Expression of Pinellia pedatisecta Lectin Gene in Transgenic Wheat Enhances Resistance to Wheat Aphids


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
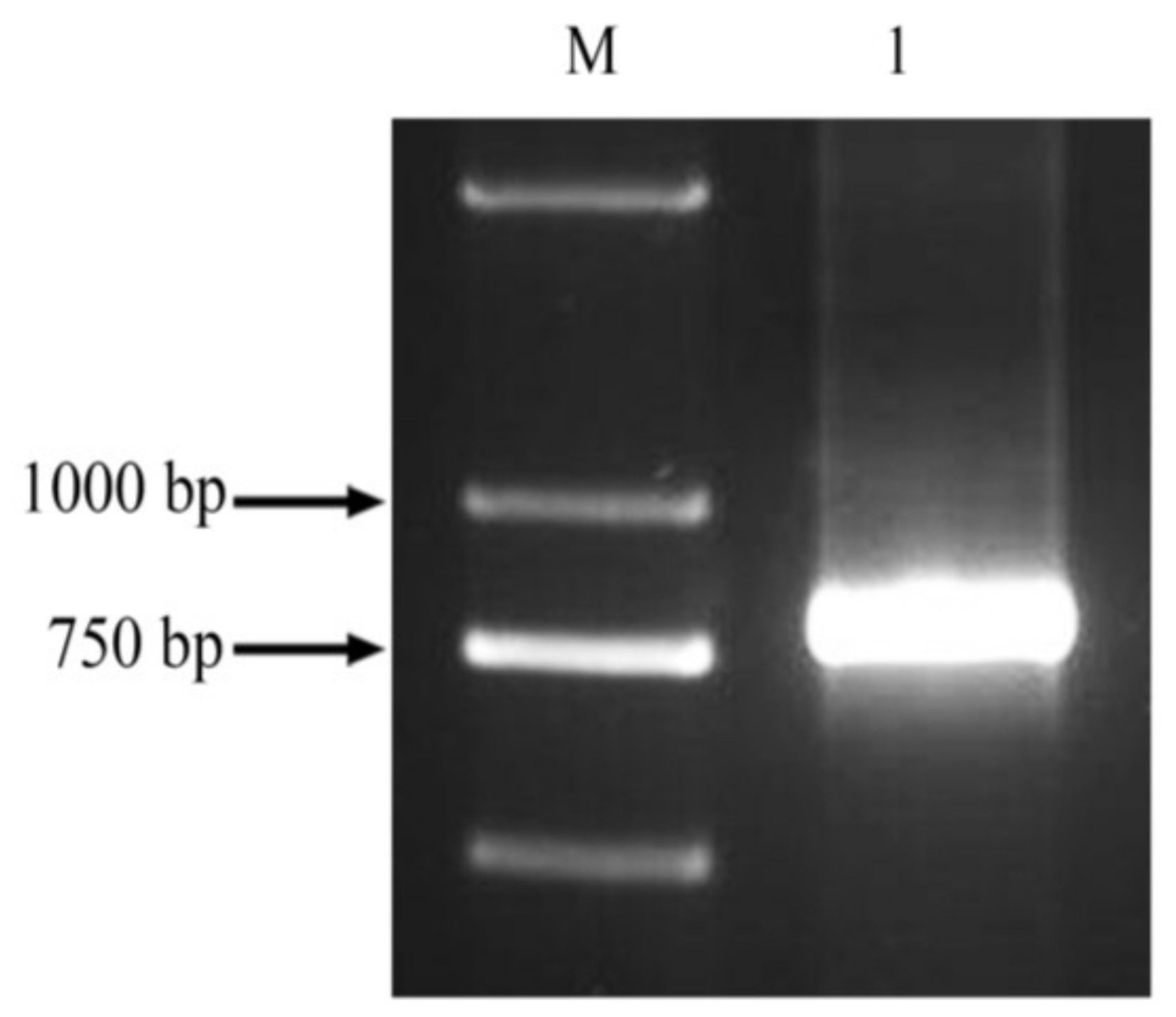
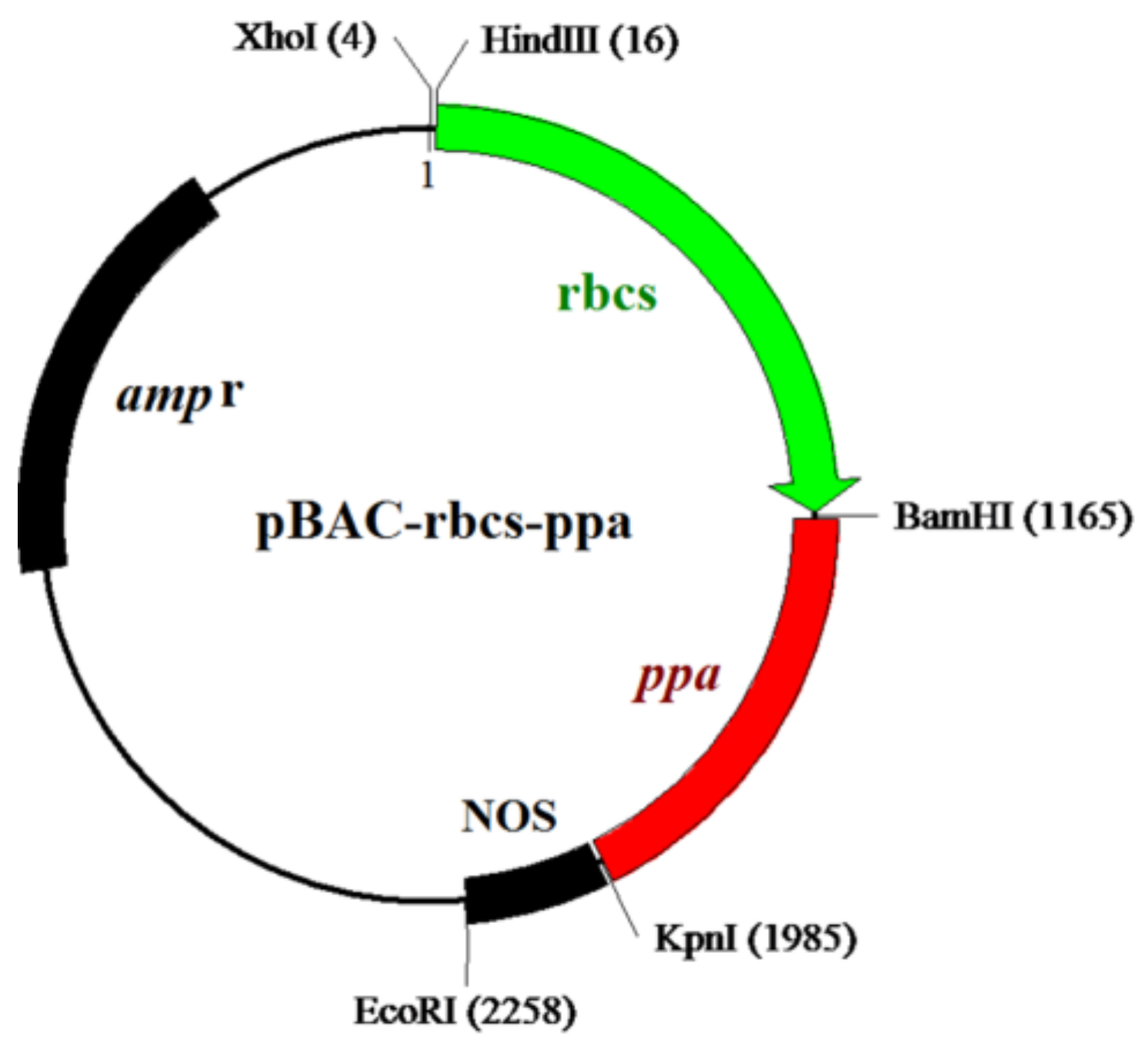
2.1. Cloning of the ppa Fragment and Construction of the Expression Vector
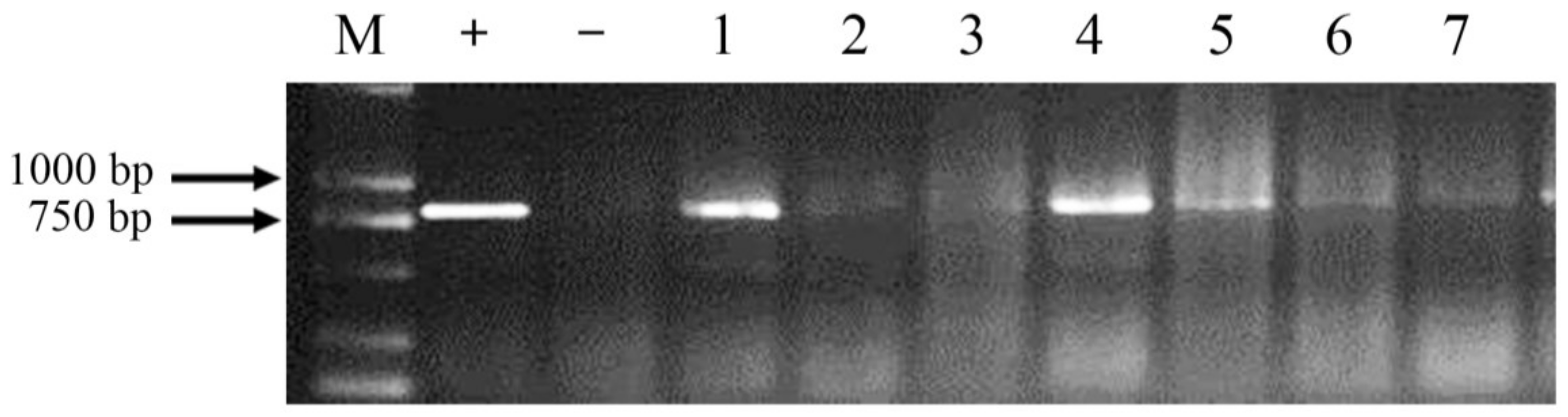
2.2. Production of Transgenic Lines and Expression of the ppa Gene
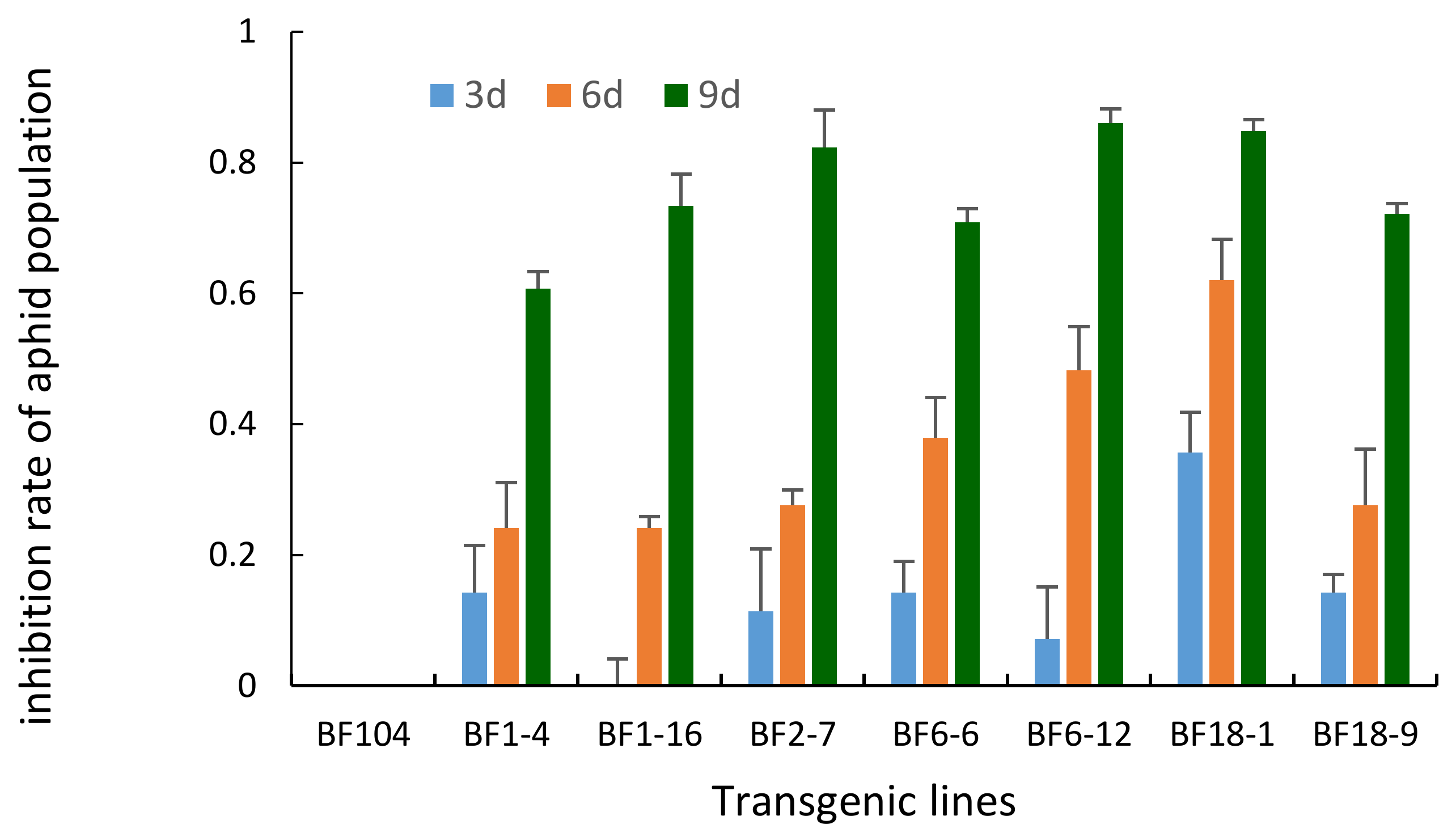
2.3. Aphid Bioassay on Detached Leaf Segments
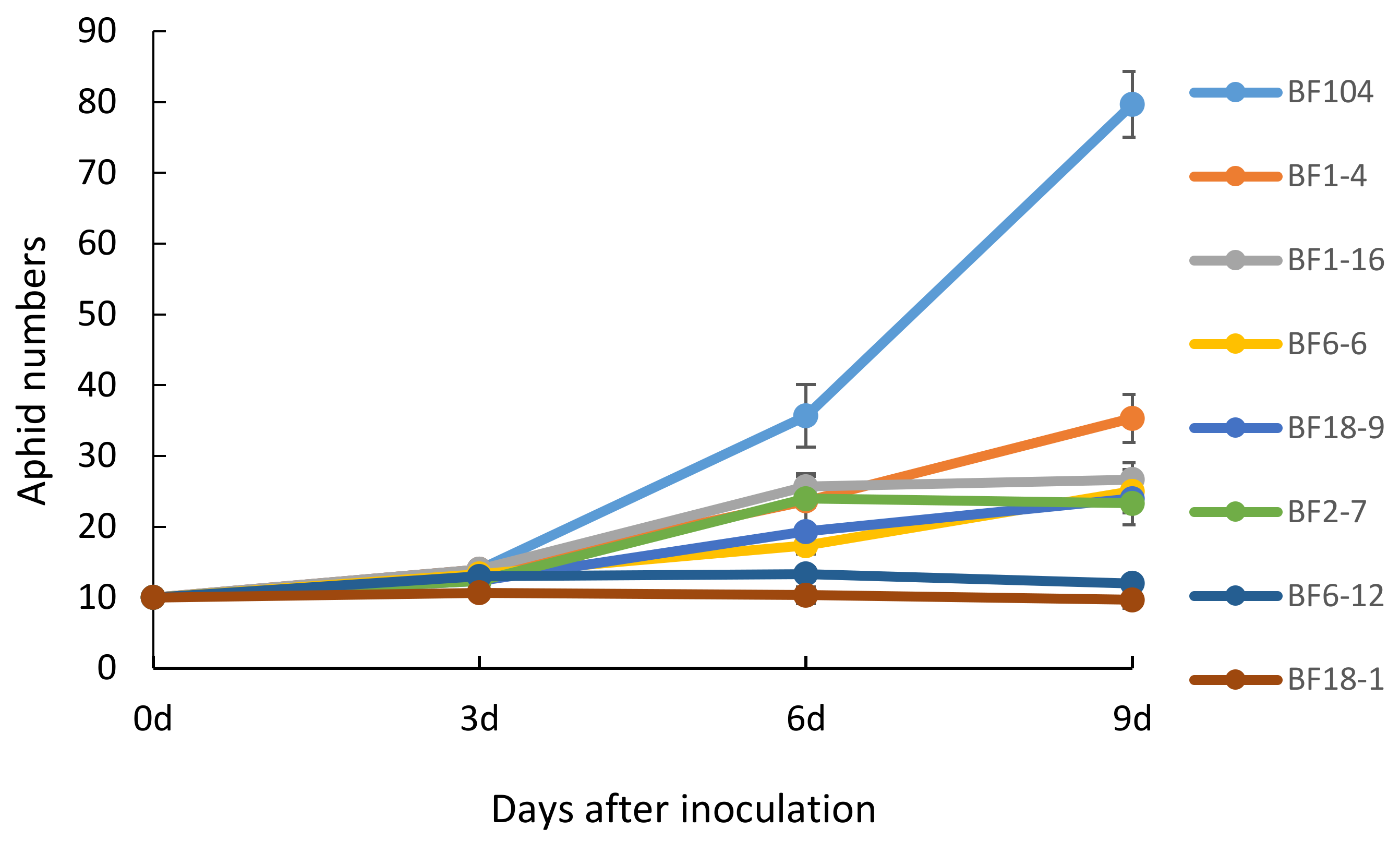
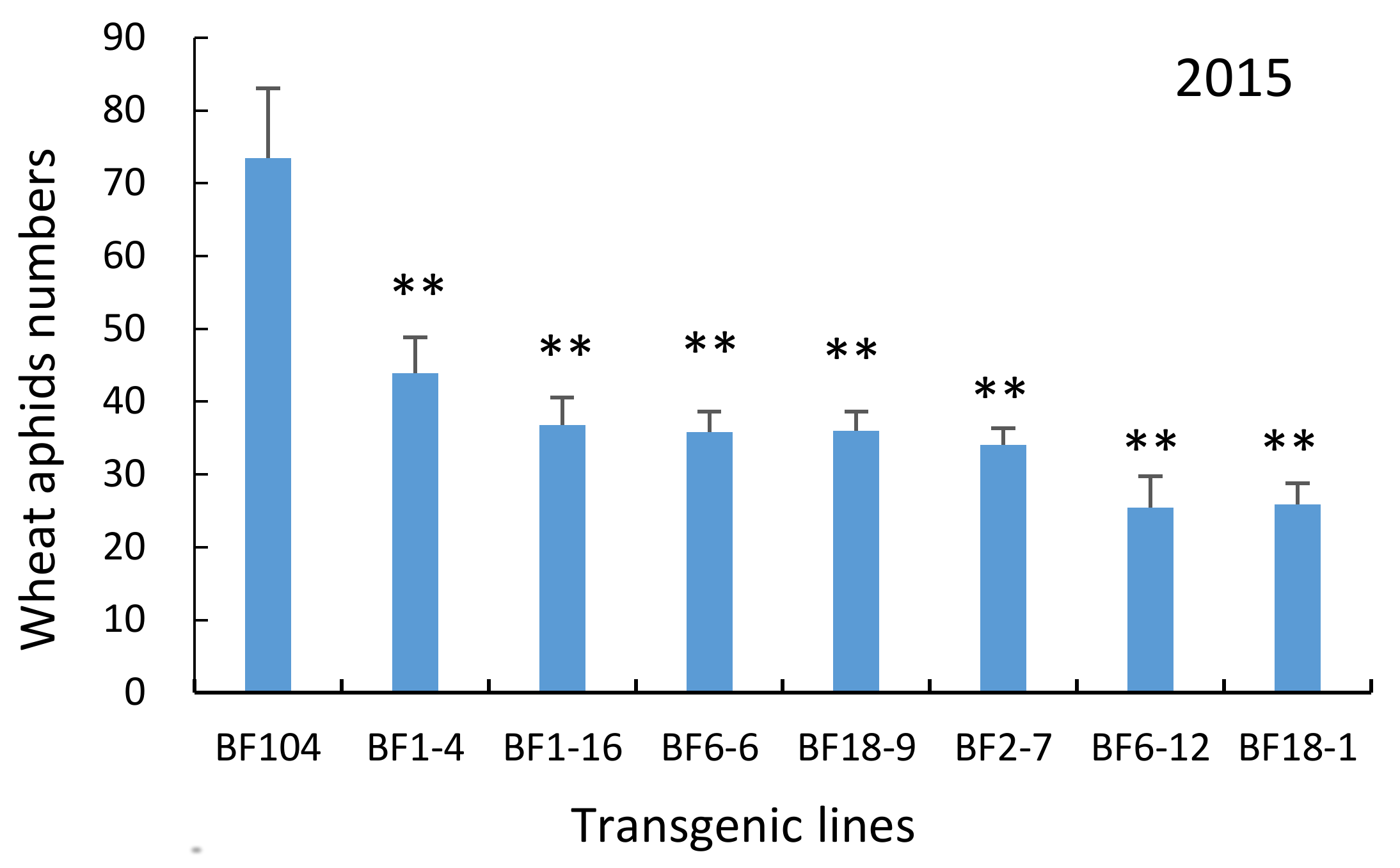
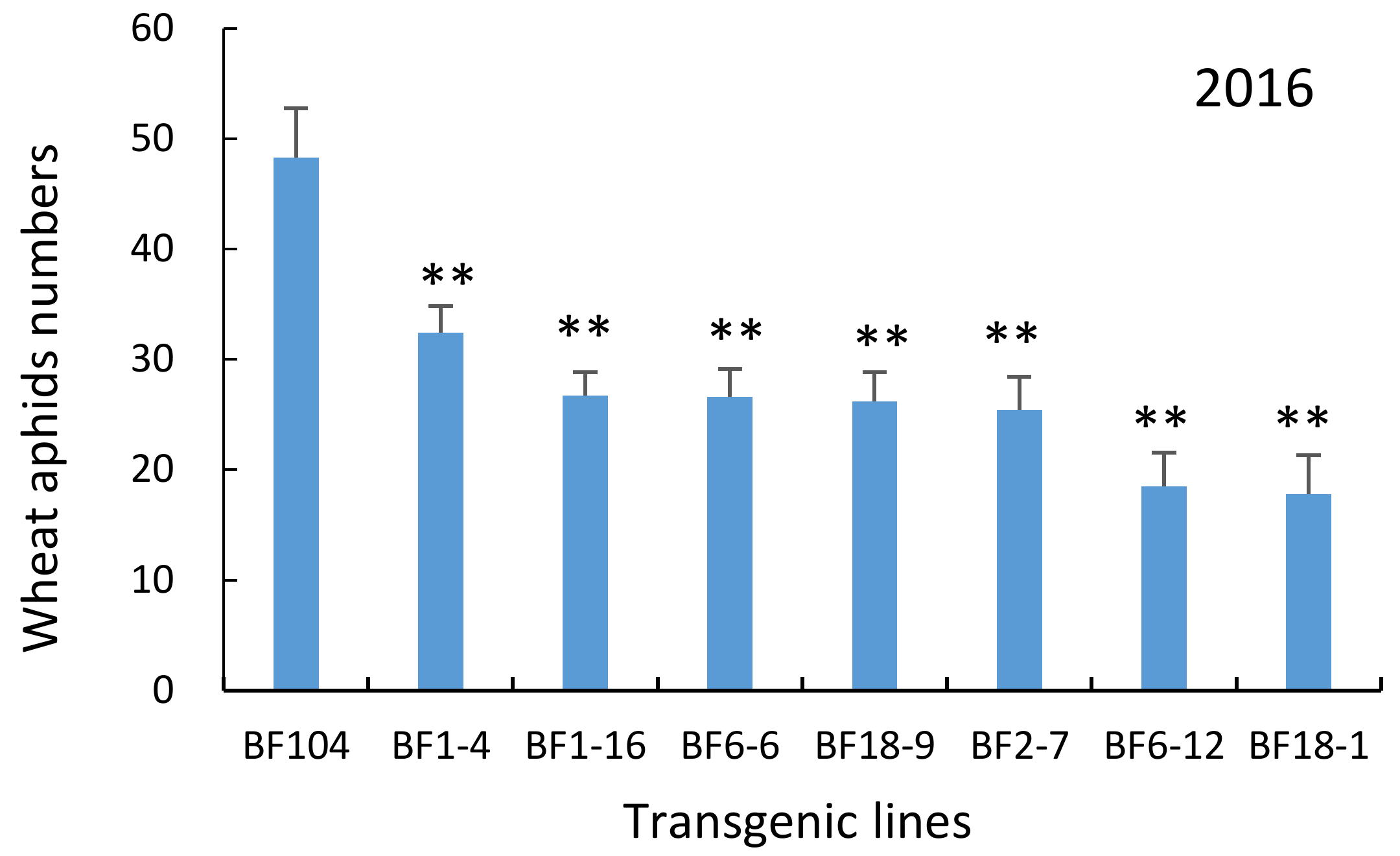
2.4. T4 and T5 Transgenic Wheat Lines in the Field Showed Lower Aphid Numbers
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Aphids
4.2. Gene Cloning and Sequencing of the ppa Gene Fragment
4.3. Construction of Expression Vector pBAC-rbcs-ppa and Production of Transgenic Wheat
4.4. PCR and Screening of Homozygous Transgenic Lines
4.5. Expression of ppa in Transgenic Wheat
4.6. Aphid Bioassay on Detached Leaf Segments
4.7. Aphid Resistance Bioassay under Field Conditions
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PPA | Pinellia pedatisecta agglutinin |
| PTA | Pinellia ternata agglutinin |
| GNA | Galanthus nivalis agglutinin |
| ORF | Open Reading Frame; |
| DAF | Days After Flowering; |
| PCR | Polymerase Chain Reaction |
| RT-PCR | Reverse Transcriptional Polymerase Chain Reaction |
References
- Feng, M.; Johnson, J.B.; Halbert, S.E. Parasitoids (Hymenoptera: Aphidiidae and Aphelinidae) and Their Effect on Aphid (Homoptera: Aphididae) Populations in Irrigated Grain in southwestern Idaho. Environ. Entomol. 1992, 21, 1433–1440. [Google Scholar] [CrossRef]
- Alkhedir, H.; Karlovsky, P.; Vidal, S. Effect of light intensity on colour morph formation and performance of the grain aphid Sitobion avenae F. (Homoptera: Aphididae). J. Insect Physiol. 2010, 56, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Tanguy, S.; Dedryver, C.A. Reduced BYDV-PAV transmission by the grain aphid in a Triticum monococcum line. Eur. J. Plant Pathol. 2009, 123, 281–289. [Google Scholar] [CrossRef]
- Chougule, N.P.; Bonning, B.C. Toxins for transgenic resistance to hemipteran pests. Toxins 2012, 4, 405–429. [Google Scholar] [CrossRef] [PubMed]
- Gatehouse, A.M.R.; Gatehouse, J.A. Identifying proteins with insecticidal activity: Use of encoding genes to produce insect-resistant transgenic crops. Pest Manag. Sci. 2015, 52, 165–175. [Google Scholar] [CrossRef]
- Habibi, J.; Backus, E.A.; Czapla, T.H. Plant lectins affect survival of the potato leaf hopper (Homoptera: Cicadellidae). J. Econ. Entomol. 1993, 86, 945–951. [Google Scholar] [CrossRef]
- Jouanin, L.; Bonadé-Bottino, M.; Girard, C.; Morrot, G.; Giband, M. Transgenic plants for insect resistance. Plant Sci. 1998, 131, 1–11. [Google Scholar] [CrossRef]
- Vasconcelos, I.M.; Oliveira, J.T.A. Antinutritional properties of plant lectins. Toxicon 2004, 44, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Hilder, V.A.; Powell, K.S.; Gatehouse, A.M.R.; Gatehouse, J.A.; Gatehouse, L.N.; Shi, Y.; Hamilton, W.D.O.; Merryweather, A.; Newell, C.A.; Timans, J.C. Expression of snowdrop lectin in transgenic tobacco plants results in added protection against aphids. Transgen. Res. 1995, 4, 18–25. [Google Scholar] [CrossRef]
- Longstaff, M.; Powell, K.S.; Gatehouse, J.A.; Raemaekers, R.; Newell, C.A.; Hamilton, W.D. Production and purification of active snowdrop lectin in Escherichia coli. Eur. J. Biochem. 1998, 252, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Powell, K.S.; Spence, J.; Bharathi, M.; Gatehouse, J.A.; Gatehouse, A.M.R. Transgenic rice plants expressing the snowdrop lectin gene (gna) exhibit high-level resistance to the whitebacked planthopper (Sogatella furcifera). Theor. Appl. Genet. 2004, 109, 1399–1405. [Google Scholar]
- Down, R.E.; Gatehouse, A.M.R.; Hamilton, W.D.O.; Gatehouse, J.A. Snowdrop lectin inhibits development and decreases fecundity of the Glasshouse Potato Aphid (Aulacorthum solani) when administered in vitro and via transgenic plants both in laboratory and glasshouse trials. J. Insect Physiol. 1996, 42, 1035–1045. [Google Scholar] [CrossRef]
- Gao, Z.; Zheng, B.; Wang, W.; Li, Q.; Yuan, Q. Cloning and functional characterization of a GNA-like lectin from Chinese Narcissus (Narcissus tazetta var. Chinensis Roem). Physiol. Plant. 2011, 142, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, K.; Ding, R.; Zhang, H.; Di, P.; Chen, J.; Zhang, L.; Chen, W. Transgenic tetraploid Isatis indigotica expressing Bt Cry1Ac and Pinellia ternata agglutinin showed enhanced resistance to moths and aphids. Mol. Biol. Rep. 2012, 39, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Pang, Y.; Qi, H.; Wan, B.; Zhao, X.; Kong, W.; Sun, X.; Tang, K. Transgenic tobacco expressing Pinellia ternata agglutinin confers enhanced resistance to aphids. Transgen. Res. 2003, 12, 715–722. [Google Scholar] [CrossRef]
- Yao, J.; Zhao, X.; Qi, H.; Wan, B.; Chen, F.; Sun, X.; Yu, S.; Tang, K.X. Transgenic tobacco expressing an Arisaema heterophyllum agglutinin gene displays enhanced resistance to aphids. Can. J. Plant Sci. 2004, 84, 785–790. [Google Scholar]
- Yu, Y.; Wei, Z. Increased oriental armyworm and aphid resistance in transgenic wheat stably expressing Bacillus thuringiensis (Bt) endotoxin and Pinellia ternate agglutinin (PTA). Plant Cell Tissue Organ Cult. 2008, 94, 33–44. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, X.; Tang, K.; Wang, X.; Sun, X.; Zhou, K. A primary study of transferring the Pinellia tenata agglutinin (PTA) gene into rice and expression. Acta Genet. Sin. 2003, 30, 1013–1019. [Google Scholar]
- Zhao, X.; Yao, J.; Liao, Z.; Zhang, H.; Chen, F.; Wang, L.; Lu, Y.; Sun, X.; Yu, S.; Tang, K. Molecular cloning of a novel mannose-binding lectin gene from Arisaema heterophyllum. Plant Sci. 2003, 165, 55–60. [Google Scholar] [CrossRef]
- Duan, X.; Hou, Q.; Liang, R. Expression of Two Synthetic Lectin Genes sGNA and sNTL in Transgenic Wheat enchanced resistance to Aphids. Res. J. Biotechnol. 2015, 10, 11–18. [Google Scholar]
- Li, G.; Jiang, W.; Xia, Q.; Chen, S.; Ge, X.; Gui, S.; Xu, C. HPV E6 down-regulation and apoptosis induction of human cervical cancer cells by a novel lipid-soluble extract (PE) from Pinellia pedatisecta Schott in vitro. J. Ethnopharmacol. 2010, 132, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Stoger, E.; Williams, S.; Christou, P.; Down, R.E.; Gatehouse, J.A. Expression of the insecticidal lectin from snowdrop (Galanthus nivalis agglutinin; GNA) in transgenic wheat plants: Effects on predation by the grain aphids Sitobion avenae. Mol. Breed. 1999, 5, 65–73. [Google Scholar] [CrossRef]
- Liang, H.; Zhu, Y.; Zhu, Z.; Sun, D.; Jia, X. Obtainment of transgenic wheat with the insecticidal lectin from snowdrop (Galanthus nivalis agglutinin; GNA) gene and analysis of resistance to aphid. Acta Genet. Sin. 2004, 31, 189–194. [Google Scholar] [PubMed]
- Xu, L.; Duan, X.; Lv, Y.; Zhang, X.; Nie, Z.; Xie, C.; Ni, Z.; Liang, R. Silencing of an aphid carboxylesterase gene by use of plant-mediated RNAi impairs Sitobion avenae tolerance of Phoxim insecticides. Transgen. Res. 2014, 23, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Z.; Wang, X.; Ding, H.; Ni, H.; Sun, J.; Cheng, D.; Chen, J. Evaluation of resistance of wheat varieties (lines) to aphids using fuzzy recognition. Plant Prot. 1998, 24, 15–16. [Google Scholar]
- Liu, X.; Wang, C.; Wang, Y.; Zhang, H.; Ji, W. Screening and Evaluation of different Wheat cultivars for resistance to Sitobion avenae at seedling and adult-plant stages. Agric. Sci. Technol. 2015, 16, 1686–1692. [Google Scholar]
- Zhou, H.; Chen, J.; Cheng, D.; Francis, F.; Liu, Y.; Sun, J.; Huang, Y.; Wang, X.; Liu, X.; Liu, X.; et al. Evaluation on the resistance to aphids of wheat germplasm resources in China. Afr. J. Biotechnol. 2011, 10, 13930–13935. [Google Scholar]
Sample Availability: Samples of pBAC-rbcs-ppa expression vector and BF104 are available from the authors. |










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, X.; Hou, Q.; Liu, G.; Pang, X.; Niu, Z.; Wang, X.; Zhang, Y.; Li, B.; Liang, R. Expression of Pinellia pedatisecta Lectin Gene in Transgenic Wheat Enhances Resistance to Wheat Aphids. Molecules 2018, 23, 748. https://doi.org/10.3390/molecules23040748
Duan X, Hou Q, Liu G, Pang X, Niu Z, Wang X, Zhang Y, Li B, Liang R. Expression of Pinellia pedatisecta Lectin Gene in Transgenic Wheat Enhances Resistance to Wheat Aphids. Molecules. 2018; 23(4):748. https://doi.org/10.3390/molecules23040748
Chicago/Turabian StyleDuan, Xiaoliang, Qiling Hou, Guoyu Liu, Xiaomeng Pang, Zhenli Niu, Xiao Wang, Yufeng Zhang, Baoyun Li, and Rongqi Liang. 2018. "Expression of Pinellia pedatisecta Lectin Gene in Transgenic Wheat Enhances Resistance to Wheat Aphids" Molecules 23, no. 4: 748. https://doi.org/10.3390/molecules23040748