Red Pepper (Capsicum annuum L.) Seed Extract Improves Glycemic Control by Inhibiting Hepatic Gluconeogenesis via Phosphorylation of FOXO1 and AMPK in Obese Diabetic db/db Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Fasting Blood Glucose and Oral Glucose Tolerance Test (OGTT)
2.3. Body Composition
2.4. Blood and Tissue Sampling
2.5. Blood Biomarkers Assay
2.6. Quantitative RT-PCR
2.7. Immunoblotting
2.8. Statistical Analysis
3. Results
3.1. RPSE Did Not Alter Growth Performance or Body Composition in ObD Mice
3.2. RPSE Improved Risk Factors for Type 2 Diabetes Mellitus and Lipid Profiles in ObD Mice
3.3. RPSE Reduced Pro-Inflammatory Cytokines without Hepatotoxicity in the ObD Mice
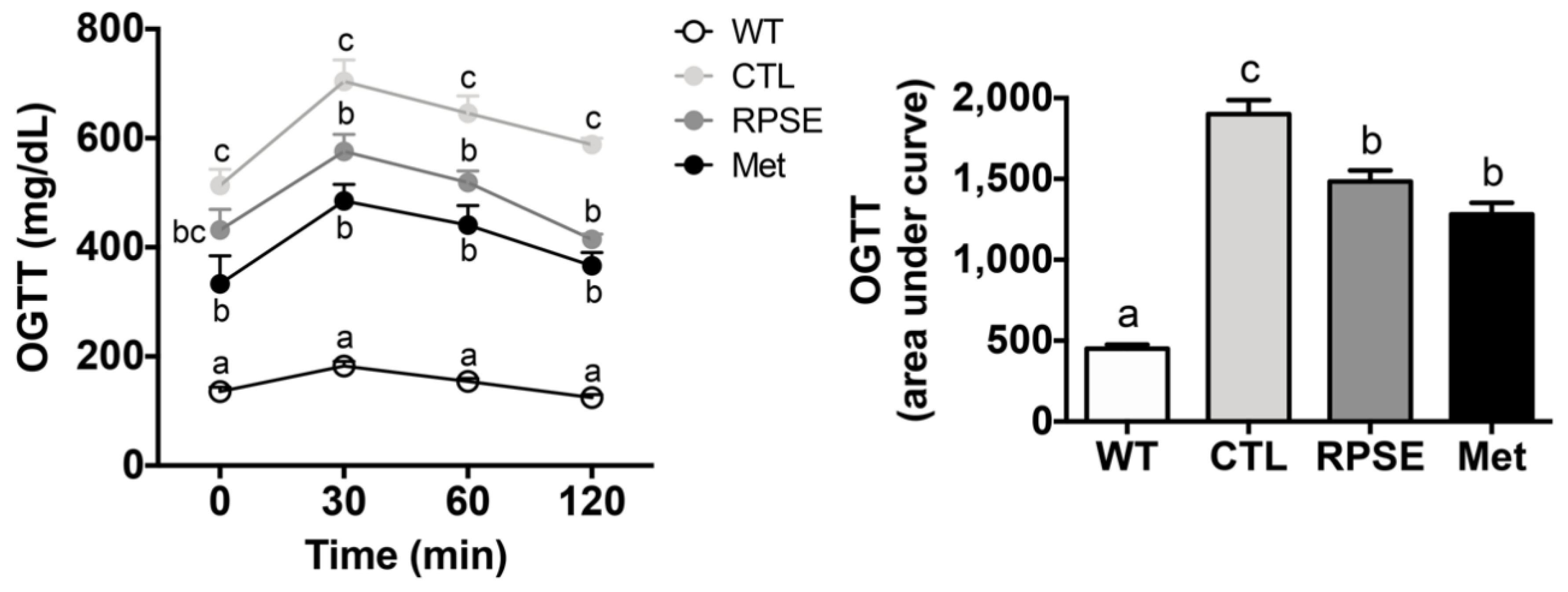
3.4. RPSE Improved Glucose Uptake by Amending Insulin Sensitivity in ObD Mice
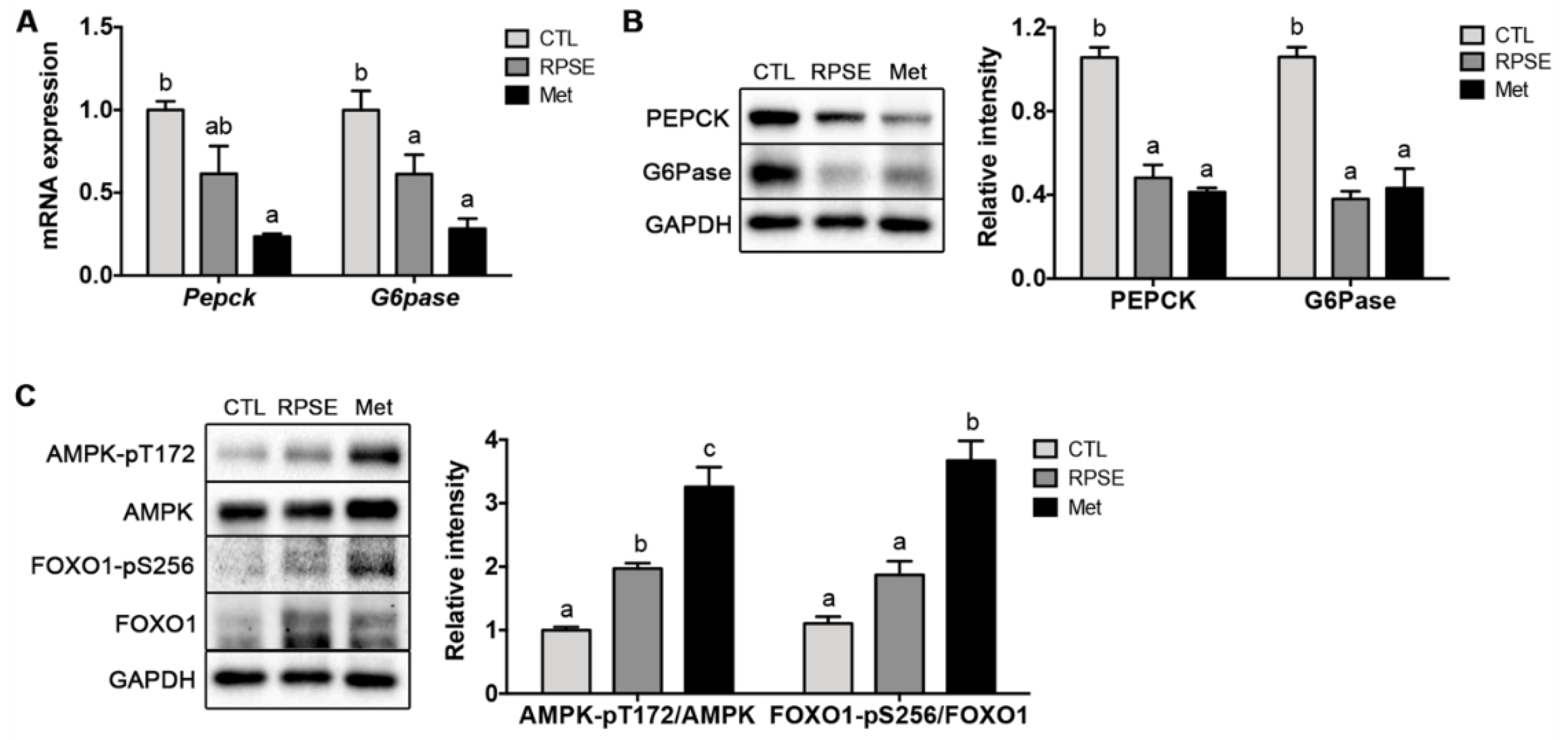
3.5. RPSE Downregulated Hepatic Gluconeogenic Genes and Proteins via Phosphorylating FOXO1 and AMPK
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42, S13–S28. [Google Scholar] [CrossRef] [Green Version]
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Battisti, W.P.; Palmisano, J.; Keane, W.F. Dyslipidemia in patients with type 2 diabetes. Relationships between lipids, kidney disease and cardiovascular disease. Clin. Chem. Lab. Med. 2003, 41, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Available online: https://www.who.int/dietphysicalactivity/media/en/gsfs_obesity.pdf (accessed on 10 July 2020).
- U.S. National Plant Germplasm System. Available online: https://npgsweb.ars-grin.gov/gringlobal/taxonomydetail.aspx?id=8904 (accessed on 10 July 2020).
- Gnayfeed, M.H.; Daood, H.G.; Biacs, P.A.; Alcaraz, C.F. Content of bioactive compounds in pungent spice red pepper (paprika) as affected by ripening and genotype. J. Sci. Food Agric. 2001, 81, 1580–1585. [Google Scholar] [CrossRef]
- Imran, M.; Butt, M.S.; Suleria, H.A.R. Capsicum annuum bioactive compounds: Health promotion perspectives. In Bioactive Molecules in Food; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2018; pp. 1–22. ISBN 978-3-319-54528-8. [Google Scholar]
- Blanco-Ríos, A.K.; Medina-Juárez, L.Á.; González-Aguilar, G.A.; Gámez-Meza, N. Antioxidant activity of the phenolic and oily fractions of different sweet bell peppers. J. Mex. Chem. Soc. 2013, 57, 137–143. [Google Scholar] [CrossRef]
- Oboh, G.; Rocha, J.B. Distribution and antioxidant activity of polyphenols in ripe and unripe tree pepper (Capsicum pubescens). J. Food Biochem. 2007, 31, 456–473. [Google Scholar] [CrossRef]
- Chu, Y.F.; Sun, J.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar] [CrossRef]
- Özyildiz, F.; Karagönlü, S.; Basal, G.; Uzel, A.; Bayraktar, O. Micro-encapsulation of ozonated red pepper seed oil with antimicrobial activity and application to nonwoven fabric. Lett. Appl. Microbiol. 2013, 56, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Ahn, I.S.; Do, M.S.; Kim, S.O.; Jung, H.S.; Kim, Y.I.; Kim, H.J.; Park, K.Y. Antiobesity effect of kochujang (Korean fermented red pepper paste) extract in 3T3-L1 adipocytes. J. Med. Food 2006, 9, 15–21. [Google Scholar] [CrossRef]
- Jeon, G.; Choi, Y.; Lee, S.M.; Kim, Y.; Jeong, H.S.; Lee, J. Anti-obesity activity of methanol extract from hot pepper (Capsicum annuum L.) seeds in 3T3-L1 adipocyte. Food Sci. Biotechnol. 2010, 19, 1123–1127. [Google Scholar] [CrossRef]
- Sim, K.H.; Han, Y.S. The antimutagenic and antioxidant effects of red pepper seed and red pepper pericarp (Capsicum annuum L.). Prev. Nutr. Food Sci. 2007, 12, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Song, W.Y.; Ku, K.H.; Choi, J.H. Effect of ethanol extracts from red pepper seeds on antioxidative defense system and oxidative stress in rats fed high-fat, high-cholesterol diet. Nutr. Res. Pract. 2010, 4, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Ku, K.H.; Choi, E.J.; Park, J.B. Chemical component analysis of red pepper (Capsicum annuum L.) seeds with various cultivars. J. Korean Soc. Food Sci. Nutr. 2008, 37, 1084–1089. [Google Scholar] [CrossRef]
- Youn, S.H.; Yin, J.; Ahn, H.S.; Tam, L.T.; Kwon, S.H.; Min, B.; Yun, S.H.; Kim, H.T.; Lee, M.W. Quantitative analysis of Icariside E5 and vanilloyl Icariside E5 from the seed of Capsicum annuum. L. Korean J. Pharmacogn. 2017, 48, 160–165. [Google Scholar]
- Kim, H.; Cho, K.W.; Jeong, J.; Park, K.; Ryu, Y.; Moyo, K.M.; Kim, H.K.; Go, G.W. Red pepper (Capsicum annuum L.) seed extract decreased hepatic gluconeogenesis and increased muscle glucose uptake in vitro. J. Med. Food 2018, 21, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; You, M.K.; Wang, Z.; Lee, Y.H.; Kim, H.A. Red pepper seed water extract suppresses high-fat diet-induced obesity in C57BL/6 mice. Food Sci. Biotechnol. 2020, 29, 275–281. [Google Scholar] [CrossRef]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schürmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Chandrasekera, P.C.; Pippin, J.J. Leptin-and leptin receptor-deficient rodent models: Relevance for human type 2 diabetes. Curr. Diabetes Rev. 2014, 10, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; Yamashita, H.; Qiao, L.; Friedman, J.E. Decreased Akt kinase activity and insulin resistance in C57BL/KsJ-Leprdb/db mice. J. Endocrinol. 2000, 167, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Yerevanian, A.; Soukas, A.A. Metformin: Mechanisms in human obesity and weight loss. Curr. Obes. Rep. 2019, 8, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Malin, S.K.; Kashyap, S.R. Effects of metformin on weight loss: Potential mechanisms. Curr. Opin. Endocrinol. Diabetes Obes. 2014, 21, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Golay, A. Metformin and body weight. Int. J. Obes. 2008, 32, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starling, S. Metformin-induced hepatokine reduces appetite. Nat. Rev. Endocrinol. 2020, 16, 131. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Yang, J.A.; Ku, K.H.; Choi, J.H. Effect of red pepper seeds powder on antioxidative system and oxidative damage in rats fed high-fat, high-cholesterol diet. J. Korean Soc. Food Sci. Nutr. 2009, 38, 1161–1166. [Google Scholar] [CrossRef]
- Sung, J.; Jeong, H.S.; Lee, J. Effect of the capsicoside G-rich fraction from pepper (Capsicum annuum L.) seeds on high-fat diet-induced obesity in mice. Phytother. Res. 2016, 30, 1848–1855. [Google Scholar] [CrossRef]
- Burke, S.J.; Batdorf, H.M.; Burk, D.H.; Noland, R.C.; Eder, A.E.; Boulos, M.S.; Karistad, M.D.; Collier, J.J. db/db mice exhibit features of human type 2 diabetes that are not present in weight-matched C57BL/6J mice fed a western diet. J. Diabetes Res. 2017, 2017, 8503754. [Google Scholar] [CrossRef] [Green Version]
- Sherwani, S.I.; Khan, H.A.; Ekhzaimy, A.; Masood, A.; Sakharkar, M.K. Significance of HbA1c test in diagnosis and prognosis of diabetic patients. Biomark. Insights 2016, 11, BMI–S38440. [Google Scholar] [CrossRef]
- Sun, F.; Xiong, S.; Zhu, Z. Dietary capsaicin protects cardiometabolic organs from dysfunction. Nutrients 2016, 8, 174. [Google Scholar] [CrossRef]
- Magied, M.M.A.; Salama, N.A.R.; Ali, M.R. Hypoglycemic and hypocholesterolemia effects of intragastric administration of dried red chili pepper (Capsicum annum) in alloxan-induced diabetic male albino rats fed with high-fat-diet. J. Food Nutr. Res. 2014, 2, 850–856. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Kim, Y.S.; Ryu, S.Y.; Cha, M.R.; Yon, G.H.; Yang, H.J.; Kim, S.K.; Kang, S.; Park, S. Capsiate improves glucose metabolism by improving insulin sensitivity better than capsaicin in diabetic rats. J. Nutr. Biochem. 2013, 24, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.J.; Qin, Y.; Wang, L.; Zeng, Y.; Chang, H.; Wang, J.; Wang, B.; Wan, J.; Chen, S.H.; Zhang, Q.Y.; et al. Capsaicin-containing chili improved postprandial hyperglycemia, hyperinsulinemia, and fasting lipid disorders in women with gestational diabetes mellitus and lowered the incidence of large-for-gestational-age newborns. Clin. Nutr. 2016, 35, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Kim, Y.N.; Chun, S.S.; Ku, K.H.; Choi, J.H. Effects of ethanol extracts from red pepper (Capsicum annuum L.) seeds on cholesterol adsorption capacity and UDP-glucuronyl transferase activity. J. Life Sci. 2011, 21, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Morigny, P.; Houssier, M.; Mouisel, E.; Langin, D. Adipocyte lipolysis and insulin resistance. Biochimie 2016, 125, 259–266. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, B.; Meng, X.; Yao, S.; Jin, L.; Yang, J.; Wang, J.; Zhang, H.; Zhang, Z.; Cai, D.; et al. Berberine prevents progression from hepatic steatosis to steatohepatitis and fibrosis by reducing endoplasmic reticulum stress. Sci. Rep. 2016, 6, 20848. [Google Scholar] [CrossRef] [Green Version]
- Su, M.L.; He, Y.; Li, Q.S.; Zhu, B.H. Efficacy of acetylshikonin in preventing obesity and hepatic steatosis in db/db mice. Molecules 2016, 21, 976. [Google Scholar] [CrossRef]
- Allemand, A.; Leonardi, B.F.; Zimmer, A.R.; Moreno, S.; Romao, P.R.T.; Gosmann, G. Red pepper (Capsicum baccatum) extracts present anti-inflammatory effects in vivo and inhibit the production of TNF-α and NO in vitro. J. Med. Food 2016, 19, 759–767. [Google Scholar] [CrossRef]
- Spiller, F.; Alves, M.K.; Vieira, S.M.; Carvalho, T.A.; Leite, C.E.; Lunardelli, A.; Poloni, J.A.; Cunha, F.Q.; de Oliveira, J.R. Anti-inflammatory effects of red pepper (Capsicum baccatum) on carrageenan-and antigen-induced inflammation. J. Pharm. Pharmacol. 2008, 60, 473–478. [Google Scholar] [CrossRef]
- Yang, H.J.; Kwon, D.Y.; Kim, M.J.; Kim, D.S.; Kang, S.; Shin, B.K.; Lee, J.J.; Moon, N.R.; Daily, J.W.; Park, S. Red peppers with different pungencies and bioactive compounds differentially modulate energy and glucose metabolism in ovariectomized rats fed high fat diets. J. Funct. Foods 2014, 7, 246–256. [Google Scholar] [CrossRef]
- Manning, B.D.; Toker, A. AKT/PKB signaling: Navigating the network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Dashty, M. A quick look at biochemistry: Carbohydrate metabolism. Clin. Biochem. 2013, 46, 1339–1352. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Vasquez, D.S.; Ravnskjaer, K.; Denechaud, P.D.; Ruth, T.Y.; Alvarez, J.G.; Downes, M.; Evans, R.M.; Montminy, M.; Shaw, R.J. Class IIa histone deacetylases are hormone-activated regulators of FOXO and mammalian glucose homeostasis. Cell 2011, 145, 607–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclerc, I.; Lenzner, C.; Gourdon, L.; Vaulont, S.; Kahn, A.; Viollet, B. Hepatocyte nuclear factor-4α involved in type 1 maturity-onset diabetes of the young is a novel target of AMP-activated protein kinase. Diabetes 2001, 50, 1515–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, S.H.; Flechner, L.; Qi, L.; Zhang, X.; Screaton, R.A.; Jeffries, S.; Hedrick, S.; Xu, W.; Boussouar, F.; Brindle, P.; et al. The CREB coactivator TORC2 is a key regulator of fasting glucose metabolism. Nature 2005, 437, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Bort, A.; Sánchez, B.G.; Spínola, E.; Mateos-Gómez, P.A.; Rodríguez-Henche, N.; Díaz-Laviada, I. The red pepper’s spicy ingredient capsaicin activates AMPK in HepG2 cells through CaMKKβ. PLoS ONE 2019, 14, e0211420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bort, A.; Sánchez, B.G.; Mateos-Gómez, P.A.; Díaz-Laviada, I.; Rodríguez-Henche, N. Capsaicin targets lipogenesis in HepG2 cells through AMPK activation, AKT inhibition and PPARs regulation. Int. J. Mol. Sci. 2019, 20, 1660. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.J.; Jang, D.J.; Hwang, J.T. Anti-diabetic effects of Korean red pepper via AMPK and PPAR-γ activation in C2C12 myotubes. J. Funct. Foods 2012, 4, 552–558. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| db/+ 1 | Control | RPSE | Metformin | |
|---|---|---|---|---|
| Growth performance | ||||
| Initial body weight (g) | 21.19 ± 0.32 a | 29.20 ± 0.23 b | 28.82 ± 0.27 b | 31.58 ± 2.72 b |
| Final body weight (g) | 24.74 ± 0.66 a | 40.85 ± 0.48 c | 41.30 ± 1.18 c | 33.95 ± 2.87 b |
| Weight gain (g/week) | 3.54 ± 0.54 a | 11.65 ± 0.48 b | 12.48 ± 1.30 b | 2.36 ± 5.51 a |
| Total energy intake (kcal) | 504.50 ± 2.52 a | 991.50 ± 2.11 b | 1040.00 ± 53.80 b | 890.70 ± 9.92 b |
| Body composition | ||||
| Fat in tissue (%) | 20.77 ± 1.17 a | 44.28 ± 0.86 b | 45.29 ± 1.74 b | 40.38 ± 6.89 b |
| Lean mass (%) | 77.26 ± 1.12 a | 67.09 ± 0.82 b | 64.89 ± 1.58 b | 66.00 ± 0.95 b |
| Bone mineral density (mg/cm2) | 86.85 ± 0.57 | 82.02 ± 1.76 | 89.04 ± 3.16 | 84.87 ± 7.76 |
| db/+ 1 | Control | RPSE | Metformin | |
|---|---|---|---|---|
| Fasting blood glucose (mg/dL) | 139.80 ± 6.74 a | 544.20 ± 20.88 d | 429.40 ± 22.85 c | 296.30 ± 46.23 b |
| HbA1c 2 (%) | 3.86 ± 0.30 a | 8.58 ± 0.41 c | 6.72 ± 0.45 b | 4.33 ± 0.67 a |
| Insulin (ng/mL) | 1.09 ± 0.04 a | 2.10 ± 0.08 c | 1.76 ± 0.08 b | 1.49 ± 0.09 b |
| Triglyceride (mg/dL) | 72.02 ± 4.08 a | 135.40 ± 9.94 c | 103.90 ± 3.01 b | 48.32 ± 6.81 a |
| Non-esterified fatty acid (mEq/L) | 0.54 ± 0.07 a | 1.21 ± 0.04 b | 0.99 ± 0.06 b | 0.56 ± 0.07 a |
| db/+ 1 | Control | RPSE | Metformin | |
|---|---|---|---|---|
| ALT 2 (IU/L) | 37.35 ± 6.14 | 45.40 ± 5.00 | 40.81 ± 2.69 | 46.31 ± 2.64 |
| AST (IU/L) | 33.17 ± 3.20 a | 46.29 ± 4.80 ab | 49.45 ± 5.39 b | 38.47 ± 3.41 ab |
| TNF-α (pg/mL) | 94.39 ± 8.13 a | 188.20 ± 11.79 c | 146.70 ± 7.44 b | 119.60 ± 11.52 ab |
| IL-6 (pg/mL) | 62.33 ± 4.52 a | 164.10 ± 15.24 b | 129.10 ± 11.28 b | 73.84 ± 9.21 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.K.; Jeong, J.; Kang, E.Y.; Go, G.-w. Red Pepper (Capsicum annuum L.) Seed Extract Improves Glycemic Control by Inhibiting Hepatic Gluconeogenesis via Phosphorylation of FOXO1 and AMPK in Obese Diabetic db/db Mice. Nutrients 2020, 12, 2546. https://doi.org/10.3390/nu12092546
Kim HK, Jeong J, Kang EY, Go G-w. Red Pepper (Capsicum annuum L.) Seed Extract Improves Glycemic Control by Inhibiting Hepatic Gluconeogenesis via Phosphorylation of FOXO1 and AMPK in Obese Diabetic db/db Mice. Nutrients. 2020; 12(9):2546. https://doi.org/10.3390/nu12092546
Chicago/Turabian StyleKim, Hyun Kyung, Jeongho Jeong, Eun Young Kang, and Gwang-woong Go. 2020. "Red Pepper (Capsicum annuum L.) Seed Extract Improves Glycemic Control by Inhibiting Hepatic Gluconeogenesis via Phosphorylation of FOXO1 and AMPK in Obese Diabetic db/db Mice" Nutrients 12, no. 9: 2546. https://doi.org/10.3390/nu12092546