
Effect of Ethylene and Abscisic Acid on Steroid and Triterpenoid Synthesis in Calendula officinalis Hairy Roots and Saponin Release to the Culture Medium



Abstract
:1. Introduction
2. Results
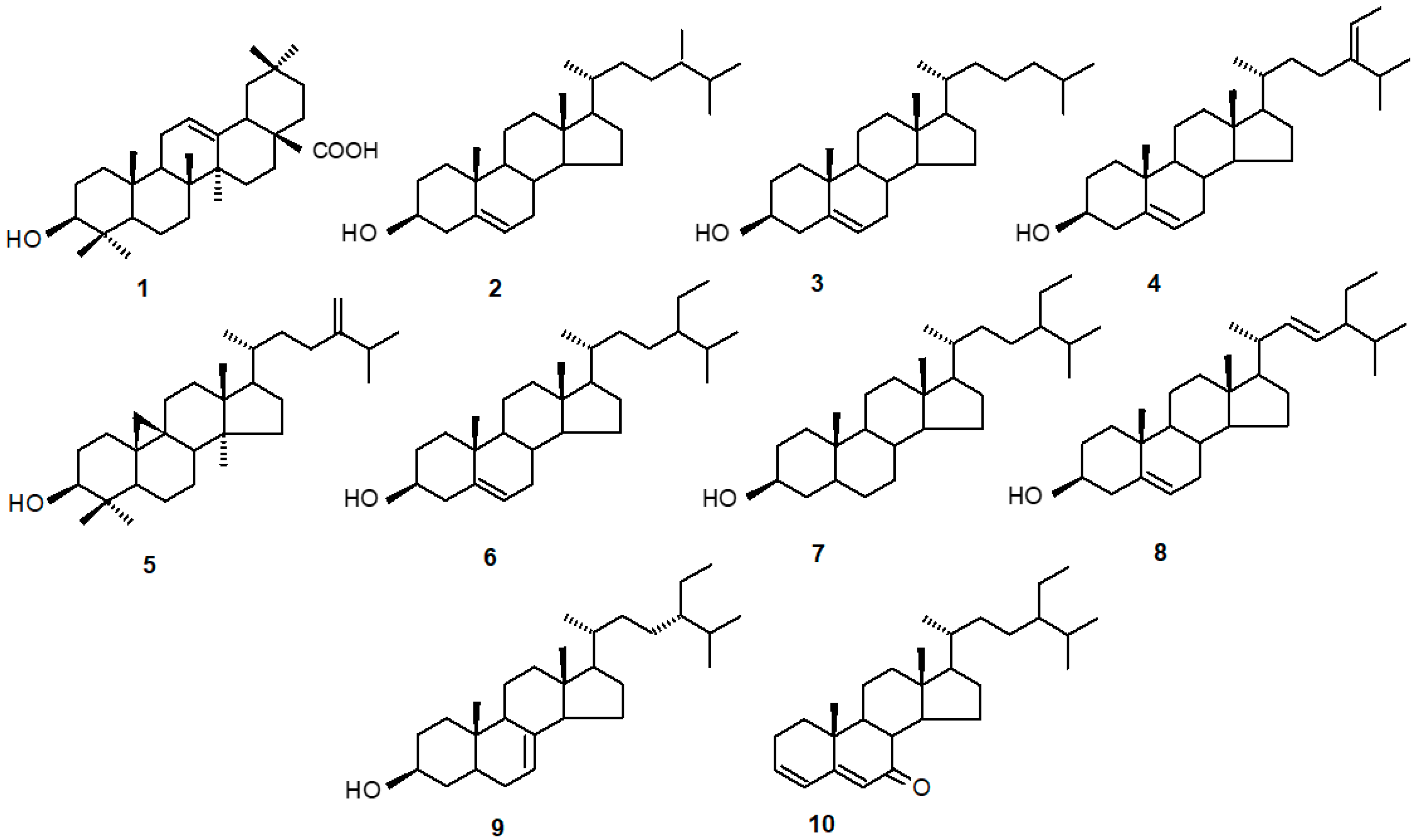
2.1. The Identification of Triterpenoids and Steroids in C. officinalis HRCs
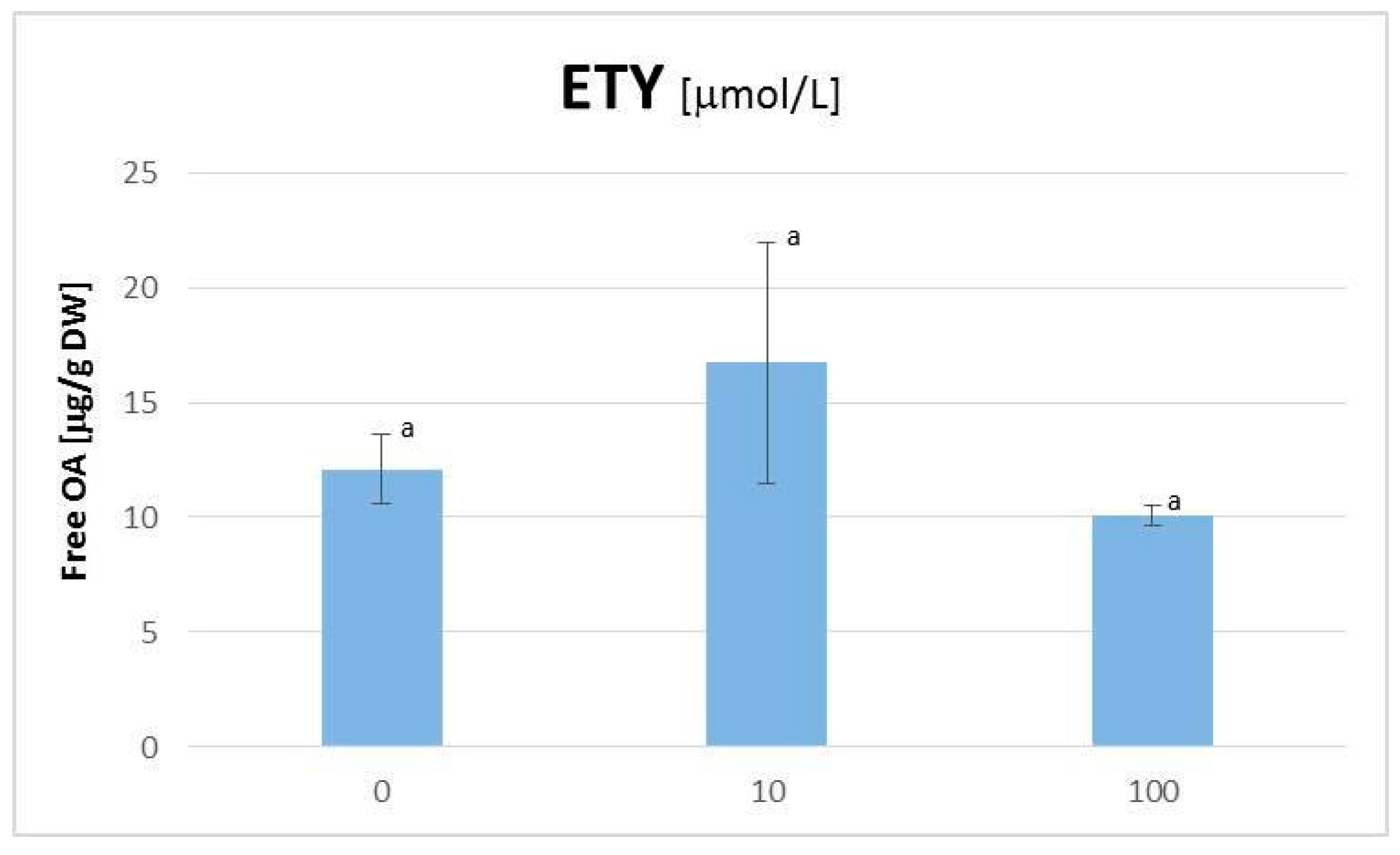
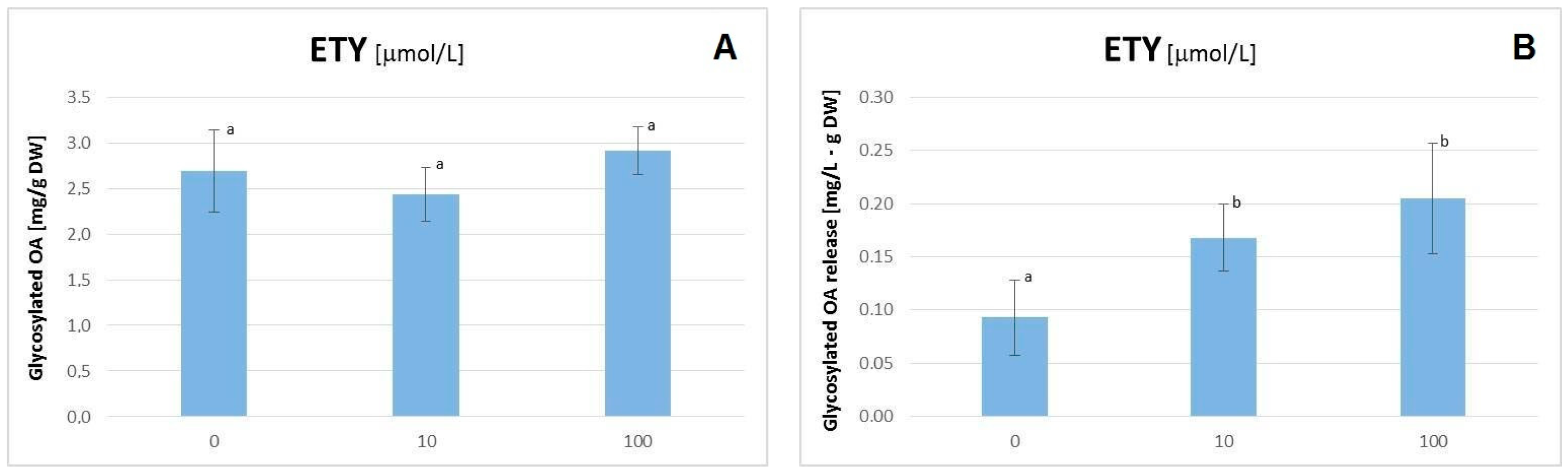
2.2. The Influence of ETYon the C. officinalis Hairy Root Growth and Productivity
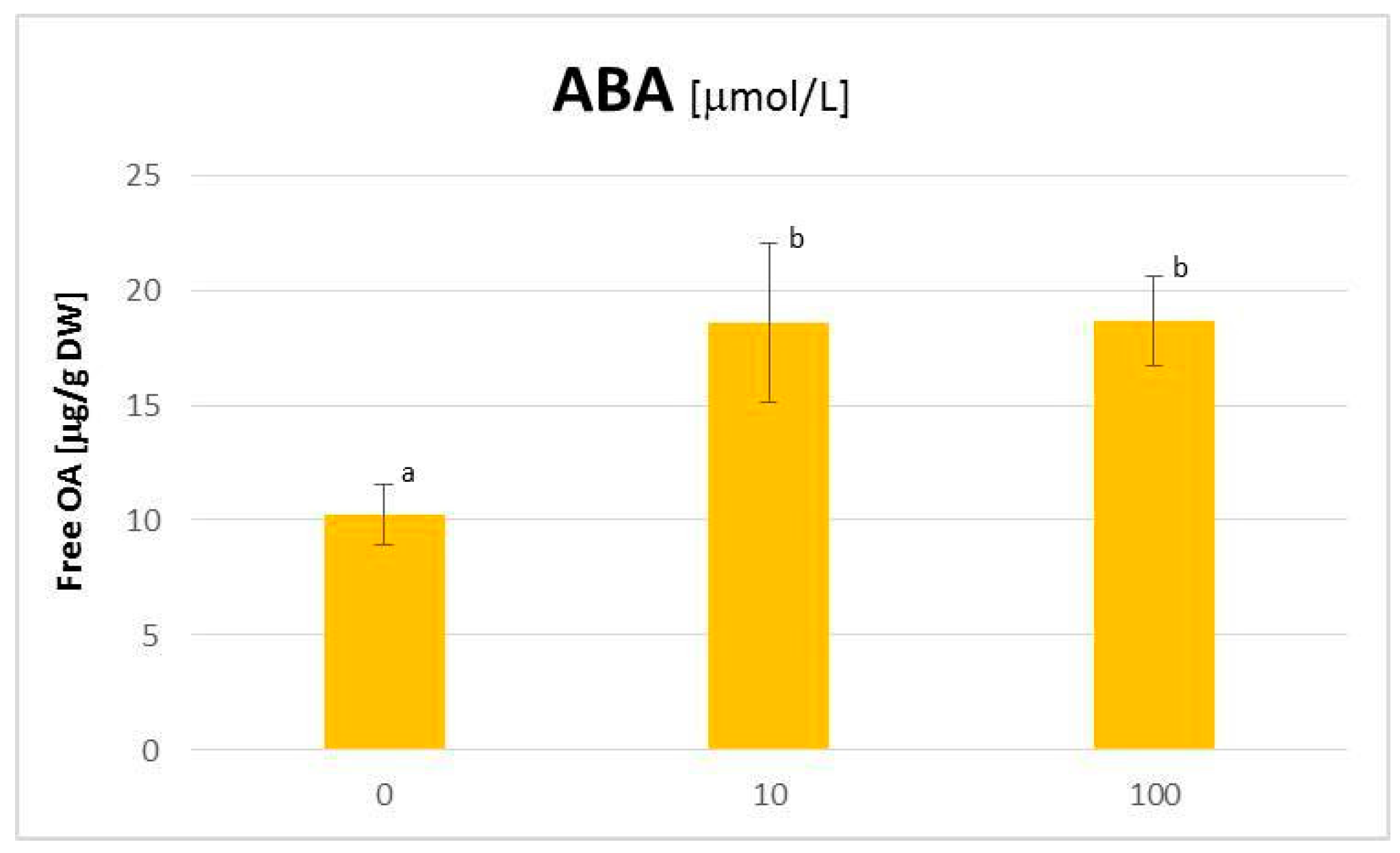
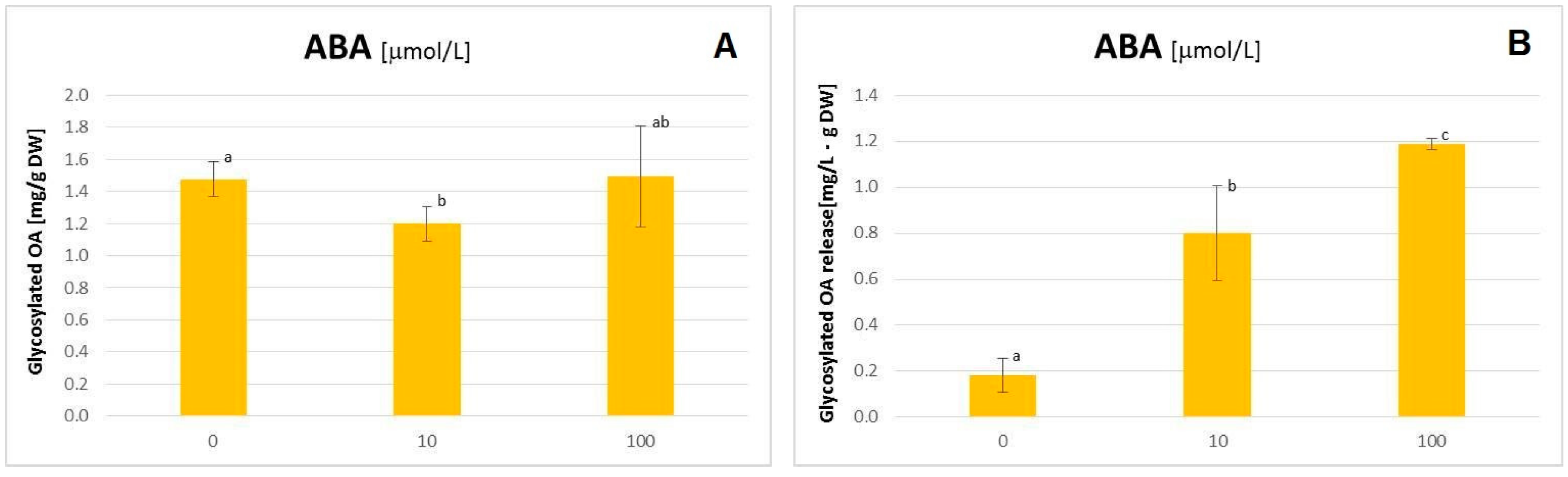
2.3. The Influence of ABA on the C. officinalis Hairy Root Growth and Productivity
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Elicitation of HRCs
4.3. Extraction of the Hairy Roots and the Culture Medium
4.4. Fractionation of Diethyl Ether Extracts
4.5. Hydrolysis of Methanol and n-Butanol Extracts
4.6. Quantification of OA by Gas Chromatography
4.7. Identification and Quantification of Steroids by Gas Chromatography–Mass Spectrometry
4.8. Statistical Analysis of Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ono, N.N.; Tian, L. The multiplicity of hairy root cultures: Prolific possibilities. Plant Sci. 2011, 180, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, M.I.; Agostini, E.; Ludwig-Müller, J.; Xu, J. Genetically transformed roots: From plant disease to biotechnological resource. Trends Biotechnol. 2012, 30, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.M.; Ritala, A.; Cardon, F. Hairy root cultures—A versatile tool with multiple applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, S.; Srivastava, V.; Rahman, L.U.; Kukreja, A.K. Hairy root biotechnology—Indicative timeline to understand missing links and future outlook. Protoplasma 2015, 252, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Chandra, R. Engineering secondary metabolite production in hairy roots. Phytochem. Rev. 2011, 10, 371–375. [Google Scholar] [CrossRef]
- Hussain, M.S.; Fareed, S.; Ansari, S.; Rahman, M.A.; Ahmad, I.Z.; Saeed, M. Current approaches toward production of secondary plant metabolites. J. Pharm. Bioallied Sci. 2012, 4, 10–20. [Google Scholar] [CrossRef]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.P.; Rajasekharan, P.E.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell Tissue Organ Cult. 2018, 132, 239–265. [Google Scholar] [CrossRef]
- Hussain, A.; Quarsi, I.A.; Nazir, H.; Ullah, I. Plant tissue culture: Current status and opportunities. In Recent Advances in Plant In Vitro Culture; Leva, A., Rinaldi, L.M.R., Eds.; InTech Open-Access: Rijeka, Croatia, 2012; pp. 1–29. [Google Scholar]
- Patel, H.; Krishnamurthy, R. Elicitors in plant tissue culture. J. Pharmacogn. Phytochem. 2013, 2, 60–65. [Google Scholar]
- Naik, P.M.; Al–Khayri, J.M. Abiotic and biotic elicitors–role in secondary metabolites production through in vitro culture of medicinal plants. In Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; InTech: Rijeka, Croatia, 2016; pp. 247–278. [Google Scholar]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Goleniowski, M.; Cusidó, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Alsoufi, A.S.M.; Pączkowski, C.; Szakiel, A.; Długosz, M. Effect of jasmonic acid and chitosan on triterpenoid production in Calendula officinalis hairyroot cultures. Phytochem. Lett. 2019, 31, 5–11. [Google Scholar] [CrossRef]
- Alsoufi, A.S.M.; Pączkowski, C.; Długosz, M.; Szakiel, A. Influence of selected abiotic factors on triterpenoid biosynthesis and saponin secretion in marigold (Calendula officinalis L.) in vitro hairy root cultures. Molecules 2019, 24, 2907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, C.M.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamińska, M. Role and activity of jasmonates in plants under in vitro conditions. Plant Cell Tissue Organ Cult. 2021, 146, 425–447. [Google Scholar] [CrossRef]
- Ali, M.B.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate and salicylic acid elicitation induces ginsenoside accumulation, enzymatic and non-enzymatic antioxidant in suspension culture Panax ginseng roots in bioreactors. Plant Cell Rep. 2006, 25, 613–620. [Google Scholar] [CrossRef]
- Chang, C. How do plant respond to ethylene and what is its importance? BMC Biol. 2016, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Int. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Moses, T.; Pollier, J.; Thevelein, J.M.; Goossens, A. Bioengineering of plant triterpenoids: From metabolic engineering of plants to synthetic biology in vivo and in vitro. New Phytol. 2013, 200, 27–43. [Google Scholar] [CrossRef]
- Rogowska, A.; Szakiel, A. Enhancement of phytosterol and triterpenoid production in plant hairy root cultures—Simultaneous stimulation or competition? Plants 2021, 10, 2028. [Google Scholar] [CrossRef]
- Długosz, M.; Wiktorowska, E.; Wiśniwska, A.; Pączkowski, C. Production of oleanolic acid glycosides by hairy root established cultures of Calendula officinalis L. Acta Biochim. Pol. 2013, 60, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Długosz, M.; Markowski, M.; Pączkowski, C. Source of nitrogen as a factor limiting saponin production by hairy root and suspension cultures of Calendula officinalis L. Acta Physiol. Plant. 2018, 40, 35. [Google Scholar] [CrossRef] [Green Version]
- Alsoufi, A.S.M.; Staśkiewicz, K.; Markowski, M. Alterations in oleanolic acid and sterol content in marigold (Calendula officinalis) hairy root cultures in response to stimulation by selected phytohormones. Acta Physiol. Plant. 2021, 43, 44. [Google Scholar] [CrossRef]
- Abidi, S.L. Chromatographic analysis of plant sterols in foods and vegetable oils. J. Chromatogr. A 2001, 935, 173–201. [Google Scholar] [CrossRef]
- Rogowska, A.; Styczyński, M.; Pączkowski, C.; Szakiel, A.; Pinheiro de Carvalho, M.Â.A. GC-MS analysis of steroids and triterpenoids occurring in leaves and tubers of Tamus edulis Lowe. Phytochem. Lett. 2019, 30, 231–234. [Google Scholar] [CrossRef]
- Woźniak, Ł.; Szakiel, A.; Pączkowski, C.; Marszałek, K.; Skąpska, S.; Kowalska, H.; Jędrzejczak, R. Extraction of triterpenic acids and phytosterols from apple pomace with supercritical carbon dioxide: Impact of process parameters, modeling of kinetics, and scaling-up study. Molecules 2018, 23, 2790. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Ma, Y.; Xu, T.; Cu, B.; Liu, Y.; Guo, Z. Effects of abscisic acid, gibberelin, ethylene and their interactions on production of phenolic acids in Salvia miltiorhhiza Bunge hairy roots. PLoS ONE 2013, 8, e72806. [Google Scholar] [CrossRef] [Green Version]
- Rogowska, A.; Szakiel, A. The role of sterols in plant response to abiotic stress. Phyt. Rev. 2020, 19, 1525–1538. [Google Scholar] [CrossRef]
- Aboobucker, S.I.; Suza, W.P. Why do plants convert sitosterol to stigmasterol? Front. Plant Sci. 2019, 10, 354. [Google Scholar] [CrossRef] [Green Version]
- De, D.; De, B. Elicitation of diosgenin production in Dioscorea floribunda by ethylene-generating agent. Fitoterapia 2005, 76, 153–156. [Google Scholar] [CrossRef]
- Bae, K.H.; Choi, Y.E.; Shin, C.G.; Kim, Y.Y.; Kim, Y.S. Enhanced ginsenoside productivity by combination of ethephon and methyl jasmoante in ginseng (Panax ginseng CA Meyer) adventitious root cultures. Biotechnol. Lett. 2006, 28, 1163–1166. [Google Scholar] [CrossRef] [PubMed]
- Saw, N.M.M.T.; Riedel, H.; Kütük, O.; Ravichanran, K.; Smetanska, I. Effect of elicitors and precursors on the synthesis of anthocyanin in grape Vitisvinifers cell cultures. Energy Res. J. 2010, 1, 189–192. [Google Scholar]
- Qiao, J.; Luo, Z.; Li, Y.; Ren, G.; Liu, C.; Ma, X. Effect of abscisic acid on accumulation of five active components in root of Glycyrrhiza uralensis. Molecules 2017, 22, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, R.; Berli, F.J.; Bottini, R.; Piccoli, P. Acclimation mechanisms elicited by sprayed abscisic acid, solar UV-B and water deficit in leaf tissues of field-grown grapevines. Plant Physiol. Biochem. 2015, 91, 56–60. [Google Scholar] [CrossRef]
- Rai, M.K.; Shekhawat, N.S.; Harish; Gupta, A.K.; Phulwaria, M.; Ram, K.; Jaiswal, U. The role of abscisic acid in plant tissue culture: A review of recent progress. Plant Cell Tissue Organ Cult. 2011, 106, 179–190. [Google Scholar] [CrossRef]
- Vaičiukynė, M.; Žiauka, J.; Žūkienė, R.; Vertelkaitė, L.; Kuusienė, S. Abscisic acid promotes root system development in birch tissue culture: A comparison to aspen culture and conventional rooting-related growth regulators. Physiol. Plant 2019, 165, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Markowski, M.; Długosz, M.; Szakiel, A.; Durli, M.; Poinsignon, S.; Bouguet-Bonnet, S.; Henry, M. Increased synthesis of a new oleanane-type saponin in hairy roots of marigold (Calendula officinalis) after treatment with jasmonic acid. Nat. Prod. Res. 2019, 33, 1218–1222. [Google Scholar] [CrossRef]
- Sharma, P.; Padh, H.; Shrivastava, N. Hairy root cultures: A suitable biological system for studying metabolic pathways in plants. Eng. Life Sci. 2013, 13, 62–75. [Google Scholar] [CrossRef]
- Da Silva Magedans, Y.V.; Philips, M.A. Production of plant bioactive triterpenoid saponins: From metabolites to genes and back. Phytochem. Rev. 2020, 20, 461–482. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ETY Concentration (µmol/L) | ||
|---|---|---|---|
| 0 | 10 | 100 | |
| campesterol | 52.6 ± 1.6 a | 41.3 ± 7.3 a | 58.1 ± 3.0 a |
| cholesterol | 2.9 ± 0.4 a | 6.1 ± 0.9 b | 7.2 ± 0.5 b |
| isofucosterol | 6.7 ± 0.8 a | 3.3 ± 1.1 b | 7.8 ± 0.7 a |
| 24-methylenecycloartanol | 11.4 ± 2.5 a | 11.5 ± 1.9 a | 7.1 ± 0.7 b |
| sitosterol | 144.0 ± 25.4 a | 70.7 ± 3.1 b | 71.7 ± 7.9 b |
| sitostanol | 15.6 ± 2.9 a | 10.7 ± 1.4 a | 9.5 ± 0.8 a |
| stigmasterol | 460.3 ± 4.2 a | 398.9 ± 53.4 a | 581.8 ± 21.9 b |
| stigmast-7-en-3-ol | 8.1 ± 2.5 a | 3.0 ± 0.7 b | 2.8 ± 0.2 b |
| tremulone | 13.2 ± 3.3 a | 9.7 ± 2.4 a | 2.1 ± 0.6 b |
| Total | 714.7 | 555.2 | 748.1 |
| Compound | ABA Concentration (µmol/L) | ||
|---|---|---|---|
| 0 | 10 | 100 | |
| campesterol | 43.8 ± 1.1 a | 33.6 ± 3.2 b | 35.5 ± 2.0 b |
| cholesterol | 4.0 ± 0.4 a | 8.3 ± 2.6 b | 7.8 ± 2.1 b |
| isofucosterol | 9.8 ± 1.4 a | 5.0 ± 2.9 a | 2.8 ± 1.8 b |
| 24-methylenecycloartanol | 12.4 ± 1.1 a | 19.8 ± 1.9 b | 24.4 ± 2.7 b |
| sitosterol | 119.4 ± 13.8 a | 103.1 ± 8.9 a | 99.3 ± 20.6 a |
| sitostanol | 18.4 ± 1.4 a | 27.5 ± 1.3 b | 25.4 ± 8.8 a |
| stigmasterol | 460.9 ± 13.4 a | 355.5 ± 29.3 b | 370.7 ± 18.4 b |
| stigmast-7-en-3-ol | 7.9 ± 2.0 a | 8.6 ± 3.1 a | 11.0 ± 4.3 a |
| tremulone | 15.3 ± 5.9 a | 34.8 ± 4.5 b | 6.4 ± 2.4 c |
| Total | 691.9 | 596.1 | 583.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markowski, M.; Alsoufi, A.S.M.; Szakiel, A.; Długosz, M. Effect of Ethylene and Abscisic Acid on Steroid and Triterpenoid Synthesis in Calendula officinalis Hairy Roots and Saponin Release to the Culture Medium. Plants 2022, 11, 303. https://doi.org/10.3390/plants11030303
Markowski M, Alsoufi ASM, Szakiel A, Długosz M. Effect of Ethylene and Abscisic Acid on Steroid and Triterpenoid Synthesis in Calendula officinalis Hairy Roots and Saponin Release to the Culture Medium. Plants. 2022; 11(3):303. https://doi.org/10.3390/plants11030303
Chicago/Turabian StyleMarkowski, Michał, Abdulwadood Shakir Mahmood Alsoufi, Anna Szakiel, and Marek Długosz. 2022. "Effect of Ethylene and Abscisic Acid on Steroid and Triterpenoid Synthesis in Calendula officinalis Hairy Roots and Saponin Release to the Culture Medium" Plants 11, no. 3: 303. https://doi.org/10.3390/plants11030303