
Alkaloidal Phytoconstituents for Diabetes Management: Exploring the Unrevealed Potential
 , ,
, , 
Abstract
:1. Introduction
2. Role of Medicinal Plants in the Management of Diabetes Mellitus
2.1. Alkaloids for Treatment of Diabetes Mellitus
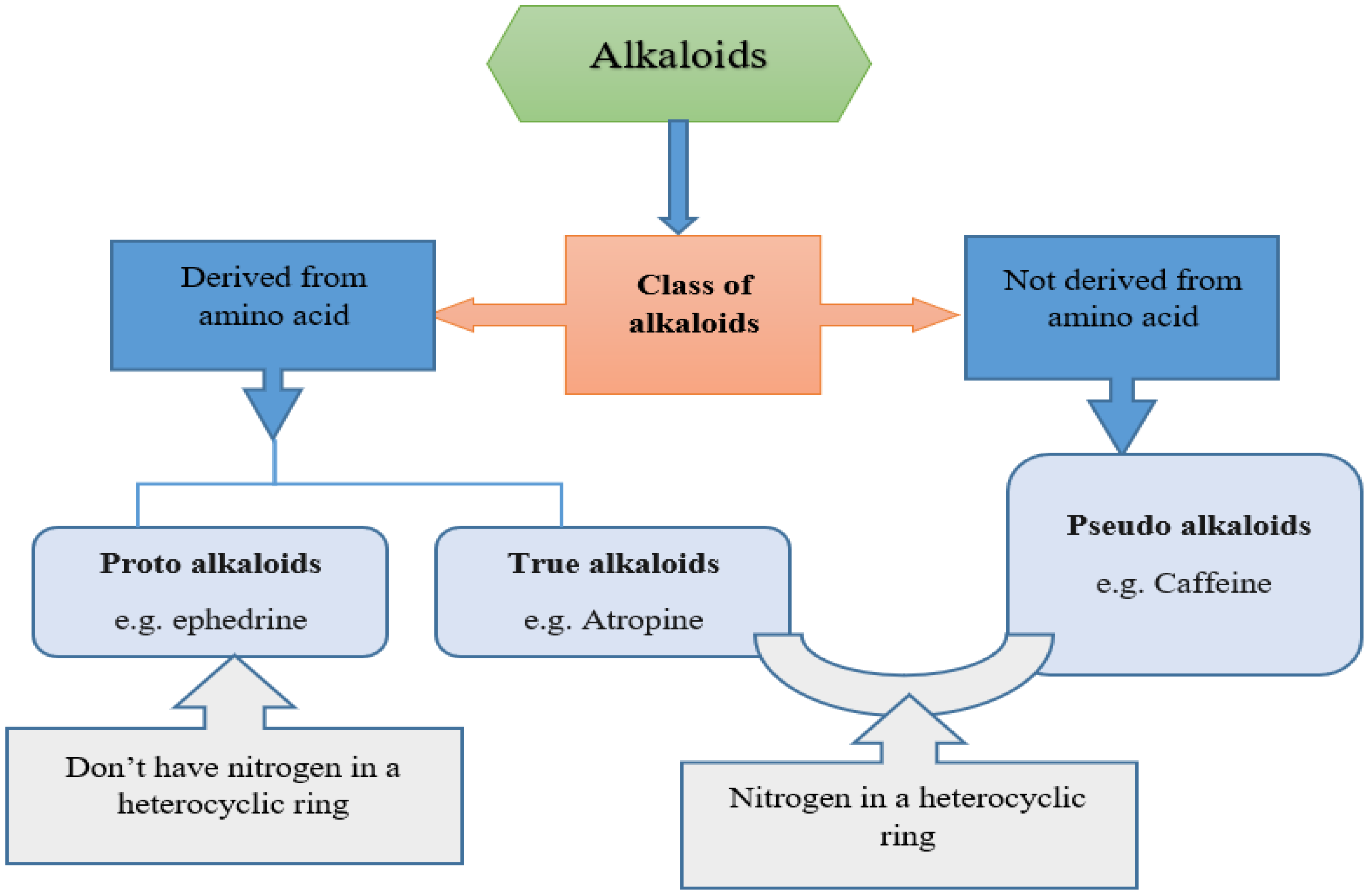
2.2. Classification of Alkaloids
2.3. Source of Various Alkaloidal Phytoconstituents Used in the Management of Diabetes Mellitus
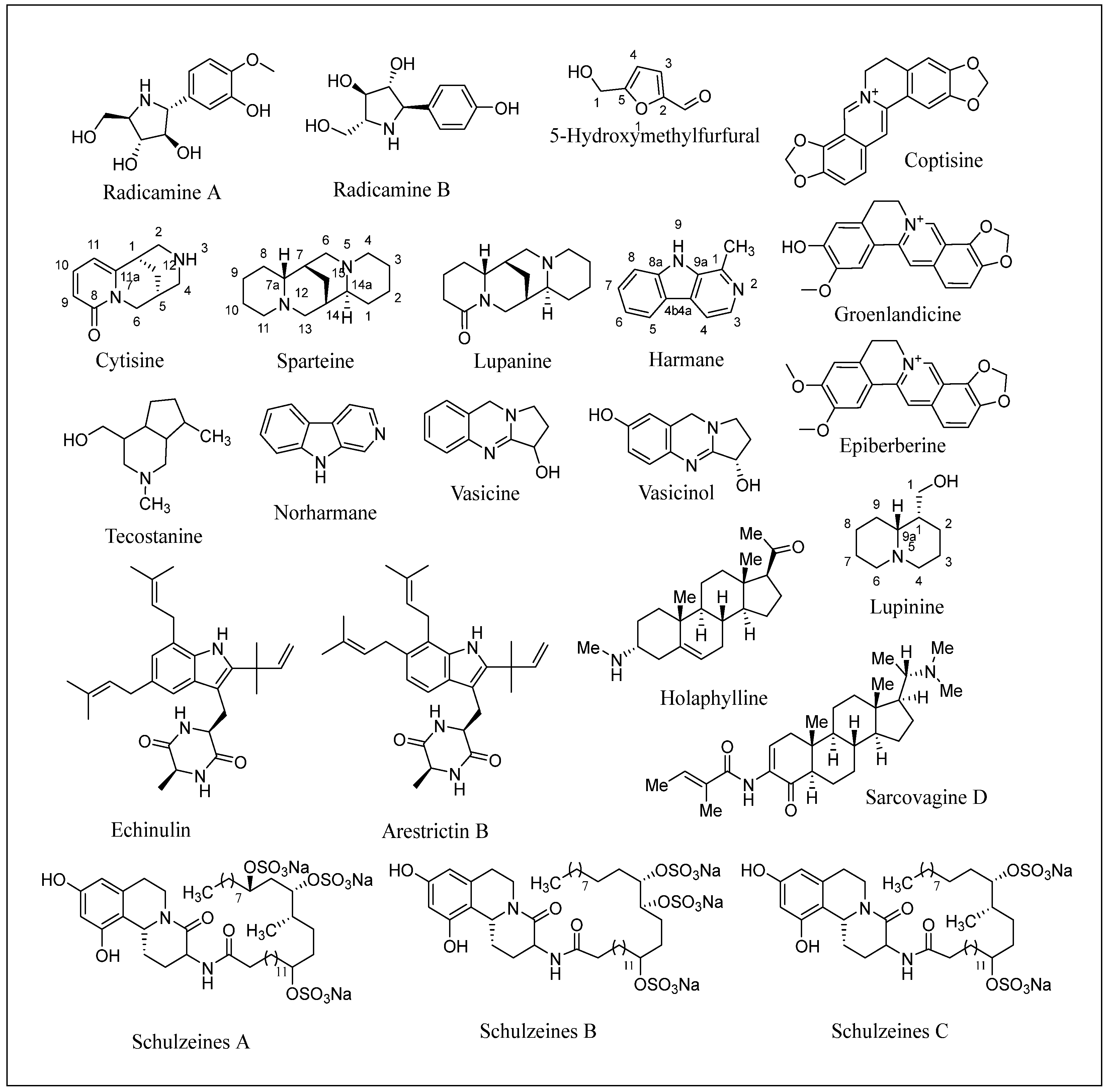
2.3.1. Berberine
2.3.2. Catharanthine, Vindoline and Vindolinine
2.3.3. Calystegine B2
2.3.4. Cryptolepine
2.3.5. Harmane, Norharmane, Pinoline and Jambosine
2.3.6. Jatrorrhizine, Magnoflorine, Palmatine, Tembetarine
2.3.7. Lepidine and Semilepidine
2.3.8. Mahanimbine
2.3.9. Piperumbellactam A, B, and C
2.3.10. Radicamines A and B
2.3.11. Schulzeines A, B, and C
2.3.12. Other Sources
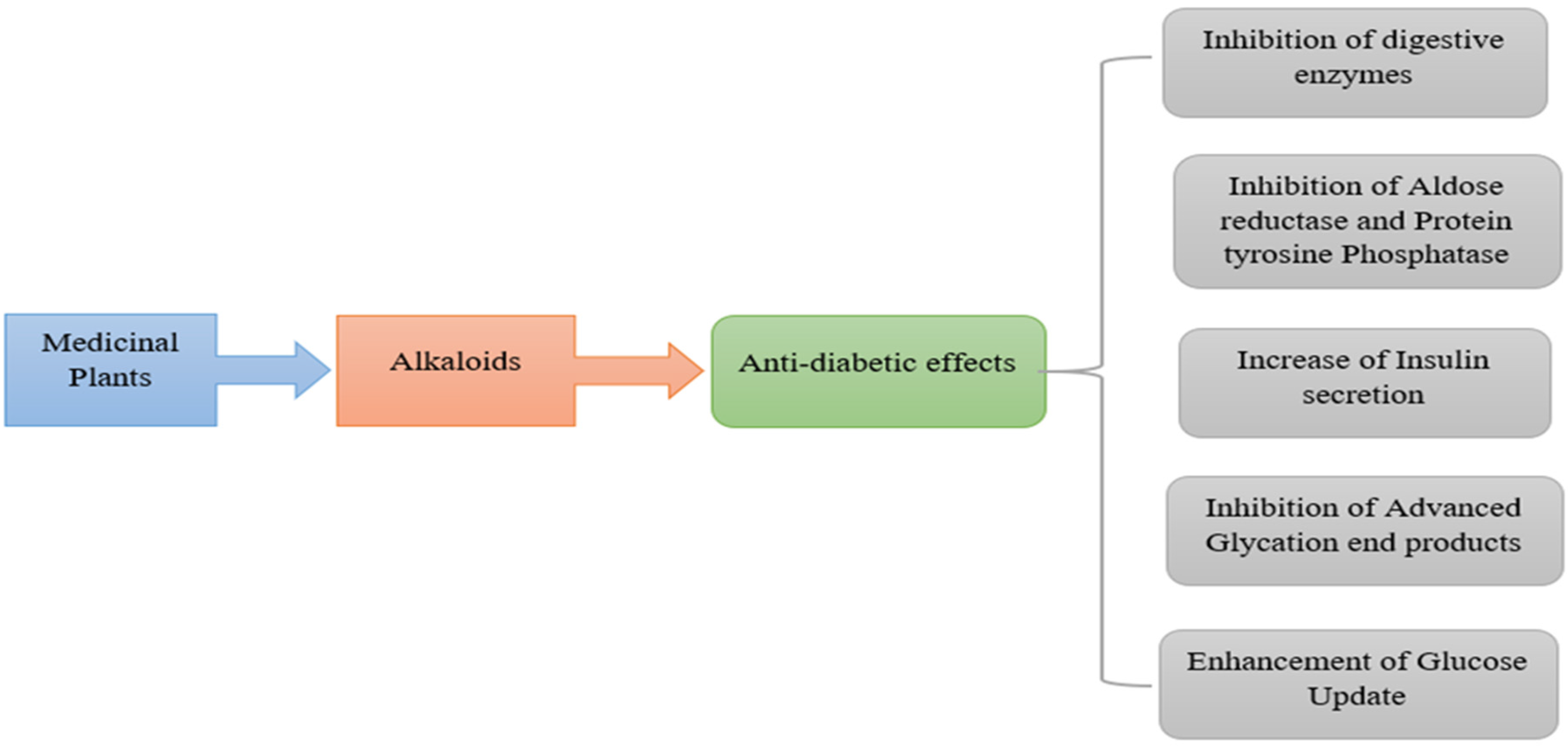
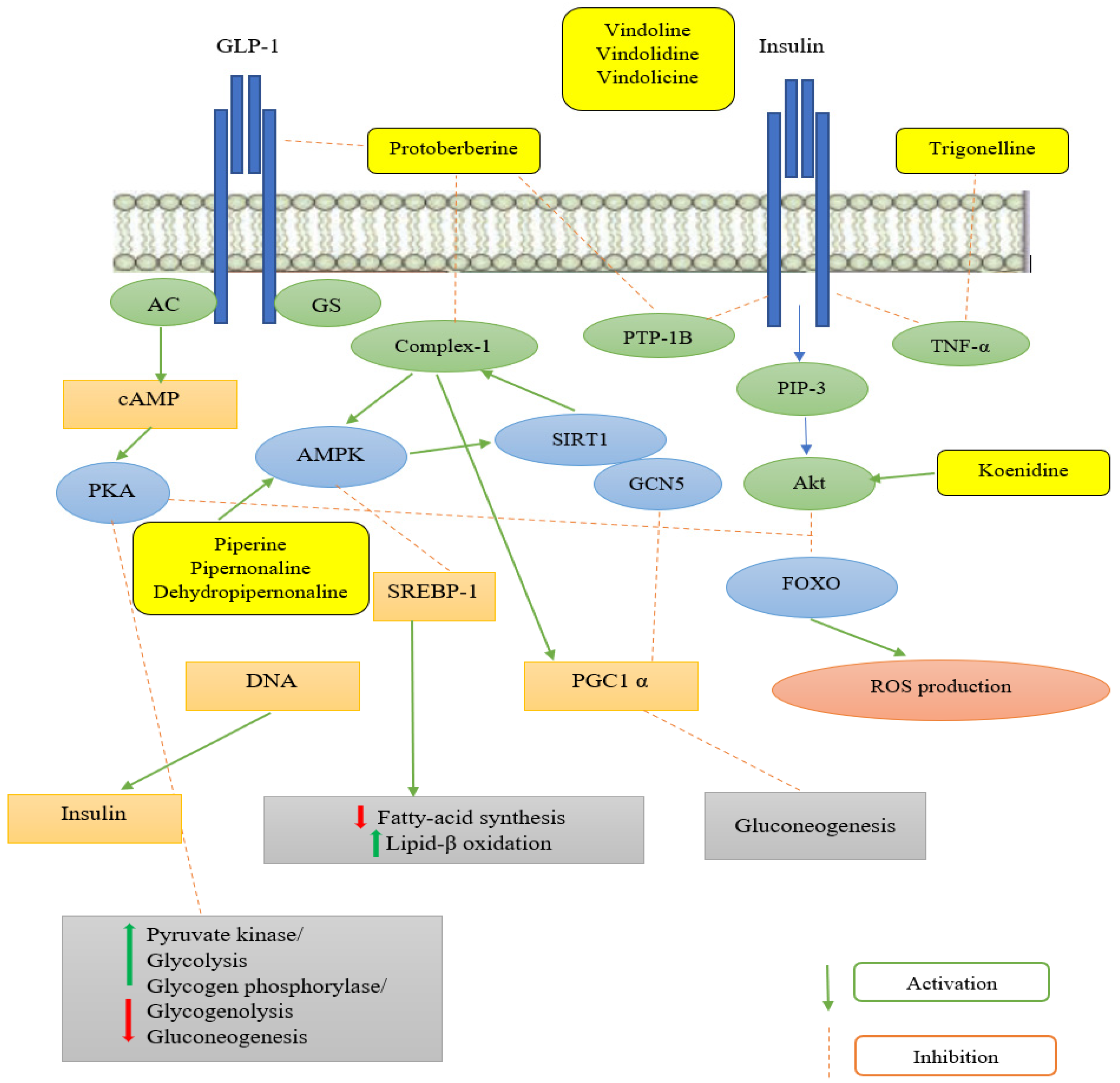
3. Mechanism of Alkaloidal Phytoconstituents in Diabetic Complications
3.1. Inhibition of Digestive Enzymes
3.2. Suppression of Aldose Reductase and Protein Tyrosine Phosphatase–1B
3.3. Enhancement of Insulin Release
3.4. Inhibition of Advanced Glycation End Products
3.5. Enhancement of Glucose Uptake
4. Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| DM | Diabetes mellitus |
| DPP-4 | Dipeptidyl peptidase-4 |
| G-6Pase | Glucose-6-phosphatase |
| GIP | Glucose-based insulin tropic polypeptide |
| GLP-1 | Glucagon-like peptide-1 |
| GLUT-4 | Glucose transporter type 4 |
| GSK3B | Glycogen synthase kinase-3 beta |
| INSR | Insulin receptor |
| MAPK1 | Mitogen-activated protein kinase 1 |
| NIDDM | Non-insulin-dependent diabetes mellitus |
| PTP1B | Protein tyrosine phosphatase1B |
| T1D | Type 1 diabetes |
| T2D | Type 2 diabetes |
| TNF | Tumor necrosis factor |
| WHO | World Health Organization |
References
- Definition, Diagnosis and Classification of Diabetes Mellitus and Its Complications: Report of a WHO Consultation. Part 1, Diagnosis and Classification of Diabetes Mellitus. Available online: https://apps.who.int/iris/handle/10665/66040 (accessed on 1 August 2022).
- Gaikwad, S.B.; Krishna Mohan, G.; Rani, M.S. Phytochemicals for Diabetes Management. Pharm. Crop. 2014, 5, 11–28. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Bai, K.J.; Lin, H.H.; Chien, S.T.; Lee, J.J.; Enarson, D.A.; Lee, T.I.; Yu, M.C. The Influence of Diabetes, Glycemic Control, and Diabetes-Related Comorbidities on Pulmonary Tuberculosis. PLoS ONE 2015, 10, e0121698. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.W.; Morrato, E.H.; Ghushchyan, V.; Wyatt, H.R.; Hill, J.O. Obesity, Inactivity, and the Prevalence of Diabetes and Diabetes-Related Cardiovascular Comorbidities in the U.S. 2000–2002. Diabetes Care 2005, 28, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Simmons, K.M.; Mitchell, A.M.; Alkanani, A.A.; McDaniel, K.A.; Baschal, E.E.; Armstrong, T.; Pyle, L.; Yu, L.; Michels, A.W. Failed Genetic Protection: Type 1 Diabetes in the Presence of Hla-Dqb1*06:02. Diabetes 2020, 69, 1763–1769. [Google Scholar] [CrossRef]
- Govindappa, M. A Review on Role of Plant(s) Extracts and Its Phytochemicals for the Management of Diabetes. J. Diabetes Metab. 2015, 6, 7. [Google Scholar] [CrossRef]
- Gregg, E.W.; Sattar, N.; Ali, M.K. The Changing Face of Diabetes Complications. Lancet Diabetes Endocrinol. 2016, 4, 537–547. [Google Scholar] [CrossRef]
- Pavic, M.; Premuzic, V.; Pavic, A.Z.; Bevanda, M.; Mihaljevic, S.; Oreskovic, S. Prevalence of Gestational Diabetes Mellitus and Perinatal Outcomes According to the Old Who Criteria and IADPSG Criteria. Psychiatr. Danub. 2021, 33, 30–36. [Google Scholar]
- Wu, Y.; Ding, Y.; Tanaka, Y.; Zhang, W. Risk Factors Contributing to Type 2 Diabetes and Recent Advances in the Treatment and Prevention. Int. J. Med. Sci. 2014, 11, 1185. [Google Scholar] [CrossRef]
- Chen, J.; Hui, S.T.; Couto, F.M.; Mungrue, I.N.; Davis, D.B.; Attie, A.D.; Lusis, A.J.; Davis, R.A.; Shalev, A. Thioredoxin-interacting Protein Deficiency Induces Akt/Bcl-xL Signaling and Pancreatic Beta-cell Mass and Protects against Diabetes. FASEB J. 2008, 22, 3581–3594. [Google Scholar] [CrossRef]
- Schardl, C.L.; Panaccione, D.G.; Tudzynski, P. Chapter 2 Ergot Alkaloids-Biology and Molecular Biology. Alkaloids. Chem. Biol. 2006, 63, 45–86. [Google Scholar]
- Panaccione, D.G.; Beaulieu, W.T.; Cook, D. Bioactive Alkaloids in Vertically Transmitted Fungal Endophytes. Funct. Ecol. 2014, 28, 299–314. [Google Scholar] [CrossRef]
- Helander, M.; Phillips, T.; Faeth, S.H.; Bush, L.P.; McCulley, R.; Saloniemi, I.; Saikkonen, K. Alkaloid Quantities in Endophyte-Infected Tall Fescue Are Affected by the Plant-Fungus Combination and Environment. J. Chem. Ecol. 2016, 42, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A Major Update to the DrugBank Database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Barshop, K.; Staller, K. Eluxadoline in Irritable Bowel Syndrome with Diarrhea: Rationale, Evidence and Place in Therapy. Adv. Chronic. Dis. 2017, 8, 153–160. [Google Scholar] [CrossRef]
- Prabhakar, P.K.; Kumar, A.; Doble, M. Combination Therapy: A New Strategy to Manage Diabetes and Its Complications. Phytomedicine 2014, 21, 123–130. [Google Scholar] [CrossRef]
- Rasouli, H.; Yarani, R.; Pociot, F.; Popović-Djordjević, J. Anti-Diabetic Potential of Plant Alkaloids: Revisiting Current Findings and Future Perspectives. Pharm. Res. 2020, 155, 104723. [Google Scholar] [CrossRef]
- Katiyar, D.; Singh, V.; Gilani, S.J.; Goel, R.; Grover, P.; Vats, A. Hypoglycemic Herbs and Their Polyherbal Formulations: A Comprehensive Review. Med. Chem. Res. 2015, 24, 1–21. [Google Scholar] [CrossRef]
- Singh, T.G.; Sharma, R.; Kaur, A.; Dhiman, S.; Singh, R. Evaluation of Renoprotective Potential of Ficus Religiosa in Attenuation of Diabetic Nephropathy in Rats. Obes. Med. 2020, 19, 100268. [Google Scholar] [CrossRef]
- Rizvi, S.I.; Mishra, N. Traditional Indian Medicines Used for the Management of Diabetes Mellitus. J. Diabetes Res. 2013, 2013, 712092. [Google Scholar] [CrossRef]
- Xu, F.; Wu, H.; Wang, X.; Yang, Y.; Wang, Y.; Qian, H.; Zhang, Y. RP-HPLC Characterization of Lupenone and β-Sitosterol in Rhizoma Musae and Evaluation of the Anti-Diabetic Activity of Lupenone in Diabetic Sprague-Dawley Rats. Molecules 2014, 19, 14114–14127. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Maiti, K.; Mukherjee, K.; Houghton, P.J. Leads from Indian Medicinal Plants with Hypoglycemic Potentials. J. Ethnopharmacol 2006, 106, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Belyagoubi-Benhammou, N.; Belyagoubi, L.; Gismondi, A.; di Marco, G.; Canini, A.; Atik Bekkara, F. GC/MS Analysis, and Antioxidant and Antimicrobial Activities of Alkaloids Extracted by Polar and Apolar Solvents from the Stems of Anabasis Articulata. Med. Chem. Res. 2019, 28, 754–767. [Google Scholar] [CrossRef]
- Mechanick, J.I.; Marchetti, A.; Hegazi, R.; Hamdy, O. Diabetes-Specific Nutrition Formulas in the Management of Patients with Diabetes and Cardiometabolic Risk. Nutrients 2020, 12, 3616. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.Y.; Huang, Z.J.; Wang, G.J.; Fawcett, J.P.; Liu, X.D.; Zhao, X.C.; Sun, J.G.; Xie, Y.Y. The Antihyperglycaemic Activity of Berberine Arises from a Decrease of Glucose Absorption. Planta Med. 2003, 69, 632–636. [Google Scholar] [CrossRef]
- Chattopadhyay, R.R. A Comparative Evaluation of Some Blood Sugar Lowering Agents of Plant Origin. J. Ethnopharmacol. 1999, 67, 367–372. [Google Scholar] [CrossRef]
- Shehadeh, M.B.; Suaifan, G.A.R.Y.; Abu-Odeh, A.M. Plants Secondary Metabolites as Blood Glucose-Lowering Molecules. Molecules 2021, 26, 4333. [Google Scholar] [CrossRef]
- Azam, K.; Rasheed, M.A.; Omer, M.O.; Altaf, I.; Akhlaq, A. Anti-Hyperlipidemic and Anti-Diabetic Evaluation of Ethanolic Leaf Extract of Catharanthus Roseus Alone and in Combination Therapy. Braz. J. Pharm. Sci. 2022, 58, 1–18. [Google Scholar] [CrossRef]
- Griffiths, R.C.; Watson, A.A.; Kizu, H.; Asano, N.; Sharp, H.J.; Jones, M.G.; Wormald, M.R.; Fleet, G.W.J.; Nash, R.J. The Isolation from Nicandra Physalodes and Identification of the 3-O-β-D-Glucopyranoside of 1α,2β,3α,6α-Tetrahydroxy-nor-Tropane (Calystegine B1). Tetrahedron. Lett. 1996, 37, 3207–3208. [Google Scholar] [CrossRef]
- Khacheba, I.; Boussoussa, H.; Djeridane, A.; Bekhaoua, A.; Bensayah, N.; Yousfi, M. α-Glucosidase Inhibitory Effect and Antioxidant Activity of the Extracts of Eighteen Plant Traditionally Used in Algeria for Diabetes. Curr. Enzym. Inhib. 2016, 13, 67–78. [Google Scholar] [CrossRef]
- Abesundara, K.J.M.; Matsui, T.; Matsumoto, K. α-Glucosidase Inhibitory Activity of Some Sri Lanka Plant Extracts, One of Which, Cassia Auriculata, Exerts a Strong Antihyperglycemic Effect in Rats Comparable to the Therapeutic Drug Acarbose. J. Agric. Food Chem. 2004, 52, 2541–2545. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, P.; Trivedi, N.; Jha, K.K.; Kumar, S.; Singh, B. A Review on Pharmacognostical and Pharmacological Activities of Plant Nicandra Physalodes. Pharma Res. 2014, 11, 42–47. [Google Scholar]
- Bierer, D.E.; Dubenko, L.G.; Zhang, P.; Lu, Q.; Imbach, P.A.; Garofalo, A.W.; Phuan, P.W.; Fort, D.M.; Litvak, J.; Gerber, R.E.; et al. Antihyperglycemic Activities of Cryptolepine Analogues: An Ethnobotanical Lead Structure Isolated from Cryptolepis Sanguinolenta. J. Med. Chem. 1998, 41, 2754–2764. [Google Scholar] [CrossRef]
- Osafo, N.; Mensah, K.B.; Yeboah, O.K. Phytochemical and Pharmacological Review of Cryptolepis Sanguinolenta (Lindl.) Schlechter. Adv. Pharm. Sci. 2017, 2017, 3026370. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.; Sarker, M.M.R.; Sultana, T.N.; Chowdhury, M.N.R.; Rashid, M.A.; Chaity, N.I.; Zhao, C.; Xiao, J.; Hafez, E.E.; Khan, S.A.; et al. Antidiabetic Phytochemicals From Medicinal Plants: Prospective Candidates for New Drug Discovery and Development. Front. Endocrinol. 2022, 13, 1–35. [Google Scholar] [CrossRef]
- Cooper, E.J.; Hudson, A.L.; Parker, C.A.; Morgan, N.G. Effects of the β-Carbolines, Harmane and Pinoline, on Insulin Secretion from Isolated Human Islets of Langerhans. Eur. J. Pharm. 2003, 482, 189–196. [Google Scholar] [CrossRef]
- Al-Eisa, R.A.; Tag, H.M.; ElNaggar, M.S.; Abdelrazek, H.M.A.; El-Shenawy, N.S. Evaluation of Tribulus Terrestris Extracts Relative to Metformin on Oxidative Stress and Histopathology of the Liver for Diabetic Male Rats. Diabetology 2022, 3, 4. [Google Scholar] [CrossRef]
- Sharma, P.; Dwivedee, B.P.; Bisht, D.; Dash, A.K.; Kumar, D. The Chemical Constituents and Diverse Pharmacological Importance of Tinospora Cordifolia. Heliyon 2019, 5, e02437. [Google Scholar] [CrossRef]
- Sharma, R.; Amin, H.; Galib; Prajapati, P.K. Antidiabetic Claims of Tinospora Cordifolia (Willd.) Miers: Critical Appraisal and Role in Therapy. Asian Pac. J. Trop. Biomed. 2015, 5, 68–78. [Google Scholar] [CrossRef]
- Khanal, P.; Patil, B.M.; Mandar, B.K.; Dey, Y.N.; Duyu, T. Network Pharmacology-Based Assessment to Elucidate the Molecular Mechanism of Anti-Diabetic Action of Tinospora Cordifolia. Clin. Phytosci. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Shukla, A.; Singh, C.S.; Bigoniya, P. Phytochemical and CNS Activity of Lepidium Sativum Linn. Seeds Total Alkaloid. Pharm. Lett. 2011, 3, 226–237. [Google Scholar]
- Attia, E.S.; Amer, A.H.; Hasanein, M.A. The Hypoglycemic and Antioxidant Activities of Garden Cress (Lepidium Sativum, L.) Seed on Alloxan-Induced Diabetic Male Rats. Nat. Prod. Res. 2019, 33, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.d.; Krishnakumar, K.; Jaganathan, S.; Mandal, M. Effect of Mangiferin and Mahanimbine on Glucose Utilization in 3T3-L1 Cells. Pharm. Mag. 2013, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Dineshkumar, B.; Mitra, A.; Mahadevappa, M. Antidiabetic and Hypolipidemic Effects of Mahanimbine (Carbazole Alkaloid) from Murraya Koenigii (Rutaceae) Leaves. Int. J. Phytomed. 2010, 2, 22–30. [Google Scholar] [CrossRef]
- Ge, Q.; Chen, L.; Yuan, Y.; Liu, L.; Feng, F.; Lv, P.; Ma, S.; Chen, K.; Yao, Q. Network Pharmacology-Based Dissection of the Anti-Diabetic Mechanism of Lobelia Chinensis. Front. Pharm. 2020, 11, 347. [Google Scholar] [CrossRef] [Green Version]
- Shibano, M.; Tsukamoto, D.; Masuda, A.; Tanaka, Y.; Kusano, G. Two New Pyrrolidine Alkaloids, Radicamines A and B, as Inhibitors of α-Glucosidase from Lobelia Chinensis Lour. Chem. Pharm. Bull. 2001, 49, 1362–1365. [Google Scholar] [CrossRef]
- Njateng, G.S.S.; Zaib, S.; Chimi, L.; Feudjio, C.; Mouokeu, R.S.; Gatsing, D.; Kuiate, J.R.; Adewole, E.; Iqbal, J. Antidiabetic Potential of Methanol Extracts from Leaves of Piper umbellatum L. and Persea Americana Mill. Asian Pac. J. Trop. Biomed. 2018, 8, 160. [Google Scholar] [CrossRef]
- Orhan, D.D.; Orhan, N.; Demir, O.; Konuklugil, B. Phenolic Content, Antioxidant and in Vitro Antidiabetic Effects of Thirteen Marine Organisms from Mediterranean Sea. Farmacia 2021, 69, 68–74. [Google Scholar] [CrossRef]
- Shakman, E.A.; ben Abdalha, A.; Talha, F.; Al-Faturi, A.; Bariche, M. First Records of Seven Marine Organisms of Different Origins from Libya (Mediterranean Sea). Bioinvasions Rec. 2017, 6, 377–382. [Google Scholar] [CrossRef]
- Marmouzi, I.; Ezzat, S.M.; Mostafa, E.S.; el Jemli, M.; Radwan, R.A.; Faouzi, M.E.A.; Tamsouri, N.; Kharbach, M. Isolation of Secondary Metabolites from the Mediterranean Sponge Species; Hemimycale Columella and Its Biological Properties. SN Appl. Sci. 2021, 3, 1–3. [Google Scholar] [CrossRef]
- Bajpai, M.B.; Asthana, R.K.; Sharma, N.K.; Chatterjee, S.K.; Mukherjee, S.K. Hypoglycemic Effect of Swerchirin from the Hexane Fraction of Swertia Chirayita. Planta Med. 1991, 57, 102–104. [Google Scholar] [CrossRef]
- Thomson, H.; Ojo, O.; Flatt, P.; AbdelWahab, Y. Antidiabetic Actions of Aqueous Bark Extract of Swertia Chirayita on Insulin Secretion, Cellular Glucose Uptake and Protein Glycation. J. Exp. Integr. Med. 2014, 4, 268–272. [Google Scholar] [CrossRef]
- Mishkinsky, J.; Joseph, B.; Sulman, F.G. Hypoglycaemic Effect of Trigonelline. Lancet 1967, 290, 1311–1312. [Google Scholar] [CrossRef]
- Geberemeskel, G.A.; Debebe, Y.G.; Nguse, N.A. Antidiabetic Effect of Fenugreek Seed Powder Solution (Trigonella Foenum-Graecum L.) on Hyperlipidemia in Diabetic Patients. J. Diabetes Res. 2019, 2019, 8507453. [Google Scholar] [CrossRef]
- Aguilar-Santamaría, L.; Ramírez, G.; Nicasio, P.; Alegría-Reyes, C.; Herrera-Arellano, A. Antidiabetic Activities of Tecoma Stans (L.) Juss. Ex Kunth. J. Ethnopharmacol. 2009, 124, 284–288. [Google Scholar] [CrossRef]
- Cushnie, T.P.T.; Cushnie, B.; Lamb, A.J. Alkaloids: An Overview of Their Antibacterial, Antibiotic-Enhancing and Antivirulence Activities. Int J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef]
- Costantino, L.; Raimondi, L.; Pirisino, R.; Brunetti, T.; Pessotto, P.; Giannessi, F.; Lins, A.P.; Barlocco, D.; Antolini, L.; El-Abady, S.A. Isolation and Pharmacological Activities of the Tecoma Stans Alkaloids. Farmaco 2003, 58, 781–785. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.; Yang, H.; Zhou, B. Chloroplast Genome of the Folk Medicine and Vegetable Plant Talinum paniculatum (Jacq.) Gaertn.: Gene Organization, Comparative and Phylogenetic Analysis. Molecules 2018, 23, 857. [Google Scholar] [CrossRef]
- Nagalievska, M.; Hachkova, H.; Sybirna, N. Galega officinalis, L. and Immunological Status in Diabetes Mellitus. Metformin 2020, 5, 75. [Google Scholar]
- Patel, D.K.; Kumar, R.; Laloo, D.; Hemalatha, S. Natural Medicines from Plant Source Used for Therapy of Diabetes Mellitus: An Overview of Its Pharmacological Aspects. Asian Pac. J. Trop. Dis. 2012, 2, 239–250. [Google Scholar] [CrossRef]
- Zaharudin, N.; Staerk, D.; Dragsted, L.O. Inhibition of α-Glucosidase Activity by Selected Edible Seaweeds and Fucoxanthin. Food Chem. 2019, 270, 481–486. [Google Scholar] [CrossRef]
- Uvarani, C.; Jaivel, N.; Sankaran, M.; Chandraprakash, K.; Ata, A.; Mohan, P.S. Axially Chiral Biscarbazoles and Biological Evaluation of the Constituents from Murraya Koenigii. Fitoterapia 2014, 94, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Ul Hassan, S.S.; Muhammad, I.; Abbas, S.Q.; Hassan, M.; Majid, M.; Jin, H.Z.; Bungau, S. Stress Driven Discovery of Natural Products from Actinobacteria with Anti-Oxidant and Cytotoxic Activities Including Docking and Admet Properties. Int. J. Mol. Sci. 2021, 22, 11432. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B. Roles of Alkaloids from Medicinal Plants in the Management of Diabetes Mellitus. J. Chem. 2021, 2021, 2691525. [Google Scholar] [CrossRef]
- Gao, H.; Huang, Y.N.; Gao, B.; Li, P.; Inagaki, C.; Kawabata, J. Inhibitory Effect on α-Glucosidase by Adhatoda Vasica Nees. Food Chem. 2008, 108, 965–972. [Google Scholar] [CrossRef]
- Oates, P. Aldose Reductase, Still a Compelling Target for Diabetic Neuropathy. Curr. Drug. Targets 2008, 9, 14–36. [Google Scholar] [CrossRef]
- Singh Grewal, A.; Bhardwaj, S.; Pandita, D.; Lather, V.; Singh Sekhon, B. Updates on Aldose Reductase Inhibitors for Management of Diabetic Complications and Non-Diabetic Diseases. Mini-Rev. Med. Chem. 2015, 16, 120–162. [Google Scholar] [CrossRef]
- Jung, H.A.; Yoon, N.Y.; Bae, H.J.; Min, B.S.; Choi, J.S. Inhibitory Activities of the Alkaloids from Coptidis Rhizoma against Aldose Reductase. Arch. Pharm. Res. 2008, 31, 1405–1412. [Google Scholar] [CrossRef]
- Patel, M.B.; Mishra, S. Isoquinoline Alkaloids from Tinospora Cordifolia Inhibit Rat Lens Aldose Reductase. Phytother. Res. 2012, 26, 1342–1347. [Google Scholar] [CrossRef]
- Sasaki, T.; Li, W.; Higai, K.; Koike, K. Canthinone Alkaloids Are Novel Protein Tyrosine Phosphatase 1B Inhibitors. Bioorg. Med. Chem. Lett. 2015, 25, 1979–1981. [Google Scholar] [CrossRef]
- Tang, D.; Chen, Q.-B.; Xin, X.L.; Aisa, H.A. Anti-Diabetic Effect of Three New Norditerpenoid Alkaloids in Vitro and Potential Mechanism via PI3K/Akt Signaling Pathway. Biomed. Pharmacother. 2017, 87, 145–152. [Google Scholar] [CrossRef]
- Ruud, J.; Steculorum, S.M.; Bruning, J.C. Neuronal Control of Peripheral Insulin Sensitivity and Glucose Metabolism. Nat. Commun. 2017, 8, 1–2. [Google Scholar] [CrossRef]
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the Two Incretin Hormones: Similarities and Differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 80. [Google Scholar] [CrossRef]
- Al-Masri, I.M.; Mohammad, M.K.; Tahaa, M.O. Inhibition of Dipeptidyl Peptidase IV (DPP IV) Is One of the Mechanisms Explaining the Hypoglycemic Effect of Berberine. J. Enzym. Inhib. Med. Chem. 2009, 24, 1061–1066. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced Glycation End Products and Diabetic Complications. Korean J. Physiol. Pharmacol. 2014, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Wen, W.; Qi, C.L.; Zhao, R.X.; Lü, J.H.; Zhong, C.Y.; Chen, Y.Y. Ameliorative Effect of Berberine on Renal Damage in Rats with Diabetes Induced by High-Fat Diet and Streptozotocin. Phytomedicine 2012, 19, 712–718. [Google Scholar] [CrossRef]
- Tiong, S.H.; Looi, C.Y.; Hazni, H.; Arya, A.; Paydar, M.; Wong, W.F.; Cheah, S.C.; Mustafa, M.R.; Awang, K. Antidiabetic and Antioxidant Properties of Alkaloids from Catharanthus Roseus (L.) G. Don. Molecules 2013, 18, 9770–9784. [Google Scholar] [CrossRef]
- Morita, M.; Yano, S.; Yamaguchi, T.; Sugimoto, T. Advanced Glycation End Products-Induced Reactive Oxygen Species Generation Is Partly through NF-Kappa B Activation in Human Aortic Endothelial Cells. J. Diabetes Complicat. 2013, 27, 11–15. [Google Scholar] [CrossRef]
- Salehi, B.; Ata, A.; Kumar, N.V.A.; Sharopov, F.; Ramírez-Alarcón, K.; Ruiz-Ortega, A.; Ayatollahi, S.A.; Fokou, P.V.T.; Kobarfard, F.; Zakaria, Z.A.; et al. Antidiabetic Potential of Medicinal Plants and Their Active Components. Biomolecules 2019, 9, 551. [Google Scholar] [CrossRef]
- Mastinu, A.; Bonini, S.A.; Premoli, M.; Maccarinelli, G.; Mac Sweeney, E.; Zhang, L.; Lucini, L.; Memo, M. Protective Effects of Gynostemma Pentaphyllum (Var. Ginpent) against Lipopolysaccharide-Induced Inflammation and Motor Alteration in Mice. Molecules 2021, 26, 570. [Google Scholar] [CrossRef]
- El-Nashar, H.A.S.; Mostafa, N.M.; El-Shazly, M.; Eldahshan, O.A. The Role of Plant-Derived Compounds in Managing Diabetes Mellitus: A Review of Literature from 2014 To 2019. Curr. Med. Chem. 2020, 28, 4694–4730. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytoconstituent (Alkaloids) Source | Chemical Structure | Mechanism of action |
|---|---|---|
| Avarol Dysidea avara |  | Inhibition of α-glucosidase enzyme can help in delaying digestion of carbohydrates, thereby reducing the levels of glucose in blood [49]. IC50 value for various avarol derivatives: 0.05–0.12 mM |
| Berberine Berberis spp. Tinospora cordifolia |  | Berberine is known as an AMP-activated protein kinase (AMPK) activator. Its insulin-independent hypoglycemic effect is related to inhibition of mitochondrial function, stimulation of glycolysis and activation of AMPK pathway, which inhibits alpha-glucosidase [25]. IC50 value: 0.68 μM |
| Casuarine 6-o-a-glucoside Syzygium malaccense |  | It acts by stimulating insulin secretion, inhibiting intestinal α-amylase activity, and increasing muscle basal glucose uptake along with antioxidant activity [60]. IC50 value of casuarine compounds: 9.7 μM |
| Catharanthine, Vindoline, and Vindolinine Catharanthus roseus |  Catharanthine  Vindoline  Vindolinine | Vindoline exhibits an insulinotropic effect by enhancing glucose-stimulated insulin secretion (GSIS). It was also found to increase plasma insulin in STZ-induced diabetic rats. In a recent study, vindoline reduced the voltage-dependent outward potassium currents through Kv2.1 inhibition. The combined effects resulted in fasting plasma glucose, improved oral glucose tolerance, and lowered serum glycated hemoglobin (HbA1c) and triglyceride (TG) levels [26]. IC 50 values: 59.6 μM, >30 µM, and >50 μg/mL, respectively. |
| Calystegine B2 Nicandra physalodes |  | In an in vitro study, calystegine B2 inhibited mainly sucrose activity by β-glucosidase alpha inhibitor and intestinal glucose absorption [30]. IC50 value: range 4.6 µM |
| Cryptolepine Cryptolepis sanguinolenta |  | Exhibits similar effects as of glibenclamide by inhibiting the ATP-sensitive potassium channels inhibitory regulatory subunit sulfonylurea receptor 1 and by activating AMP-activated protein kinase [33]. IC50 value of cryptoleptine derivatives: 27–41 μM |
| Harmane, Norharmane, Pinoline Tribulus terrestris |   Ponoline | Stimulatory action on insulin secretion by the activation of imidazoline-I binding sites in the pancreatic cell [36]. IC50 values: 5 μM; 51–58 μM and 0.11 μM |
| Jambosine Syzygium cumini |  | Reduces free radicals, improves the functioning of beta-pancreatic cells, and upregulates the PPARγ and PPARα [36]. IC50 value: 2.5 nM |
| Jatrorrhizine, Magnoflorine, Palmatine, Tembetarine Tinospora cordifolia |  Jatrorrhizine  Magnoflorine  Palmitine  Tembetarine | Lowering of blood glucose, increase in insulin sensitivity, inhibition of a-amylase and a-glucosidase activities, direct effect on carbohydrate metabolism [39,40]. IC50 value (derivatives): ~1.05 μM |
| Javaberine A, Javaberine B Talinum paniculatum |  Javaberine A  Javaberine B | Very limited data is available for these molecules; however, inhibitory activity in an in-vitro setting was noted by inhibition of TNF-α [39,40]. IC50 value: 23.5 µg/mL |
| Lepidine and semilepidine Lepidium sativum |  Lepidine  Semilepidine | Reduction in oxidative damage and modulation of antioxidant enzymes, potentiation of pancreatic secretion of insulin from the remaining islet β cells [42]. IC50 value: 1.42 ± 0.04 mg/mL |
| Mahanimbine Murraya koenigii |  | Inhibits alpha-amylase and alpha-glucosidase [44]. IC50 value: ranges from 3.5 to 64 μM |
| Piperumbellactam A Piper umbellatum |  | Inhibits α -glucosidase, maltase-glucoamylase, aldose reductase, and aldehyde reductase [45]. IC50 value of derivatives: Range 29.64 ± 0.46–98.07 ± 0.44 |
| Swerchirin Swertia chirayita |  | Lowers blood glucose level by stimulating insulin release from islets of Langerhans [51]. IC50 value: 20 μM |
| Radicamine A, B Lobelia Chinensis Lour |   | Suppressive activity against a glucosidase [46]. IC50 value: 54.6 µg/mL |
| Schulzeines A, B, and C Penares Schulzei |    | Inhibitors of glucosidase activity [48]. IC50 value range: 48−170 nM |
| Trigonelline Trigonella foenum-graecum |  | Glucose transport, carbohydrate digestion, and absorption [53]. IC50 value: 233 ± 0.12 µM |
| Tecomine Tecostanine Tecoma stans |  Tecomine  Tecostanine | Boosting effect on glucose absorption [57]. IC50 value: Not available |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behl, T.; Gupta, A.; Albratty, M.; Najmi, A.; Meraya, A.M.; Alhazmi, H.A.; Anwer, M.K.; Bhatia, S.; Bungau, S.G. Alkaloidal Phytoconstituents for Diabetes Management: Exploring the Unrevealed Potential. Molecules 2022, 27, 5851. https://doi.org/10.3390/molecules27185851
Behl T, Gupta A, Albratty M, Najmi A, Meraya AM, Alhazmi HA, Anwer MK, Bhatia S, Bungau SG. Alkaloidal Phytoconstituents for Diabetes Management: Exploring the Unrevealed Potential. Molecules. 2022; 27(18):5851. https://doi.org/10.3390/molecules27185851
Chicago/Turabian StyleBehl, Tapan, Amit Gupta, Mohammed Albratty, Asim Najmi, Abdulkarim M. Meraya, Hassan A. Alhazmi, Md. Khalid Anwer, Saurabh Bhatia, and Simona Gabriela Bungau. 2022. "Alkaloidal Phytoconstituents for Diabetes Management: Exploring the Unrevealed Potential" Molecules 27, no. 18: 5851. https://doi.org/10.3390/molecules27185851