
Purification of Polysaccharide Produced by the Haploid Yeast Strain of Tremella sanguinea and Its Antioxidant and Prebiotic Activities

Abstract
:1. Introduction
2. Results and Discussion
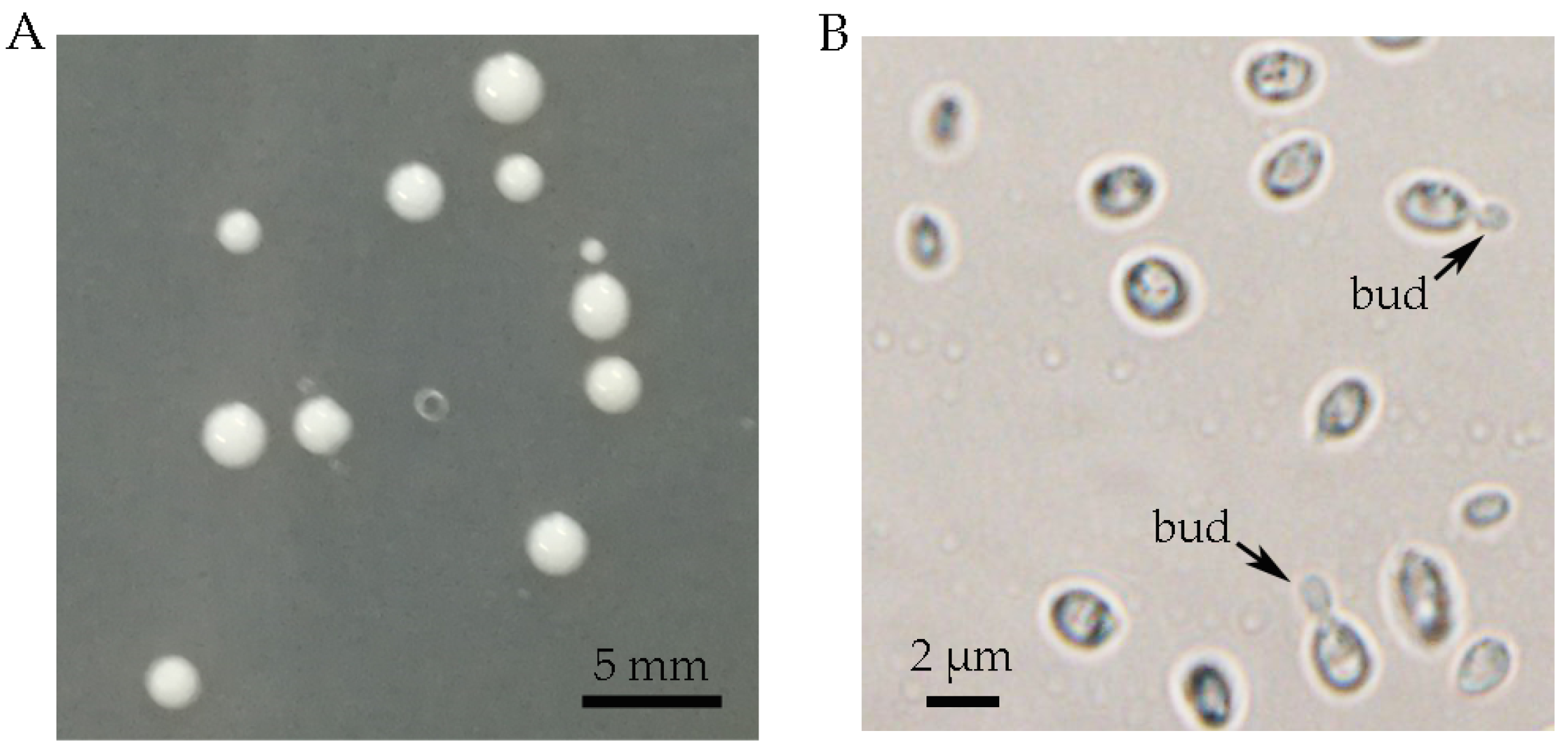
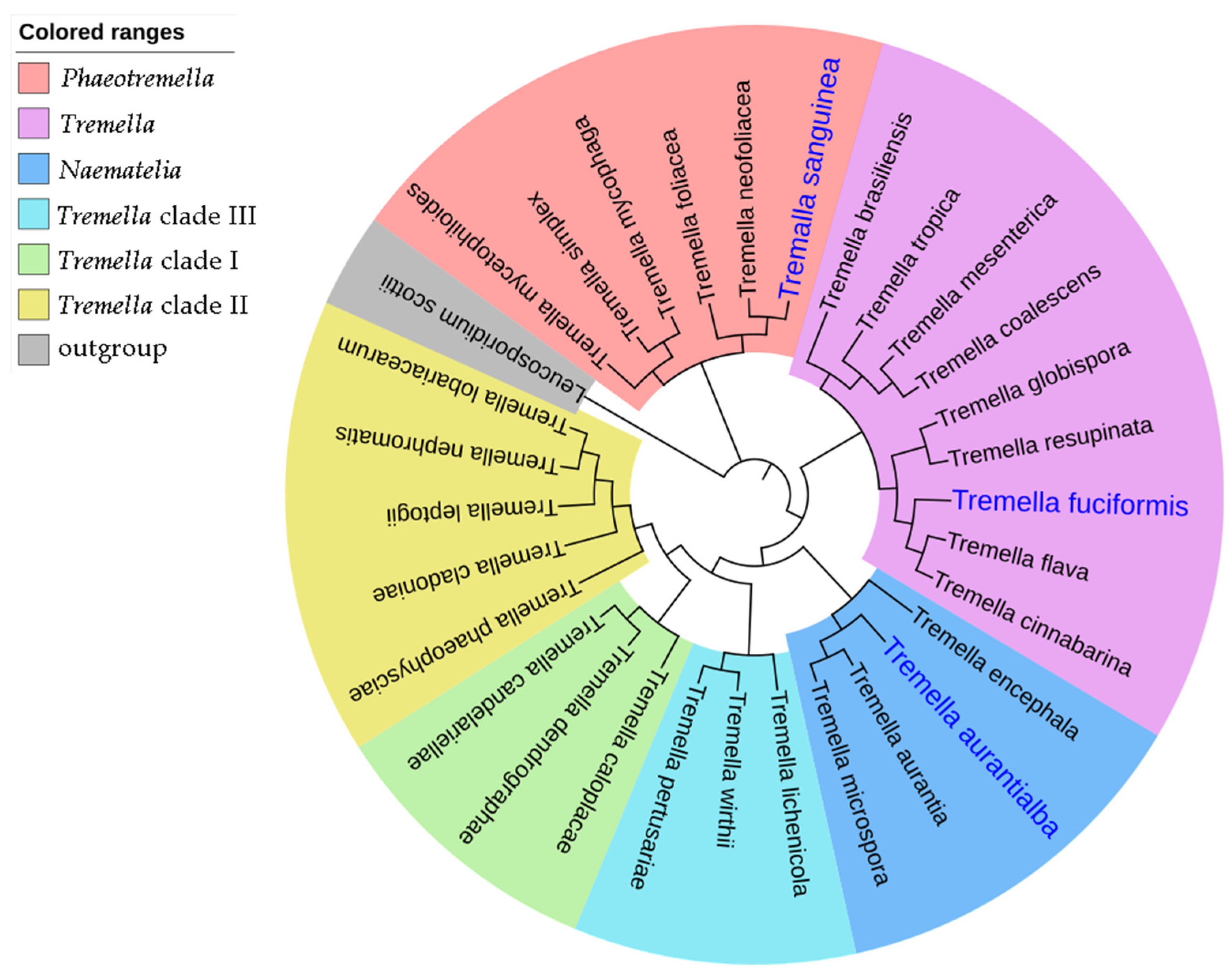
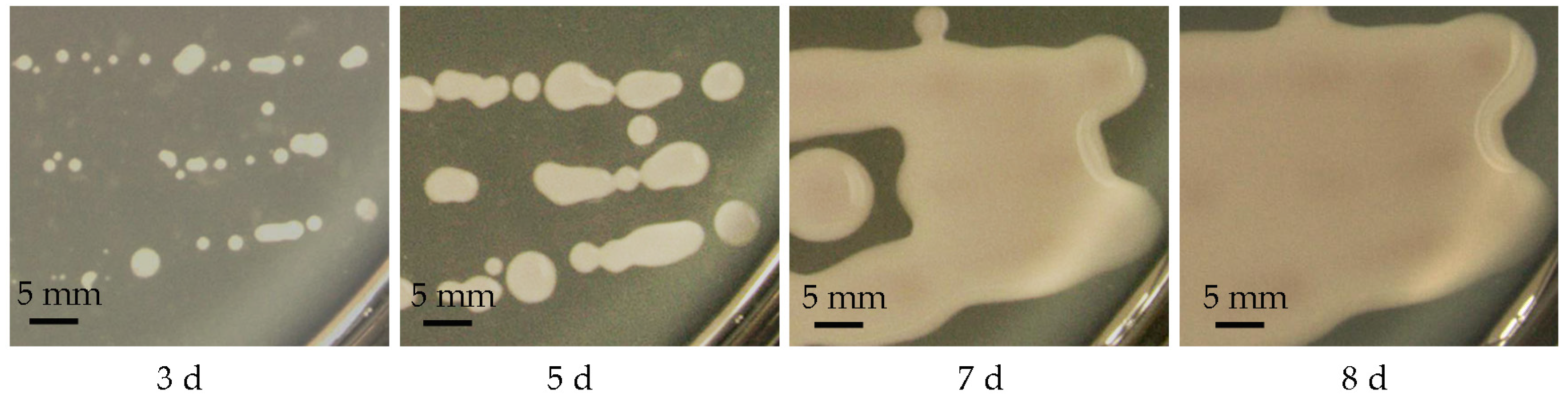
2.1. Isolation of the Haploid Yeast Strain of T. sanguinea
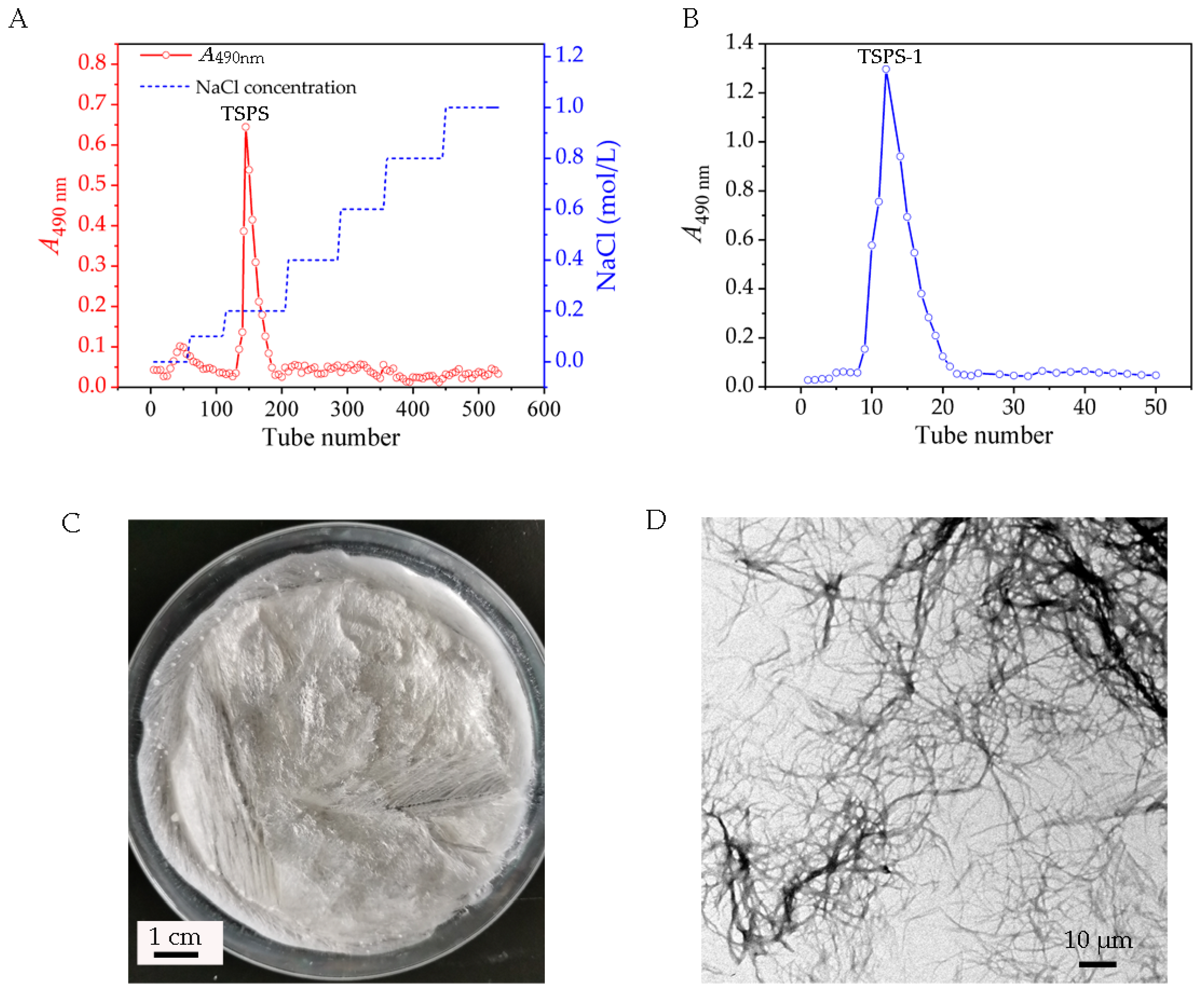
2.2. Extraction and Purification of EPS from the Haploid Yeast Strain of T. sanguinea
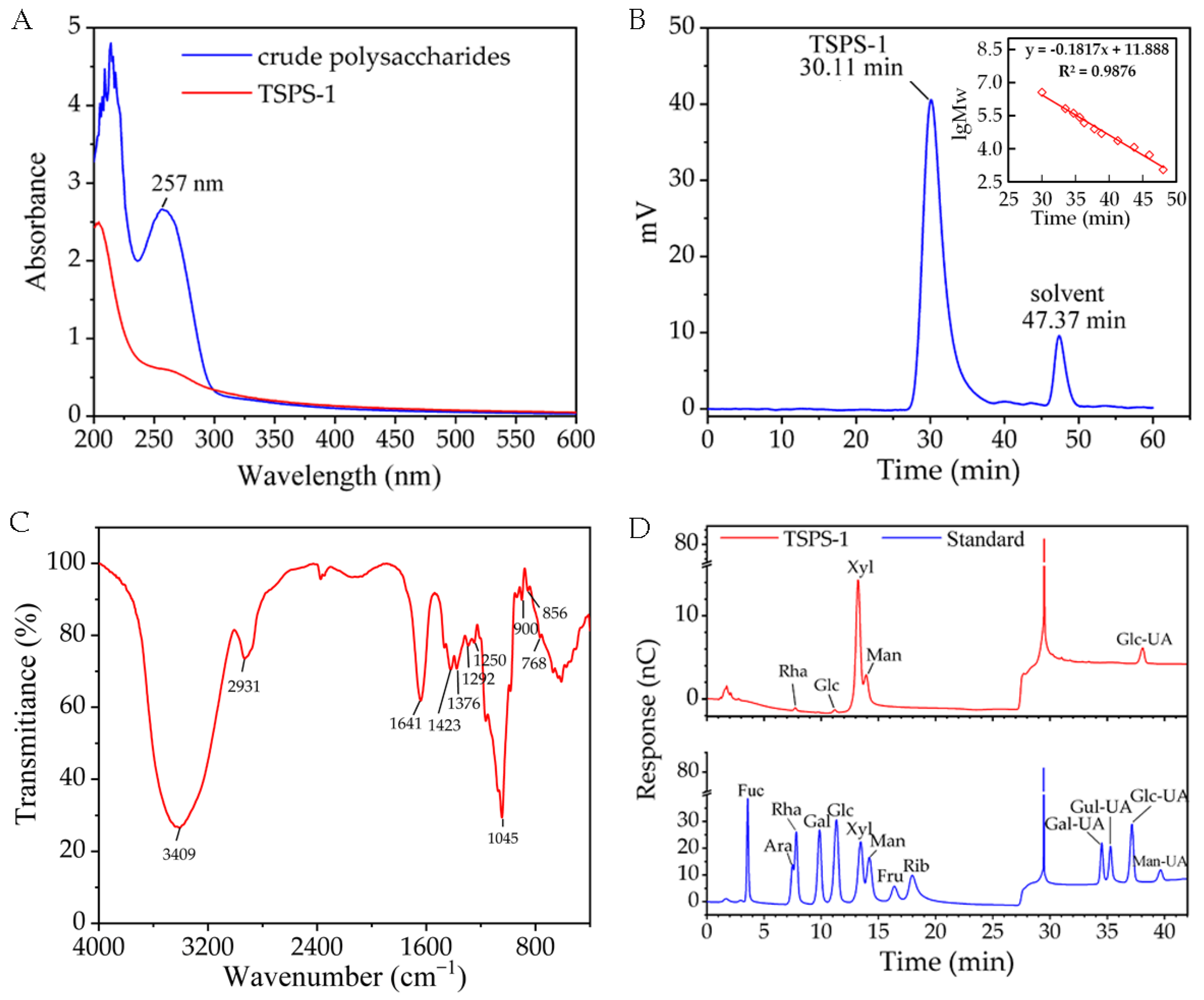
2.3. Structural Analysis of the Polysaccharide TSPS-1
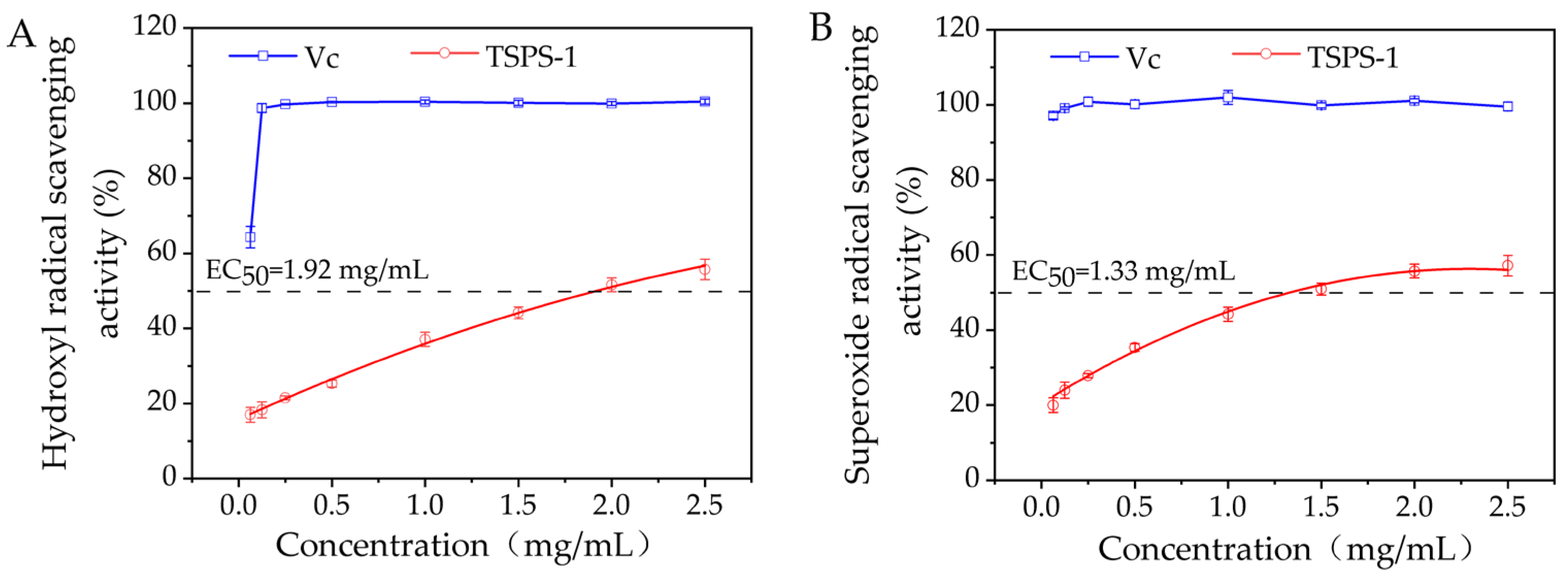
2.4. Antioxidant Activity of the Polysaccharide TSPS-1
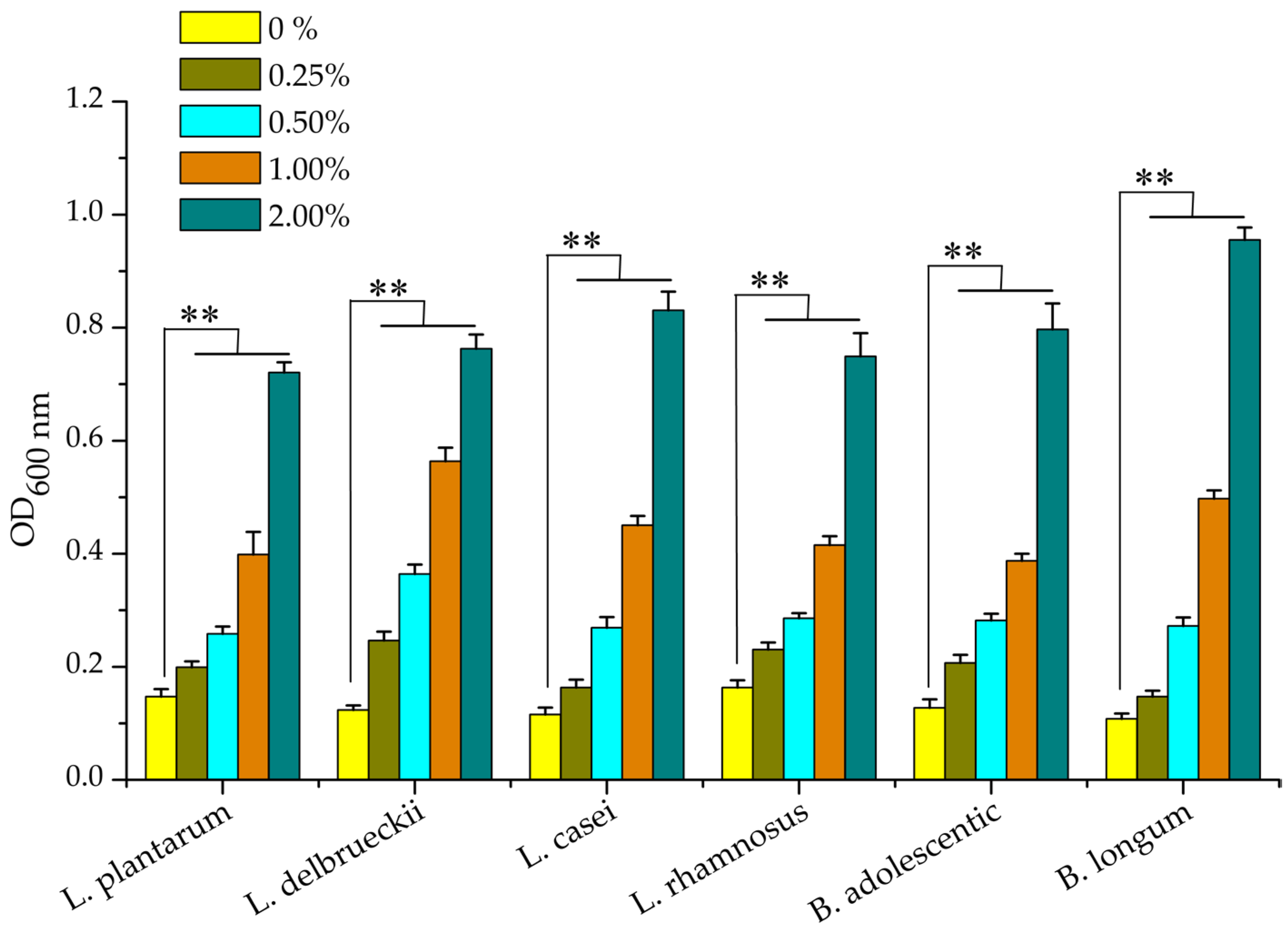
2.5. Effect of the Polysaccharide TSPS-1 on the Proliferation of Probiotics
3. Materials and Methods
3.1. Isolation of the Haploid Yeast Strain of T. sanguinea
3.2. Phylogenetic Analysis
3.3. Preparation of Crude Polysaccharides
3.4. Purification of the Polysaccharide TSPS-1
3.5. UV-Vis and FT-IR Analysis
3.6. Microscopic Analysis
3.7. Determination of Molecular Weight
3.8. Analysis of Monosaccharide Composition
3.9. Determination of Antioxidant Activities in Vitro
3.10. Probiotic Bacteria Growth Stimulation
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Wang, Z.; Zeng, Y.; Luo, D. Structure elucidation of a non-branched and entangled heteropolysaccharide from Tremella sanguinea Peng and its antioxidant activity. Carbohydr. Polym. 2016, 151, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Ding, B.M.; Wu, Q.H.; Yang, H.L.; Wang, Y.; Li, L. Optimization of ultrasound-assisted extraction of pigment from Tremella sanguinea. Int. J. Agric. Bio. 2019, 22, 900–906. [Google Scholar]
- Sun, T.; Wang, R.; Sun, D.; Li, S.; Xu, H.; Qiu, Y.; Lei, P.; Sun, L.; Xu, X.; Zhu, Y. High-efficiency production of Tremella aurantialba polysaccharide through basidiospore fermentation. Bioresour. Technol. 2020, 318, 124268. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yang, M.; He, Y.; Zhai, C.; Li, C. A review on the production, structure, bioactivities and applications of Tremella polysaccharides. Int. J. Immunopathol. Pharmacol. 2021, 35, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Gao, Y.Q.; Zhang, M.M.; Lai, L.; Liu, Y.P.; Bao, H.Y.; Fu, Y.H. Chemical constituents from fruiting bodies of Tremella sanguinea. China J. Chin. Mater. Medica 2019, 44, 1836–1841. [Google Scholar] [CrossRef]
- Yang, D.; Liu, Y.; Zhang, L. Tremella polysaccharide: The molecular mechanisms of its drug action. Prog. Mol. Biol. Transl. Sci. 2019, 163, 383–421. [Google Scholar] [CrossRef]
- Wu, D.T.; An, L.Y.; Liu, W.; Hu, Y.C.; Wang, S.P.; Zou, L. In vitro fecal fermentation properties of polysaccharides from Tremella fuciformis and related modulation effects on gut microbiota. Food Res. Int. 2022, 156, 111185. [Google Scholar] [CrossRef]
- Yan, Y.H.; Wang, M.T.; Chen, N.; Wang, X.; Fu, C.H.; Li, Y.M.; Gan, X.R.; Lv, P.; Zhang, Y. Isolation, structures, bioactivities, application and future prospective for polysaccharides from Tremella aurantialba: A review. Front. Immunol. 2022, 13, 1091210. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, S.; Zeng, Y.; Luo, D. Extraction optimization, microstructure and anti-inflammatory properties of a polysaccharide from Tremella samguinea Peng. J. Chin. Inst. Food Sci. Technol. 2019, 19, 96–102. [Google Scholar] [CrossRef]
- Niu, Y.; Wang, X.; Yao, X.; Wang, J.; Bao, H. Immunomodulatory activities of polysaccharides from Tremella samguinea Peng. Farm Prod. Process. 2022, 550, 1–3. [Google Scholar] [CrossRef]
- Liu, X.Z.; Wang, Q.M.; Göker, M.; Groenewald, M.; Kachalkin, A.V.; Lumbsch, H.T.; Millanes, A.M.; Wedin, M.; Yurkov, A.M.; Boekhout, T.; et al. Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Z.; Wang, Q.M.; Theelen, B.; Groenewald, M.; Bai, F.Y.; Boekhout, T. Phylogeny of tremellomycetous yeasts and related dimorphic and filamentous basidiomycetes reconstructed from multiple gene sequence analyses. Stud. Mycol. 2015, 81, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Spirin, V.; Malysheva, V.; Yurkov, A.; Miettinen, O.; Larsson, K.H. Studies in the Phaeotremella foliacea group (Tremellomycetes, Basidiomycota). Myc. Mycol. 2018, 17, 451–466. [Google Scholar] [CrossRef]
- Zheng, Q.W.; He, B.L.; Wang, J.Y.; Huang, S.S.; Zou, Y.; Wei, T.; Ye, Z.W.; Guo, L.Q.; Lin, J.F. Structural analysis and antioxidant activity of extracellular polysaccharides extracted from vulinary-medicinal white jelly mushroom Tremella fuciformis (Tremellomycetes) conidium cells. Int. J. Med. Mushrooms 2020, 22, 489–500. [Google Scholar] [CrossRef]
- Zhu, H.; Cao, C.X.; Zhang, S.S.; Zhang, Y.; Zou, W.S. pH-control modes in a 5-L stirred-tank bioreactor for cell biomass and exopolysaccharide production by Tremella fuciformis spore. Bioresour. Technol. 2011, 102, 9175–9178. [Google Scholar] [CrossRef]
- Zhu, H.; Sun, S. Effect of constant glucose feeding on the production of exopolysaccharides by Tremella fuciformis spores. Appl. Biochem. Biotech. 2009, 152, 366–371. [Google Scholar] [CrossRef]
- Peng, Y. A new species of the genus Tremella from China. Acta Sci. Nat. Univ. Norm. 1990, 13, 253–256. [Google Scholar]
- Joseph, S. Characterization of a Pigmented Yeast and Its Slime. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 1959. [Google Scholar]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. Yeast exopolysaccharides and their physiological functions. Folia Microbiol. 2021, 66, 171–182. [Google Scholar] [CrossRef]
- Gientka, I.; Blazejak, S.; Stasiak-Rozanska, L.; Chlebowska-Smigiel, A. Exopolysaccharides from yeast: Insight into optimal conditions for biosynthesis, chemical composition and functional properties-review. Acta Sci. Pol. Technol. Aliment. 2015, 14, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zou, P.; Zhao, H.; Qiu, J.; Regenstein, J.M.; Yang, X. Isolation, purification, structure and antioxidant activity of polysaccharide from pinecones of Pinus koraiensis. Carbohydr. Polym. 2020, 251, 117078. [Google Scholar] [CrossRef]
- Hu, D.; Bao, T.; Lu, Y.; Su, H.; Chen, W. Polysaccharide from mulberry mruit (Morus alba L.) protects against palmitic-acid-induced hepatocyte lipotoxicity by activating the Nrf2/ARE signaling pathway. J. Agr. Food Chem. 2020, 68, 13016–13024. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Yan, L.; Zhang, Q.; Zhu, J.; Na, H.; Wang, Z. Chemical compositions and antioxidant activities of polysaccharides from the sporophores and cultured products of Armillaria mellea. Molecules 2015, 20, 5680–5697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Liu, Z.; Zhao, J.; Chen, R.; Meng, F.; Zhang, M.; Ge, W. Antioxidant and immunobiological activity of water-soluble polysaccharide fractions purified from Acanthopanax senticosu. Food Chem. 2011, 127, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Yan, Y.; Mi, J.; Zhang, H.; Lu, L.; Luo, Q.; Li, X.; Zeng, X.; Cao, Y. Simulated digestion and fermentation in vitro by human gut microbiota of polysaccharides from bee collected pollen of Chinese wolfberry. J. Agr. Food Chem. 2018, 66, 898–907. [Google Scholar] [CrossRef]
- Feng, K.; Chen, W.; Sun, L.; Liu, J.; Zhao, Y.; Li, L.; Wang, Y.; Zhang, W. Optimization extraction, preliminary characterization and antioxidant activity in vitro of polysaccharides from Stachys sieboldii Miq. tubers. Carbohydr. Polym. 2015, 125, 45–52. [Google Scholar] [CrossRef]
- Natalia, N.J.; Renata, N.; Marek, J.; Anna, M.; Marta, L.; Wojciech, R.; Zbigniew, K. New biological activity of the polysaccharide fraction from Cantharellus cibarius and its structural characterization. Food Chem. 2018, 268, 355–361. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strains | GenBank Numbers |
|---|---|---|
| Tremella mycetophiloides * (Phaeotremella mycetophiloides) | DSM 5728 | MF580587.1 |
| T. mycophaga * (P. mycophaga) | RJB6539-4 | AF042431.1 |
| T. simplex * (P. simplex) | FO31782 | AF042428.1 |
| T. foliacea * (P. simplex) | CBS 6969 | AF444431.1 |
| T. neofoliacea * (P. simplex) | CCJ1204 | AF042415.1 |
| T. brasiliensis | CBS 6966 | AF444429.1 |
| T. mesenterica | CBS 6973 | AF444433.1 |
| T. tropica | CCJ1355 | AF042433.1 |
| T. coalescens | CBS6967 | KF036601.1 |
| T. resupinata | CCJ1458 | AF042421.1 |
| T. fuciformis | CBS 6970 | NR_155936.1 |
| T. cinnabarina | CBS 8234 | NR_155933.1 |
| T. globispora | CBS 6972 | NR_155889.1 |
| T. flava | CBS 8471 | NR_155935.1 |
| T. aurantia * (Naematelia aurantia) | CBS 6965 | NR_155873.1 |
| T. encephala * (N. encephala) | CCJ925 | AF042404.1 |
| T. microspora * (N. microspora) | BPI702328 | AF042435.1 |
| T. aurantialba * (N. aurantialba) | 9901 | DQ400104.1 |
| T. lichenicola | AM18 | JN053504.1 |
| T. pertusariae | AM2 | JN053494.1 |
| T. wirthii | AM90 | JN053492.1 |
| T. caloplacae | AM31 | JN053468.1 |
| T. candelariellae | AM34 | JN053470.1 |
| T. dendrographae | AM39 | JN053471.1 |
| T. leptogii | AM81 | JN053476.1 |
| T. lobariacearum | AM118 | JN053474.1 |
| T. nephromatis | AM133 | JN053475.1 |
| T. phaeophysciae | AM121 | JN053480.1 |
| T. cladoniae | AM84 | JN053478.1 |
| Leucosporidium scottii | CBS 5930 | NR_073267.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Chen, S.; Zhang, J.; Gao, M.; Li, L. Purification of Polysaccharide Produced by the Haploid Yeast Strain of Tremella sanguinea and Its Antioxidant and Prebiotic Activities. Molecules 2023, 28, 5391. https://doi.org/10.3390/molecules28145391
Liu Y, Chen S, Zhang J, Gao M, Li L. Purification of Polysaccharide Produced by the Haploid Yeast Strain of Tremella sanguinea and Its Antioxidant and Prebiotic Activities. Molecules. 2023; 28(14):5391. https://doi.org/10.3390/molecules28145391
Chicago/Turabian StyleLiu, Yichi, Suo Chen, Jialan Zhang, Mengxiang Gao, and Li Li. 2023. "Purification of Polysaccharide Produced by the Haploid Yeast Strain of Tremella sanguinea and Its Antioxidant and Prebiotic Activities" Molecules 28, no. 14: 5391. https://doi.org/10.3390/molecules28145391