
A Nosocomial Outbreak of Invasive Listeriosis in An Italian Hospital: Epidemiological and Genomic Features

 , , and
, , and 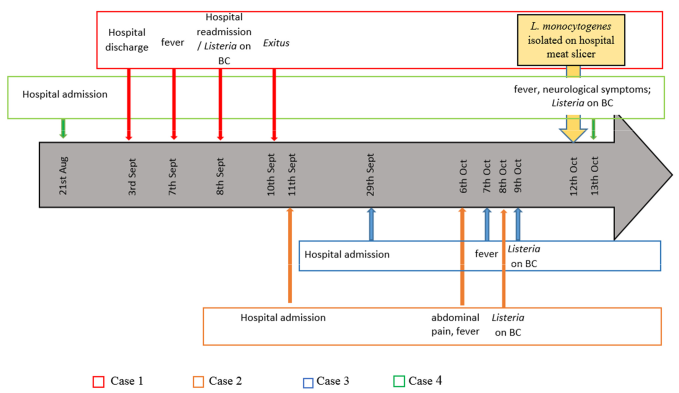{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Description of the Clinical Cases
2.1.1. Case 1
2.1.2. Case 2
2.1.3. Case 3
2.1.4. Case 4
2.2. Epidemiological Investigation Results
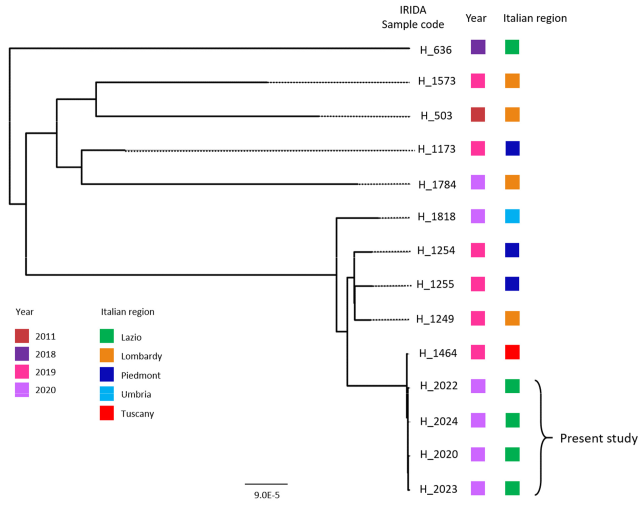
2.3. Bacterial Identification and Genomic Analysis
2.4. In-Depth Study of L. monocytogenes ST451 in a National and International Context
3. Discussion
4. Materials and Methods
4.1. Outbreak Investigation and Sampling
4.2. Microbiological Methods for Bacterial Identification and Characterization
4.3. Whole Genome Sequencing of Bacterial Isolates
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kathariou, S. Listeria monocytogenes Virulence and Pathogenicity, a Food Safety Perspective. J. Food Prot. 2002, 65, 1811–1829. [Google Scholar] [CrossRef] [PubMed]
- Lorber, B. Listeria monocytogenes. In Principles and Practice of Infectious Diseases, 5th ed.; Mandell, G.L., Bennett, J.E., Dolin, R., Eds.; Churchill Livingstone: New York, NY, USA, 2000. [Google Scholar]
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes Persistence in Food-Associated Environments: Epidemiology, Strain Characteristics, and Implications for Public Health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef]
- Desai, A.N.; Anyoha, A.; Madoff, L.C.; Lassmann, B. Changing epidemiology of Listeria monocytogenes outbreaks, sporadic cases, and recalls globally: A review of ProMED reports from 1996 to 2018. Int. J. Infect. Dis. 2019, 84, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Tortajada, C.; Porta, R.; Riba, M.; Santoma, M.J.; Palacín, E.; Español, M. Nosocomial outbreak due to Listeria monocytogenes in a neonatal unit. Enferm. Infecc. Microbiol. Clin. 2012, 30, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Schuchat, A.; Lizano, C.; Broome, C.V.; Swaminathan, B.; Kim, C.; Winn, K. Outbreak of neonatal listeriosis associated with mineral oil. Pediatr. Infect. Dis. J. 1991, 10, 183–189. [Google Scholar] [CrossRef]
- Elsner, H.A.; Kaulfers, P.M.; Tenschert, W.; Fischer, L. Nosocomial infections by Listeria monocytogenes: Analysis of a cluster of septicemias in immunocompromised patients. Infection 1997, 25, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, H.; Jones, D. Listeria. Bergey’s Man. Syst. Bacteriol. 1986, 2, 1235–1245. [Google Scholar]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the Major Listeria monocytogenes Serovars by Multiplex PCR. J. Clin. Microbiol. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Ragon, M.; Wirth, T.; Hollandt, F.; Lavenir, R.; Lecuit, M.; Monnier, A.L.; Brisse, S. A new perspective on Listeria monocytogenes evolution. PLoS Pathog. 2008, 4, e1000146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, A.; Tourdjman, M.; Leclercq, A.; Hamelin, E.; Laurent, E.; Fredriksen, N.; van Cauteren, D.; Bracq-Dieye, H.; Thouvenot, P.; Vales, G.; et al. Real-time whole-genome sequencing for surveillance of Listeria monocytogenes, France. Emerg. Infect. Dis. 2017, 23, 1462–1470. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Maiden, M.C.J. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef] [Green Version]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef]
- Jean, D.; Croize, J.; Hirtz, P.; Legeais, C.; Pelloux, I.; Favier, M.; Mallaret, M.R.; Le Noc, P.; Rambaud, P. Infection nosocomiale à Listeria monocytogenes en maternité. Arch. Fr. Pediatr. 1991, 48, 419–422. [Google Scholar]
- Simmons, B.T.; Dixon, R.E.; Bennett, J.V. Listeria monocytogenes infection in a renal transplant unit in an Alabama hospital. CDC EPI; 1980; 80, p. 16-2. [Google Scholar]
- Gaul, L.K.; Farag, N.H.; Shim, T.; Kingsley, M.A.; Silk, B.J.; Hyytia-Trees, E. Hospital-Acquired Listeriosis Outbreak Caused by Contaminated Diced Celery—Texas, 2010. Clin. Infect. Dis. 2013, 56, 20–26. [Google Scholar] [CrossRef]
- Shetty, A.; McLauchlin, J.; Grant, K.; O’Brien, D.; Howard, T.; Davies, E.M. Outbreak of Listeria monocytogenes in an oncology unit associated with sandwiches consumed in hospital. J. Hosp. Infect. 2009, 72, 332–336. [Google Scholar] [CrossRef]
- Johnsen, B.O.; Lingaas, E.; Torfoss, D.; Strøm, E.H.; Nordøy, I. A large outbreak of Listeria monocytogenes infection with short incubation period in a tertiary care hospital. J. Infect. 2010, 61, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Rietberg, K.; Lloyd, J.; Melius, B.; Wyman, P.; Treadwell, R.; Olson, G.; Kang, M.G.; Duchin, J.S. Outbreak of Listeria monocytogenes infections linked to a pasteurized ice cream product served to hospitalized patients. Epidemiol. Infect. 2016, 144, 2728–2731. [Google Scholar] [CrossRef] [Green Version]
- Mazengia, E.; Kawakami, V.; Rietberg, K.; Kay, M.; Wyman, P.; Skilton, C.; Aberra, A.; Boonyaratanakornkit, J.; Limaye, A.P.; Pergam, S.A.; et al. Hospital-acquired listeriosis linked to a persistently contaminated milkshake machine. Epidemiol. Infect. 2017, 145, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Jacks, A.; Pihlajasaari, A.; Vahe, M.; Myntti, A.; Kaukoranta, S.S.; Elomaa, N.; Salmenlinna, S.; Rantala, L.; Lahti, K.; Huusko, S.; et al. Outbreak of hospital-acquired gastroenteritis and invasive infection caused by Listeria monocytogenes, Finland, 2012. Epidemiol. Infect. 2016, 144, 2732–2742. [Google Scholar] [CrossRef] [Green Version]
- Lachmann, R.; Halbedel, S.; Adler, M.; Becker, N.; Allerberger, F.; Holzer, A.; Boone, I.; Falkenhorst, G.; Kleta, S.; Al Dahouk, S.; et al. Nationwide outbreak of invasive listeriosis associated with consumption of meat products in health care facilities, Germany, 2014–2019. Clin. Microbiol. Infect. 2020. [Google Scholar] [CrossRef]
- Li, Z.; Pérez-Osorio, A.; Wang, Y.; Eckmann, K.; Glover, W.A.; Allard, M.W.; Brown, E.W.; Chen, Y. Whole genome sequencing analyses of Listeria monocytogenes that persisted in a milkshake machine for a year and caused illnesses in Washington State. BMC Microbiol. 2017, 17, 134. [Google Scholar] [CrossRef]
- Poimenidou, S.V.; Chrysadakou, M.; Tzakoniati, A.; Bikouli, V.C.; Nychas, G.J.; Skandamis, P.N. Variability of Listeria monocytogenes strains in biofilm formation on stainless steel and polystyrene materials and resistance to peracetic acid and quaternary ammonium compounds. Int. J. Food Microbiol. 2016, 237, 164–171. [Google Scholar] [CrossRef]
- Ronner, A.B.; Wong, A.C.L. Biofilm Development and Sanitizer Inactivation of Listeria monocytogenes and Salmonella typhimurium on Stainless Steel and Buna-n Rubber. J. Food Prot. 1993, 56, 750–758. [Google Scholar] [CrossRef]
- Kushwaha, K.; Muriana, P.M. Adherence characteristics of Listeria strains isolated from three ready-to-eat meat processing plants. J. Food Prot. 2009, 72, 2125–2131. [Google Scholar] [CrossRef]
- Gaillard, J.L.; Berche, P.; Frehel, C.; Gouln, E.; Cossart, P. Entry of L. monocytogenes into cells is mediated by internalin, a repeat protein reminiscent of surface antigens from gram-positive cocci. Cell 1991, 65, 1127–1141. [Google Scholar] [CrossRef]
- Mengaud, J.; Ohayon, H.; Gounon, P.; Mège, R.-M.; Cossart, P. E-Cadherin Is the Receptor for Internalin, a Surface Protein Required for Entry of L. monocytogenes into Epithelial Cells. Cell 1996, 84, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Bierne, H.; Sabet, C.; Personnic, N.; Cossart, P. Internalins: A complex family of leucine-rich repeat-containing proteins in Listeria monocytogenes. Microbes Infect. 2007, 9, 1156–1166. [Google Scholar] [CrossRef]
- Matereke, L.T.; Okoh, A.I. Listeria monocytogenes Virulence, Antimicrobial Resistance and Environmental Persistence: A Review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef]
- Lecuit, M.; Vandormael-Pournin, S.; Lefort, J.; Huerre, M.; Gounon, P.; Dupuy, C.; Babinet, C.; Cossart, P. A transgenic model for listeriosis: Role of internalin in crossing the intestinal barrier. Science 2001, 292, 1722–1725. [Google Scholar] [CrossRef] [Green Version]
- Halbedel, S.; Prager, R.; Fuchs, S.; Trost, E.; Werner, G.; Flieger, A. Whole-Genome Sequencing of Recent Listeria monocytogenes Isolates from Germany Reveals Population Structure and Disease Clusters. Am. Soc. Microbiol 2018, 56, e00119-18. [Google Scholar] [CrossRef] [Green Version]
- Kuch, A.; Goc, A.; Belkiewicz, K.; Filipello, V.; Ronkiewicz, P.; Gołębiewska, A.; Wróbel, I.; Kiedrowska, M.; Waśko, I.; Hryniewicz, W.; et al. Molecular diversity and antimicrobial susceptibility of Listeria monocytogenes isolates from invasive infections in Poland (1997–2013). Sci. Rep. 2018, 8, 14562. [Google Scholar] [CrossRef]
- Ebner, R.; Stephan, R.; Althaus, D.; Brisse, S.; Maury, M.; Tasara, T. Phenotypic and genotypic characteristics of Listeria monocytogenes strains isolated during 2011–2014 from different food matrices in Switzerland. Food Control. 2015, 57, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Dreyer, M.; Aguilar-Bultet, L.; Rupp, S.; Guldimann, C.; Stephan, R.; Schock, A.; Otter, A.; Schüpbach, G.; Brisse, S.; Lecuit, M.; et al. Listeria monocytogenes sequence type 1 is predominant in ruminant rhombencephalitis. Sci. Rep. 2016, 6, 36419. [Google Scholar] [CrossRef] [Green Version]
- Cabal, A.; Pietzka, A.; Huhulescu, S.; Allerberger, F.; Ruppitsch, W.; Schmid, D. Isolate-Based Surveillance of Listeria monocytogenes by Whole Genome Sequencing in Austria. Front. Microbiol. 2019, 10, 2282. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. Babraham Bioinformatics-FastQC a Quality Control tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 January 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-P.; Wang, S.-F.; Hou, P.-B.; Liu, J.; Du, P.; Bai, L.; Fanning, S.; Zhang, H.-N.; Chen, Y.-Z.; Zhang, Y.-K.; et al. Nosocomial cross-infection of hypervirulent Listeria monocytogenes sequence type 87 in China. Ann. Transl. Med. 2020, 8, 603. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Alikhan, N.-F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. GrapeTree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Clausen, P.T.L.C.; Aarestrup, F.M.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19. [Google Scholar] [CrossRef]
- The Integrated Rapid Infectious Disease Analysis (IRIDA) Advanced Research Infrastructure for Experimentation in GenomicS (ARIES) Platform. Available online: https://irida.iss.it (accessed on 1 March 2021).
- Knijn, A.; Michelacci, V.; Orsini, M.; Morabito, S. Advanced research infrastructure for experimentation in genomicS (ARIES): A lustrum of galaxy experience. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lees, J.A.; Harris, S.R.; Tonkin-Hill, G.; Gladstone, R.A.; Lo, S.W.; Weiser, J.N.; Corander, J.; Bentley, S.D.; Croucher, N.J. Fast and flexible bacterial genomic epidemiology with PopPUNK. Genome Res. 2019, 29, 304–316. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russini, V.; Spaziante, M.; Zottola, T.; Fermani, A.G.; Di Giampietro, G.; Blanco, G.; Fabietti, P.; Marrone, R.; Parisella, R.; Parrocchia, S.; et al. A Nosocomial Outbreak of Invasive Listeriosis in An Italian Hospital: Epidemiological and Genomic Features. Pathogens 2021, 10, 591. https://doi.org/10.3390/pathogens10050591
Russini V, Spaziante M, Zottola T, Fermani AG, Di Giampietro G, Blanco G, Fabietti P, Marrone R, Parisella R, Parrocchia S, et al. A Nosocomial Outbreak of Invasive Listeriosis in An Italian Hospital: Epidemiological and Genomic Features. Pathogens. 2021; 10(5):591. https://doi.org/10.3390/pathogens10050591
Chicago/Turabian StyleRussini, Valeria, Martina Spaziante, Tiziana Zottola, Anna Giovanna Fermani, Gina Di Giampietro, Giovanni Blanco, Paolo Fabietti, Riccardo Marrone, Roberta Parisella, Sergio Parrocchia, and et al. 2021. "A Nosocomial Outbreak of Invasive Listeriosis in An Italian Hospital: Epidemiological and Genomic Features" Pathogens 10, no. 5: 591. https://doi.org/10.3390/pathogens10050591