A Non-Targeted Capillary Electrophoresis-Mass Spectrometry Strategy to Study Metabolic Differences in an In Vitro Model of High-Glucose Induced Changes in Human Proximal Tubular HK-2 Cells

 , ,
, ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Optimization of the Sample Treatment and Analysis by CE-MS
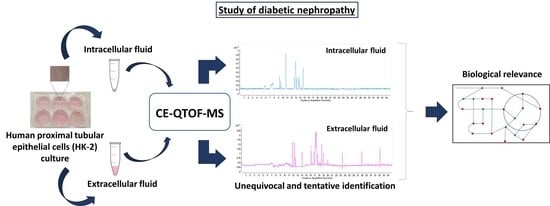
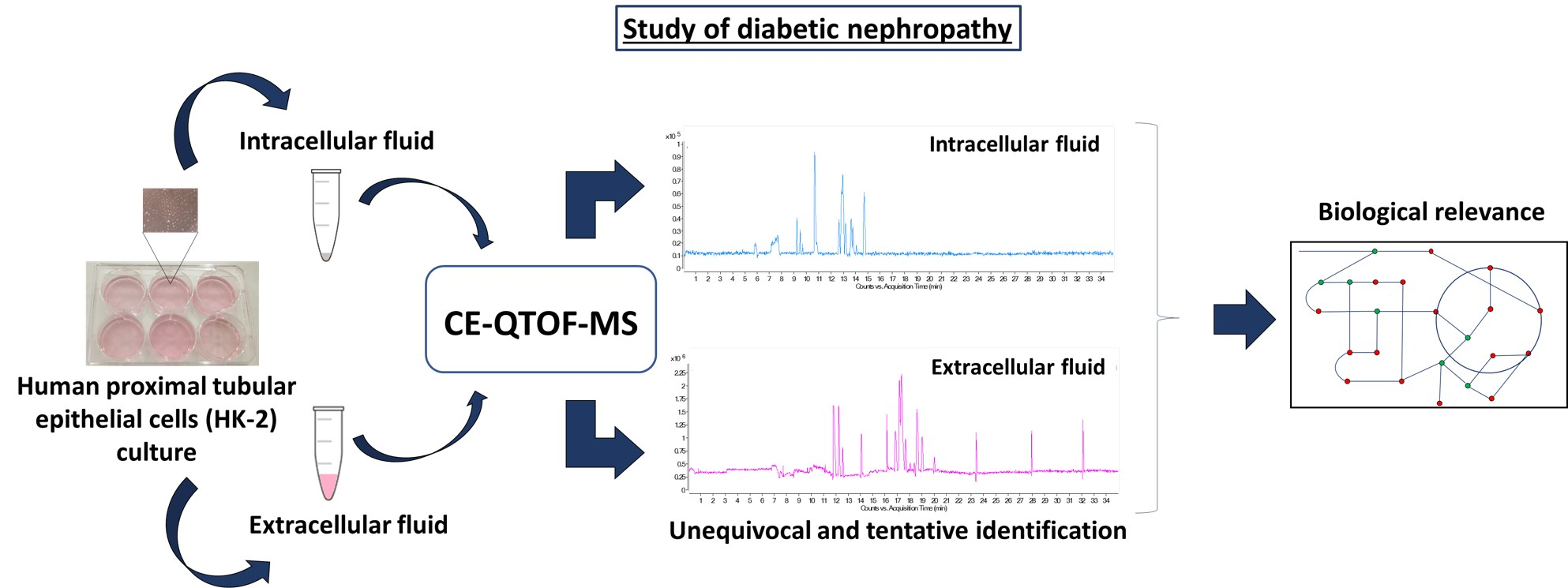
2.2. Non-Targeted Metabolomics Analysis of An in Vitro Model of High Glucose in HK-2 Cells
2.3. Identification of Affected Metabolites
2.4. Biological Interpretation of the Results
3. Materials and Methods
3.1. Reagents and Solvents
3.2. HK-2 Cells Culture
3.3. Metabolite Extraction from HK-2 Cells and Culture Media
3.4. Capillary Electrophoresis–Mass Spectrometry Analysis
3.5. Metabolomic Sequence
3.6. Data Treatment and Analysis
3.7. Identification of Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes atlas: Global estimates of diabetes prevalence for 2017 and for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2012, 35, S64–S71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, M.A.; Tak Park, J.; Natarajan, R. Epigenetic modifications in the pathogenesis of diabetic nephropathy. Semin. Nephrol. 2013, 33, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Lim, A.K.H. Diabetic nephropathy—Complications and treatment. Int. J. Nephrol. Renovasc. Dis. 2014, 7, 361–381. [Google Scholar] [CrossRef] [Green Version]
- León, Z.; García-Cañaveras, J.C.; Donato, M.T.; Lahoz, A. Mammalian cell metabolomics: Experimental design and sample preparation. Electrophoresis 2013, 34, 2762–2775. [Google Scholar] [CrossRef]
- Gilbert, R.E. Proximal tubulopathy: Prime mover and key therapeutic in diabetic kidney disease. Diabetes 2017, 66, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Bernardo-Bermejo, S.; Sánchez-López, E.; Castro-Puyana, M.; Benito, S.; Lucio-Cazaña, F.J.; Marina, M.L. An untargeted metabolomic strategy based on liquid chromatography- mass spectrometry to study high glucose-induced changes in HK-2 cells. J. Chromatogr. A 2019, 1596, 124–133. [Google Scholar] [CrossRef]
- So, E.J.; Kim, H.J.; Kim, C.W. Proteomic analysis of human proximal tubular cells exposed to high glucose concentrations. Proteomics Clin. Appl. 2008, 2, 1118–1126. [Google Scholar] [CrossRef]
- Ramautar, R.; Somsen, G.W.; de Jong, G.J. CE-MS for metabolomics: Developments and applications in the period 2016–2018. Electrophoresis 2019, 40, 165–179. [Google Scholar] [CrossRef] [Green Version]
- González-Riano, C.; Dudzik, D.; Garcia, A.; Gil-de-la-Fuente, A.; Gradillas, A.; Godzien, J.; López-Gonzálvez, Á.; Rey-Stolle, F.; Rojo, D.; Ruperez, F.J.; et al. Recent developments along the analytical process for metabolomics workflows. Anal. Chem. 2019, 1, 203–216. [Google Scholar] [CrossRef]
- Begou, O.; Gika, H.G.; Wilson, I.D.; Theodoridis, G. Hyphenated MS-based targeted approaches in metabolomics. Analyst 2017, 142, 3079–3100. [Google Scholar] [CrossRef]
- Wei, P.Z.; Fung, W.W.; Ng, J.K.; Lai, K.; Luk, C.C.; Chow, K.M.; Li, P.K.; Szeto, C.C. Metabolomic Changes of Human Proximal Tubular Cell Line in High Glucose Environment. Sci. Rep. 2019, 9, 16617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soga, T.; Ohashi, Y.; Ueno, Y.; Naraoka, H.; Tomita, M.; Nishioka, T. Quantitative metabolome analysis using capillary electrophoresis mass spectrometry. J. Proteome. Res. 2003, 2, 488–499. [Google Scholar] [CrossRef]
- Soga, T.; Ueno, Y.; Naraoka, H.; Ohashi, Y.; Tomita, M.; Nishioka, T. Simultaneous determination of anionic intermediates for Bacillus subtilis metabolomic pathways by capillary electrophoresis electrospray ionization mass spectrometry. Anal. Chem. 2002, 74, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Heiger, D.N. Amino acid analysis by capillary electrophoresis electrospray ionization mass spectrometry. Anal. Chem. 2000, 72, 1236–1241. [Google Scholar] [CrossRef]
- Kimura, T.; Hamase, K.; Miyoshi, Y.; Miyoshi, Y.; Yamamoto, R.; Yasuda, K.; Mita, M.; Rakugi, H.; Hayashi, T.; Isaka, Y. Chiral amino acid metabolomics for novel biomarker screening in the prognosis of chronic kidney disease. Sci Rep. 2016, 6, 26137. [Google Scholar] [CrossRef] [PubMed]
- Kurosaki, Y.; Imoto, A.; Kawakami, F.; Yokoba, M.; Takenaka, T.; Ichikawa, T.; Katagiri, M.; Ishii, N. Oxidative stress increases megalin expression in the renal proximal tubules during the normoalbuminuric stage of diabetes mellitus. Am. J. Physiol. Renal Physiol. 2018, 314, F462–F470. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M., Jr.; Gao, T.; Cooper, T.K.; Kepka-Lenhart, D.; Awad, A.S. Arginase-2 mediates diabetic renal injury. Diabetes 2011, 60, 3015–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, W.; Cao, R.; He, Y.C.; Guan, Y.F.; Ruan, X.Z. Crosstalk of Hyperglycemia and Dyslipidemia in Diabetic Kidney Disease. Kidney Dis (Basel). 2017, 3, 171–180. [Google Scholar] [CrossRef]
- Kuehnbaum, N.L.; Gillen, J.B.; Kormendi, A.; Lam, K.P.; DiBattista, A.; Gibala, M.J.; Britz-McKibbin, P. Multiplexed separations for biomarker discovery in metabolomics: Elucidating adaptive responses to exercise training. Electophoresis 2015, 36, 2226–2236. [Google Scholar] [CrossRef]
- Britz McKibbin, P. Capillary electrophoresis electrospray ionization-mass spectrometry (CE-ESI-MS)-based metabolomics. Methods. Mol. Biol. 2011, 708, 229–246. [Google Scholar] [PubMed]
- Baeuml, F.; Welsch, T. Improvement of the long-term stability of polyimide-coated fused-silica capillaries used in capillary electrophoresis and capillary electrochromatography. J. Chromatogr. A. 2002, 96, 35–44. [Google Scholar] [CrossRef]
- Wan, H.; Holmén, A.; Någård, M.; Lindberg, W. Rapid screening of pK (a) values of pharmaceuticals by pressure-assisted capillary electrophoresis combined with short-end injection. J. Chromatogr. A. 2002, 979, 369–377. [Google Scholar] [CrossRef]
- Kohler, I.; Verhoeven, A.; Derks, R.J.; Giera, M. Analytical pitfalls and challenges in clinical metabolomics. Bioanalysis 2016, 8, 1509–1532. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.T.; Cieslarová, Z.; Tavares, M.F.; Simionato, A.V. Strategies involving mass spectrometry combined with capillary electrophoresis in metabolomics. Adv. Exp. Med. Biol. 2017, 965, 99–141. [Google Scholar] [PubMed]
- Bonvin, G.; Schappler, J.; Rudaz, S. Capillary electrophoresis-electrospray ionization-mass spectrometry interfaces: Fundamental concepts and technical developments. J. Chromatogr. A. 2012, 1267, 17–31. [Google Scholar] [CrossRef]
- Nilsson, S.L.; Bylund, D.; Jornten-Karlsson, M.; Peteresson, P.; Markides, K.E. A chemometric study of active parameters and their interaction effects in a nebulized sheath-liquid electrospray interface for capillary electrophoresis-mass spectrometry. Electrophoresis 2004, 25, 2100–2107. [Google Scholar] [CrossRef]
- Ramautar, R. Capillary electrophoresis-mass spectrometry for clinical metabolomics. Adv. Clin. Chem. 2016, 74, 1–34. [Google Scholar]
- Mokaddem, M.; Gareil, P.; Belgaied, J.E.; Varenne, A. A new insight into suction and dilution effects in capillary electrophoresis coupled to mass spectrometry via an electrospray ionization interface. Part I-Suction effect. Electrophoresis 2008, 29, 1957–1964. [Google Scholar] [CrossRef]
- Duan, Q.; Cao, J.; Zhang, J. Analysis of phenolic acids and their antioxidant activity by capillary electrophoresis-mass spectrometry with field-amplified sample injection. Anal. Methods 2012, 4, 3027–3032. [Google Scholar] [CrossRef]
- Aluksanasuwan, S.; Khamchun, S.; Thongboonkerd, V. Targeted functional investigations guided by integrative proteome network analysis revealed significant perturbations of renal tubular cell functions induced by high-glucose. Proteomics 2017, 17, 1700151. [Google Scholar] [CrossRef] [PubMed]
- Ramautar, R.; Berger, R.; van der Greef, J.; Hankemeier, T. Human metabolomics: Strategies to understand biology. Curr. Opin. Chem. Biol. 2013, 17, 841–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armitage, E.G.; Godzien, J.; Peña, I.; López-Gonzálvez, A.; Angulo, S.; Gradillas, A.; Alonso-Herranz, V.; Martín, J.; Fiandor, M.J.; Barret, M.P.; et al. Clustering analysis as a strategy for compound selection in the drug discovery pipeline for leishmaniasis. ACS Chem. Biol. 2018, 13, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Sun, K.; Lan, Z.; Song, W.; Cheng, L.; Chi, W.; Chen, J.; Huo, Y.; Xu, L.; Liu, X.; et al. Tenofovir and adefovir down-regulate mitochondrial chaperone TRAP1 and succinate dehydrogenase subunit B to metabolically reprogram glucose metabolism and induce nephrotoxicity. Sci. Rep. 2017, 7, 46344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Ge, M.; Lan, Z.; Ma, Z.; Chi, W.; Kuang, W.; Sun, K.; Zhao, X.; Liu, Y.; Feng, Y.; et al. Zoledronate dysregulates fatty acid metabolism in renal tubular epithelial cells to induce nephrotoxicity. Arch. Toxicol. 2018, 92, 469–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hau, J.; Devaud, S.; Blank, I. Detection of Amadori compounds by capillary electrophoresis coupled to tandem mass spectrometry. Electrophoresis 2004, 25, 2077–2083. [Google Scholar] [CrossRef] [PubMed]
- Mather, A.; Pollock, C. Glucose handling by the kidney. Kidney Int. 2011, 79, S1–S6. [Google Scholar] [CrossRef] [Green Version]
- Dunn, W.B.; Broadhurst, D.I.; Atherton, H.J.; Goodacre, R.; Griffin, J.L. Systems level studies of mammalian metabolomes: The roles of mass spectrometry and nuclear magnetic resonance spectroscopy. Chem. Soc. Rev. 2011, 40, 387–426. [Google Scholar] [CrossRef]
- Morris, S.M. Recent advances in arginine metabolism roles and regulation of the arginases. Br. J. Pharmacol. 2009, 157, 922–930. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.J.; Lai, F.J.; Wen, J.D.; Hsiao, P.J.; Hsieh, M.C.; Tzeng, T.F.; Chen, H.C.; Guh, J.Y.; Tsai, J.H. Neuronal and endothelial nitric oxide synthase expression in outer medulla of streptozotocin-induced diabetic rat kidney. Diabetologia 2000, 43, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.M., Jr.; Bhamidipati, D.; Kepka-Lenhart, D. Human type II arginase: Sequence analysis and tissue-specific expression. Gene 1997, 193, 157–161. [Google Scholar] [CrossRef]
- Morgan, T.K.; Montgomery, K.; Mason, V.; Mason, V.; West, R.B.; Wang, L.; van de Rijn, M.; Higgins, J.P. Upregulation of histidine decarboxylase expression in superficial cortical nephrons during pregnancy in mice and women. Kidney Int. 2006, 70, 306–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pini, A.; Verta, R.; Grange, C.; Gurrieri, M.; Rosa, A.C. Histamine and diabetic nephropathy: An up-to-date overview. Clin. Sci. 2019, 133, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, D.; Chen, Y.; Sun, H.; He, S.; Wang, C.; Yang, G.; Shi, M.; Zhang, J.; Ren, Y.; et al. 1H NMR-based metabonomic analysis of serum and urine in a nonhuman primate model of diabetic nephropathy. Mol. Biosyst. 2013, 9, 2645–2652. [Google Scholar] [CrossRef]
- Salvucci, M.; Neufeld, Z.; Newsholme, P. Mathematical model of metabolism and electrophysiology of amino acid and glucose stimulated insulin secretion: In vitro validation using a β-cell line. PLoS ONE 2013, 8, e5261. [Google Scholar] [CrossRef] [Green Version]
- Stancáková, A.; Civelek, M.; Saleem, N.K.; Soininen, P.; Kangas, A.J.; Cederberg, H.; Paananen, J.; Pihlajamäki, J.; Bonnycastle, L.L.; Morken, M.A.; et al. Hyperglycemia and a common variant of GCKR are associated with the levels of eight amino acids in 9369 Finnish men. Diabetes 2012, 61, 1895–1902. [Google Scholar] [CrossRef] [Green Version]
- Curthoys, N.P.; Moe, O.W. Proximal tubule function and response to acidosis. Clin. J. Am. Soc. Nephrol. 2014, 9, 1627–1638. [Google Scholar] [CrossRef] [Green Version]
- Mircheff, A.K.; Kippen, I.; Hirayama, B.; Wright, E.M. Delineation of sodium-stimulated amino acid transport pathways in rabbit kidney brush border vesicles. J Membr Biol. 1982, 64, 113–122. [Google Scholar] [CrossRef]
- Bröer, S. Amino acid transport across mammalian intestinal and renal epithelia. Physiol Rev. 2008, 88, 249–286. [Google Scholar] [CrossRef]
- Zhang, H.; Qi, L.; Lin, Y.; Mao, L.; Chen, Y. Study on the decrease of renal d-amino acid oxidase activity in the rat after renal ischemia by chiral ligand exchange capillary electrophoresis. Amino Acids 2012, 42, 337–345. [Google Scholar] [CrossRef]
- Pastore, A.; Noce, A.; Di Giovamberardino, G.; De Stefano, A.; Callà, C.; Zenobi, R.; Dessì, M.; Di Daniele, N. Homocystine, cystine, folate and vitamin B12status in type2 diabetic patients with chronic kidney disease. J. Nephrol. 2015, 28, 571–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargrove, J.L.; Wichman, R.D. A cystine-dependent inactivator of tyrosine amino-transferase co-purifies with gamma-cystathionase (cystine desulfurase). J. Biol. Chem. 1987, 262, 7351–7357. [Google Scholar] [PubMed]
- Libert, D.M.; Nowacki, A.S.; Natowicz, M.R. Metabolomic analysis of obesity, metabolic syndrome, and type 2 diabetes: Amino acid and acylcarnitine levels change along a spectrum of metabolic wellness. PeerJ 2018, 6, e5410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pena, M.J.; Lambers Heerspink, H.J.; Hellemons, M.E.; Friedrich, T.; Dallmann, G.; Bakker, S.J.; Gansevoort, R.T.; Rossing, P.; de Zeeuw, D.; Roscioni, S.S. Urine and plasma metabolites predict the development of diabetic nephropathy in individuals with Type 2 diabetes mellitus. Diabet. Med. 2014, 31, 1138–1147. [Google Scholar] [CrossRef]
- Sánchez, C.F.; Peiró, C.; Rodríguez, L. Los productos de amadori como mediadores de disfunción endothelial en la diabetes mellitus. Endocrinol Nutr. 2004, 51, 497–505. [Google Scholar] [CrossRef]
- Nagamori, S.; Wiriyasermkul, P.; Guarch, M.E.; Okuyama, H.; Nakagomi, S.; Tadagaki, K.; Nishinaka, Y.; Bodoy, S.; Takafuji, K.; Okuda, S.; et al. Novel cystine transporter in renal proximal tubule identified as a missing partner of cystinuria-related plasma membrane protein rBAT/SLC3A1. Proc. Natl. Acad. Sci. USA 2016, 113, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Rawat, A.; Misra, G.; Saxena, M.; Tripathi, S.; Dubey, D.; Saxena, S.; Aggarwal, A.; Gupta, V.; Khan, M.Y.; Prakash, A. 1H NMR based serum metabolic profiling reveals differentiating biomarkers in patients with diabetes and diabetes-related complication. Diabetes Metab. Syndr. 2019, 13, 290–298. [Google Scholar] [CrossRef]
- Gil de la Fuente, A.; Godzien, J.; Fernández López, M.; Rupérez, F.J.; Barbas, C.; Otero, A. Knowledge-based metabolite annotation tool: CEU Mass Mediator. J. Pharm. Biomed. Anal. 2018, 154, 138–149. [Google Scholar] [CrossRef]
Sample Availability: Not available. |



{kind=link}
{kind=link}
{kind=link}
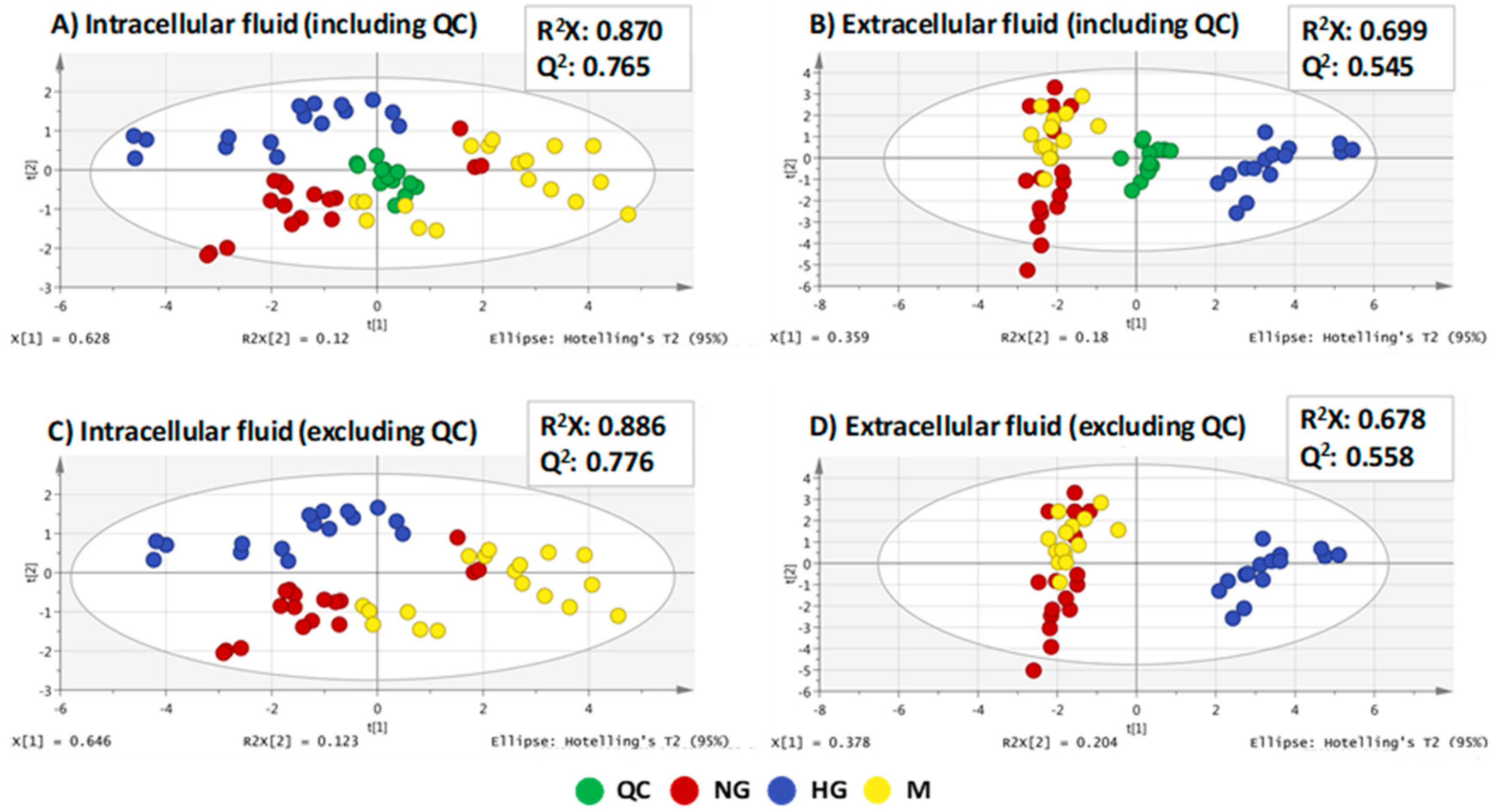
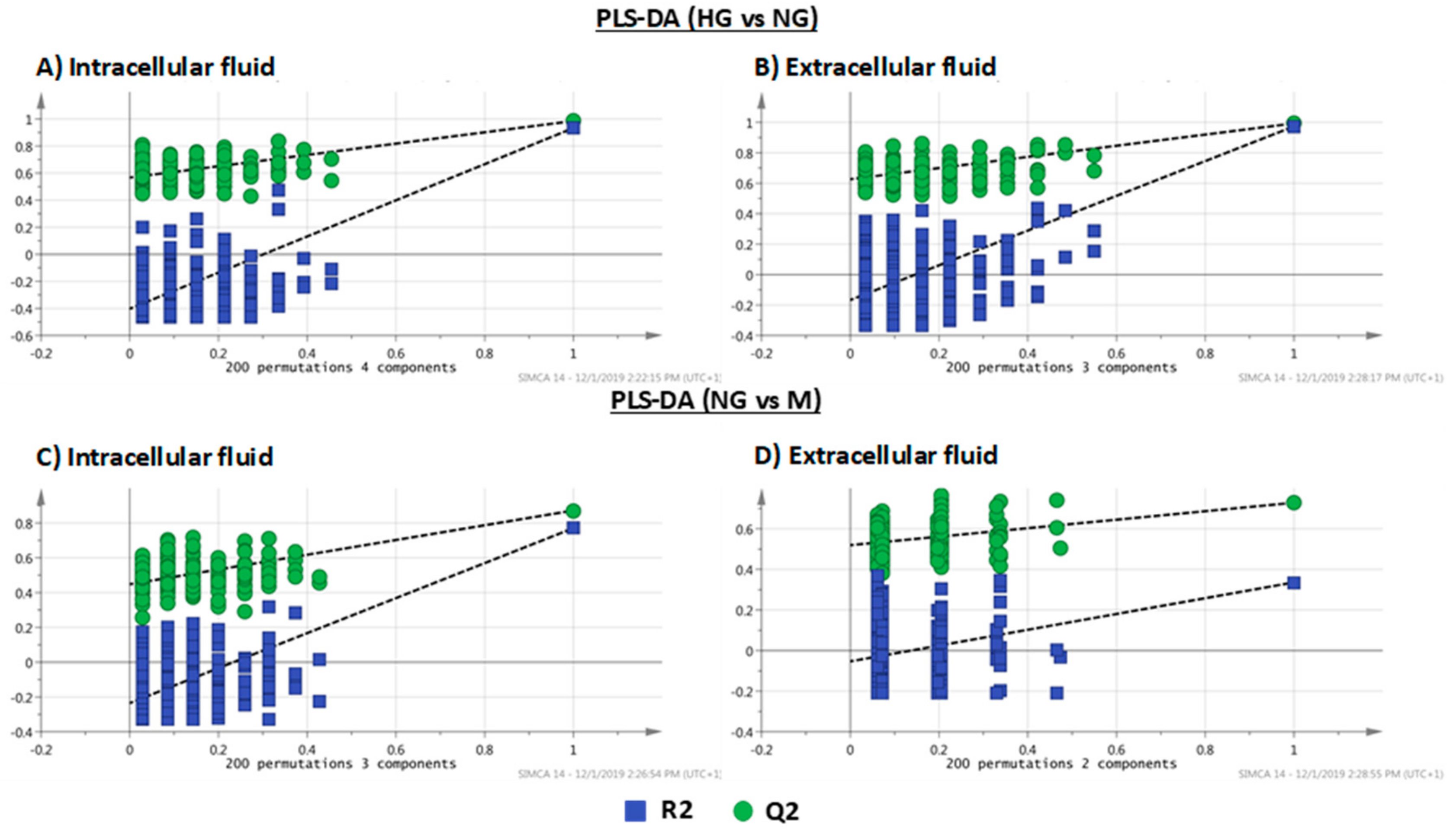
| PLS-DA Models | R2X | R2Y | Q2 | CV-ANOVA | |
|---|---|---|---|---|---|
| Intracellular Fluid | |||||
| HG vs. NG | 0.801 | 0.985 | 0.933 | F(22.5) | p(2.3 × 10−9) |
| NG vs. M | 0.827 | 0.872 | 0.771 | F(15.8) | p(7.7 × 10−8) |
| Extracellular Fluid | |||||
| HG vs. NG | 0.670 | 0.993 | 0.974 | F(149.2) | p(8.7 × 10−18) |
| NG vs. M | 0.457 | 0.728 | 0.336 | F(3.1) | p(3.3 × 10−2) |
| # | MT (min) | Molecular Formula | Identification | [M] Monoisotopic Mass (Da) | Mass Error (ppm) | Main Fragments (MS/MS) | p-Value ** | Trend *** | |
|---|---|---|---|---|---|---|---|---|---|
| HG vs. NG | NG vs. M | ||||||||
| Intracellular Fluid | |||||||||
| 1 | 9.7 | C6H14N4O2 | Arginine * | 174.1116 | 0.6 | 130.0975 ([M + H-NH3-CO]+) | 1.33 × 10−2 | 3.53 ×∙10−5 | ↓ |
| 116.0702 ([M + H-CH5N3]+ | |||||||||
| 112.0873 ([M + H-NH3-H2O-CO]+) | |||||||||
| 158.0950 ([M + H-NH3]+) | |||||||||
| 2 | 13.5 | C5H9NO4 | Glutamic acid * | 147.0531 | 0.7 | 84.0446 ([M + H-2H2O-CO]+) | 7.85 ×∙10−8 | 3.81 ×∙10−6 | ↓ |
| 56.0497 ([M + H-2H2O-2CO]+) | |||||||||
| 3 | 23.8 | C5H7NO3 | Pyroglutamic acid * | 129.0424 | 1.5 | 56.0493 ([M + H-H2O-2CO]+) | 5.49∙× 10−3 | 2.90∙× 10−3 | ↑ |
| 84.0443 ([M + H-H2O-CO] +) | |||||||||
| Extracellular Fluid | |||||||||
| 4 | 12.9 | C6H9N3O2 | Histidine * | 155.0696 | 0.6 | 110.0713 ([M + H-H2O-CO]+) | 1.20∙× 10−2 | 0.79 | ↑ |
| 83.0602 ([M + H-H2O-CO-HCN]+) | |||||||||
| 93.0444 ([M + H-H2O-CO-NH3]+) | |||||||||
| 5 | 15.8 | C3H7NO2 | Alanine * | 89.0476 | 0.9 | 58.0619 | 5.94∙× 10−7 | 0.71 | ↑ |
| 60.0784 ([M + H-CH2O]+) | |||||||||
| 74.0927 ([M + H-NH2]+) | |||||||||
| 60.9827 ([M + H-CHO]+) | |||||||||
| 72.0752 ([M + H-H2O]+) | |||||||||
| 6 | 19.0 | C5H9NO2 | Proline * | 115.0629 | 3.5 | 70.0638 ([M + H-H2O-CO]+) | 1.11∙× 10−4 | 0.25 | ↑ |
| 7 | 19.1 | C5H9NO4 | Glutamic acid * | 147.0529 | 2.0 | 84.0429 ([M + H-2H2O-CO]+) | 6.38 × 10−5 | 6.62∙× 10−2 | ↑ |
| 56.0480 ([M + H-2H2O-2CO]+) | |||||||||
| 8 | 19.3 | C6H12N2O4S2 | Cystine * | 240.0215 | 9.6 | 74.0216 ([M + H-C3H7NO2-CH2S2]]) | 1.00 ×∙10−3 | 0.29 | ↑ |
| 120.0109 ([M + H-C3H7NO2S]+) | |||||||||
| 122.0247 ([M + H-C3H5NO2S]+) | |||||||||
| 151.9848 ([M + H-C3H7NO2]+) | |||||||||
| 9 | 19.9 | C9H11NO3 | Tyrosine * | 181.0731 | 4.4 | 91.0501 ([M + H-NH3-H2O-2CO]+) | 2.28∙× 10−2 | 8.27∙× 10−4 | ↑ |
| 136.0711 ([M + H-H2O-CO]+) | |||||||||
| 119.0445 ([M + H-NH3-H2O-CO]+) | |||||||||
| 123.0398 ([M + H-NH3-CH2CO]+) | |||||||||
| 95.0450 ([M + H-NH3-CH2CO-CO]+) | |||||||||
| 10 | 24.6 | C12H23NO7 | N-(1-Deoxy-1-fructosyl)leucine or N-(1-Deoxy-1-fructosyl)isoleucine | 293.1472 | 1.0 | 230.1350 ([M + H-2H2O-CO]+) 258.1301 ([M + H-2H2O]+) | 8.87∙× 10−8 | 0.30 | ↑ |
| 276.1404 ([M + H-H2O]+) | |||||||||
| 11 | 26.3 | C15H21NO7 | N-(1-Deoxy-1-fructosyl)phenylalanine | 327.1316 | 0.6 | 310.1226 ([M + H-H2O]+) | 8.51∙× 10−6 | 0.28 | ↑ |
| 292.1189 ([M + H-2H2O]+) | |||||||||
| 264.1281 ([M + H-H2O-C2H5OH]+) | |||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardo-Bermejo, S.; Sánchez-López, E.; Castro-Puyana, M.; Benito-Martínez, S.; Lucio-Cazaña, F.J.; Marina, M.L. A Non-Targeted Capillary Electrophoresis-Mass Spectrometry Strategy to Study Metabolic Differences in an In Vitro Model of High-Glucose Induced Changes in Human Proximal Tubular HK-2 Cells. Molecules 2020, 25, 512. https://doi.org/10.3390/molecules25030512
Bernardo-Bermejo S, Sánchez-López E, Castro-Puyana M, Benito-Martínez S, Lucio-Cazaña FJ, Marina ML. A Non-Targeted Capillary Electrophoresis-Mass Spectrometry Strategy to Study Metabolic Differences in an In Vitro Model of High-Glucose Induced Changes in Human Proximal Tubular HK-2 Cells. Molecules. 2020; 25(3):512. https://doi.org/10.3390/molecules25030512
Chicago/Turabian StyleBernardo-Bermejo, Samuel, Elena Sánchez-López, María Castro-Puyana, Selma Benito-Martínez, Francisco Javier Lucio-Cazaña, and María Luisa Marina. 2020. "A Non-Targeted Capillary Electrophoresis-Mass Spectrometry Strategy to Study Metabolic Differences in an In Vitro Model of High-Glucose Induced Changes in Human Proximal Tubular HK-2 Cells" Molecules 25, no. 3: 512. https://doi.org/10.3390/molecules25030512