
Renoprotective Effect of Pediococcus acidilactici GKA4 on Cisplatin-Induced Acute Kidney Injury by Mitigating Inflammation and Oxidative Stress and Regulating the MAPK, AMPK/SIRT1/NF-κB, and PI3K/AKT Pathways

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Samples
2.2. Reagents
2.3. Animals
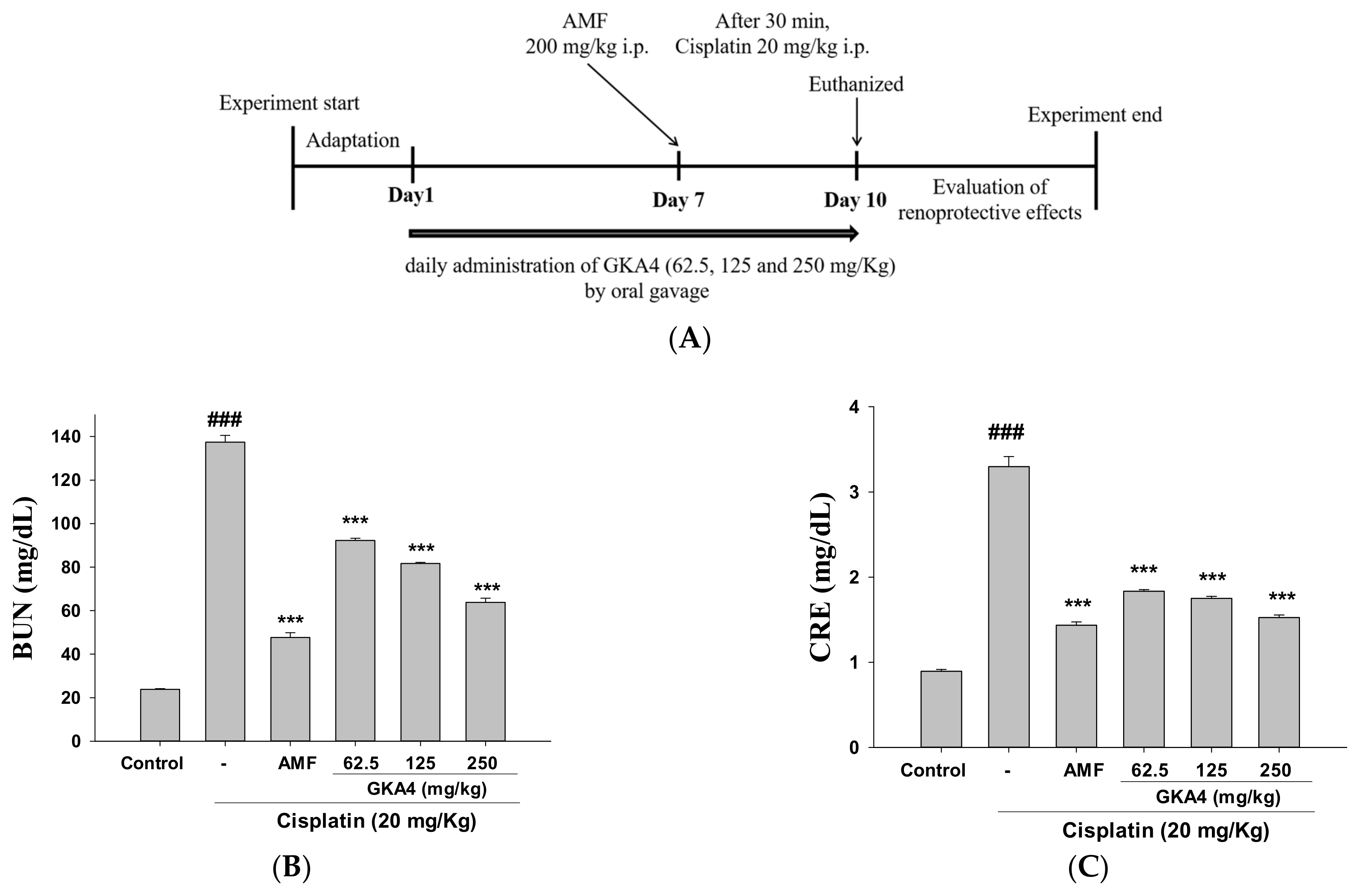
2.4. Research Design
2.5. Kidney Index
2.6. Renal Biomarker Measurements
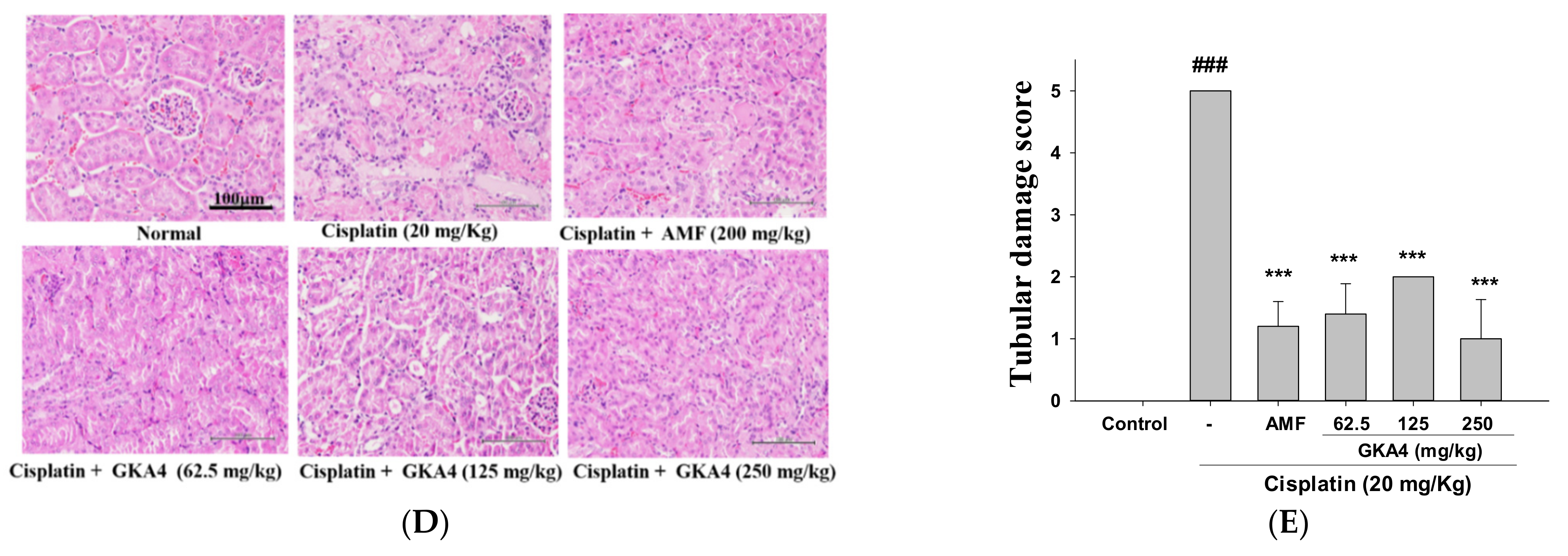
2.7. Histological Examination
2.8. The TBARS (Thiobarbituric Acid Reactive Substance) Assay
2.9. Measurement of Serum Cytokine Levels
2.10. Measurement of Serum NO Levels
2.11. Glutathione (GSH) Asaay
2.12. Western Blot Analysis
2.13. Statistical Analysis
3. Results
3.1. GKA4 Inhibits Kidney Failure and Improves Kidney Activity after Cisplatin-Challenged AKI
3.2. GKA4 Changes in the Renal Index against Cisplatin-Treated Mice
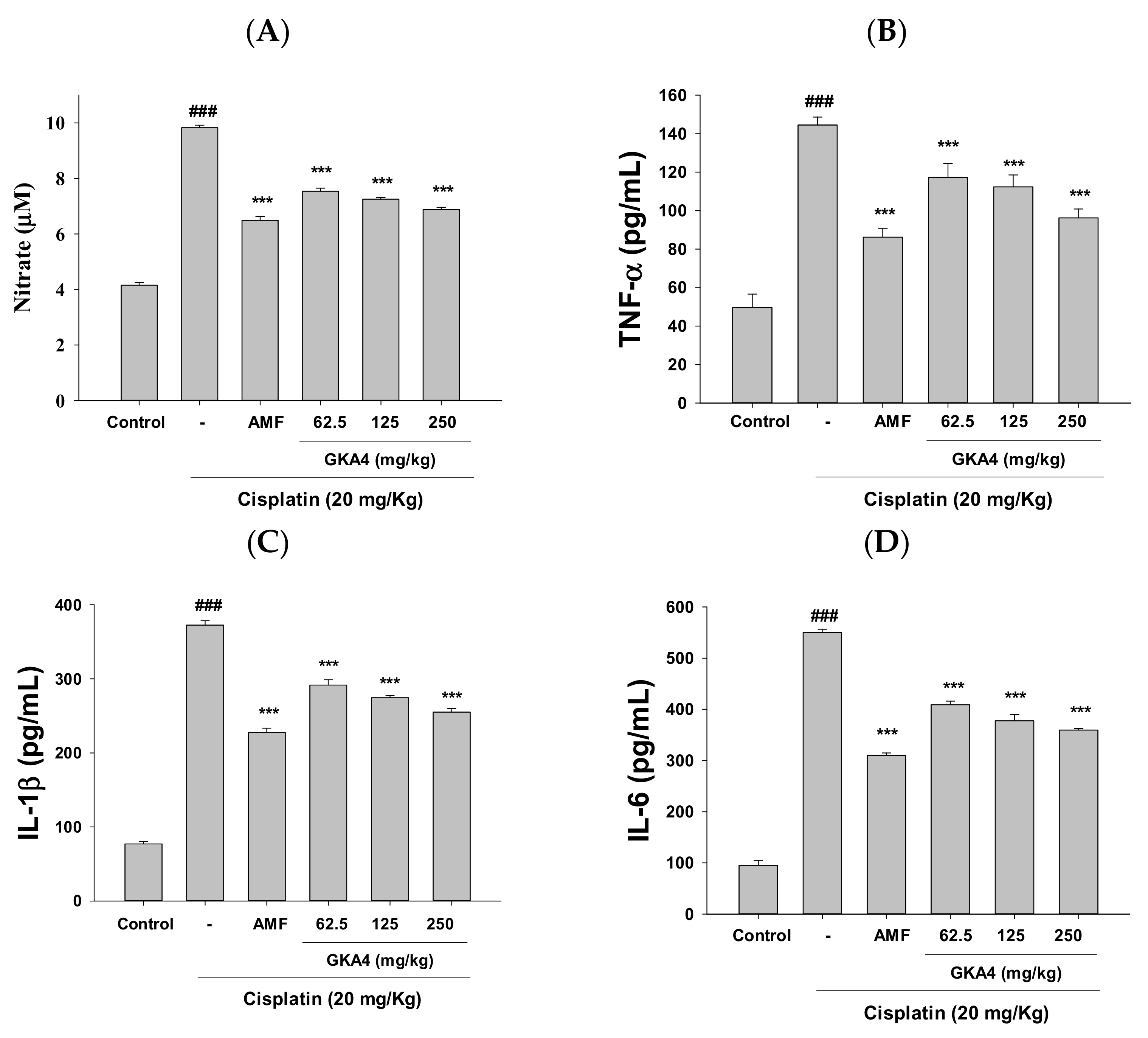
3.3. GKA4 Reduces NO and Pro-Inflammatory Cytokine Levels in Cisplatin-Related Nephrotoxicity
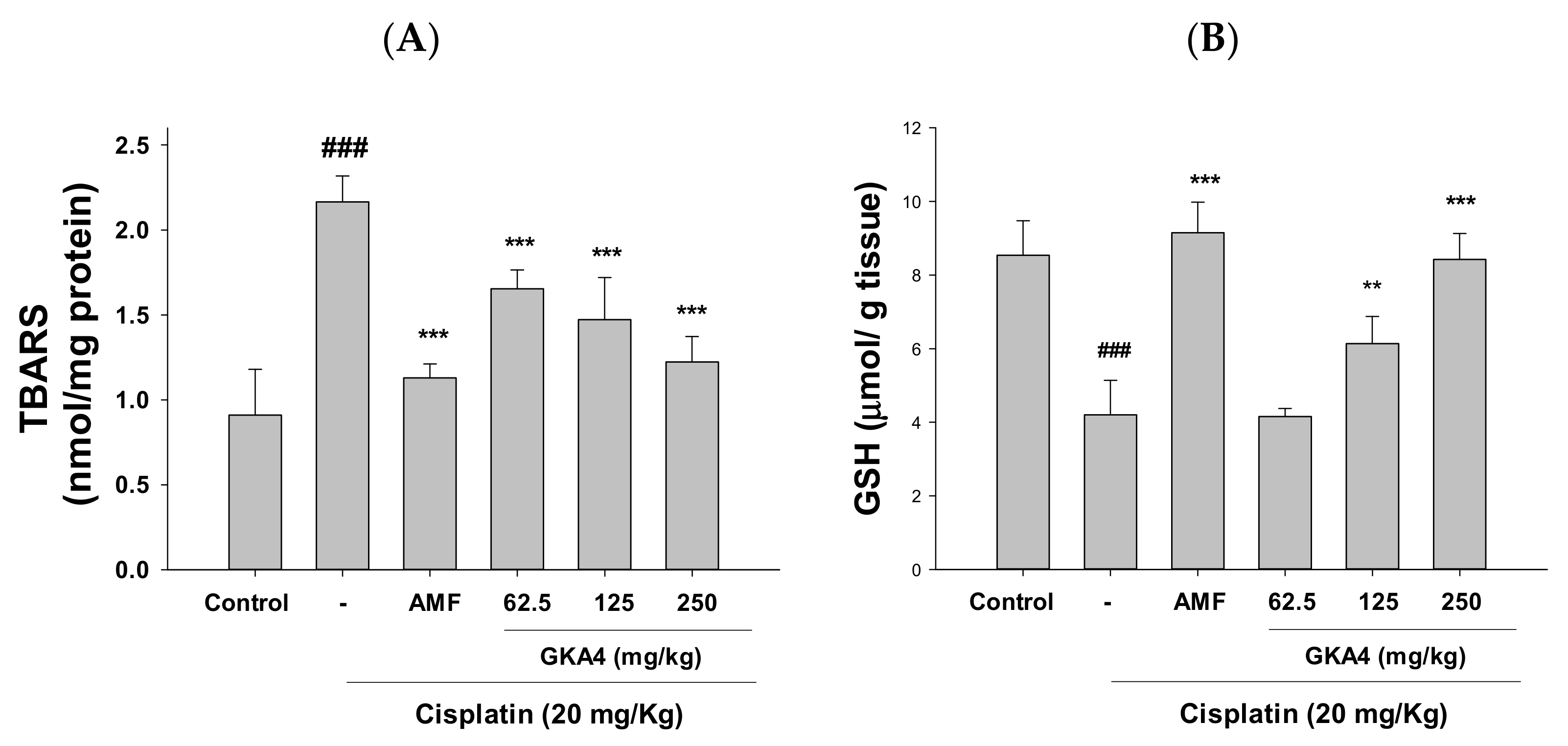
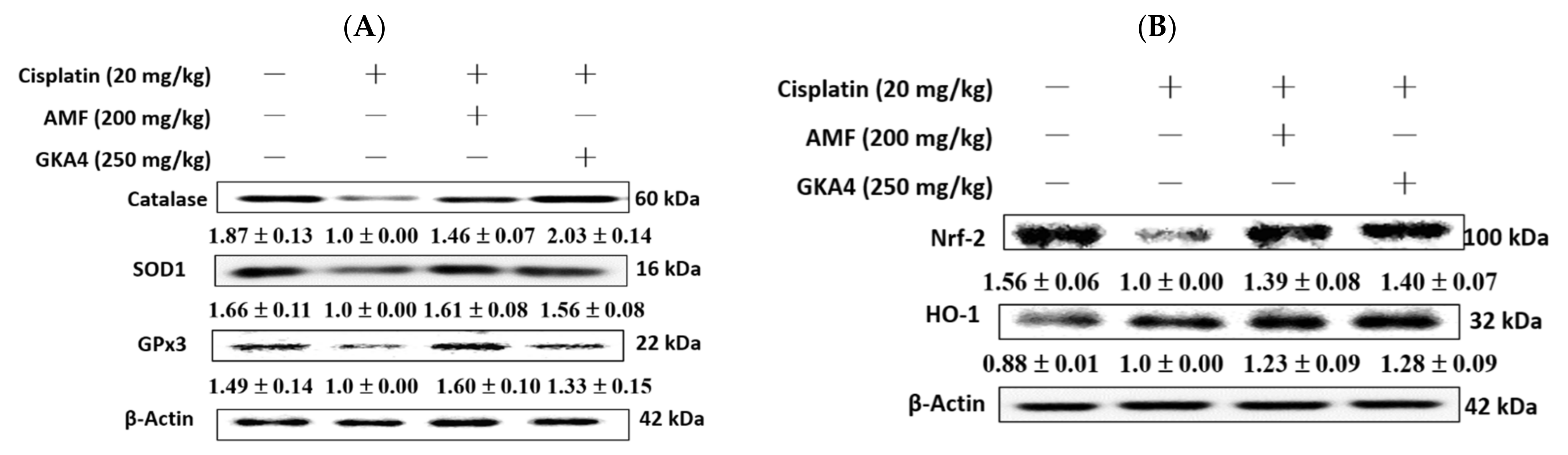
3.4. GKA4 Inhibits Oxidative Stress in Cisplatin-Related AKI
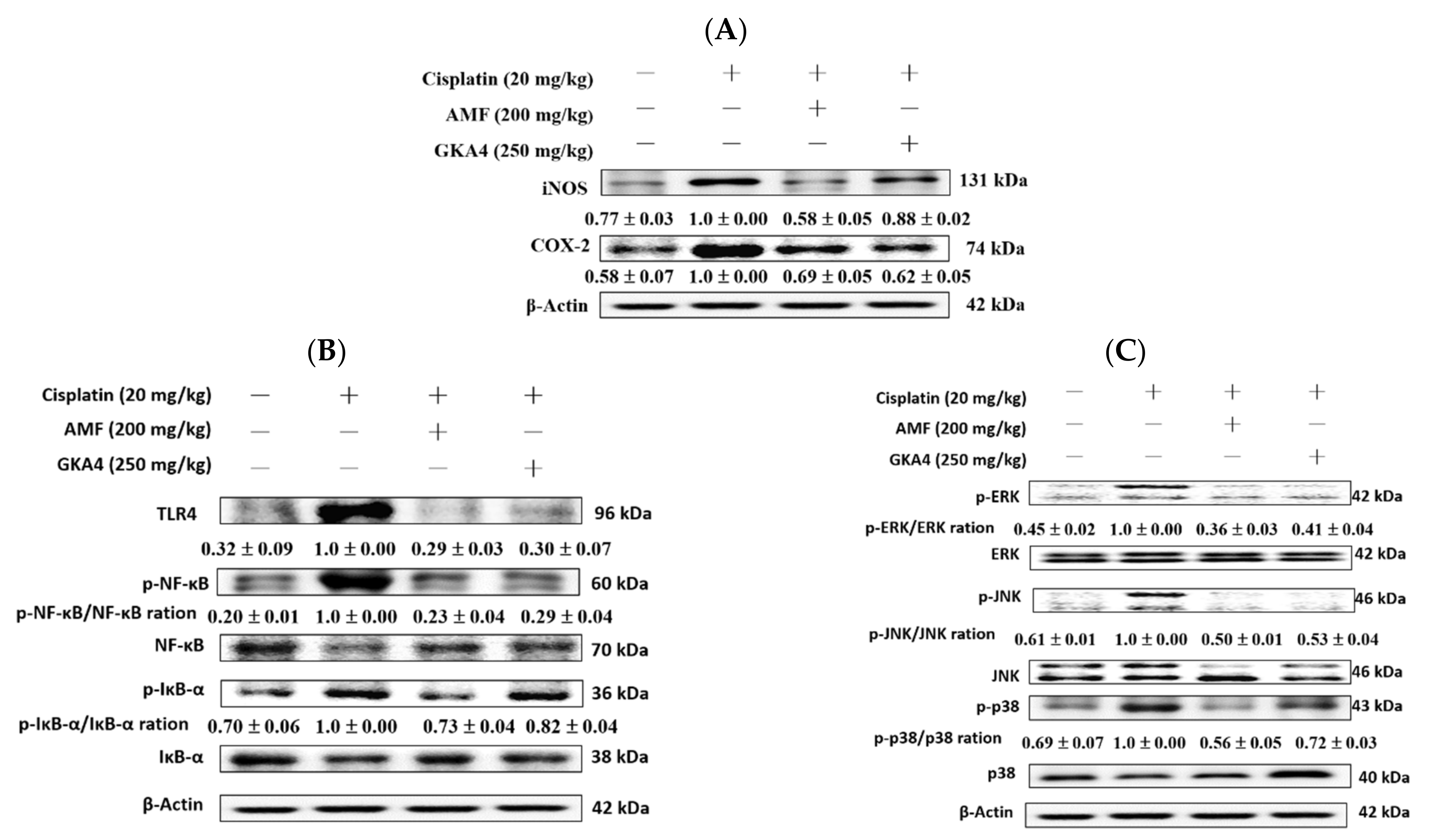
3.5. GKA4 Reduced the Inflammatory Response in Cisplatin-Induced AKI
3.6. GKA4-Inactivated Cisplatin Induces the MAPK Pathway in Kidneys
3.7. GKA4 Restores Renal Antioxidant Defense and the HO-1/Nrf2 Signaling Pathway in Cisplatin-Associated Nephrotoxicity
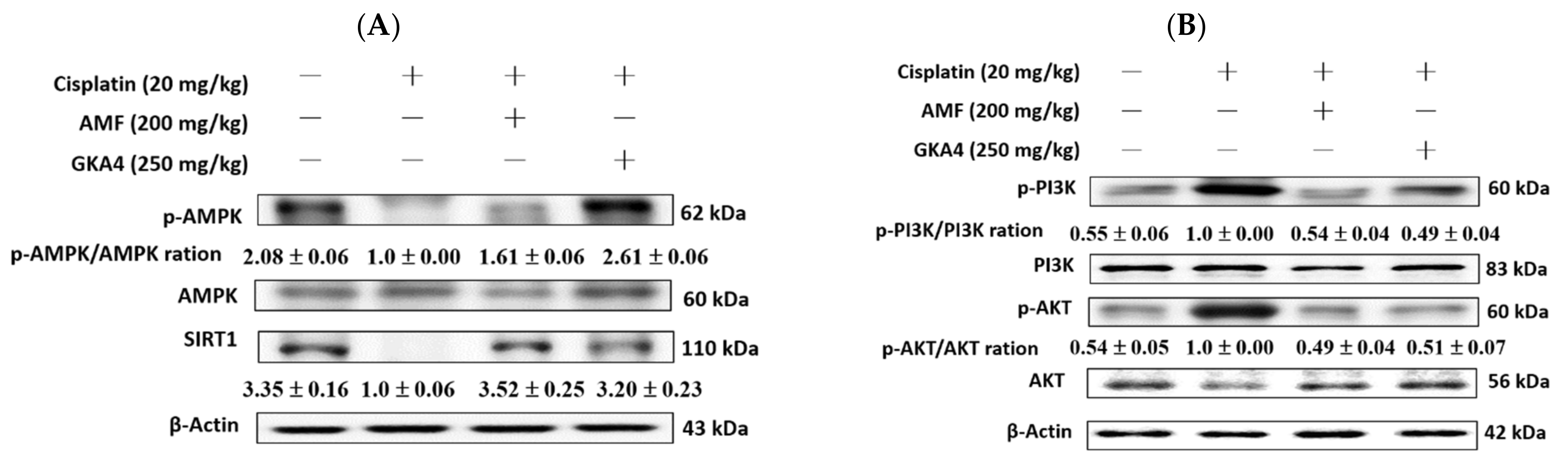
3.8. GKA4 Alleviates the Cisplatin-Induced AMPK, SIRT1, PI3K, and AKT Expressions
3.9. GKA4 Decreases the Cisplatin-Induced Autophagy Signaling Pathway
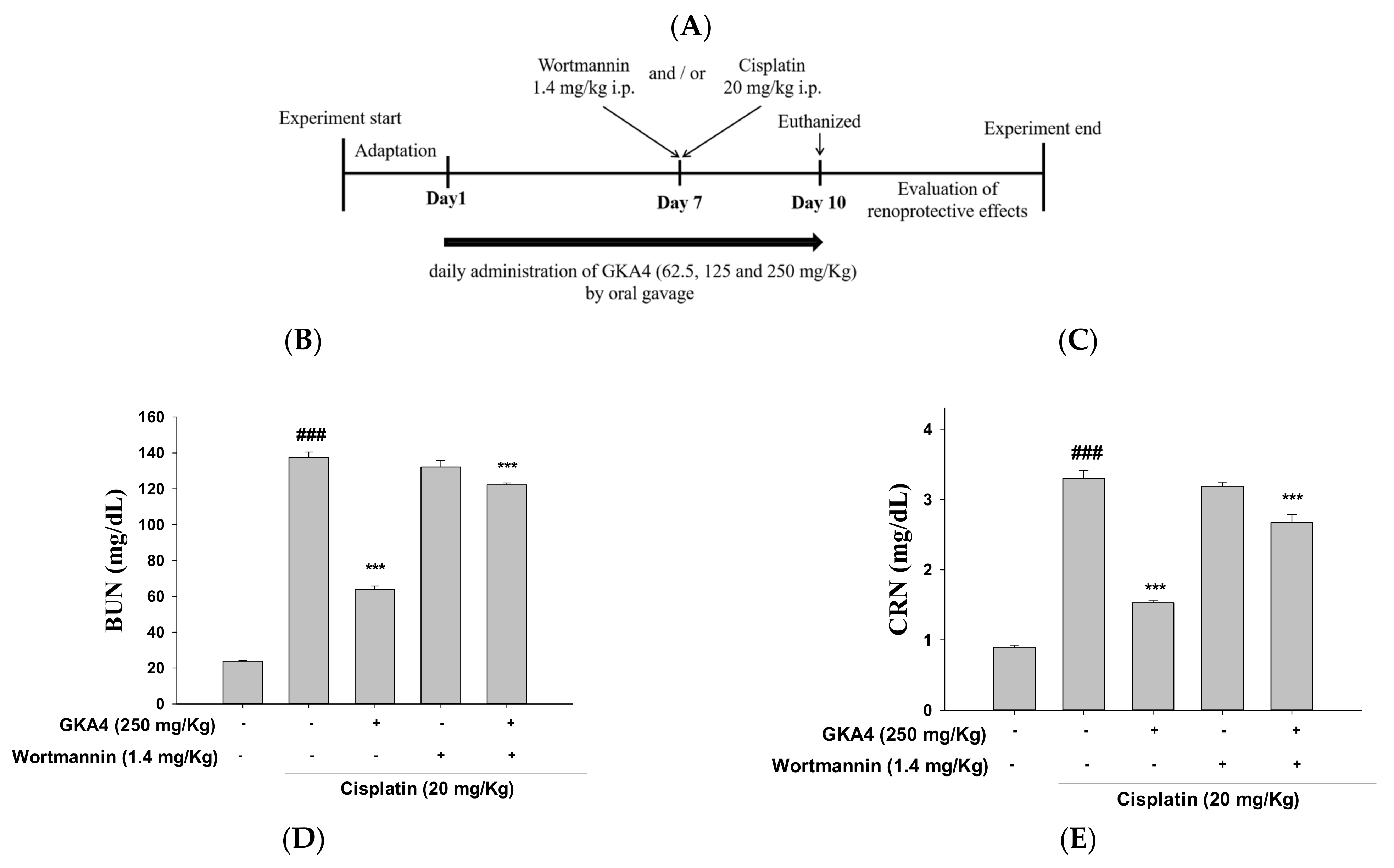
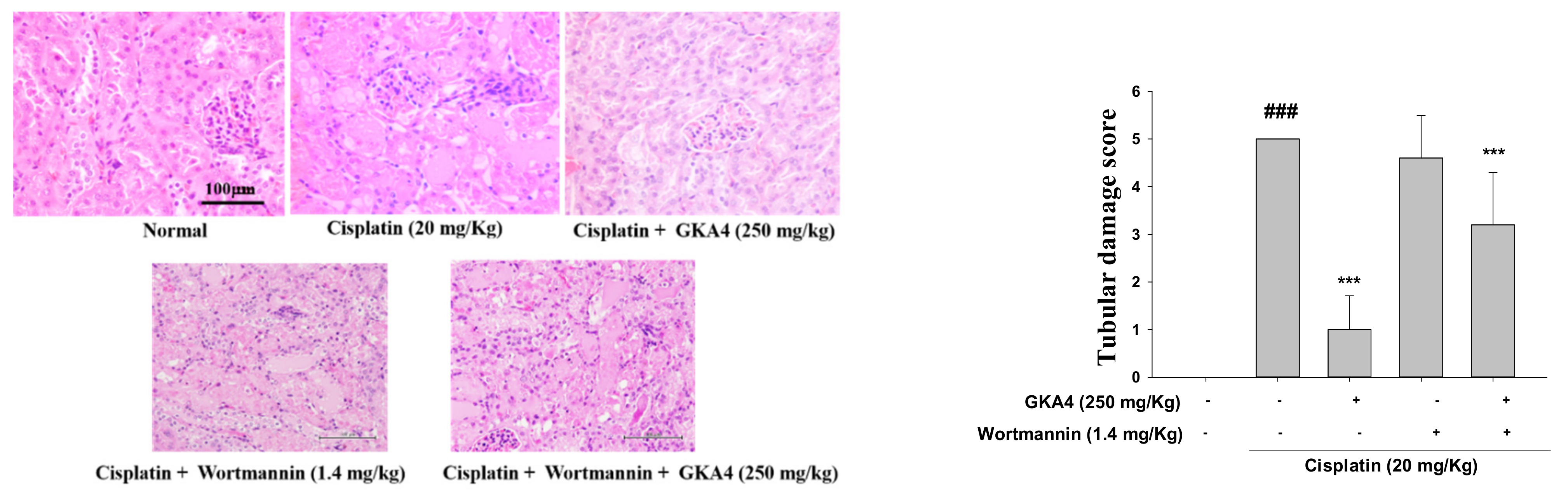
3.10. Blocking the Synergy of PI3K with Wortmannin Increases Renal Damage after Cisplatin Exposure
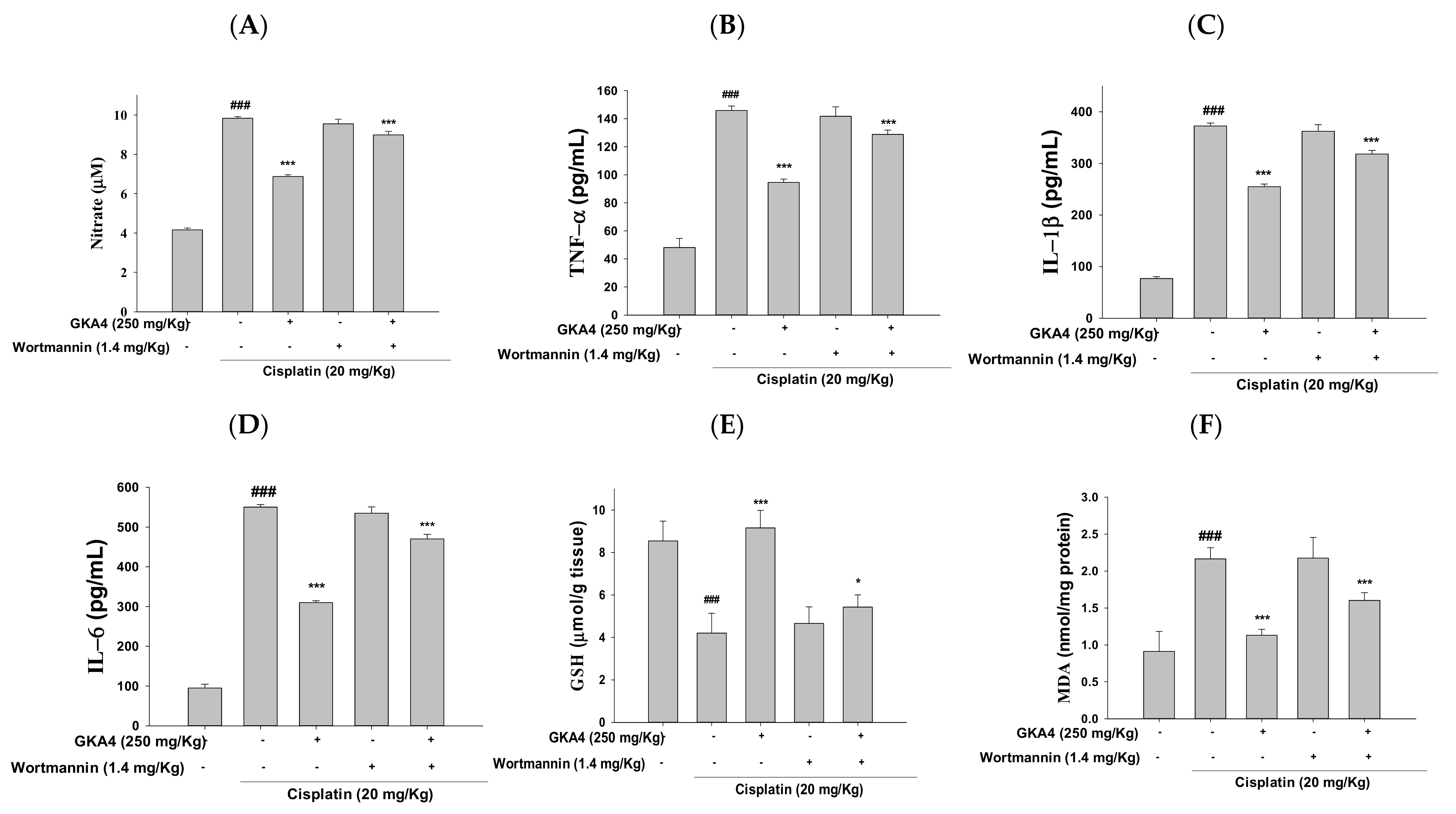
3.11. GKA4 and/or Wortmannin Changes the Anti-Inflammatory Response
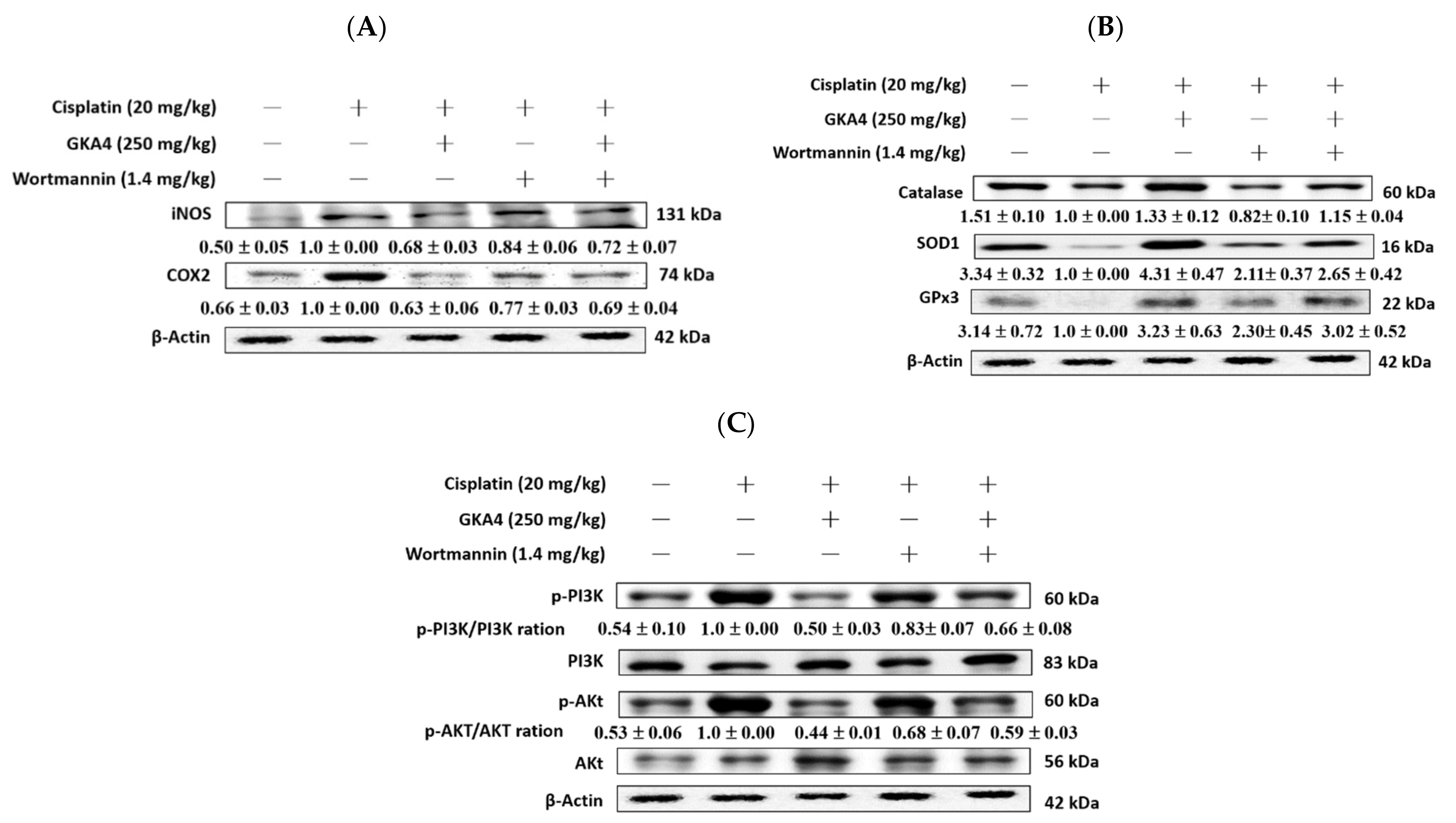
3.12. GKA4 and Wortmannin Diminish the Inflammatory Secretions, Oxidative Stress, and PI3K/AKT Signal-Related Proteins
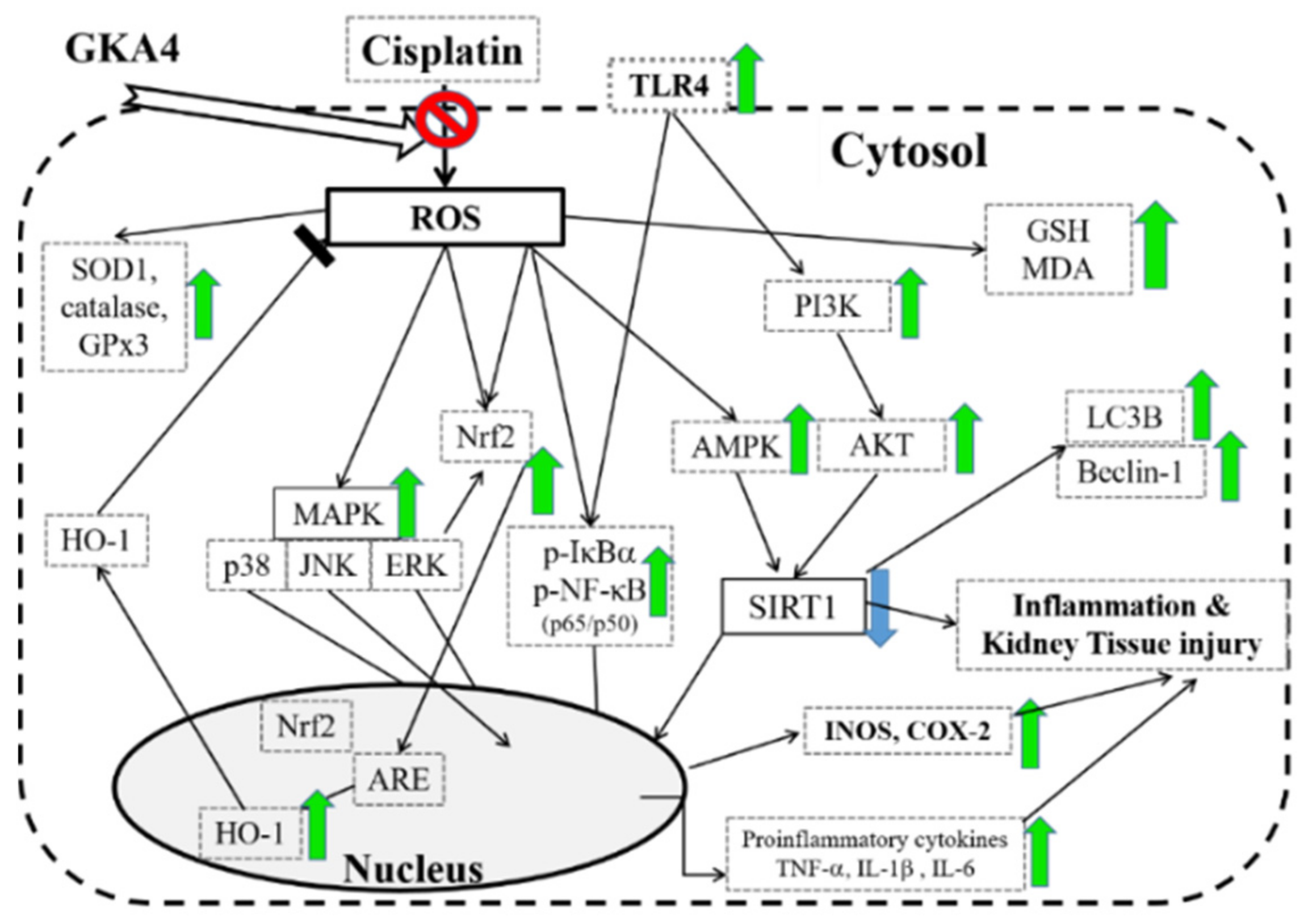
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holditch, S.J.; Brown, C.N.; Lombardi, A.M.; Nguyen, K.N.; Edelstein, C.L. Recent Advances in Models, Mechanisms, Biomarkers, and Interventions in Cisplatin-Induced Acute Kidney Injury. Int. J. Mol. Sci. 2019, 20, 3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perazella, M.A. Drug-induced acute kidney injury: Diverse mechanisms of tubular injury. Curr. Opin. Crit. Care 2019, 25, 550–557. [Google Scholar] [CrossRef]
- McSweeney, K.R.; Gadanec, L.K.; Qaradakhi, T.; Ali, B.A.; Zulli, A.; Apostolopoulos, V. Mechanisms of cisplatin-induced acute kidney injury: Pathological mechanisms, pharmacological interventions, and genetic mitigations. Cancers 2021, 13, 1572. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Jiang, X.; Li, A.; Zhao, Z.; Li, S. S-Allylmercaptocysteine attenuates cisplatin-induced nephrotoxicity through suppression of apoptosis, oxidative stress, and inflammation. Nutrients 2017, 9, 166. [Google Scholar] [CrossRef] [Green Version]
- Volarevic, V.; Djokovic, B.; Jankovic, M.G.; Harrell, C.R.; Fellabaum, C.; Djonov, V.; Arsenijevic, N. Molecular mechanisms of cisplatin-induced nephrotoxicity: A balance on the knife-edge between renoprotection and tumor toxicity. J. Biomed. Sci. 2019, 26, 25. [Google Scholar] [CrossRef] [Green Version]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant mechanisms in renal injury and disease. Antioxid. Redox Signal. 2016, 25, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Atherton, S. Tumor necrosis factor-α in cisplatin nephrotoxicity: A homebred for? Kidney Int. 2007, 72, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Akcay, A.; Nguyen, Q.; Edelstein, C.L. Mediators of inflammation in acute kidney injury. Mediat. Inflamm. 2009, 2009, 137072. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhao, T.; Xiao, H. The implication of oxidative stress and AMPK-Nrf2 antioxidative signaling in pneumonia pathogenesis. Front. Endocrinol. 2020, 11, 400. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [Green Version]
- Min, S.W.; Sohn, P.D.; Cho, S.H.; Swanson, R.A.; Gan, L. Sirtuins in neurodegenerative diseases: An update on potential mechanisms. Front. Aging Neurosci. 2013, 5, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porto, M.C.W.; Kuniyoshi, T.M.; Azevedo, P.; Vitolo, M.; Oliveira, R.S. Pediococcus spp.: An important genus of lactic acid bacteria and pediocin producers. Biotechnol. Adv. 2017, 35, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Qiu, Z.; Tian, F.; Yu, L.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Pediococcus acidilactici strains improve constipation symptoms and regulate intestinal flora in mice. Front. Cell. Infect. Microbiol. 2021, 11, 655258. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Wu, C. Modulation of gut microbiota and immune system by probiotics, prebiotics, and post-biotics. Front. Nutr. 2021, 8, 634897. [Google Scholar] [CrossRef]
- Miranda, C.; Contente, D.; Igrejas, G.; Câmara, S.; Dapkevicius, M.d.L.E.; Poeta, P. Role of exposure to lactic acid bacteria from foods of animal origin in human health. Foods 2021, 10, 2092. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wang, C.S.; Tsai, Y.S.; Lin, S.W.; Wu, W.S.; Chen, Y.L.; Chen, C.C. Screening and evaluation of probiotics for reducing intestinal inflammation. Hans J. Food Nutr. Sci. 2022, 11, 44–55. [Google Scholar] [CrossRef]
- Shackelford, C.; Long, G.; Wolf, J.; Okerberg, C.; Herbert, R. Qualitative and quantitative analysis of nonneoplastic lesions in toxicology studies. Toxicol. Pathol. 2002, 30, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B.; Gutteridge, J.M. Formation of a thiobarbituric-acid-reactive substance from deoxyribose in the presence of iron salts: The role of superoxide and hydroxyl radicals. FEBS Lett. 1981, 128, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Maya, N.I.; Padilla-Vaca, F.; Romero-González, O.E.; Rosales-Castillo, E.A.S.; Rangel-Serrano, Á.; Arias-Negrete, S.; Franco, B. Refinement of the Griess method for measuring nitrite in biological samples. J. Microbiol. Methods 2021, 187, 106260. [Google Scholar] [CrossRef]
- Fakurazi, S.; Hairuszah, I.; Nanthini, U. Moringa oleifera Lam prevents acetaminophen-induced liver injury through restoration of glutathione level. Food Chem. Toxicol. 2008, 46, 2611–2615. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.C. NF-κB in inflammation and renal diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, B.D.; Kumar, J.M.; Sistla, R. Baicalein, a bioflavonoid, prevents cisplatin-induced acute kidney injury by up-regulating antioxidant defenses and down-regulating the MAPKs and NF-κB pathways. PLoS ONE 2015, 10, e0134139. [Google Scholar] [CrossRef] [Green Version]
- Gyurászová, M.; Gurecká, R.; Bábíčková, J.; Tóthová, Ľ. Oxidative stress in the pathophysiology of kidney disease: Implications for noninvasive monitoring and identification of biomarkers. Oxid. Med. Cell. Longev. 2020, 2020, 5478708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Tan, B.; Vasan, K.; Yuan, H.; Cheng, F.; Ramos da Silva, S.; Lu, C.; Gao, S.J. SIRT1 and AMPK pathways are essential for the proliferation and survival of primary effusion lymphoma cells. J. Pathol. 2017, 242, 309–321. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.H.W.; Kuo, M.T. Role of glutathione in the regulation of cisplatin resistance in cancer chemotherapy. Met.-Based Drugs 2010, 2010, 430939. [Google Scholar] [CrossRef]
- Perše, M.; Večerić-Haler, Ž. Cisplatin-induced rodent model of kidney injury: Characteristics and challenges. Biomed. Res. Int. 2018, 2018, 1462802. [Google Scholar] [CrossRef]
- Yao, X.; Panichpisal, K.; Kurtzman, N.; Nugent, K. Cisplatin nephrotoxicity: A review. Am. J. Med. Sci. 2007, 334, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Levi, J.; Jacobs, C.; Kalman, S.M.; McTigue, M.; Weiner, M.W. Mechanism of cis-platinum nephrotoxicity: I. Effects of sulfhydryl groups in rat kidneys. J. Pharmacol. Exp. Ther. 1980, 213, 545–550. [Google Scholar]
- Perše, M. Cisplatin mouse models: Treatment, toxicity and translatability. Biomedicines 2021, 9, 1406. [Google Scholar] [CrossRef]
- Spreckelmeyer, S.; Orvig, C.; Casini, A. Cellular transport mechanisms of cytotoxic metallodrugs: An overview beyond cisplatin. Molecules 2014, 19, 15584–15610. [Google Scholar] [CrossRef] [Green Version]
- Florea, A.M.; Büsselberg, D. Cisplatin as an anti-tumor drug: Cellular mechanisms of activity, drug resistance, and induced side effects. Cancers 2011, 3, 1351–1371. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. 2018, 6, 89–94. [Google Scholar]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M. The role and potential application of antimicrobial peptides in autoimmune diseases. Front. Immunol. 2020, 11, 859. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Sureda, A.; Belll, T.; Çetinkaya, S.; Süntar, İ.; Tejada, S.; Devkota, H.P.; Ullah, H.; Aschner, M. Polyphenols in the treatment of autoimmune diseases. Autoimmun. Rev. 2019, 18, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chung, J.Y.; Kim, K.Y.; Im, W.; Kim, M. Two-weeks repeated-dose oral toxicity study of Pediococcus acidilactici J9 in a mice model. BMC Microbiol. 2020, 20, 372. [Google Scholar] [CrossRef]
- Ma, N.; Wei, W.; Fan, X.; Ci, X. Farrerol attenuates cisplatin-induced nephrotoxicity by inhibiting the reactive oxygen species-mediated oxidation, inflammation, and apoptotic signaling pathways. Front. Physiol. 2019, 10, 1419. [Google Scholar] [CrossRef]
- Feng, M.; Smith, D.E.; Normolle, D.P.; Knol, J.A.; Pan, C.C.; Ben-Josef, E.; Lu, Z.; Feng, M.R.; Chen, J.; Ensminger, W. A phase I clinical and pharmacology study using amifostine as a radioprotector in dose-escalated whole liver radiation therapy. Int. J. Radiat. Oncol. Biol. Phys. 2012, 83, 1441–1447. [Google Scholar] [CrossRef] [Green Version]
- Dubey, V.; Mishra, A.K.; Ghosh, A.R.; Mandal, B.K. Probiotic Pediococcus pentosaceus GS 4 shields brush border membrane and alleviates liver toxicity imposed by chronic cadmium exposure in Swiss albino mice. J. Appl. Microbiol. 2019, 126, 1233–1244. [Google Scholar] [CrossRef]
- Di Cerbo, A.; Palmieri, B.; Aponte, M.; Morales-Medina, J.C.; Iannitti, T. Mechanisms and therapeutic effectiveness of lactobacilli. J. Clin. Pathol. 2016, 69, 187–203. [Google Scholar] [CrossRef]
- Kaur, B.; Garg, N.; Sachdev, A.; Kumar, B. Effect of the oral intake of probiotic Pediococcus acidilactici BA28 on Helicobacter pylori causing peptic ulcer in C57BL/6 mice models. Appl. Biochem. Biotechnol. 2014, 172, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Bikheet, M.M.; Mahmoud, M.E.; Yassien, E.E.; Hassan, H.M. Effect of lactic acid bacteria isolated from some fermented dairy products on carbon tetrachloride-induced hepatotoxicity and nephrotoxicity of albino rats. Environ. Sci. Pollut. Res. Int. 2022, 29, 11790–11800. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.P.; Tadagavadi, R.K.; Ramesh, G.; Reeves, W.B. Mechanisms of cisplatin nephrotoxicity. Toxins 2010, 2, 2490–2518. [Google Scholar] [CrossRef] [Green Version]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Ramesh, G.; Uematsu, S.; Akira, S.; Reeves, W.B. TLR4 signaling mediates inflammation and tissue injury in nephrotoxicity. J. Am. Soc. Nephrol. 2008, 19, 923–932. [Google Scholar] [CrossRef]
- Oh, G.-S.; Kim, H.J.; Choi, J.H.; Shen, A.; Kim, C.H.; Kim, S.J.; Shin, S.R.; Hong, S.H.; Kim, Y.; Park, C. Activation of lipopolysaccharide–TLR4 signaling accelerates the ototoxic potential of cisplatin in mice. J. Immunol. 2011, 186, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yan, L.; Zhu, Q.; Shao, F. Puerarin attenuates cisplatin-induced rat nephrotoxicity: The involvement of TLR4/NF-κB signaling pathway. PLoS ONE 2017, 12, e0171612. [Google Scholar] [CrossRef] [Green Version]
- De Almeida Chuffa, L.G.; de Moura Ferreira, G.; Lupi, L.A.; da Silva Nunes, I.; Fávaro, W.J. P-MAPA immunotherapy potentiates the effect of cisplatin on serous ovarian carcinoma through targeting TLR4 signaling. J. Ovarian Res. 2018, 11, 8. [Google Scholar] [CrossRef]
- Cassidy, H.; Radford, R.; Slyne, J.; O’Connell, S.; Slattery, C.; Ryan, M.P.; McMorrow, T. The role of MAPK in drug-induced kidney injury. J. Signal. Transduct. 2012, 2012, 463617. [Google Scholar] [CrossRef] [Green Version]
- Sung, M.J.; Kim, D.H.; Jung, Y.J.; Kang, K.P.; Lee, A.S.; Lee, S.; Kim, W.; Davaatseren, M.; Hwang, J.T.; Kim, H.J. Genistein protects the kidney from cisplatin-induced injury. Kidney Int. 2008, 74, 1538–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa-Diez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sánchez-Pérez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Ramalho, J.B.; Soares, M.B.; Spiazzi, C.C.; Bicca, D.F.; Soares, V.M.; Pereira, J.G.; Da Silva, W.P.; Sehn, C.P.; Cibin, F.W. In vitro probiotic and antioxidant potential of Lactococcus lactis subsp. cremoris LL95 and its effect on mice behavior. Nutrients 2019, 11, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duann, P.; Lianos, E.A.; Ma, J.; Lin, P.H. Autophagy, innate immunity and tissue repair in acute kidney injury. Int. J. Mol. Sci. 2016, 17, 662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Zhang, Y.Y.; Lan, J.N.; Liu, H.M.; Li, W.; Wu, Y.; Leng, Y.; Tang, L.H.; Hou, J.B.; Sun, Q. Ischemic postconditioning alleviates intestinal ischemia-reperfusion injury by enhancing autophagy and suppressing oxidative stress through the Akt/GSK-3β/Nrf2 pathway in mice. Oxid. Med. Cell. Longev. 2020, 2020, 6954764. [Google Scholar] [CrossRef] [Green Version]
- Juszczak, F.; Caron, N.; Mathew, A.V.; Declèves, A.E. Critical role for AMPK in metabolic disease-induced chronic kidney disease. Int. J. Mol. Sci. 2020, 21, 7994. [Google Scholar] [CrossRef]
- Ogura, Y.; Kitada, M.; Koya, D. Sirtuins and renal oxidative stress. Antioxidants 2021, 10, 1198. [Google Scholar] [CrossRef]
- Elibol, B.; Kilic, U. High levels of SIRT1 expression as a protective mechanism against disease-related conditions. Front. Endocrinol. 2018, 9, 614. [Google Scholar] [CrossRef]
- Liao, Y.; Li, H.; Pi, Y.; Li, Z.; Jin, S. Cardioprotective effect of IGF-1 against myocardial ischemia/reperfusion injury through activation of PI3K/Akt pathway in rats in vivo. Int. J. Mol. Sci. 2019, 47, 3886–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.B.; Meng, Q.H.; Huang, C.; Wang, J.B.; Liu, X.W. Nephroprotective effects of polydatin against ischemia/reperfusion injury: A role for the PI3K/Akt signal pathway. Oxid. Med. Cell. Longev. 2015, 2015, 362158. [Google Scholar] [CrossRef] [PubMed] [Green Version]












 : inhibition;
: inhibition;  : up-regulation;
: up-regulation;  : down-regulation.
: inhibition; : up-regulation; : down-regulation.
: down-regulation.
: inhibition; : up-regulation; : down-regulation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Initial Body Weight (g) | Final Body Weight (g) | Kidney Index (mg/g) |
|---|---|---|---|
| Control | 34.12 ± 0.82 | 39.52 ± 0.47 | 1.36 ± 0.08 |
| Cisplatin (20 mg/kg) | 34.14 ± 0.15 | 32.50 ± 0.16 ### | 2.36 ± 0.09 ### |
| Cisplatin (20 mg/kg) + AMF (200 mg/kg) | 34.08 ± 0.30 | 36.56 ± 0.15 *** | 1.51 ± 0.08 *** |
| Cisplatin (20 mg/kg) + GKA4 (62.5 mg/kg) | 34.06 ± 0.44 | 35.48 ± 0.22 *** | 1.85 ± 0.09 *** |
| Cisplatin (20 mg/kg) + GKA4 (125 mg/kg) | 34.14 ± 0.35 | 35.84 ± 0.15 *** | 1.75 ± 0.04 *** |
| Cisplatin (20 mg/kg) + GKA4 (250 mg/kg) | 34.06 ± 0.21 | 36.36 ± 0.21 *** | 1.61 ± 0.06 *** |
| Groups | Initial Body (g) | Final Body (g) | Kidney Index (mg/g) |
|---|---|---|---|
| Control | 34.00 ± 0.69 | 39.56 ± 0.55 | 1.36 ± 0.08 |
| Cisplatin (20 mg/kg) | 33.90 ± 0.16 | 36.50 ±0.19 ### | 2.36 ± 0.09 ### |
| Cisplatin (20 mg/kg) + GKA4 (250 mg/kg) | 34.10 ± 0.13 | 34.30 ± 0.53 *** | 1.61 ± 0.04 *** |
| Cisplatin (20 mg/kg) + wortmannin (1.4 mg/kg) | 34.40 ± 0.26 | 33.40 ± 0.51 * | 2.26 ± 0.08 |
| Cisplatin (20 mg/kg) + GKA4 (250 mg/kg) + wortmannin (1.4 mg/kg) | 33.88 ± 0.26 | 34.32 ± 0.19 *** | 2.05 ± 0.06 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, W.-H.; Jiang, W.-P.; Chen, C.-C.; Lee, L.-Y.; Tsai, Y.-S.; Chien, L.-H.; Chou, Y.-N.; Deng, J.-S.; Huang, G.-J. Renoprotective Effect of Pediococcus acidilactici GKA4 on Cisplatin-Induced Acute Kidney Injury by Mitigating Inflammation and Oxidative Stress and Regulating the MAPK, AMPK/SIRT1/NF-κB, and PI3K/AKT Pathways. Nutrients 2022, 14, 2877. https://doi.org/10.3390/nu14142877
Lin W-H, Jiang W-P, Chen C-C, Lee L-Y, Tsai Y-S, Chien L-H, Chou Y-N, Deng J-S, Huang G-J. Renoprotective Effect of Pediococcus acidilactici GKA4 on Cisplatin-Induced Acute Kidney Injury by Mitigating Inflammation and Oxidative Stress and Regulating the MAPK, AMPK/SIRT1/NF-κB, and PI3K/AKT Pathways. Nutrients. 2022; 14(14):2877. https://doi.org/10.3390/nu14142877
Chicago/Turabian StyleLin, Wen-Hsin, Wen-Ping Jiang, Chin-Chu Chen, Li-Ya Lee, You-Shan Tsai, Liang-Hsuan Chien, Ya-Ni Chou, Jeng-Shyan Deng, and Guan-Jhong Huang. 2022. "Renoprotective Effect of Pediococcus acidilactici GKA4 on Cisplatin-Induced Acute Kidney Injury by Mitigating Inflammation and Oxidative Stress and Regulating the MAPK, AMPK/SIRT1/NF-κB, and PI3K/AKT Pathways" Nutrients 14, no. 14: 2877. https://doi.org/10.3390/nu14142877