1. Introduction
Yams are herbaceous perennial vine plants in the genus
Dioscorea, comprising over 600 species [
1,
2]. They grow in the tropics and sub-tropics of Africa, the Caribbean, Latin America, Asia, and Oceania as a source of dietary food and ingredients for pharmaceuticals and traditional medicine. In West Africa, where over 92% of its global production occurs [
3], yam is involved in many key life ceremonies [
4].
Dioscorea alata, also known by the common names ‘water yam’ or ‘greater yam’, is one of the most important food yams. It is the world’s most widely distributed cultivated yam, though it is not cultivated on the same magnitude as
D. rotundata due to traditional bias that overlooks its nutritional and agronomic potential [
5].
End-use quality significantly influences the acceptance of yam varieties by farmers and consumers [
6]. In effect, the success of newly developed yam varieties depends not only on their agronomic attributes but also on their acceptability to consumers in terms of both sensory and utilization characteristics [
7]. The yam tuber has diverse uses as a result of the wide variation in organoleptic, culinary and nutritional properties, making some yam cultivars more appropriate for certain types of food preparation than others. Water yam cultivars with good eating qualities are characterized by high dry matter, starch, and amylose contents [
8]. The rapid change in colour of the tissue from white to yellow or brown after the tuber is cut, which is the result of the oxidation of polyphenols, influences its processing and utilization [
9]. Polyphenolic oxidation is also linked to bitterness, off-flavours and in some instances necessitates special preparation processes to make acceptable dishes [
10].
Boiled yam, pounded yam (also known as “fufu”), and “amala” (prepared from cooking and stirring of the fermented yam flour—“elubo”) are the three leading food forms of yams in West and Central African regions [
11,
12]. In this region,
Dioscorea rotundata (white yam) is preferred to water yam for “fufu” and “amala” due to its ease of dough formation when pounded. However, some collections and advanced breeding lines of water yam at the International Institute of Tropical Agriculture (IITA, Ibadan, Nigeria) have displayed the capacity to form good dough, similar to or even superior to that of some genotypes of
D. rotundata [
7]. Additionally, some water yam clones have higher total dietary fibre and amylose content than that reported for brown rice and whole wheat flour together with low sodium but high potassium content, indicating the nutritional role that water yam could play in managing chronic diseases including diabetes [
13].
This highlights the potential of water yam for food and nutritional security in West and Central Africa, particularly as it has a very wide adaptation, high genetic potential to produce fairly high yield under low to average soil fertility, early vigour for weed suppression, ease of propagation through the production of bulbils resulting in a high multiplication ratio, low post-harvest losses, good processing quality and high nutritive value [
5,
14].
An improvement in the food quality of
D. alata remains a key objective in yam breeding programmes [
7] and is critical for increasing the adoption level of newly developed varieties. To date, the genetic improvement efforts to develop water yam varieties with enhanced tuber quality attributes are mainly by conventional breeding strategies based on phenotypic records/data. This is, however, arduous and slow due to the lengthy screening process of identifying superior individuals from clonal populations [
5,
15]. In addition, the genetic basis of traits that define tuber quality has not received much attention. With the advent of molecular markers, tremendous progress has been made to understand the genetic diversity and relationships in
Dioscorea species including
D. alata (water yam) [
16]. The use of genotypic and phenotypic data has also proven to elucidate genetic diversity in different
Dioscorea spp. [
17].
The advancements in next-generation sequencing technologies have led to the rapid development of DNA-informed breeding techniques such as marker-assisted breeding and genomic selection through which many crops have recorded fast genetic gains. The increasing availability of molecular markers enables researchers to tag regions of the genome associated with specific phenotypes of interest in Quantitative trait locus (QTL) mapping and genome-wide association studies [
18]. The genetic mapping of loci underlying important tuber quality traits of water yam has not been conducted yet to assist selection decisions in the breeding process. Mapping based on genome-wide associations has become increasingly popular and powerful because of the emergence of more cost-effective, high-throughput genotyping platforms. Using molecular approaches, such as the candidate gene technique, to unravel the causal gene(s) would hasten efforts in introgressing tuber quality traits into preferred genetic backgrounds of
D. alata. The precision and speed of crop breeding have been improved by the evolution of genetic linkage and association mapping of the quantitative traits. This has been clearly demonstrated by Pétro et al. [
19] in water yam. Genomic resources for
D. alata are being rapidly developed [
20]. These include the recent pre-release of a chromosome-scale “v2” assembly of
Dioscorea alata see release notes and assembly [
20]. This genome sequence enables genome-wide association studies of key traits in water yam.
To elucidate the genetic factors for tuber dry matter content and oxidative browning, a diversity panel consisting of 100 water yam clones was genotyped by diversity arrays technology sequencing. While whole-genome sequencing provides the highest resolution, it still remains expensive for non-model species such as yams. A genome-wide association study was conducted to identify single nucleotide polymorphism loci or QTL regions and genes associated with tuber dry matter and oxidative browning. The SNP loci and associated candidate genes, when validated, would be a valuable resource for marker-assisted selection in the breeding process to develop new water yam varieties with acceptable end-user qualities.
3. Discussion
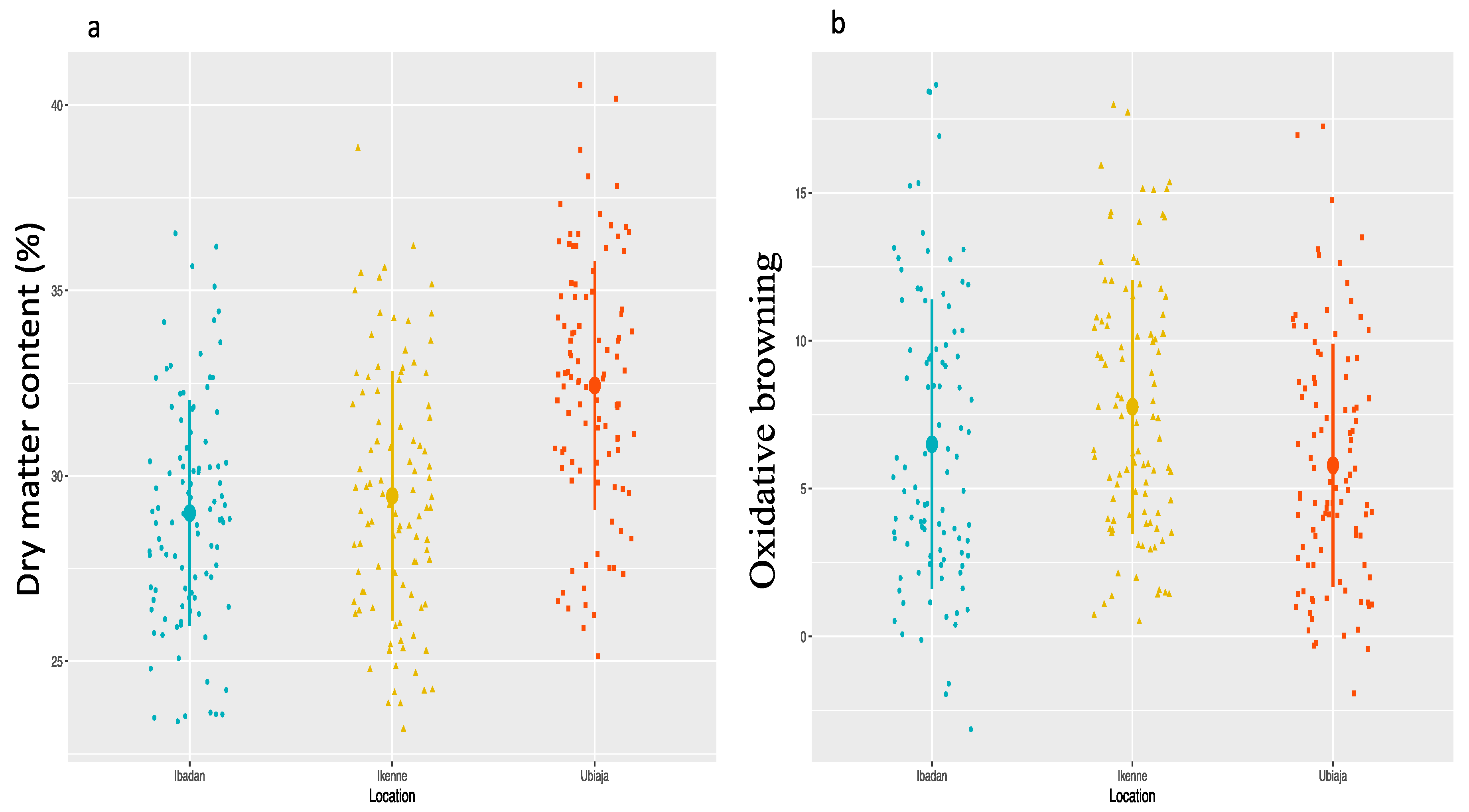
Dry matter content and oxidation properties of yam tubers are very important quality traits that influence the rate of adoption of new clones for cultivation and consumption. Yam improvement efforts worldwide, especially in West Africa, have tested several clones within yam populations for DMC and none or minimal oxidation of fresh tuber using conventional selective breeding based on phenotypic records [
8,
22,
23], an approach that is slow and arduous. For quality traits, DNA-based strategies such as GWAS reported in this study have advantages over the conventional selection breeding approach because it has the potential to fast-track the development and delivery of improved yam varieties with acceptable end-user attributes. The potential of GWAS to dissect complex traits has been proven in root and tuber crops such as cassava [
24,
25,
26] and potatoes [
27,
28]. The present GWAS sought to identify QTL (s) and putative candidate genes associated with genetic variation in DMC and OxB in water yam. The highly significant genotype variance for DMC content and OxB in the current
D. alata panel warrant further analysis for the dissection of the genetic basis of variation for these two traits.
Detailed knowledge of population structure and familial relationships (kinship) in the association panel is crucial to prevent sham associations in GWAS [
29]. Population structure and admixture for this population were reported in a previous study [
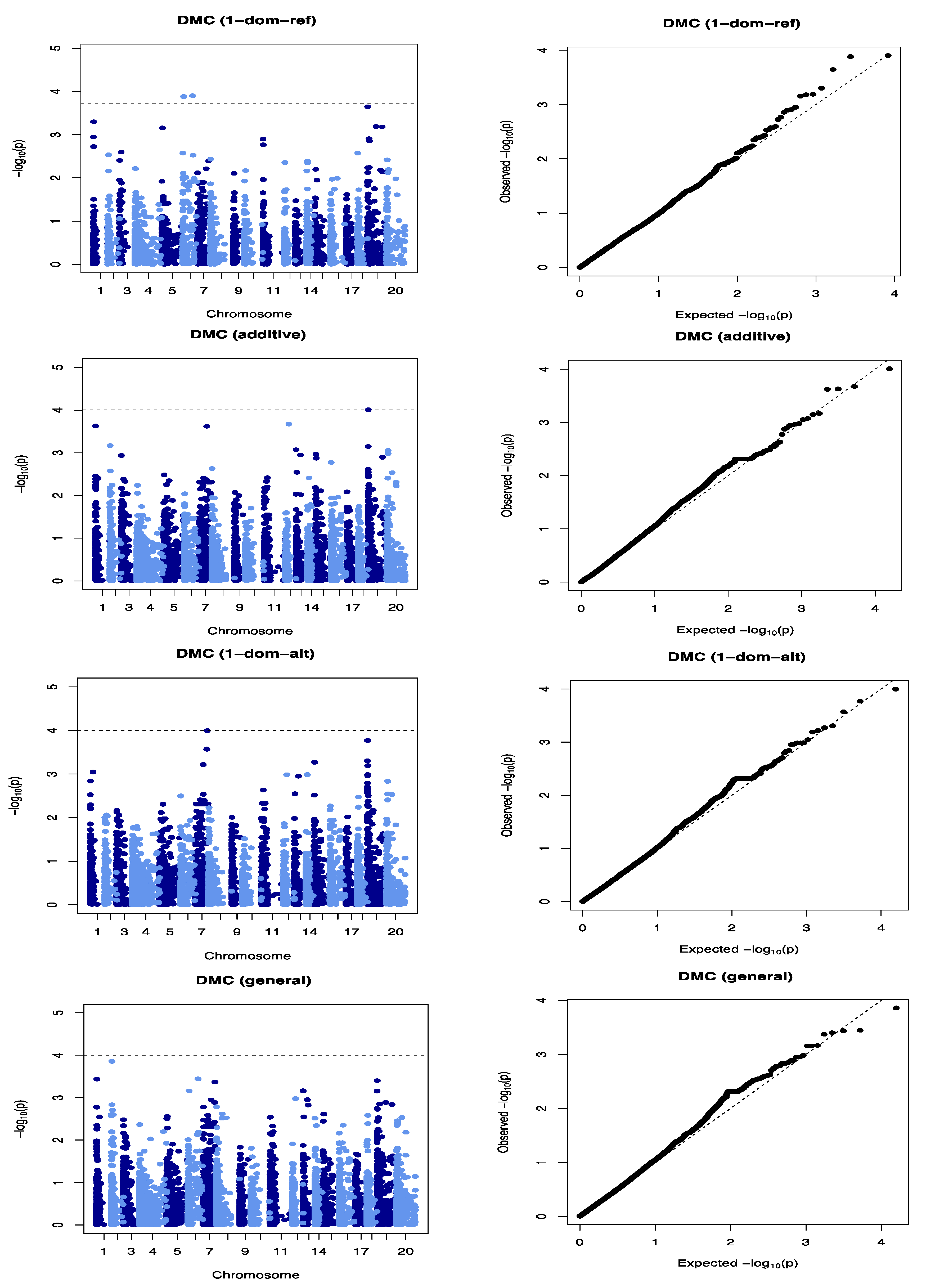
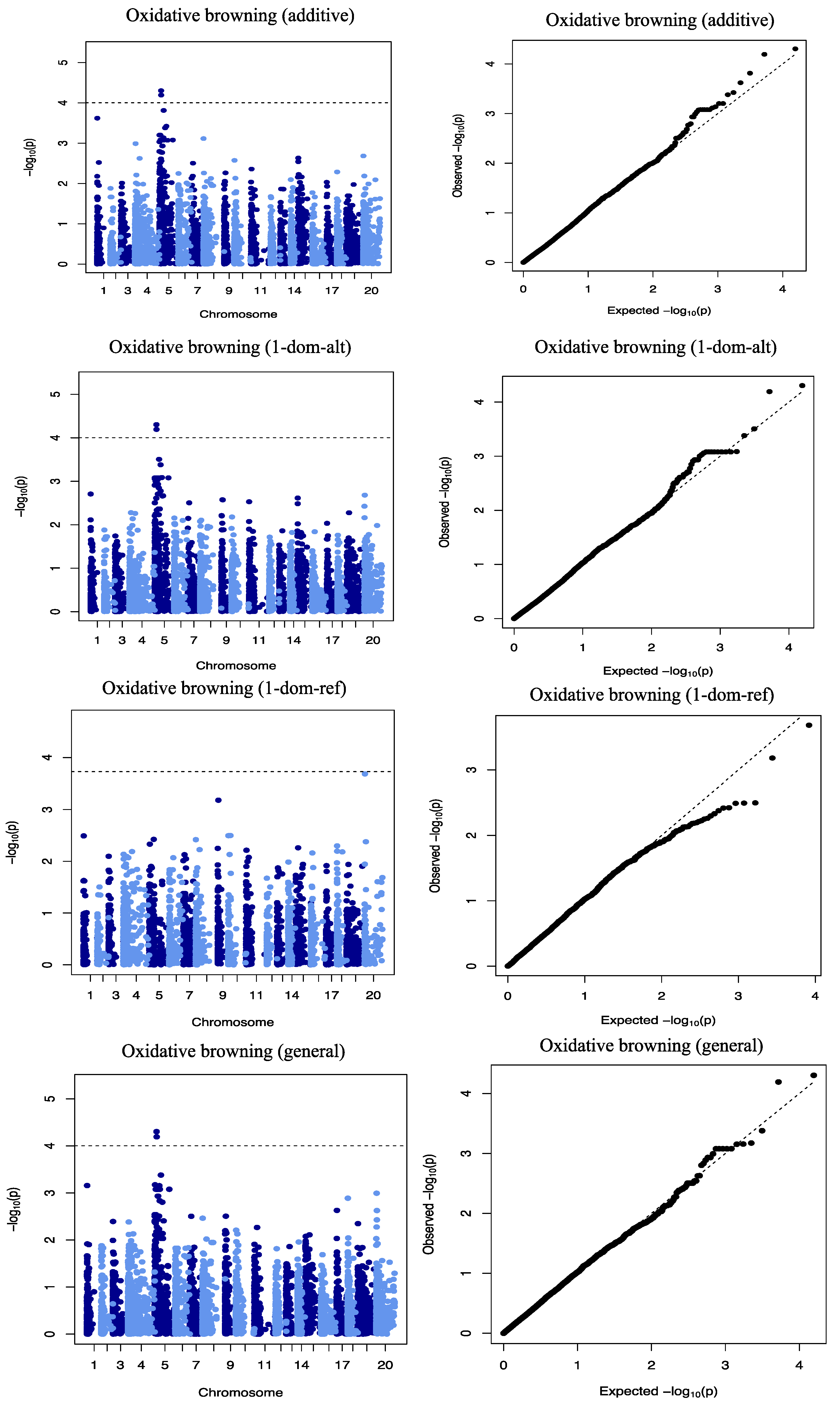
21]. The Q matrix (population structure) and K matrix (Admixture) were used as covariates in a mixed linear model for the association analysis to reduce false-positive associations. The reducing −log10 (
p-values) toward the expected level for both traits on the quantile–quantile plots is a sign that the model successfully accounted for population structure and familial relationships in the GWAS analysis.
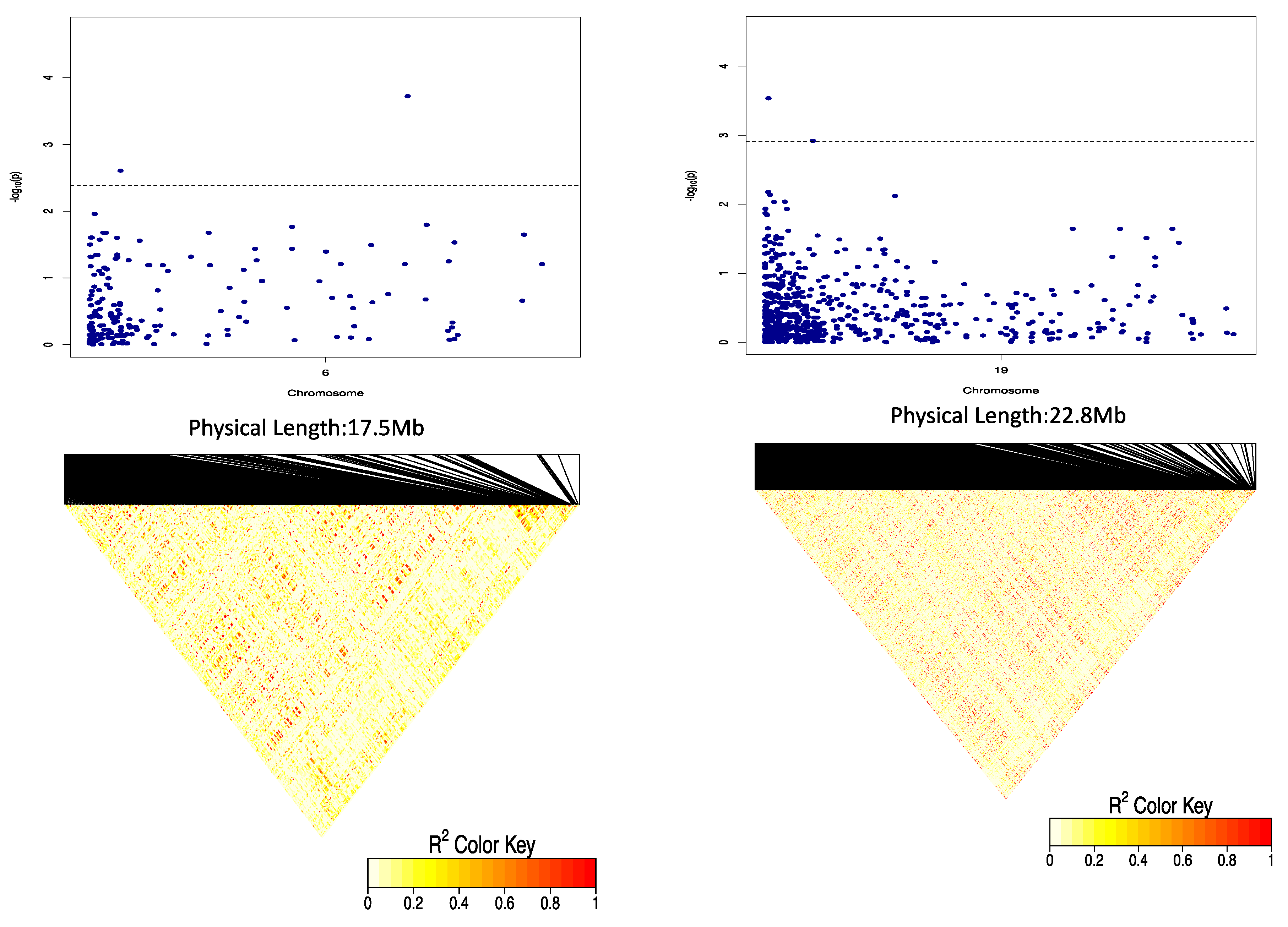
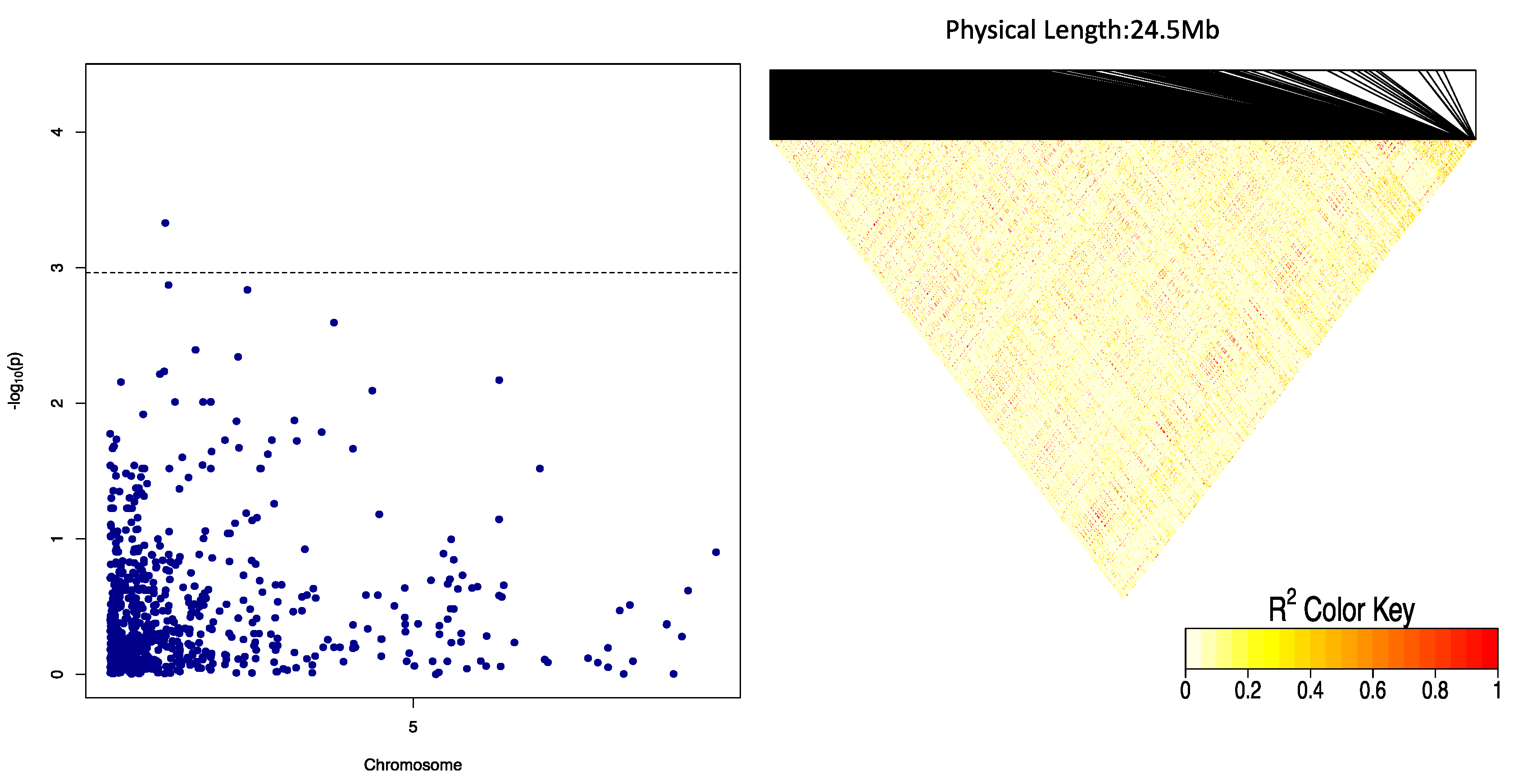
Three QTLs were identified to be associated with DMC, 2 of which showed significant QTL-by-Environment interactions (QEI). In a study to identify QTLs associated with cassava brown streak disease, Kayondo et al. [
30] reported similar results with QTLs identified at different locations and highlighted the effects of many factors such as the panel size, harvest time and environmental conditions influencing QTL identification. To address this, the best approach for increasing the resolution of associations of traits and QTLs is to combine multi-location genotypic and phenotypic scores from different diversity panels [
30,
31]
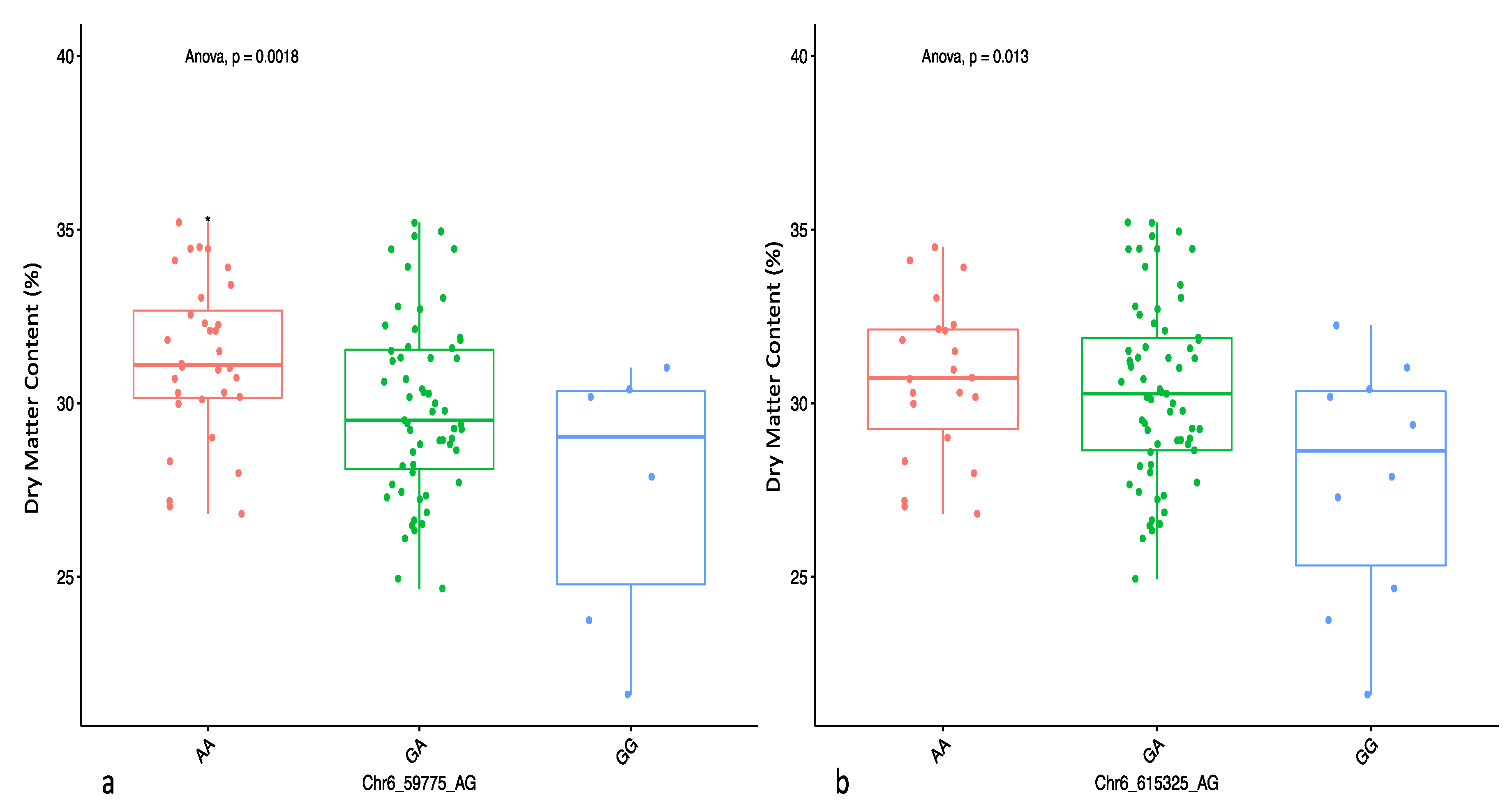
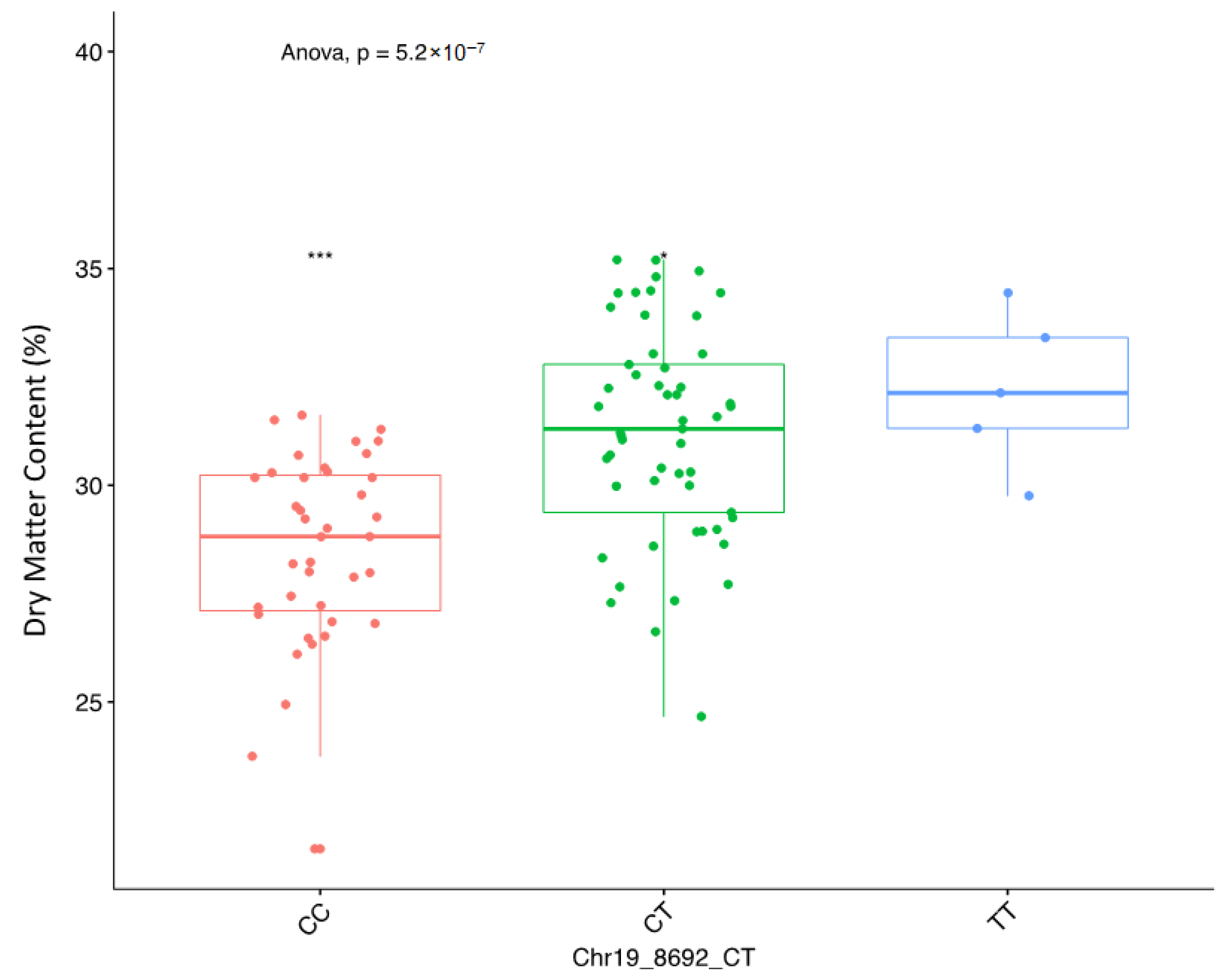
Using the marker effect, we observed allele AA on chromosome 6 and allele TT on chromosome 19 to be responsible for high DMC in the diversity panel used in the study. Information on marker effect through the segregation pattern is fundamental for marker validation and deployment in a breeding programme [
32,
33,
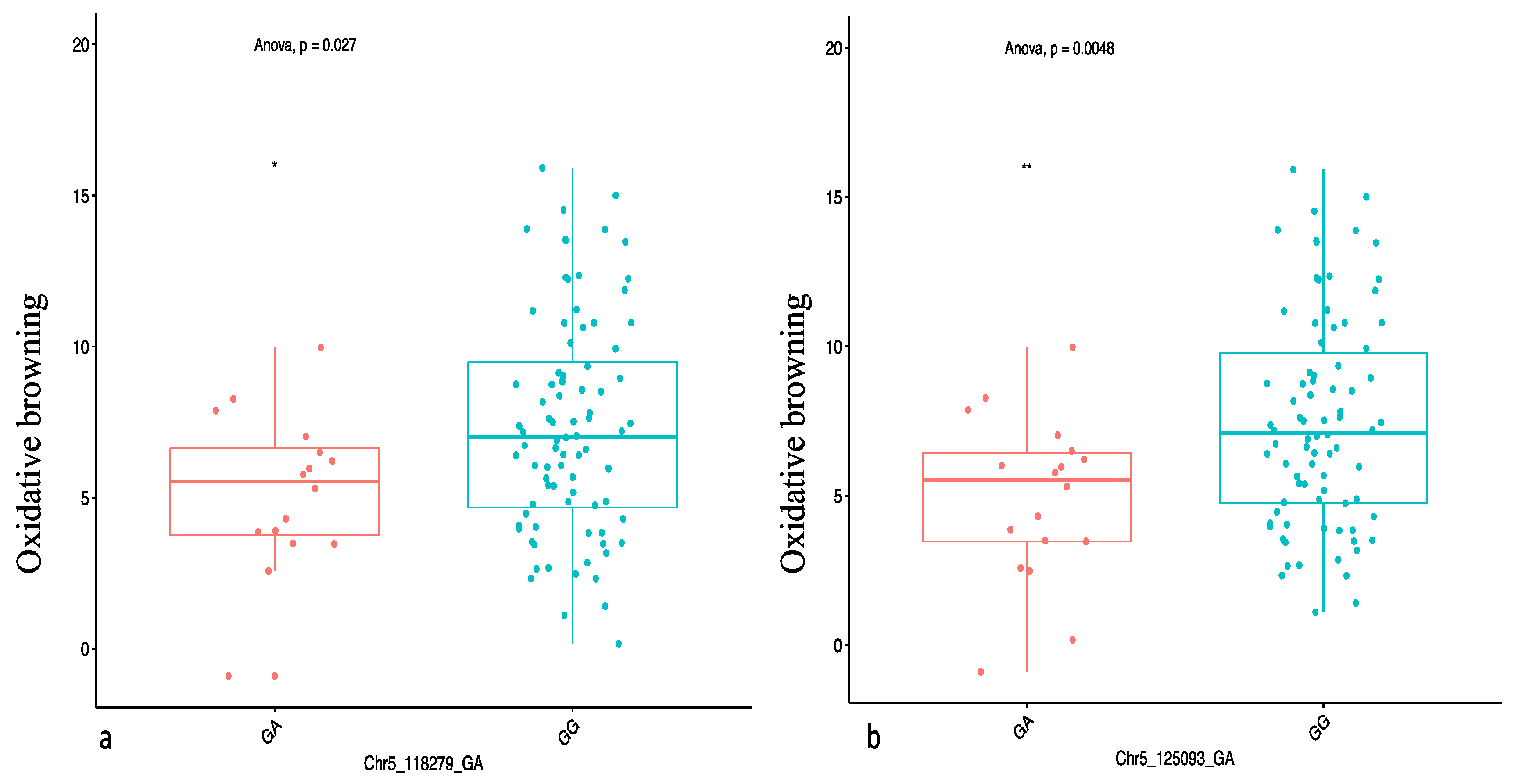
34]. We also identified the heterozygous allele GA to be significantly associated with low OxB.
Our study also identified putative candidate genes within the QTL regions of the targeted traits. A total of 26 putative candidate genes were detected upstream and downstream of the SNP associated with DMC, of which 3 genes (Serine/threonine-protein kinase, Tetratricopeptide-like helical domain superfamily and Glycoside hydrolase family 9) were reported to play important roles in DMC. Serine/threonine-protein kinase (SnRK1) was reported to participate in the process of starch and sugar biosynthesis in potatoes and stimulated glucose pyrophosphorylase [
35,
36,
37]. In potato (
Solanum tuberosum) and wheat (
Triticum aestivum), SnRK1 was reported to stimulate some enzymes in the starch biosynthesis pathways [
38,
39]. The Tetratricopeptide-like helical domain superfamily genes were reported to mediate protein–protein interactions and involved in the production of protein and starch that are the principal storage carbohydrates in plants [
40,
41]. The third important putative gene “Glycoside hydrolase family 9” was reported to be involved in diverse enzymatic metabolisms of carbohydrate compounds available in many plant tissues [
42].
Six putative candidate genes were identified within the QTL regions of the peak SNPs detected for OxB. Of these 6 genes, the Thioesterase domain (IPR006683) has been reported to play a major role in tuber oxidative browning pathway [
43]. The thioesterase domain is part of peroxisomes that contain soluble thioesterases (oxalate oxidase), which play a significant role in regulating flux of various substrates by releasing CoA from β-oxidation intermediates and products [
44]. The oxalate oxidase is an enzyme containing manganese that catalyse the oxidation of oxalate to carbon dioxide by reducing the oxygen to hydrogen peroxide [
45].
Tuber dry matter had a negative correlation with OxB. This desirable correlation suggests that selection for high DMC is expected to reduce enzymatic browning following oxidation of the tuber flesh. The genotypic factors greatly contribute for trait linkages and the negative correlations between the two phenotypic traits may be an indication of pleiotropism, genetic coupling and/or linkage disequilibrium with population structure effects [
46]. However, our GWAS result did not reveal co-localized SNP markers for both traits to confirm that such an association is due to pleiotropy. In any case, the observed trait association provided the opportunity to select superior yam clones that indeed offer increased tuber dry matter and less tuber flesh enzymatic oxidation simultaneously. Our study identified significant and functional SNPs established at the genome level and revealed the known genes, which showed the presence of the disequilibrium between SNP markers and causative variants of DMC and OxB within or near identified genes. It is plausible that the allelic variation for oxidation observed in our study, which is associated with browning, is the result of enzymatic activities in yam, which includes polyphenol oxidase and peroxidase activities that play a major role in the phenolic content of yam tubers [
47,
48].
The functions and characteristics of some identified genes have not been explored. However, the identified genes from this study may provide new intuition into the genetic fundamentals of DMC and OxB in D. alata.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}