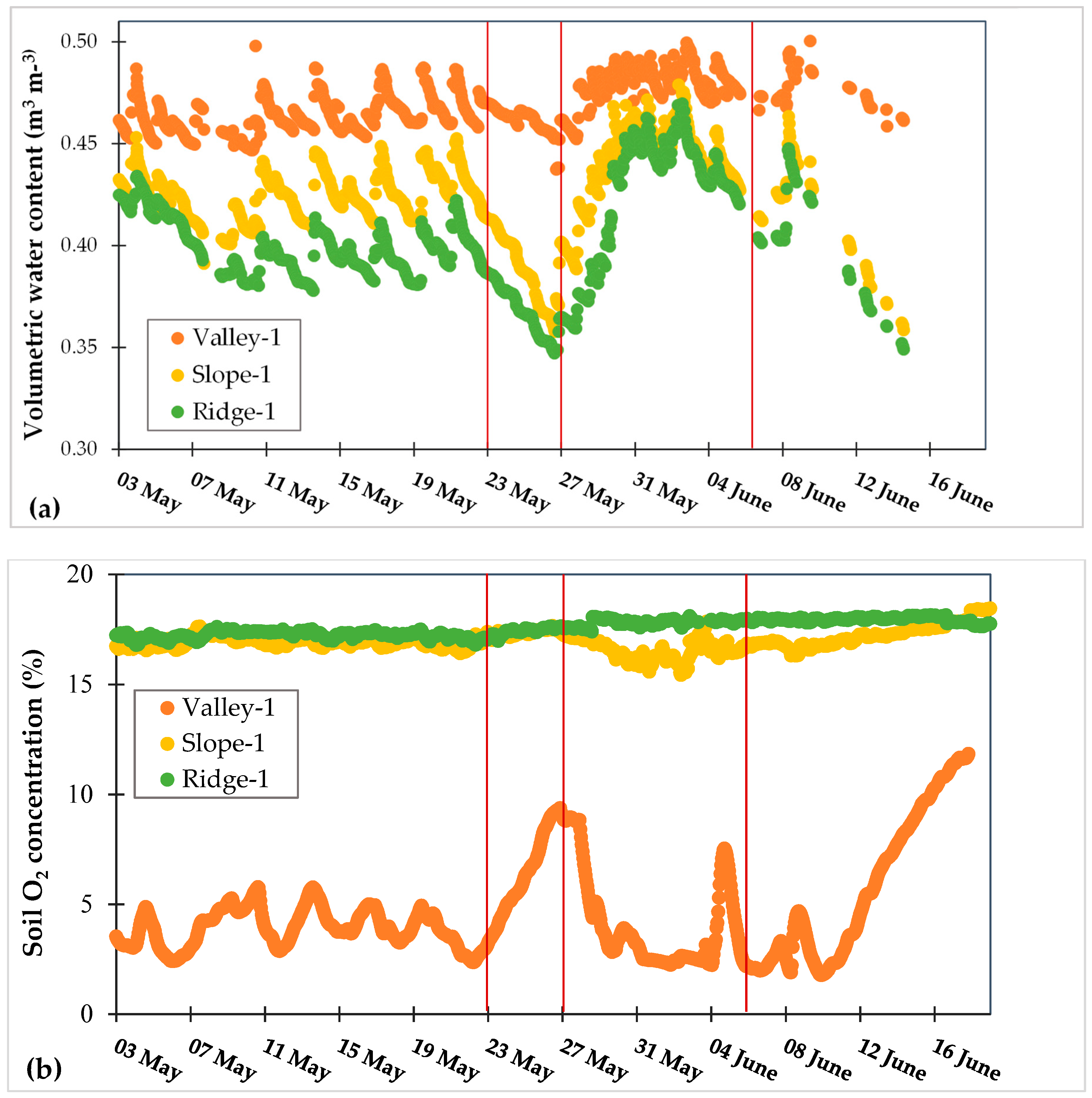
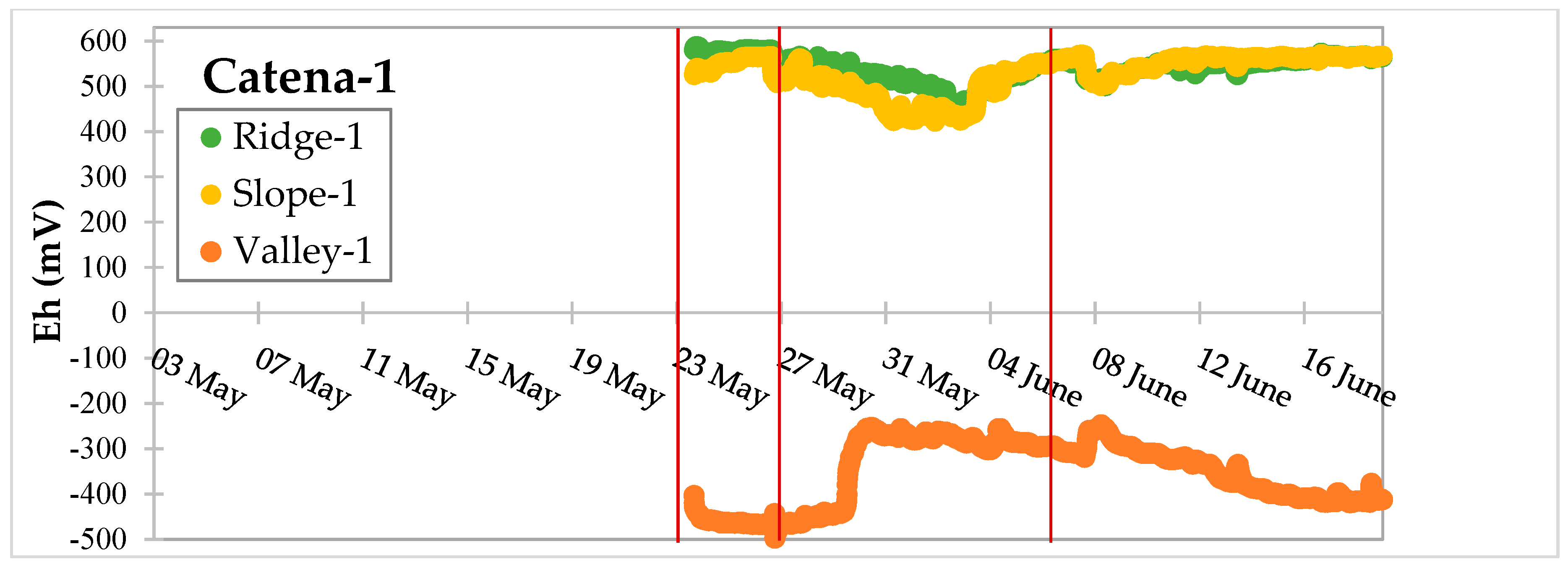
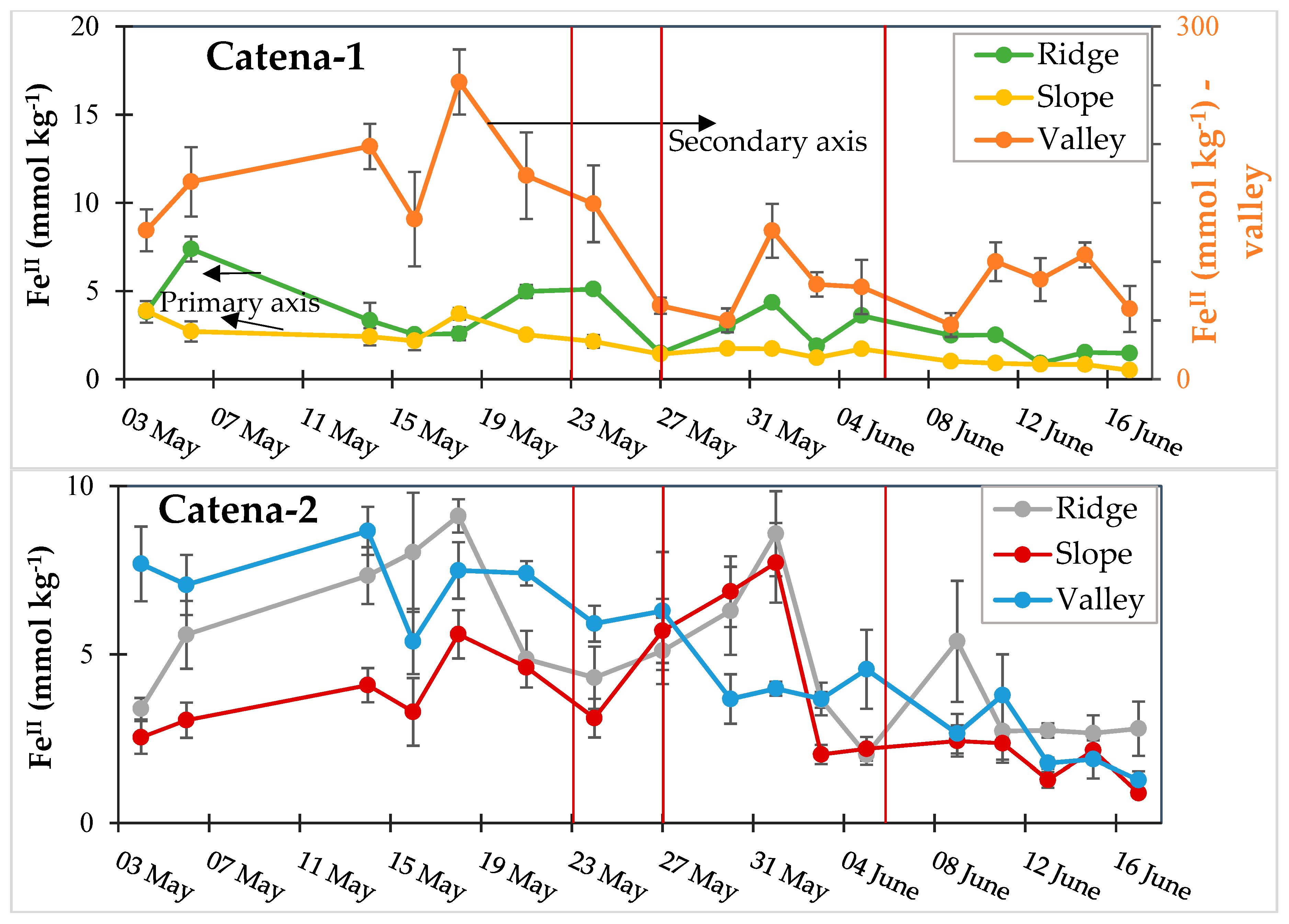
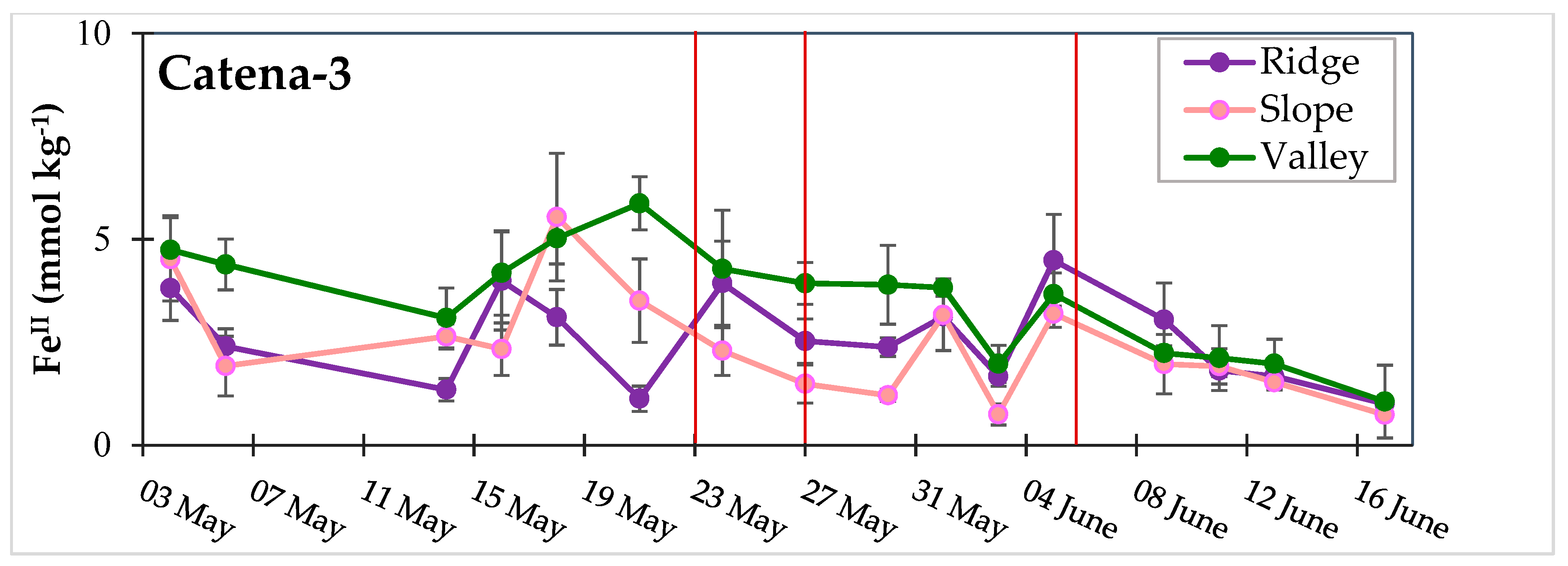
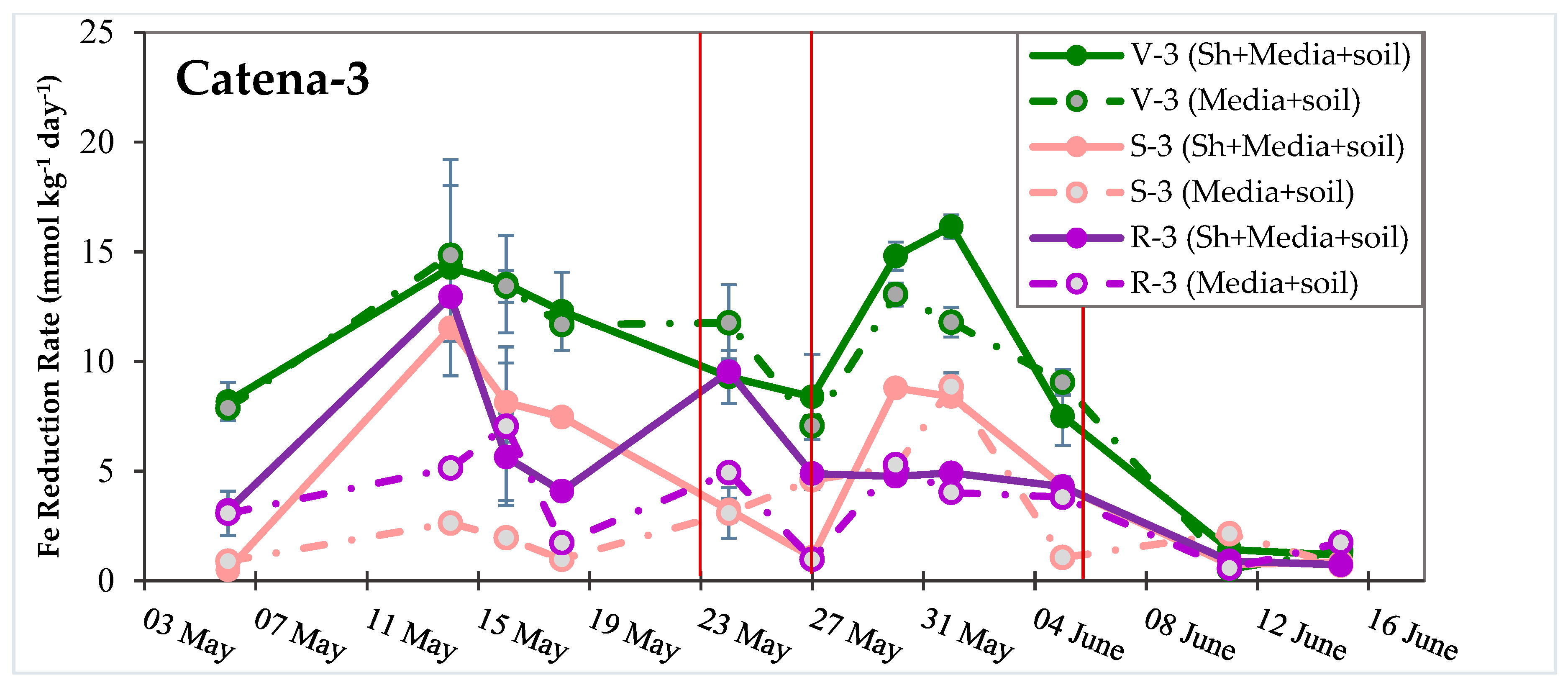
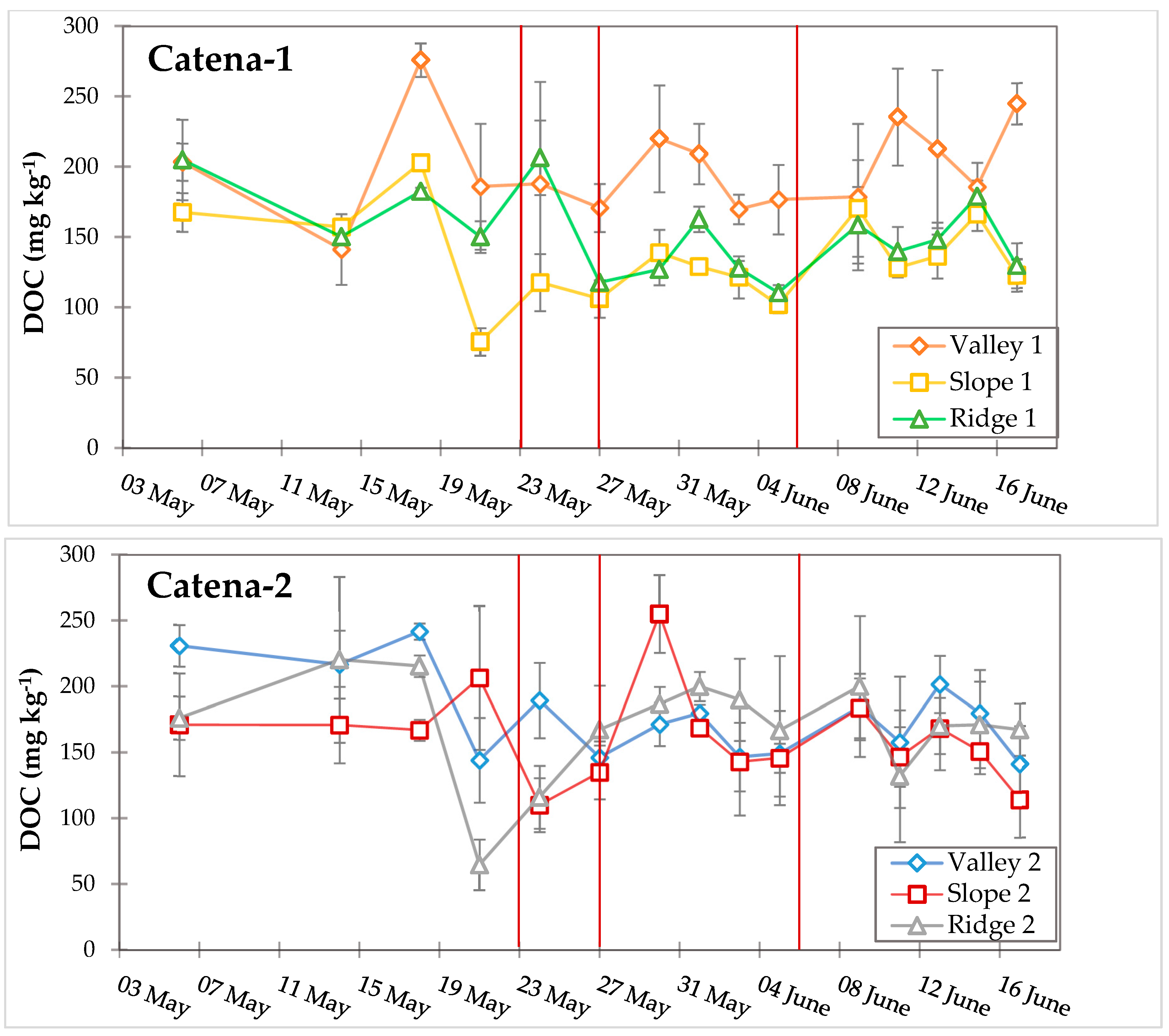
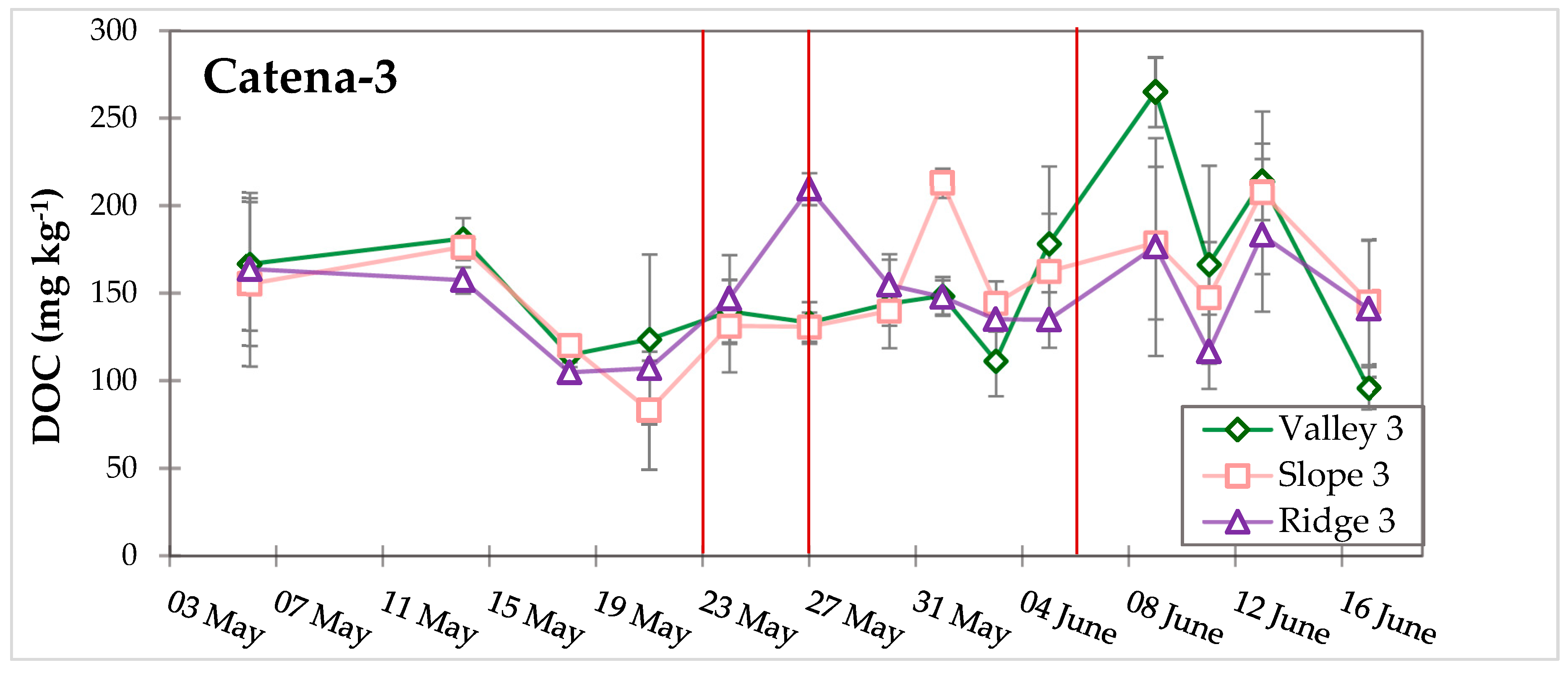
3.3. Dynamic Biogeochemical Soil Measurements
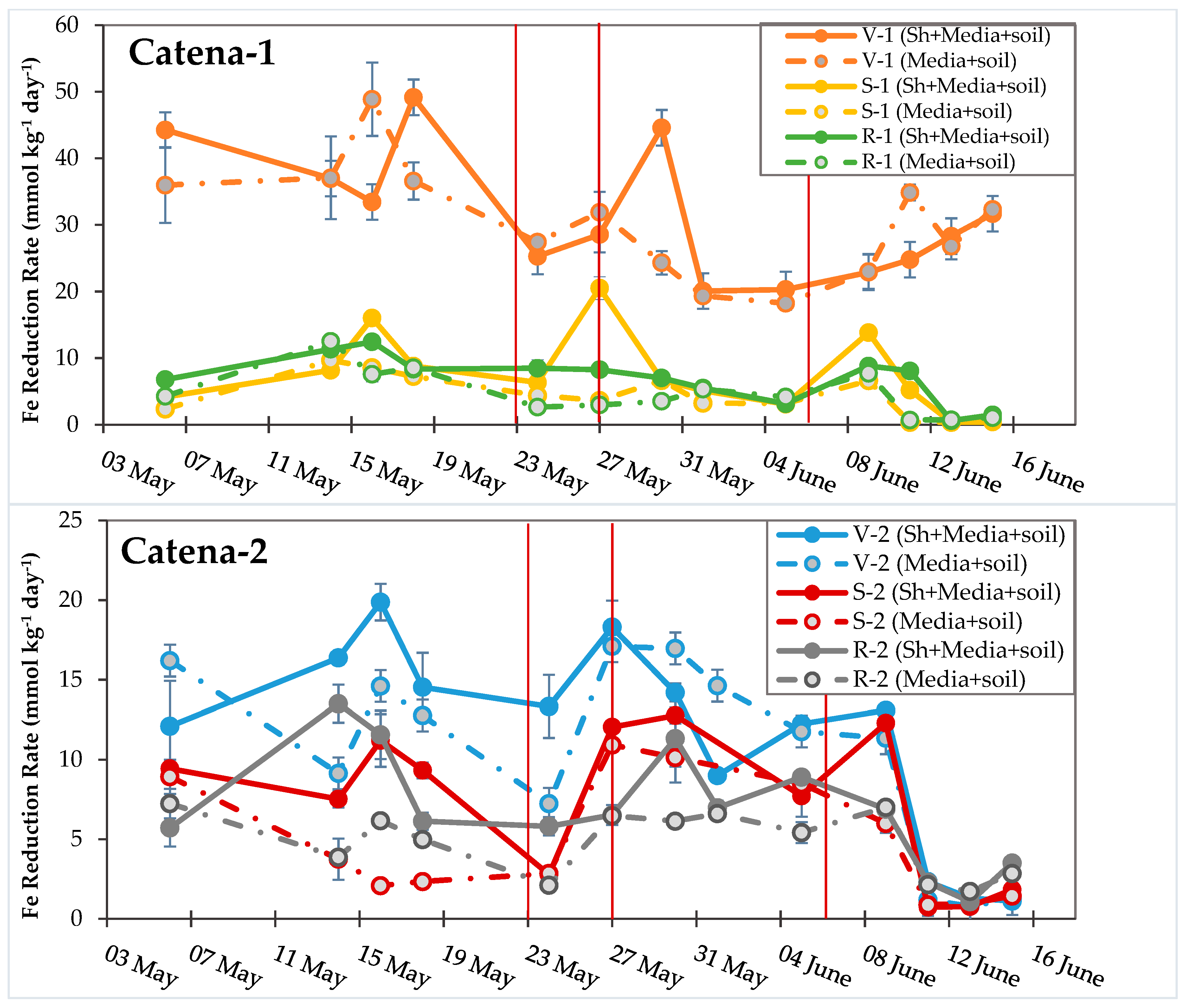
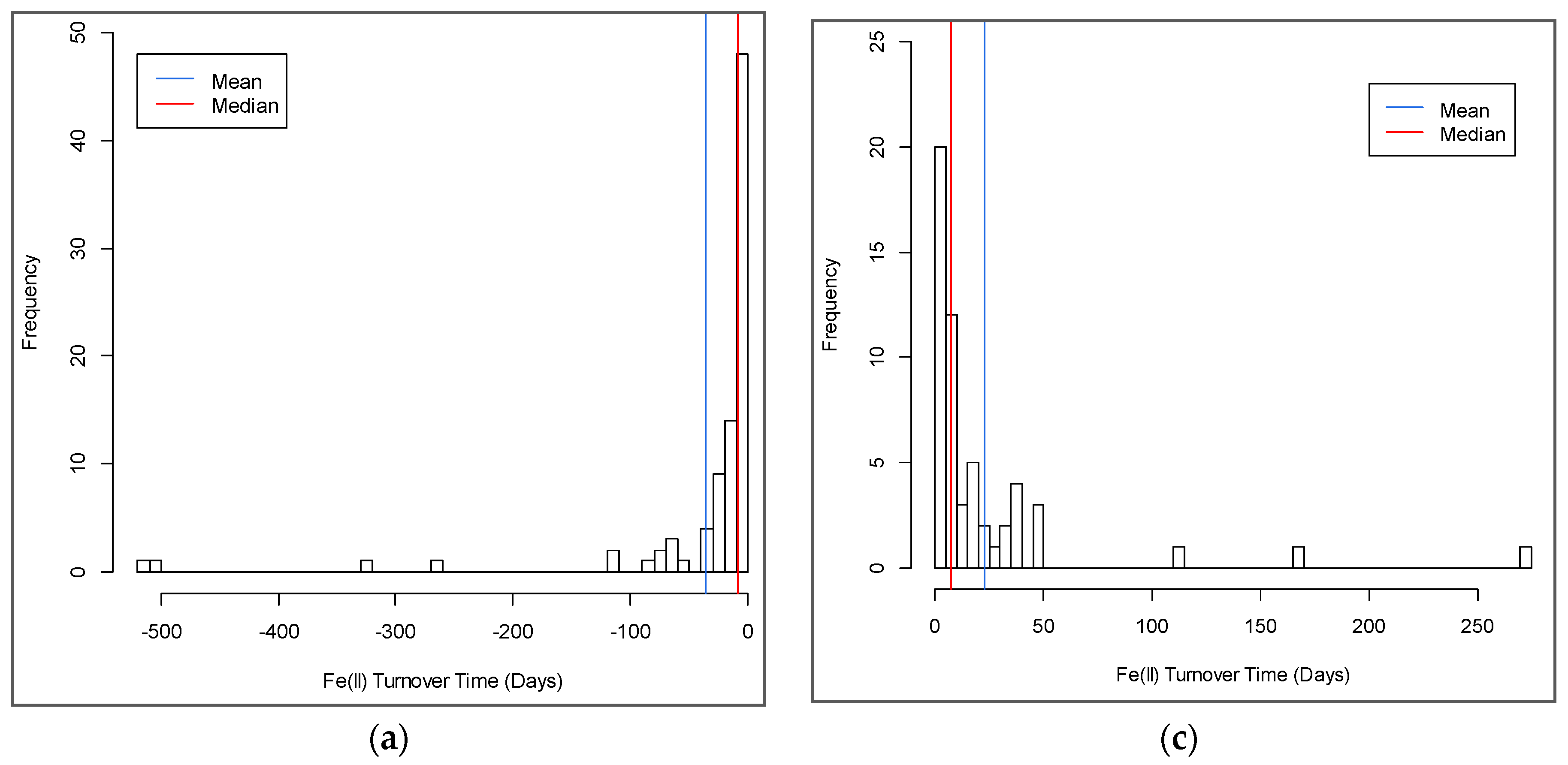
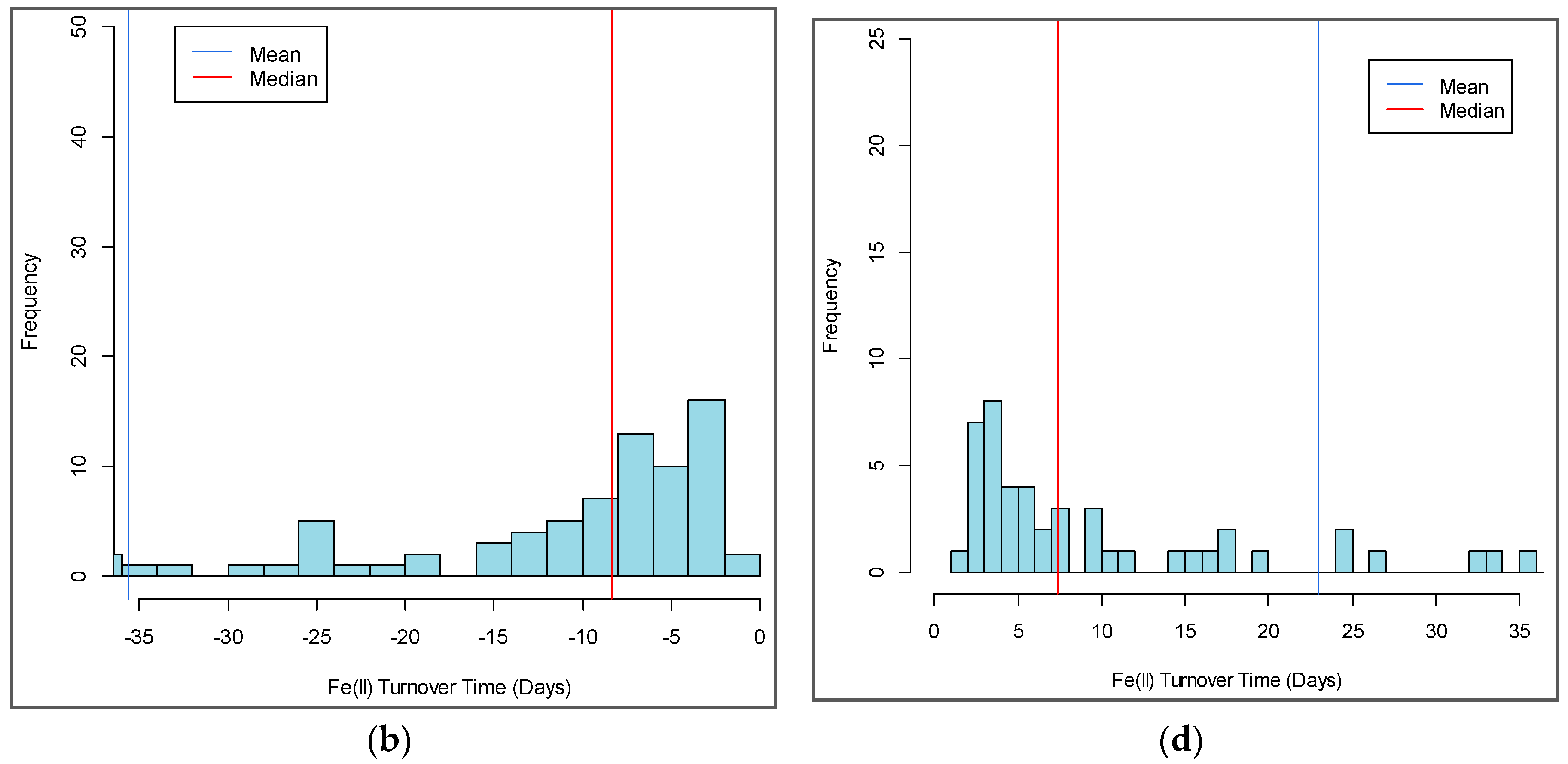
3.4. Turnover Timescales for HCl-Extractable FeII Pool and Statistical Models to Predict Soil Biogeochemical Parameters
4. Discussion
4.1. Turnover Times for HCl-Extractable FeII Pool and Drivers of Fe Redox Processes
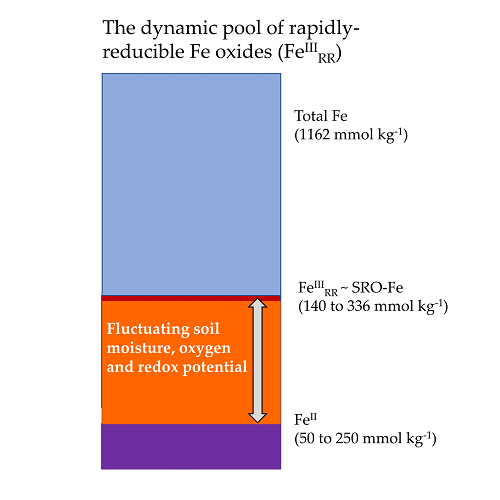
4.2. The Dynamic Pool of Rapidly Reducible Fe Oxides (FeIIIRR)
4.3. Soil Moisture and Precipitation Influence Fe–C Redox Cycling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kleber, M.; Eusterhues, K.; Keiluweit, M.; Mikutta, C.; Mikutta, R.; Nico, P.S. Mineral–organic associations: Formation, properties, and relevance in soil environments. Adv. Agron. 2015, 130, 1–140. [Google Scholar]
- FAO. Status of the World’s Soil Resources (SWSR)–Main Report. In Food and Agriculture Organization of the United Nations and Intergovernmental Technical Panel on Soils; FAO: Rome, Italy, 2015; Volume 650. [Google Scholar]
- Broedel, E.; Tomasella, J.; Cândido, L.A.; Randow, C. Deep soil water dynamics in an undisturbed primary forest in central Amazonia: Differences between normal years and the 2005 drought. Hydrol. Process. 2017, 31, 1749–1759. [Google Scholar] [CrossRef]
- Hall, S.J.; McDowell, W.H.; Silver, W.L. When wet gets wetter: Decoupling of moisture, redox biogeochemistry, and greenhouse gas fluxes in a humid tropical forest soil. Ecosystems 2013, 16, 576–589. [Google Scholar] [CrossRef]
- Saleska, S.R.; Miller, S.D.; Matross, D.M.; Goulden, M.L.; Wofsy, S.C.; Da Rocha, H.R.; De Camargo, P.B.; Crill, P.; Daube, B.C.; De Freitas, H.C. Carbon in Amazon forests: Unexpected seasonal fluxes and disturbance-induced losses. Science 2003, 302, 1554–1557. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Dávila, E.; Cayuela, L.; González-Caro, S.; Aldana, A.M.; Stevenson, P.R.; Phillips, O.; Cogollo, Á.; Peñuela, M.C.; von Hildebrand, P.; Jiménez, E. Forest biomass density across large climate gradients in northern South America is related to water availability but not with temperature. PLoS ONE 2017, 12, e0171072. [Google Scholar] [CrossRef] [PubMed]
- Brokaw, N.; Crowl, T.A.; Lugo, A.; McDowell, W.H.; Scatena, F.; Waide, R.B.; Willig, M.R. A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response; Oxford University Press: New York, NY, USA, 2012. [Google Scholar]
- Muggler, C.C.; van Griethuysen, C.; Buurman, P.; Pape, T. Aggregation, organic matter, and iron oxide morphology in Oxisols from Minas Gerais, Brazil. Soil Sci. 1999, 164, 759–770. [Google Scholar] [CrossRef]
- Rezende, M.; Curi, N.; Rezende, S.; Corrêa, G. Pedologia: Base Para Distinção de Ambientes, 5th ed.; Editora UFLA: Lavras, Brazil, 2007; p. 322. [Google Scholar]
- Cleveland, C. Nitrogen and phosphorus additions cause substantial losses of soil carbon from a lowland tropical rain forest. Proc. Natl. Acad. Sci. USA 2006, 103, 10316–10321. [Google Scholar] [CrossRef] [PubMed]
- Silver, W.L.; Lugo, A.; Keller, M. Soil oxygen availability and biogeochemistry along rainfall and topographic gradients in upland wet tropical forest soils. Biogeochemistry 1999, 44, 301–328. [Google Scholar] [CrossRef]
- Martin, S.T. Precipitation and dissolution of iron and manganese oxides. In Environmental Catalysis; CRC Press: Boca Raton, FL, USA, 2005; pp. 61–81. [Google Scholar]
- Peters, V.; Conrad, R. Sequential reduction processes and initiation of CH4 production upon flooding of oxic upland soils. Soil Biol. Biochem. 1996, 28, 371–382. [Google Scholar] [CrossRef]
- Hall, S.J.; Huang, W. Iron reduction: A mechanism for dynamic cycling of occluded cations in tropical forest soils? Biogeochemistry 2017, 136, 91–102. [Google Scholar] [CrossRef]
- Lovley, D.R. Fe (III) and Mn (IV) reduction. In Environmental Microbe-Metal Interactions; American Society of Microbiology: Washington, DC, USA, 2000; pp. 3–30. [Google Scholar]
- Mikutta, R.; Kleber, M.; Torn, M.S.; Jahn, R. Stabilization of soil organic matter: Association with minerals or chemical recalcitrance? Biogeochemistry 2006, 77, 25–56. [Google Scholar] [CrossRef]
- Lugo, A.E.; Brown, S.; Brinson, M.M. Concepts in wetland ecology. Ecosyst. World 1990, 15, 53–85. [Google Scholar]
- Ginn, B.; Meile, C.; Wilmoth, J.; Tang, Y.; Thompson, A. Rapid Iron Reduction Rates Are Stimulated by High-Amplitude Redox Fluctuations in a Tropical Forest Soil. Environ. Sci. Technol. 2017, 51, 3250–3259. [Google Scholar] [CrossRef]
- Weber, K.A.; Achenbach, L.A.; Coates, J.D. Microorganisms pumping iron: Anaerobic microbial iron oxidation and reduction. Nat. Rev. Microbiol. 2006, 4, 752–764. [Google Scholar]
- Cornell, R.M.; Schwertmann, U. The Iron Oxides: Structure, Properties, Reactions, Occurrences and Uses; Wiley-VCH: Weinheim, Germany, 2003; p. 667. [Google Scholar]
- Neff, J.C.; Asner, G.P. Dissolved organic carbon in terrestrial ecosystems: Synthesis and a model. Ecosystems 2001, 4, 29–48. [Google Scholar] [CrossRef]
- Gottschalk, P.; Smith, J.U.; Wattenbach, M.; Bellarby, J.; Stehfest, E.; Arnell, N.; Osborn, T.; Jones, C.; Smith, P. How will organic carbon stocks in mineral soils evolve under future climate? Global projections using RothC for a range of climate change scenarios. Biogeosciences 2012, 9, 3151. [Google Scholar] [CrossRef]
- Lovley, D.R.; Holmes, D.E.; Nevin, K.P. Dissimilatory fe (iii) and mn (iv) reduction. Adv. Microb. Physiol. 2004, 49, 219–286. [Google Scholar] [PubMed]
- Carmo, J.B.D.; Keller, M.; Dias, J.D.; Camargo, P.B.D.; Crill, P. A source of methane from upland forests in the Brazilian Amazon. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, C.; Throckmorton, H.; Liles, G.; Heckman, K.; Meding, S.; Horwath, W. Controls on Soil Organic Carbon Partitioning and Stabilization in the California Sierra Nevada. Soil Syst. 2018, 2, 41. [Google Scholar]
- Huang, W.; Hall, S.J. Elevated moisture stimulates carbon loss from mineral soils by releasing protected organic matter. Nat. Commun. 2017, 8, 1774. [Google Scholar] [Green Version]
- Hall, S.J.; Silver, W.L. Iron oxidation stimulates organic matter decomposition in humid tropical forest soils. Glob. Chang. Biol. 2013, 19, 2804–2813. [Google Scholar] [CrossRef] [PubMed]
- Knorr, K.-H. DOC-dynamics in a small headwater catchment as driven by redox fluctuations and hydrological flow paths-are DOC exports mediated by iron reduction/oxidation cycles? Biogeosciences 2013, 10, 891. [Google Scholar] [CrossRef]
- Blodau, C.; Fulda, B.; Bauer, M.; Knorr, K.-H. Arsenic speciation and turnover in intact organic soil mesocosms during experimental drought and rewetting. Geochim. Cosmochim. Acta 2008, 72, 3991–4007. [Google Scholar] [CrossRef]
- Chen, C.; Thompson, A. Ferrous Iron Oxidation under Varying pO2 Levels: The Effect of Fe (III)/Al (III) Oxide Minerals and Organic Matter. Environ. Sci. Technol. 2018, 52, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Thaymuang, W.; Kheoruenromne, I.; Suddhipraharn, A.; Sparks, D.L. The role of mineralogy in organic matter stabilization in tropical soils. Soil Sci. 2013, 178, 308–315. [Google Scholar] [CrossRef]
- Souza, I.F.; Almeida, L.F.; Jesus, G.L.; Pett-Ridge, J.; Nico, P.S.; Kleber, M.; Silva, I.R. Carbon Sink Strength of Subsurface Horizons in Brazilian Oxisols. Soil Sci. Soc. Am. J. 2018, 82, 76–86. [Google Scholar] [CrossRef]
- Silva, L.C.; Doane, T.A.; Corrêa, R.S.; Valverde, V.; Pereira, E.I.; Horwath, W.R. Iron-mediated stabilization of soil carbon amplifies the benefits of ecological restoration in degraded lands. Ecol. Appl. 2015, 25, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Silva Neto, L.F.; Vasconcellos Inda, A.; Bayer, C.; Pinheiro Dick, D.; Tiago Tonin, A. Óxidos de ferro em latossolos tropicais e subtropicais brasileiros em plantio direto. Rev. Bras. Ciênc. Solo 2008, 32, 1873–1881. [Google Scholar] [CrossRef]
- Hall, S.J.; Silver, W.L. Reducing conditions, reactive metals, and their interactions can explain spatial patterns of surface soil carbon in a humid tropical forest. Biogeochemistry 2015, 125, 149–165. [Google Scholar] [CrossRef]
- O’Connell, C.S.; Ruan, L.; Silver, W.L. Drought drives rapid shifts in tropical rainforest soil biogeochemistry and greenhouse gas emissions. Nat. Commun. 2018, 9, 1348. [Google Scholar] [CrossRef] [PubMed]
- Liptzin, D.; Silver, W.L.; Detto, M. Temporal Dynamics in Soil Oxygen and Greenhouse Gases in Two Humid Tropical Forests. Ecosystems 2011, 14, 171–182. [Google Scholar] [CrossRef]
- Hook, P.B.; Burke, I.C. Biogeochemistry in a shortgrass landscape: Control by topography, soil texture, and microclimate. Ecology 2000, 81, 2686–2703. [Google Scholar]
- Bernhardt, E.S.; Blaszczak, J.R.; Ficken, C.D.; Fork, M.L.; Kaiser, K.E.; Seybold, E.C. Control points in ecosystems: Moving beyond the hot spot hot moment concept. Ecosystems 2017, 20, 665–682. [Google Scholar] [CrossRef]
- McClain, M.E.; Boyer, E.W.; Dent, C.L.; Gergel, S.E.; Grimm, N.B.; Groffman, P.M.; Hart, S.C.; Harvey, J.W.; Johnston, C.A.; Mayorga, E. Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems 2003, 6, 301–312. [Google Scholar] [CrossRef]
- Vargas, R.; Sánchez-Cañete, P.E.; Serrano-Ortiz, P.; Curiel Yuste, J.; Domingo, F.; López-Ballesteros, A.; Oyonarte, C. Hot-Moments of Soil CO2 Efflux in a Water-Limited Grassland. Soil Syst. 2018, 2, 47. [Google Scholar] [CrossRef]
- Seiders, V.M. Cretaceous and Lower Tertiary Stratigraphy of the Gurabo and El Yunque Quadrangles, Puerto Rico. 1971. Available online: http://agris.fao.org/agris-search/search.do?recordID=US201300472921 (accessed on 14 June 2017).
- Hodge, E.T. Geology of the Coamo-Guayama District; New York Academy of Sciences: New York, NY, USA, 1920; Volume 28. [Google Scholar]
- Heartsill-Scalley, T.; Scatena, F.N.; Estrada, C.; McDowell, W.; Lugo, A.E. Disturbance and long-term patterns of rainfall and throughfall nutrient fluxes in a subtropical wet forest in Puerto Rico. J. Hydrol. 2007, 333, 472–485. [Google Scholar] [CrossRef]
- Scatena, F.N. An Introduction to the Physiography and History of the Bisley Experimental Watersheds in the Luquillo Mountains of Puerto Rico; Gen. Tech. Report SO-72; US Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1989; Volume 72.
- Almaraz, M. Nitrogen Availability and Loss from Unmanaged and Managed Ecosystems. Ph.D. Thesis, Brown University, Providence, RI, USA, 2017. [Google Scholar]
- McDowell, W.; Estrada-Pinto, A. Rainfall at the El Verde Field Station, 1964–1986; CEER T-228; University of Puerto Rico: San Juan, Puerto Rico, 1988. [Google Scholar]
- Ginn, B.R.; Habteselassie, M.Y.; Meile, C.; Thompson, A. Effects of sample storage on microbial Fe-reduction in tropical rainforest soils. Soil Biol. Biochem. 2014, 68, 44–51. [Google Scholar] [CrossRef]
- Barcellos, D.; Cyle, K.T.; Thompson, A. Faster redox fluctuations can lead to higher iron reduction rates in humid forest soils. Biogeochemistry 2018, 137, 367–378. [Google Scholar] [CrossRef]
- Thompson, A.; Chadwick, O.A.; Rancourt, D.G.; Chorover, J. Iron-oxide crystallinity increases during soil redox oscillations. Geochim. Cosmochim. Acta 2006, 70, 1710–1727. [Google Scholar] [CrossRef]
- Huang, W.; Hall, S.J. Optimized high-throughput methods for quantifying iron biogeochemical dynamics in soil. Geoderma 2017, 306, 67–72. [Google Scholar] [CrossRef]
- Haynes, R.; Francis, G. Changes in microbial biomass C, soil carbohydrate composition and aggregate stability induced by growth of selected crop and forage species under field conditions. Eur. J. Soil Sci. 1993, 44, 665–675. [Google Scholar] [CrossRef]
- Boyer, J.; Groffman, P. Bioavailability of water extractable organic carbon fractions in forest and agricultural soil profiles. Soil Biol. Biochem. 1996, 28, 783–790. [Google Scholar] [CrossRef]
- Ghani, A.; Dexter, M.; Perrott, K. Hot-water extractable carbon in soils: A sensitive measurement for determining impacts of fertilisation, grazing and cultivation. Soil Biol. Biochem. 2003, 35, 1231–1243. [Google Scholar] [CrossRef]
- Guigue, J.; Mathieu, O.; Lévêque, J.; Mounier, S.; Laffont, R.; Maron, P.-A.; Navarro, N.; Chateau, C.; Amiotte-Suchet, P.; Lucas, Y. A comparison of extraction procedures for water-extractable organic matter in soils. Eur. J. Soil Sci. 2014, 65, 520–530. [Google Scholar] [CrossRef]
- Pachon, J.C.; Kowalski, K.; Butterick, J.; Bacon, A.R. The quantified effect of real refractive index assumptions on laser diffraction mechanical analysis. Soil Sci. Soc. Am. J. 2018, in press. [Google Scholar]
- Bonneville, S.; Van Cappellen, P.; Behrends, T. Microbial reduction of iron (III) oxyhydroxides: Effects of mineral solubility and availability. Chem. Geol. 2004, 212, 255–268. [Google Scholar] [CrossRef]
- Bonneville, S.; Behrends, T.; Van Cappellen, P. Solubility and dissimilatory reduction kinetics of iron (III) oxyhydroxides: A linear free energy relationship. Geochim. Cosmochim. Acta 2009, 73, 5273–5282. [Google Scholar] [CrossRef]
- Eusterhues, K.; Hädrich, A.; Neidhardt, J.; Küsel, K.; Keller, T.; Jandt, K.; Totsche, K. Reduction of ferrihydrite with adsorbed and coprecipitated organic matter: Microbial reduction by Geobacter bremensis vs. abiotic reduction by Na-dithionite. Biogeosciences 2014, 11, 4953. [Google Scholar] [CrossRef]
- Fiedler, S.; Vepraskas, M.J.; Richardson, J. Soil redox potential: Importance, field measurements, and observations. Adv. Agron. 2007, 94, 1–54. [Google Scholar]
- Husson, O. Redox potential (Eh) and pH as drivers of soil/plant/microorganism systems: A transdisciplinary overview pointing to integrative opportunities for agronomy. Plant Soil 2013, 362, 389–417. [Google Scholar] [CrossRef] [Green Version]
- Patrick, W.; Gambrell, R.; Faulkner, S. Redox measurements of soils. In Methods of Soil Analysis Part 3—Chemical Methods; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1255–1273. [Google Scholar]
- Sparks, D.L. Environmental Soil Chemistry; Academic Press: San Diego, CA, USA, 2003; p. 351. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Johnson, P.C. Extension of Nakagawa & Schielzeth’s R2GLMM to random slopes models. Methods Ecol. Evol. 2014, 5, 944–946. [Google Scholar] [PubMed]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Ponnamperuma, F.N. The chemistry of submerged soils. Adv. Agron. 1972, 24, 29–96. [Google Scholar]
- Barcellos, D. Biogeochemical Cycling of Iron and Carbon in Humid (Sub)tropical Forest Soils under Fluctuating Redox Conditions. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2018. [Google Scholar]
- Wang, Y.; Van Cappellen, P. A multicomponent reactive transport model of early diagenesis: Application to redox cycling in coastal marine sediments. Geochim. Cosmochim. Acta 1996, 60, 2993–3014. [Google Scholar] [CrossRef]
- Berg, P.; Rysgaard, S.; Thamdrup, B. Dynamic modeling of early diagenesis and nutrient cycling. A case study in an artic marine sediment. Am. J. Sci. 2003, 303, 905–955. [Google Scholar] [CrossRef]
- Boudreau, B.P. A method-of-lines code for carbon and nutrient diagenesis in aquatic sediments. Comput. Geosci. 1996, 22, 479–496. [Google Scholar] [CrossRef]
- Hall, S.J.; Liptzin, D.; Buss, H.L.; DeAngelis, K.; Silver, W.L. Drivers and patterns of iron redox cycling from surface to bedrock in a deep tropical forest soil: A new conceptual model. Biogeochemistry 2016, 130, 177–190. [Google Scholar] [CrossRef]
- Chen, C.; Meile, C.; Wilmoth, J.; Barcellos, D.; Thompson, A. Influence of PO2 on Iron Redox Cycling and Anaerobic Organic Carbon Mineralization in a Humid Tropical Forest Soil. Environ. Sci. Technol. 2018, 52, 7709–7719. [Google Scholar] [CrossRef] [PubMed]
- Wilmoth, J.L.; Moran, M.A.; Thompson, A. Transient O2 pulses direct Fe crystallinity and Fe(III)-reducer gene expression within a soil microbiome. Microbiome 2018, 6, 189. [Google Scholar] [CrossRef] [PubMed]
- Coward, E.K.; Thompson, A.T.; Plante, A.F. Iron-mediated mineralogical control of organic matter accumulation in tropical soils. Geoderma 2017, 306, 206–216. [Google Scholar] [CrossRef]
- Hodges, C.; King, E.; Pett-Ridge, J.; Thompson, A. Potential for Iron Reduction Increases with Rainfall in Montane Basaltic Soils of Hawaii. Soil Sci. Soc. Am. J. 2018, 82, 176–185. [Google Scholar] [CrossRef]
- Hodges, C.A. Drivers and Variability of Iron Reduction in Upland Soils. Master’s Thesis, University of Georgia, Athens, GA, USA, 2017. [Google Scholar]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter: Fourteen years on. Soil Biol. Biochem. 2017, 105, A3–A8. [Google Scholar] [CrossRef]
- Six, J.; Conant, R.; Paul, E.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Buettner, S.W.; Kramer, M.G.; Chadwick, O.A.; Thompson, A. Mobilization of colloidal carbon during iron reduction in basaltic soils. Geoderma 2014, 221, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Grybos, M.; Davranche, M.; Gruau, G.; Petitjean, P.; Pédrot, M. Increasing pH drives organic matter solubilization from wetland soils under reducing conditions. Geoderma 2009, 154, 13–19. [Google Scholar] [CrossRef]
- Keiluweit, M.; Nico, P.S.; Kleber, M.; Fendorf, S. Are oxygen limitations under recognized regulators of organic carbon turnover in upland soils? Biogeochemistry 2016, 127, 157–171. [Google Scholar] [CrossRef]
- Manabe, S.; Wetherald, R.T. Reduction in summer soil wetness induced by an increase in atmospheric carbon dioxide. Science 1986, 232, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Xu, M.; Qi, Y. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Glob. Chang. Biol. 2001, 7, 667–677. [Google Scholar] [CrossRef]
- Wang, Z.; Delaune, R.; Patrick, W.; Masscheleyn, P. Soil redox and pH effects on methane production in a flooded rice soil. Soil Sci. Soc. Am. J. 1993, 57, 382–385. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | SRO-Fe † mmol kg−1 | Total-Fe mmol kg−1 | Carbon (%) | Clay (%) | Sand (%) | Silt (%) | pH ‡ |
|---|---|---|---|---|---|---|---|
| Valley-1 | 336 (32) a | 1162 (10) a | 4.6 (0.1) a | 7 (0.6) a | 45 (3.6) a | 48 (1.9) a | 6.1 (0.2) a |
| Valley-2 | 305 (21) a | 1353 (12) b | 4.0 (0.2) a | 8 (0.7) a | 36 (2.7) a | 56 (2.1) b | 5.3 (0.2) b |
| Valley-3 | 195 (10) b | 1409 (14) c | 3.1 (0.1) b | 10 (0.9) b | 33 (2.6) b | 56 (2.3) b | 5.3 (0.1) b |
| Slope-1 | 178 (10) b | 1403 (12) c | 3.4 (0.3) b | 10 (0.6) b | 27 (3.4) b | 63 (2.0) c | 5.1 (0.2) bc |
| Slope-2 | 213 (7) b | 1503 (13) d | 3.1 (0.1) b | 12 (1.1) b | 28 (2.2) b | 60 (2.4) c | 4.8 (0.2) d |
| Slope-3 | 131 (11) c | 1434 (12) c | 3.4 (0.1) b | 8 (0.2) a | 45 (1.0) a | 47 (0.8) a | 5.1 (0.1) bc |
| Ridge-1 | 190 (17) b | 1428 (12) c | 4.5 (0.4) a | 6 (0.4) a | 43 (4.2) a | 51 (1.7) a | 4.9 (0.2) d |
| Ridge-2 | 227 (9) b | 1334 (13) b | 5.9 (0.1) c | 5 (0.5) a | 52 (2.6) ad | 43 (2.2) a | 4.8 (0.2) d |
| Ridge-3 | 206 (6) b | 1340 (12) b | 4.6 (0.1) a | 5 (0.4) a | 60 (4.8) cd | 36 (1.4) d | 5.0 (0.2) cd |
| Variable | Catenas | AIC a (R2c) b | Num. of Observ. c | Model (Significant Predictors in Bold at p < 0.05) |
|---|---|---|---|---|
| Eh | All | 3850.41 (0.944) | 324 | Eh = −0.7187 × Precipitation |
| O2 | 1 | 866.20 (0.992) | 240 | O2 = −0.0129 × Precipitation |
| θV | 1 | −880.34 (0.853) | 213 | θV = 4.894 × 10–4 × Precipitation |
| Variable | Catenas | AIC a (R2c) b | Num. of Observ. c | Model (Significant Predictors in Bold at p < 0.05) |
|---|---|---|---|---|
| FeII | All | 990.725 (0.727) | 110 | FeII = 2.130 × Media-FeIIIRR + 20.286 × pH |
| Catena 1 | 377.89 (0.789) | 39 | FeII = 2.4758 × Media-FeIIIRR − 4.3644 × O2_48 h | |
| Media-FeIIIRR | All | 413.32 (0.835) | 73 | Media-FeIIIRR ~ 0.1073 × FeII + 20.295 × θG − 0.845 × Eh_6 h |
| Shewa-FeIIIRR | All | 654.06 (0.810) | 110 | Shewa-FeIIIRR ~ 0.1202 × FeII + 21.221 × θG + 3.861 × pH |
| DOC | All | 718.68 (0.441) | 132 | DOC = 0.1909 × FeII + 12.47 × θG |
| Flux CO2 | Catena 1 | 117.2 (0.429) | 45 | F(CO2) = −10.995 × θV |
| Flux CH4 | Catena 1 | 126.01 (0.545) | 43 | F(CH4) = 2.782 × pH |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barcellos, D.; O’Connell, C.S.; Silver, W.; Meile, C.; Thompson, A. Hot Spots and Hot Moments of Soil Moisture Explain Fluctuations in Iron and Carbon Cycling in a Humid Tropical Forest Soil. Soil Syst. 2018, 2, 59. https://doi.org/10.3390/soilsystems2040059
Barcellos D, O’Connell CS, Silver W, Meile C, Thompson A. Hot Spots and Hot Moments of Soil Moisture Explain Fluctuations in Iron and Carbon Cycling in a Humid Tropical Forest Soil. Soil Systems. 2018; 2(4):59. https://doi.org/10.3390/soilsystems2040059
Chicago/Turabian StyleBarcellos, Diego, Christine S. O’Connell, Whendee Silver, Christof Meile, and Aaron Thompson. 2018. "Hot Spots and Hot Moments of Soil Moisture Explain Fluctuations in Iron and Carbon Cycling in a Humid Tropical Forest Soil" Soil Systems 2, no. 4: 59. https://doi.org/10.3390/soilsystems2040059