Structural Diversity and Biological Activities of Novel Secondary Metabolites from Endophytes


1
Department of Natural Medicine and Pharmacognosy, School of Pharmacy, Qingdao University, Qingdao 266021, China
2
Department of Natural Product Chemistry, Key Laboratory of Chemical Biology of Ministry of Education, School of Pharmaceutical Sciences, Shandong University, No. 44 West Wenhua Road, Jinan 250012, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2018, 23(3), 646; https://doi.org/10.3390/molecules23030646
Submission received: 23 February 2018
/
Revised: 10 March 2018
/
Accepted: 11 March 2018
/
Published: 13 March 2018
(This article belongs to the Collection Bioactive Compounds)
Abstract
:Exploration of structurally novel natural products greatly facilitates the discovery of biologically active pharmacophores that are biologically validated starting points for the development of new drugs. Endophytes that colonize the internal tissues of plant species, have been proven to produce a large number of structurally diverse secondary metabolites. These molecules exhibit remarkable biological activities, including antimicrobial, anticancer, anti-inflammatory and antiviral properties, to name but a few. This review surveys the structurally diverse natural products with new carbon skeletons, unusual ring systems, or rare structural moieties that have been isolated from endophytes between 1996 and 2016. It covers their structures and bioactivities. Biosynthesis and/or total syntheses of some important compounds are also highlighted. Some novel secondary metabolites with marked biological activities might deserve more attention from chemists and biologists in further studies.
1. Introduction
The last 30 years have seen tremendous successes in natural product-based drug discovery [1,2]. Natural products, their semisynthetic derivatives, and synthetic products that mimic a natural product template, represent more than half of all approved small-molecule drugs [1,3]. Diverse and biologically active pharmacophores, especially in naturally occurring novel compounds, play a pivotal role in modern drug discovery [4,5]. They possess specific steric and electronic properties for molecular recognition by a biological target [6]. Alarmingly, only a few new natural product drug pharmacophores have been discovered in the last twenty years, which poses critical issues for natural product-driven lead discovery campaigns and new drug types [7].
Many strategies have been developed to discover structurally novel natural product leads through available biological approaches [8]. Mining the largely unexplored natural sources, such as endophytes, will pave the way for chemical and biological novelties [8]. Endophytes, mainly fungi and bacteria, colonize the living, internal plant tissues without causing visible symptoms of disease [9]. There are approximately 300,000 different plant species inhabiting our planet and it can be expected that each individual one has a complex community of one to many cultivable or uncultivable endophytic microorganisms [10,11]. Endophytes are recognized to have complex associations with host plants and other organisms, including endophytic microorganisms in their ecological niches and pathogens in external environments [12,13,14]. In order to adapt to their microenvironments, endophytes typically coevolve a plethora of traits that range from production of diverse chemical defense compounds to triggers for activating cryptic biosynthetic pathways, production of precursors, quorum sensing molecules, epigenetic modulators, and even direct physical organismal interactions [15,16]. These functional biomolecules derived from endophytes are important from an ecological perspective [13]. For instance, the endophytic fungus Neotyphodium coenophialum inhabiting the tall fescue (Festuca arundinacea) was discovered to produce toxic alkaloids, defending host plants against herbivorous mammals and causing “fescue toxicosis” of livestock [17]. From the medicinal perspective, they may directly or indirectly be used as therapeutic agents against numerous diseases.
The enormous diversity of endophytes in combination with their potential biosynthetic capabilities has provided the impetus for a number of chemical investigations on endophytes. Endophytes are now well-known to biosynthesize diverse natural products with intriguing biological activities, and around ten reviews have reported on the new and known bioactive secondary metabolites of endophytes [18,19,20,21,22,23,24,25,26,27]. It should be noted that small molecules with new carbon skeletons, unusual ring systems, or rare structural moieties from endophytic fungi and bacteria have not been reviewed to the best of our knowledge. They might deserve attention from chemists and biologists and could be a potential resource of new biologically active pharmacophores for natural product-based drug development.
The target of this review is to summarize endophyte-derived secondary metabolites with new carbon skeletons, unique ring systems, or uncommon structural moieties isolated in a period between 1996 and 2016 that marks enormous progress in the chemical investigation of fungal and bacterial endophytes. Their structures and biological activities, together with the biosynthesis and total syntheses of some important molecules are described. In this review, the structures are mainly classified according to their proposed biosynthesis. They might be further arranged according to the structural features of secondary metabolites.
2. Polyketides
2.1. Macrolides
A mangrove-derived bacterial endophyte Streptomyces sp. was discovered by the Hertweck group to produce four unprecedented ansa macrolides, divergolides A–D (1–4, Figure 1) [28,29].
They were biosynthesized from a common linear polyketide precursor that underwent various reactions including an optional acyl migration to form the diverse multicyclic structures (Scheme 1). An unusual isobutylmalonyl-CoA (ibMCoA) extender unit derived from isobutyrate and acetate rather than l-leucine was involved in the divergolide polyketide pathway (Scheme 1) [30]. The remarkable structural plasticity of this kind of macrolides led to different antibacterial and cytotoxic properties [31]. Compound 1 showed the strongest antibacterial activity against Mycobacterium vaccae with an inhibitory zone of 19 mm at 50 μg per paper disk in the disk diffusion assay, while compound 4 demonstrated marked cytotoxicity against several cancer cell lines, with IC50 values ranging from 1.0 to 2.0 μM [28]. Their intriguing structures and associated antibacterial or antitumor activities have stimulated various synthetic methods towards divergolides [32,33], and scientific interest in biosynthetic gene clusters [29].
Iwatsuki and co-workers obtained a fungus Actinoallomurus fulvus harbored in the roots of Capsicum frutescens collected in Thailand [34]. Chemical investigation of this fungus led to the discovery of five unique 12-membered macrolides, actinoallolides A–E (5–9, Figure 2). Compound 5 exhibited significant anti-trypanosomal activity against Trypansoma cruzi (IC50: 0.226 μg/mL) similar to that of commonly used therapeutic drug, benznidazole (IC50: 0.418 μg/mL), indicating a promising new class of lead compounds for treating Chagas disease [34]. Bioassay-guided isolation of the ethyl acetate extract of an unidentified endophytic fungus provided an unusual C16 nonenolide, microcarpalide (10) (Figure 2) with an alky side chain. Compound 10 disrupted microfilaments in approximately half of the cells at a concentration of 0.5–1.0 μg/mL and showed weak cytotoxicity against two mammalian cell lines (KB and LoVo) [35].
2.2. Benzopyran
The Krohn group discovered seven rare chromanones, blennolides A–G (11–17, Figure 3) from an endophytic fungus Blennoria sp. occurring in Carpobrotus edulis found in the Canary Islands [36]. They displayed moderate antialgal activity against Chlorella fusca and antifungal activity against Microbotryum violaceum with radii of the zones of inhibition ranging from 5 to 9 mm with 50 μg per paper disk in the agar diffusion assay. Compounds 14–16 are unique natural products with a highly substituted γ-lactone moiety, while compound 17 is a novel heterodimer incorporating two unusual chromanone subunits, the monomer 11 and the deoxy analogue of monomer 15 [36]. Another unusual heterodimeric chromanone, noduliprevenone (18) (Figure 3), was isolated from a Mediterranean alga-derived endophyte Nodulisporium sp., and was a potential competitive inhibitor of cytochrome P450 1A with an IC50 value 6.5 ± 1.6 μM [37].
In the course of discovering novel and bioactive metabolites from endophytic fungi, the Krohn group found three more novel antimicrobial benzopyran derivatives, microsphaeropsones A–C (19–21, Figure 4) with a unique oxepino[2,3-b] chromen-6-one (ring-enlarged xanthone) skeleton from an endophyte Microsphaeropsis sp. isolated from the shoots of Lycium intricatum [38]. From an endophytic Chalara sp. isolated from the plant Artemisia vulgaris, isofusidienols A–D (22–25) (Figure 4) with an unprecedented chromone-3-oxepine moiety were found by the Zeeck group. Compounds 22 and 23 exhibited strong antibacterial activity against Bacillus subtilis with inhibition zones of 23 and 22 mm at 15 μg/disk, respectively [39]. Lycopodiellactone (26, Figure 4) with an uncommon δ-lactone and a rare 3-methylene isochromanone moiety, was obtained from a fungal endophyte Paraphaeosphaeria neglecta isolated from a Hawaiian indigenous plant, Lycopodiella cernua [40]. This metabolite might be biosynthesized by a polyketide pathway involving a key condensation of the δ-lactone and the 3-methylene isochromanone motif.
2.3. Spiro Compounds
Chemical investigation of the EtOAc extract of an endophytic fungus Pestalotiopsis virgatula led to the isolation of three cytotoxic metabolites named virgatolides A–C (27–29, Figure 5) [41]. They are new members of the rare benzannulated 6,6-spiroketal class of natural products and possess one or two γ-lactone units, representing the first occurrence of the γ-lactone units in the benzannulated 6,6-spiroketals. Jaroszewski and co-workers employed a hyphenated technique comprising HPLC-SPE-NMR to uncover some novel metabolites from Pestalotiopsis virgatula, an endophyte inhabiting the bark of Terminalia chebula [42,43]. Among them, pestalospiranes A and B (30 and 31, Figure 5) have an unprecedented 1,9,11,18-tetraoxadispiro[6.2.6.2]octadecane skeleton in addition to the characteristic benzo[c]-oxepin motif [43,44]. A bioinspired tandem dimerization-spiroketalization strategy to forge the unique dispiro skeleton of 31 has recently been described (Scheme 2) [44].
The Munro group from New Zealand disclosed the structure of spiro-mamakone A (32, Figure 5) from a non-sporulating endophytic fungus derived from the New Zealand native tree Knightia excels [45]. This compound belongs to the family of the structurally diverse spirobisnaphthalenes and represents the first spirobisnaphthalene analogue containing a new spiro-nonadiene skeleton [45]. Using feeding experiments conducted with different labeled acetates, the biosynthesis of compound 32 was investigated and found to involve the same two pentaketide-derived naphthalene units that underwent oxidative coupling and further extensive rearrangement [46]. Compound 32 exhibited significant cytotoxicity toward the P388 murine leukemia cell line (IC50 of 0.33 μM), and was also active against three selected bacteria [45]. A series of spiro-mamakone analogues have been synthesized for the investigation of structure-activity relationships, confirming the importance of the enedione moiety to bioactivities [47]. Penicillactones A–C (33–35, Figure 5) were biosynthesized by an endophytic fungus, Penicillium dangeardii residing in the plant Lysidice rhodostegia, and are novel natural products possessing a spirocyclic anhydride moiety [48]. Compounds 34 and 35 were active in inhibiting the release of β-glucuronidase from polymorphonuclear leukocytes with ED50 values of 2.58 and 1.57 μM, respectively.
2.4. Quinones
In 1995, Clardy and co-workers identified an endophytic fungus Pestalotiopsis microspora, which lived in the inner bark of the healthy host plant Torreya taxifolia but could be switched to have the pathological activity by environmental triggers [49]. Bioassay-guided investigation of the fermentation culture of P. microspora led to the isolation of an unusual dimeric quinone, (±)-torreyanic acid (36, Figure 6) [50]. Compound 36, as a cytotoxic agent, caused cell death by apoptosis with IC50 values ranging from 5.1 to 65.0 μM for 25 different human cell lines [50]. Inspired by a proposed biosynthetic scheme of 36, the Porco, Jr. and Mehta groups successfully applied a biomimetic electrocyclization/Diels-Alder dimerization cascade to construct the structure of 36 [51,52]. Ding et al. isolated and identified a novel torreyanic acid analogue (37, Figure 6) from a fungus Pestalotiopsis sp. inhabiting the lichen Clavaroids sp. [53].
2.5. Nitrogen-Containing Heterocycles
Chaetoglobins A (38) and B (39) (Figure 7), the first azaphilone alkaloid dimers formed through bonding between C-5 and C-5′ [54], were isolated from a fungus, Chaetomium globosum, residing inside the stem of Imperata cylindrical by the Tan group [55,56]. Compound 38 has been demonstrated to be significantly cytotoxic against the human breast cancer cell line MCF-7 and colon cancer cell line SW1116 with IC50 values of 42.1 and 55.7 μM, respectively. (−)-Alternarlactam (40, Figure 7), as a unique polyketide, was also described firstly by the Tan group and was obtained from a strain of Alternaria living inside the leaves of Carex aridula [57]. Compound 40 contains two important antitumor-related pharmacophores, cyclopentenone and isoquinolinone scaffolds, and was highly effective against human cervix HeLa adenocarcinoma cell and human hepatocellular carcinoma cell with IC50 of 4.2 μM and 5.9 μM, respectively. The total synthesis of 40 has been achieved through using two commercially available chemicals, 3,5-dimethoxyaniline and (±)-4-methyl-1,2-cyclo-pentanedione [57].
The Gao group reported a polyketide-derived isoquinoline alkaloid, fusarimine (41, Figure 7) containing a rare N-ethyl-4-methyl-7-carboxyisoquinoline carbon skeleton [58]. This compound can be derived biogenetically from a single hexaketide chain with an external nitrogen incorporated in the endophytic fungus Fusarium sp. occurring in the renowned insecticidal plant Melia azedarach. Duclauxamide A1 (42, Figure 7) was purified from the endophytic Penicillium manginii inhabiting the elder root of the traditional Chinese medicinal (TCM) plant Panax notoginseng by the Huang group [59]. As a polyketide-derived heptacyclic oligophenalenone dimer with an uncommon N-2-hydroxyethyl moiety [60], compound 42 demonstrated moderate cytotoxicity against HL-60, SMML-7721, A-549, MCF-7, and SW480 cancer cell lines with IC50 values ranging from 11 to 32 μM. From another TCM plant Camellia sinensis selected by Huang and co-workers, a bacterial endophyte, Streptomyces sp. was isolated and was found to produce a purple red solid, rubrolone B (43, Figure 7) with potential cardioprotection [61]. This metabolite belongs to the tropolone alkaloid family [62,63], but displays an expanded aromatic tropolone skeleton that includes a unique benzoic acid-pyridine inner salt fragment. Feeding experiments using 13C-labled acetates indicated a type-II polyketide synthase (PKS)-catalyzed biosynthesis route followed by complex oxidative rearrangements to form the tropolone ring system (Scheme 3) [61].
2.6. Others
Following an antimicrobial screening for bioactive metabolites from endophytic fungi [64], fusidilactone C (44, Figure 8) was purified and found to comprise an unusual and rigid oxoadamantane skeleton and also has two ether-bridged hemiacetals in addition to its spiro acetal structure [65,66]. A novel ketal-tethered intramolecular Diels-Alder cycloaddition has been developed for the synthesis of the 2-oxadecalin spiroketal core of 44 [67]. Cephalosol (45, Figure 8), isolated from Cephalosporium acremonium that used to reside as an endophyte in Trachelospermum jasminoides, showed strong antimicrobial activities against Escherichia coli, Pseudomonas fluorescens, Trichophyton rubrum, and Candida albicans with MIC values of 3.9, 3.9, 7.8 and 1.95 μg/mL, respectively [68]. Compound 45, with an unprecedented carbon skeleton, was proposed to be derived from a PKS pathway similar to that of alternariol and graphislactones [69], and has already been a total synthesis target [70]. An endophytic fungus from the leaves of Catharanthus roseus was identified as Penicillium sp. by the Asai group [71]. It produced citreoviripyrone A (46, Figure 8) with a bicyclo[4.2.0]octadiene arising from a key 8π-6π electrocyclization cascade route (Scheme 4) [72]. Compound 46 was toxic to human HCT 116 cells with a GI50 value of 10.4 μM. Recently, Hertweck and co-workers reported a polyketide-derived antibiotic, daldionin (47, Figure 8) with an unprecedented oxane-linked binaphthyl ring system, obtained from an orchid endophyte [73]. Another endophytic fungus Cryptosporiopsis sp. isolated from tissues of Viburnum tinus proved to produce viburspiran (48, Figure 8) [74]. It was the first eight-membered maleic anhydride natural product with potential antifungal activity against Microbotryum violaceum and Botrytis cinerea with radii of inhibition zones of 6 and 10 mm at 50 μg per paper disk, respectively [74]. Chemical investigation of the EtOAc extract of the mangrove-derived endophyte Corynespora cassiicola isolated from Laguncularia racemosa, provided five unusual octalactone derivatives, such as coryoctalactone E (49, Figure 8) [75].
Citrinals A and B (50 and 51, Figure 9) from the endophytic fungus Colletotrichum capsici represented a new compound class with a unique skeleton but displayed no cytotoxic activities [76,77]. Following a biochemical induction assay, cytoskyrins A and B (52 and 53, Figure 9) with a 1,3,6,8-tetrahydroxyanthraquinone-type carbon skeleton, were isolated from the endophytic fungus Cytospora sp. [78]. Compound 52 demonstrated strong biochemical induction assay (BIA; used to identify compounds that damage DNA or inhibit DNA synthesis) activity down to 12.5 ng while the biosynthetically related 53 was inactive. The total synthesis has already been reported by the group of Nicolaou by developing a cascade sequence called the “cytoskyrin cascade” [79,80,81].
3. Noribosomal Peptides
Aspertryptanthrins A–C (54–56, Figure 10), three new indole diketopiperazine alkaloids, were obtained from a strain of Aspergillus sp. isolated from the stem bark of Melia azedarach L. [82].
They possess a 6/5/6/6 tryptanthrin framework that is formed by a tryptophan unit and an anthranilate residue. In addition, compound 56 has an unusual 16-membered ring skeleton which was cyclized through the formation of phenylate. Spirobrocazines A and B (57 and 58, Figure 10) were isolated and identified from the mangrove-derived Penicillium brocae and possess a very rare spirocyclic skeleton [83]. Compound 57 showed moderate antibacterial activities against E. coli, S. aureus, and Vibrio harveyi with MIC values of 32.0, 16.0, and 64.0 μg/mL, respectively.
Neosartoryadins A (59) and B (60) (Figure 11) with a unique 6/6/6/5 quinazoline ring system connected directly to a 6/5/5 imidazoindolone ring system, represented a new class of quinazoline-containing indole alkaloids and displayed inhibitory effects against influenza A virus (H1N1) with IC50 values of 66 μM and 58 μM, respectively [84]. Their structures were proposed to be assembled by four amino acids l-tryptophan, anthranilic acid (ATA), l-valine, and 2-aminoisobutyric acid (Aib) in the endophytic fungus Neosartorya udagawae [84,85].
Antitumor screening of extracts of 43 endophytic fungi isolated from the leaves of the TCM plant Adenophora axilliflora enabled the discovery of a bioactive strain, Chaetomium sp. [86]. 1H-NMR and bioassay fractionation of the fungal culture led to the isolation of a tripeptide-derived alkaloidal metabolite, chaetominine (61, Figure 11) with a unique alanine-derived δ-lactam ring [87]. Compound 61 showed more potent cytotoxicity to the human leukemia K562 and colon cancer SW1116 cell lines than the positive drug 5-fluorouracil (IC50 values of 21.0 and 28.0 nM for compound 61, respectively; IC50 values of 33.0 and 76.0 nM for 5-fluorouracil, respectively) [86]. It was proposed to be biosynthesized from l-alanine, ATA, and d-tryptophan and has been a target for numerous synthetic efforts [88]. Apicidins A–C (62–64, Figure 11), three new members of a unique family of cyclic tetrapeptides, were isolated from a fungal endophyte Fusarium pallidoroseum by chemists from Merck research laboratories [89,90]. They showed a variety of potent antiprotozoal activities by reversibly inhibiting histone deacetylase (HDAC) and are attracting considerable attention for their anti-tumor effects [91,92,93]. In particular, compound 64 showed MIC values of 0.8, 101, and 69 nM against Besnoitia jellisoni, Eimeria tenella, and Plasmodium falciparum, respectively, and was slightly more active than compounds 62 and 63 [90].
4. Isoprenoids
4.1. Steroids
Solanioic acid (65, Figure 12), a degraded and rearranged steroid with an unprecedented carbon skeleton, has been isolated from the fungus Rhizoctonia solani obtained from tubers of the medicinal plant Cyperus rotundus [94]. It displayed significant inhibitory activities against B. subtilis, S. aureus, and methicillin-resistant S. aureus (MRSA) with MIC values around 1 μg/mL, and moderate antifungal activity against C. albicans with an MIC value of 16 μg/mL [94]. Asterogynins A (66) and B (67) (Figure 12), two unusual steroid-like metabolites with a tetracyclic carbocyclic ring system [95], were purified from the culture of Chalara alabamensis isolated from the host plant Asterogyne martiana [96]. More recently, four structurally related steroids, wortmannines A–C (68–70, Figure 12) and secovironolide (71, Figure 12) bearing an unusual five-membered B ring [97,98], were discovered from an endophytic fungus Talaromyces wortmannii living in Tripterygium wilfordii by the group of Yang.
4.2. Sesquiterpenoids
Chloropupukeananin (72, Figure 13) featuring a unique tricyclo-[4.3.1.03,7]-decane skeleton, was the first chlorinated pupukeananae derivative originated from a sesquiterpenoid in the plant endophyte Pestalotiopsis fici [99,100]. It inhibited the HIV-1 replication in C8166 cells at an IC50 of 14.6 μM and also exhibited weak antibacterial activity [100]. A key intermolecular Diels-Alder reaction followed by a subsequent carbonyl-ene reaction was proposed to be involved in the biosynthesis of compound 72 [100,101]. More novel pupukeananae derivatives with significant anti-HIV or anticancer activities, such as chloropestolide A (73) [102] and chloropupukeanolides A–E (74–78, Figure 13) [103,104], have also been reported from endophytes. Compounds 73 and 74 showed inhibitory effects on replication of the HIV-1 virus in C8166 cells with IC50 values of 62.4 and 6.9 μM, respectively, and inhibited the growth of HeLa cell line with IC50 values of 0.7 and 16.9 μM, respectively [102,103]. Compounds 76 and 77 demonstrated significant cytotoxicity against HeLa and HT29 cell lines with IC50 values ranging from 1.2 to 7.9 μM [104].
Periconianone A (79, Figure 13), the first member of sesquiterpenoids with a new 6/6/6 tricarbocyclic skeleton, was isolated from Periconia sp. derived from the medicinal plant Annonsa muricata [105]. An intramolecular aldol condensation for the formation of a carbon bond between C-4 and C-12 might result in the generation of the unusual six-membered carbonic ring, which has recently been utilized in the totally synthetic strategy to 79 (Scheme 5) [106].
Compound 79 exhibited significant neural anti-inflammatory activity against lipopolysaccharide (LPS)-induced NO production in mouse microglia BV2 cells with IC50 value of 0.15 μM (curcumin as a positive control, IC50 = 3.9 μM) [105]. Pestalotiopsin A (80, Figure 13), an immunosuppressive agent, was isolated from an endophytic fungus Pestalotiopsis sp. associated with Taxus brevifolia by the group of Clardy [107]. The oxatricyclic ring system in the sesquiterpenoid 80 is unprecedented among natural products. In 2015, Ding et al. isolated three plant-like sesquiterpenes, bacaryolanes A–C (81–83, Figure 13) from a mangrove-derived bacterial endophyte Streptomyces sp. [108]. They were identified as the mirror images of plant-derived caryolanes [109]. This discovery may point to complex cross-talk between plant and endophytic microorganisms [20].
4.3. Diterpenoids
From an unidentified fungus colonizing the plant Daphnopsis americana, guanacastepene A (84, Figure 14) and 14 biosynthetically related congers that comprised a unique family of diterpene natural products were found [110,111]. Compound 84 showed potent antibacterial activity against MRSA and vancomycin-resistant Enterococcus faecalis (VREF) through disrupting the cell membrane with inhibition zones of 11 and 9 mm at 100 μg per paper disk, respectively [111,112]. It has attracted numerous synthetic efforts or strategies toward the guanacastepenes [113,114]. Harziandione (85, Figure 14) and harzianone (86, Figure 14) are antimicrobial harziane diterpenes containing a unique tetracyclic scaffold from the potential biocontrol agents, Trichoderma spp. [115].
4.4. Sesterterpenoids
The group of She has been dedicated to the search for structurally unique and biologically active compounds from mangrove plant-derived fungal endophytes, especially Aspergillus spp. Five sesterterpenoids with an unprecedented carbon skeleton including asperterpenoid A (87), asperterpenols A and B (88 and 89), and aspterpenacids A and B (90 and 91), have been obtained (Figure 15) [116,117,118]. Among them, compound 87 with an unprecedented 5/7/(3)6/5 pentacyclic system, inhibited the Mycobacterium tuberculosis protein tyrosine phosphatase B with an IC50 value of 2.2 μM [116]. Compounds 88 and 89 possessing an unusual 5/8/6/6 tetracyclic ring skeleton, exhibited inhibitory activity against acetylcholinesterase (AChE) with IC50 values of 2.3 μM and 3.0 μM, respectively [118]. There were no antibacterial and cytotoxic activities for compounds 90 and 91 with a rare carbon skeleton of a 5/3/7/6/5 ring system [117].
5. Hybrid Products
5.1. PKS-NRPS
Cytochalasans are a large class of fungal secondary metabolites with biological diversity originating from a mixed PKS and nonribosomal peptide synthetase (NRPS) [119]. The group of Dai isolated an endophytic fungus Periconia sp. from the medicinal plant Annona muricata, and discovered it was cytotoxic to several human cancer cell lines. Bioassay-guided isolation of EtOAc extracts of the different fermentation media of this strain resulted in the isolation and identification of twelve novel PKS-NRPS hybrid cytochalasans [120,121,122,123,124]. Among them, periconiasins A and B (92 and 93, Figure 16) with an unprecedented 9/6/5 tricyclic ring system exhibited significant cytotoxicity against human HCT-8 cancer cells with IC50 values of 0.9 and 0.8 μM, respectively [123]. Periconiasin D (94, Figure 16) has a 5/6/6/5 tetracyclic ring skeleton, while periconiasin G (95, Figure 16) is the first cytotoxic cytochalasan with a 7/6/5 tricyclic ring system [120]. Pericoannosin A (96, Figure 16) possesses an unusual hexahydro-1H-isochromen-5-isobutylpyrrolidin-2-one skeleton and showed moderate anti-HIV activity (IC50 of 69.6 μM) [122,124]. Compounds 92–96 were proposed to be biosynthesized from an unusual seven acetate/malonate polyketide chain attached to a leucine unit by a PKS-NRPS and a key Diels-Alder reaction should be occurred in the cyclization of cytochalasans [122,123]. Owing to their structural diversity and biological activities, they have emerged as targets for bioinspired total syntheses [125].
From a fungal endophyte Trichoderma gamsii isolated from the traditional Chinese herb Panax notoginseng, three more unique cytochalasans, trichoderones A (97) and B (98) (Figure 16) together with trichodermone (99, Figure 16), were obtained by Zou and co-workers [126,127]. Their structures with an unprecedented pentacyclic or tetracyclic ring system might originate from a key intramolecular Michael 1,4-addition of the possible biosynthetic precursor aspochalasin D [126,127]. Compounds 97 and 98 showed weak inhibitory activity against the HeLa cell lines with IC50 values over 40 μM [127]. Recently, phomopchalasins A (100) and B (101) (Figure 17), two novel cytochalasans featuring unprecedented 5/6/5/8-fused tetracyclic or 5/6/6/7/5-fused pentacyclic skeletons, were isolated from the endophytic fungus Phomopsis sp., and compound 101 showed antimigratory activity against MDA-MB-231 with IC50 value of 19.1 μM [128].
Chemical investigation of a mangrove-derived endophytic fungus Campylocarpon sp., led to the isolation of four novel cytotoxic 4-hydroxy-2-pyridone alkaloids, campyridones A–D (102–105, Figure 17) [129]. Their unprecedented ring systems containing a spiro-furanone or γ-pyrone substructure were proposed to be synthesized by the PKS-NRPS hybrid involving a polyketide chain and a tyrosine moiety.
Clardy and co-workers reported the isolation of two hybrid PKS-NRPS products: phaeosphaeride A (106, Figure 18) and its inactive diastereomer phaeosphaeride B (107, Figure 18) from an endophytic fungus Phaeosphaeria avenaria [130]. They were potent inhibitors of signal transducer and activator of transcription 3 (STAT3) signaling with an IC50 of 0.61 mM [130]. Their structural elucidations were achieved by spectral data [130], total synthesis [131,132] and X-ray crystallographic analysis [133]. The diastereomers or semi-synthetic derivatives of compounds 106 and 107 exhibited in vitro cytotoxicity against MD-MB-231, PANC-1, and A549 cancer cell lines [134,135]. Another biosynthetically related hybrid PKS-NRPS product, paraphaeosphaeride A (108, Figure 18), was discovered from an endophytic Paraphaeosphaeria neglecta isolated from the stem of Hawaiian-plant Lycopodiella cernua [136]. It has an unusual 4-pyranone-γ-lactam-1,4-thiazine moiety and showed STAT3 inhibition at 10 μM. The plausible hybrid biosynthetic pathway of compound 108 involving a precursor cysteine has been shown in Scheme 6 [136,137].
A proline-pentaketide amide, penibruguieramine A (109, Figure 18) with an unprecedented 1-hydroxy-2-methylpyrrolizidin-3-one skeleton, was isolated from Penicillium sp. associated with the Chinese mangrove Bruguiera gymnorrhiza [138]. A biomimetic total synthesis of compound 109 involving a key intramolecular aldol-type reaction was accomplished by Kim et al. [139]. The endophytic fungus Cryptosporiopsis cf. quercina produced a unique functionalized tetramic acid, cryptocin (110) (Figure 18) arising from a mixed PKS-NRPS pathway [140,141]. It demonstrated significant inhibitory activity against a wide variety of plant pathogens, including the fungus Pyricularia oryzae (the causal agent of rice blast disease) with an MIC value of 0.39 μg/mL [140]. Further total synthesis, semi-synthetic and biological studies by the group of Gao suggested the importance of different tetramic acid ring systems for cytotoxicity [142]. A high-throughput screen for endophytes-derived antimalarial compounds enabled the discovery of a new tryptophan-polyketide hybrid with a polyketide decalin [141], codinaeopsin (111, Figure 18) [143]. Compound 111 had an IC50 of 4.66 μM against P. falciparum, the causative agent of the most lethal form of malaria.
5.2. NRPS-Terpene
A mangrove-derived endophyte Mucor irregularis was found to produce three novel indole-diterpenes [144], named rhizovarins A–C (112–114, Figure 19) [145]. They appeared to be chemically unique due to the complex 4,6,6,8,5,6,6,6,6-fused indole-diterpene ring system that incorporates an unusual acetal linked to a hemiketal (112) or a ketal (113 and 114). Compounds 112 and 113 were effective against the human A-549 (IC50: 11.5 μM for 112; 6.3 μM for 113) and HL-60 cancer cell lines (IC50: 9.6 μM for 112; 5.0 μM for 113) [145].
The group of Ji isolated and identified a novel prenylated indole alkaloid, aspeverin (115, Figure 19) from an endophytic strain Aspergillus versicolor harbored in the marine green alga Codium fragile [146]. It showed inhibitory activity against marine phytoplankton (Heterosigma akashiwo) with the EC50 values of 16.7 and 9.0 μM for 24 and 96 h, respectively. The structure of compound 115 containing an unprecedented cyclic carbamate linkage and a rare cyano could be assembled through a dipeptide-like precursor with dimethylallyl pyrophosphate (DMAPP) [147,148], and has promoted the attention of chemists from a totally synthetic perspective [149]. Varioxepine A (116, Figure 19), a 3H-oxepine-containing alkaloid with an unprecedented oxa-cage unit, was isolated from Paecilomyces variotii, an endophytic fungus residing in marine red alga [150]. It showed diverse antibacterial activities with MIC values ranging from 16 to 64 μg/mL and inhibited plant pathogenic fungus Fusarium graminearum with an MIC value of 4 μg/mL. Like compounds 59–60, compound 116 could be biosynthesized by the condensation of ATA, valine, phenylalanine, and DMAPP [151].
5.3. PKS-Terpene
Two novel hybrid sesquiterpene-cyclopaldic acid metabolites, named pestalotiopens A (117) and B (118) (Figure 20), were obtained from the marine endophytic fungus Pestalotiopsis sp. [152]. Compound 117 showed moderate antibacterial activity against E. faecalis whereas compound 118 containing a third, triketide-derived moiety was inactive. Three unusual polyketide-sesquiterpene metabolites peyronellins A–C (119–121, Figure 20), have been isolated from the endophytic fungus Peyronellaea coffeae-arabicae, which was isolated from the native Hawaiian plant Pritchardia lowreyana [153]. Compound 119 was active against A2780 and A2780 CisR cancer cell lines with IC50 values of 1.8 and 3.4 μM, respectively, while compounds 120 and 121 were inactive.
5.4. PKS-NRPS-Terpene
From a plant endophytic fungus Emericella nidulans, emericellolides A–C (122–124, Figure 21) with an unprecedented macrolide skeleton were found by Li and co-workers. A l-glutamate fragment, an isoindolone unit [154], and a sesquiterpene moiety might be involved in the construction of the macro-ring in compounds 122–124 [155].
An endophytic fungus Aspergillus versicolor was isolated from the rhizome of Paris polyphylla var. yunnanensis by Zhou et al. and was found to biosynthesize five highly oxygenated cyclopiazonic acid-derived alkaloids, aspergillines A–E (125–129, Figure 22) [156,157]. Compounds 125–129 with a rigid hexacyclic (6/5/6/5/5/5) indole-tetrahydro-furan-tetramic acid scaffold, were proposed to arise from a mixed biosynthetic pathway that involves a tryptophan unit, one or two molecules of acetate, and DMAPP [158,159,160]. They not only exhibited significant anti-tobacco mosaic virus (anti-TMV) activity with IC50 values of 15.4–48.6 μΜ, but also showed moderate cytotoxicity against a panel of human cancer cell lines [157].
6. Conclusions
Stereochemically complex and structurally diverse secondary metabolites play a pivotal role in discovery campaigns for new natural product drug pharmacophores. A number of structurally novel compounds are increasingly being discovered from endophytic fungi and bacteria and could comprise a powerful compound library for drug lead development. Herein, we present a comprehensive review of 129 endophyte-derived natural products with new carbon skeletons, unusual ring systems, or rare structural moieties (Table 1). Most of them were discovered from fungal endophytes in which more than 70% were isolated from terrestrial plants, especially those with an ethnobotanical history. The structural novelty and diversity of these microbial metabolites are as a result of the enormous diversity of terrestrial and marine endophytes in combination with their potential biosynthetic capabilities. In addition, they display diverse and remarkable biological activities, and frequently reported biological properties are antimicrobial and cytotoxic activities (Table 1). As shown in Figure 23, 16 secondary metabolites with marked biological activities might deserve more attention from chemists and biologists in further investigations.
Although traditional bioassay-guided chemical investigation encounters the frequent re-isolation of known compounds, it remains the most popular approach in discovering structurally novel small molecules from endophytes, especially with the aid of advanced analytical techniques, such as LC-MS. Alteration of easily accessible cultivation parameters, such as media composition, has well proven in this review to activate the silent gene clusters in endophytes and will continue to be used as a promising strategy for increasing the number of novel natural products by a single microbial strain.
Furthermore, recent advances in microbial genomics and metagenomics offer promising opportunities to access cryptic secondary metabolites. It is expected that most endophytic species are more likely to be uncultivable or poorly cultivable in standard laboratory conditions. Exploration of this largely unexplored source would provide more structurally unique compounds with properties suitable for a wide variety of biological and medicinal applications.
Acknowledgments
This work was financially supported in part by Scientific Research Foundation of Qingdao University (no. 29016010007010) and Key Laboratory of Chemical Biology (Ministry of Education) Open Projects Fund (no. CB-201705). We thankfully acknowledge James Oppong-Kyekyeku (Department of Pharmaceutical Chemistry, Kwame Nkrumah University of Science and Technology) for valuable discussions.
Author Contributions
H.G. and G.L. reviewed the literature and wrote the article. H.-X.L. guided and supervised writing of the article. All authors approved the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patridge, E.; Gareiss, P.; Kinch, M.S.; Hoyer, D. An analysis of FDA-approved drugs: Natural products and their derivatives. Drug Discov. Today 2016, 21, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Morris-Natschke, S.L.; Lee, K.H. Strategies for the optimization of natural leads to anticancer drugs or drug candidates. Med. Res. Rev. 2016, 36, 32–91. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Van Drie, J.H. Generation of three-dimensional pharmacophore models. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2013, 3, 449–464. [Google Scholar] [CrossRef]
- Butler, M.S.; Robertson, A.A.; Cooper, M.A. Natural product and natural product derived drugs in clinical trials. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Lou, H.X. Strategies to diversify natural products for drug discovery. Med. Res. Rev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Spiteller, P. Chemical ecology of fungi. Nat. Prod. Rep. 2015, 32, 971–993. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schaberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Spiteller, M. Are we ready for industrial production of bioactive plant secondary metabolites utilizing endophytes? Nat. Prod. Rep. 2011, 28, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Biol. 2012, 19, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Pandey, S.P.; Spiteller, M. Untapped mutualistic paradigms linking host plant and endophytic fungal production of similar bioactive secondary metabolites. Phytochemistry 2013, 91, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef] [PubMed]
- Scherlach, K.; Hertweck, C. Mediators of mutualistic microbe-microbe interactions. Nat. Prod. Rep. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lyons, P.C.; Plattner, R.D.; Bacon, C.W. Occurrence of peptide and clavine ergot alkaloids in tall fescue grass. Science 1986, 232, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Q.Y.; Jia, M.; Ming, Q.L.; Yue, W.; Rahman, K.; Qin, L.P.; Han, T. Endophytic fungi with antitumor activities: Their occurrence and anticancer compounds. Crit. Rev. Microbiol. 2016, 42, 454–473. [Google Scholar] [CrossRef] [PubMed]
- Nisa, H.; Kamili, A.N.; Nawchoo, I.A.; Shafi, S.; Shameem, N.; Bandh, S.A. Fungal endophytes as prolific source of phytochemicals and other bioactive natural products: A review. Microb. Pathog. 2015, 82, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Endophytic and epiphytic microbes as “sources” of bioactive agents. Front. Chem. 2015, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Verekar, S.A.; Bhave, S.V. Endophytic fungi: A reservoir of antibacterials. Front. Microbiol. 2015, 5, 715. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes—Secret producers of bioactive plant metabolites. Pharmazie 2013, 68, 499–505. [Google Scholar] [PubMed]
- Radic, N.; Strukelj, B. Endophytic fungi: The treasure chest of antibacterial substances. Phytomedicine 2012, 19, 1270–1284. [Google Scholar] [CrossRef] [PubMed]
- Kharwar, R.N.; Mishra, A.; Gond, S.K.; Stierle, A.; Stierle, D. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Song, Y.C.; Tan, R.X. Biology and chemistry of endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef] [PubMed]
- Gunatilaka, A.A.L. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity, and implications of their occurrence. J. Nat. Prod. 2006, 69, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Maier, A.; Fiebig, H.H.; Gorls, H.; Lin, W.H.; Peschel, G.; Hertweck, C. Divergolides A–D from a mangrove endophyte reveal an unparalleled plasticity in ansa-macrolide biosynthesis. Angew. Chem. Int. Ed. 2011, 50, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Li, S.R.; Zhao, G.S.; Sun, M.W.; He, H.G.; Wang, H.X.; Li, Y.Y.; Lu, C.H.; Shen, Y.M. Identification and characterization of the biosynthetic gene cluster of divergolides from Streptomyces sp. W112. Gene 2014, 544, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.L.; Ding, L.; Hertweck, C. A branched extender unit shared between two orthogonal polyketide pathways in an endophyte. Angew. Chem. Int. Ed. 2011, 50, 4667–4670. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Franke, J.; Hertweck, C. Divergolide congeners illuminate alternative reaction channels for ansamycin diversification. Org. Biomol. Chem. 2015, 13, 1618–1623. [Google Scholar] [CrossRef] [PubMed]
- Rasapalli, S.; Jarugumilli, G.; Yarrapothu, G.R.; Golen, J.A.; Rheingold, A.L. Synthesis of the naphthoquinone core of divergolides (C–D) and model studies for elaboration of the ansabridge. Tetrahedron Lett. 2013, 54, 2615–2618. [Google Scholar] [CrossRef]
- Nawrat, C.C.; Kitson, R.R.; Moody, C.J. Toward the total synthesis of hygrocin B and divergolide C: Construction of the naphthoquinone-azepinone core. Org. Lett. 2014, 16, 1896–1899. [Google Scholar] [CrossRef] [PubMed]
- Inahashi, Y.; Iwatsuki, M.; Ishiyama, A.; Matsumoto, A.; Hirose, T.; Oshita, J.; Sunazuka, T.; Panbangred, W.; Takahashi, Y.; Kaiser, M.; et al. Actinoallolides A–E, new anti-trypanosomal macrolides, produced by an endophytic actinomycete, Actinoallomurus fulvus MK10-036. Org. Lett. 2015, 17, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, A.S.; Yoshida, W.Y.; Mooberry, S.L.; Hemscheidt, T. The structure of microcarpalide, a microfilament disrupting agent from an endophytic fungus. Org. Lett. 2001, 3, 3479–3481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Krohn, K.; Zia, U.; Florke, U.; Pescitelli, G.; Di Bari, L.; Antus, S.; Kurtan, T.; Rheinheimer, J.; Draeger, S.; et al. New mono- and dimeric members of the secalonic acid family: Blennolides A–G isolated from the fungus Blennoria sp. Chem. Eur. J. 2008, 14, 4913–4923. [Google Scholar] [CrossRef] [PubMed]
- Pontius, A.; Krick, A.; Kehraus, S.; Foegen, S.E.; Muller, M.; Klimo, K.; Gerhauser, C.; Konig, G.M. Noduliprevenone: A novel heterodimeric chromanone with cancer chemopreventive potential. Chem. Eur. J. 2008, 14, 9860–9863. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Kouam, S.F.; Kuigoua, G.M.; Hussain, H.; Cludius-Brandt, S.; Florke, U.; Kurtan, T.; Pescitelli, G.; Di Bari, L.; Draeger, S.; et al. Xanthones and oxepino[2,3-b]chromones from three endophytic fungi. Chem. Eur. J. 2009, 15, 12121–12132. [Google Scholar] [CrossRef] [PubMed]
- Lösgen, S.; Magull, J.; Schulz, B.; Draeger, S.; Zeeck, A. Isofusidienols: Novel chromone-3-oxepines produced by the endophytic fungus Chalara sp. Eur. J. Org. Chem. 2008, 2008, 698–703. [Google Scholar] [CrossRef]
- Li, C.; Yang, B.; Fenstemacher, R.; Turkson, J.; Cao, S. Lycopodiellactone, an unusual δ-lactone-isochromanone from a Hawaiian plant-associated fungus Paraphaeosphaeria neglecta FT462. Tetrahedron Lett. 2015, 56, 1724–1727. [Google Scholar] [CrossRef]
- Li, J.; Li, L.; Si, Y.K.; Jiang, X.J.; Guo, L.D.; Che, Y.S. Virgatolides A–C, benzannulated spiroketals from the plant endophytic fungus Pestalotiopsis virgatula. Org. Lett. 2011, 13, 2670–2673. [Google Scholar] [CrossRef] [PubMed]
- Kesting, J.R.; Staerk, D.; Tejesvi, M.V.; Kini, K.R.; Prakash, H.S.; Jaroszewski, J.W. HPLC-SPE-NMR identification of a novel metabolite containing the benzo[c]oxepin skeleton from the endophytic fungus Pestalotiopsis virgatula culture. Planta Med. 2009, 75, 1104–1106. [Google Scholar] [CrossRef] [PubMed]
- Kesting, J.R.; Olsen, L.; Staerk, D.; Tejesvi, M.V.; Kini, K.R.; Prakash, H.S.; Jaroszewski, J.W. Production of unusual dispiro metabolites in Pestalotiopsis virgatula endophyte cultures: HPLC-SPE-NMR, electronic circular dichroism, and time-dependent density-functional computation study. J. Nat. Prod. 2011, 74, 2206–2215. [Google Scholar] [CrossRef] [PubMed]
- Badrinarayanan, S.; Squire, C.J.; Sperry, J.; Brimble, M.A. Bioinspired total synthesis and stereochemical revision of the fungal metabolite pestalospirane B. Org. Lett. 2017, 19, 3414–3417. [Google Scholar] [CrossRef] [PubMed]
- Van der Sar, S.A.; Blunt, J.W.; Munro, M.H. spiro-Mamakone A: A unique relative of the spirobisnaphthalene class of compounds. Org. Lett. 2006, 8, 2059–2061. [Google Scholar] [CrossRef] [PubMed]
- Van der Sar, S.A.; Lang, G.; Mitova, M.I.; Blunt, J.W.; Cole, A.L.; Cummings, N.; Ellis, G.; Munro, M.H. Biosynthesis of spiro-mamakone A, a structurally unprecedented fungal metabolite. J. Org. Chem. 2008, 73, 8635–8638. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.C.; Devenish, S.R.; Muscroft-Taylor, A.C.; Blunt, J.W.; Munro, M.H. Synthetic and biological studies on the spiro-mamakone system. Org. Biomol. Chem. 2008, 6, 3854–3862. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ding, G.; Li, Y.; Qu, J.; Ma, S.; Lv, H.; Liu, Y.; Wang, W.; Dai, J.; Tang, Y.; Yu, S. Structures and absolute configurations of penicillactones A–C from an endophytic microorganism, Penicillium dangeardii Pitt. Org. Lett. 2013, 15, 5206–5209. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Yang, X.S.; Schwartz, M.; Strobel, G.; Clardy, J. The Relationship between an Endangered North-American Tree and an Endophytic Fungus. Chem. Biol. 1995, 2, 721–727. [Google Scholar] [CrossRef]
- Lee, J.C.; Strobel, G.A.; Lobkovsky, E.; Clardy, J. Torreyanic acid: A selectively cytotoxic quinone dimer from the endophytic fungus Pestalotiopsis microspora. J. Org. Chem. 1996, 61, 3232–3233. [Google Scholar] [CrossRef]
- Li, C.; Johnson, R.P.; Porco, J.A. Total synthesis of the quinone epoxide dimer (+)-torreyanic acid: Application of a biomimetic oxidation/electrocyclization/Diels-Alder dimerization cascade. J. Am. Chem. Soc. 2003, 125, 5095–5106. [Google Scholar] [CrossRef] [PubMed]
- Mehta, G.; Pan, S.C. Total synthesis of the novel, biologically active epoxyquinone dimer (+/−)-torreyanic acid: A biomimetic approach. Org. Lett. 2004, 6, 3985–3988. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Li, Y.; Fu, S.; Liu, S.; Wei, J.; Che, Y. Ambuic acid and torreyanic acid derivatives from the endolichenic fungus Pestalotiopsis sp. J. Nat. Prod. 2009, 72, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.M.; Yang, S.X.; Qin, J.C. Azaphilones: Chemistry and biology. Chem. Rev. 2013, 113, 4755–4811. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Song, Y.C.; Chen, J.R.; Xu, C.; Ge, H.M.; Wang, X.T.; Tan, R.X. Chaetoglobosin U, a cytochalasan alkaloid from endophytic Chaetomium globosum IFB-E019. J. Nat. Prod. 2006, 69, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Ming Ge, H.; Yun Zhang, W.; Ding, G.; Saparpakorn, P.; Song, Y.C.; Hannongbua, S.; Tan, R.X. Chaetoglobins A and B, two unusual alkaloids from endophytic Chaetomium globosum culture. Chem. Commun. 2008. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.H.; Jiang, N.; Gu, W.; Ma, J.; Wang, Y.R.; Song, Y.C.; Tan, R.X. Characterization, synthesis and self-aggregation of (−)-alternarlactam: A new fungal cytotoxin with cyclopentenone and isoquinolinone scaffolds. Chem. Eur. J. 2010, 16, 14479–14485. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-X.; Xiao, J.; Laatsch, H.; Holstein, J.J.; Dittrich, B.; Zhang, Q.; Gao, J.-M. Fusarimine, a novel polyketide isoquinoline alkaloid, from the endophytic fungus Fusarium sp. LN12, isolated from Melia azedarach. Tetrahedron Lett. 2012, 53, 6372–6375. [Google Scholar] [CrossRef]
- Cao, P.; Yang, J.; Miao, C.P.; Yan, Y.; Ma, Y.T.; Li, X.N.; Zhao, L.X.; Huang, S.X. New duclauxamide from Penicillium manginii YIM PH30375 and structure revision of the duclauxin family. Org. Lett. 2015, 17, 1146–1149. [Google Scholar] [CrossRef] [PubMed]
- Elsebai, M.F.; Saleem, M.; Tejesvi, M.V.; Kajula, M.; Mattila, S.; Mehiri, M.; Turpeinen, A.; Pirttila, A.M. Fungal phenalenones: Chemistry, biology, biosynthesis and phylogeny. Nat. Prod. Rep. 2014, 31, 628–645. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Ma, Y.T.; Yang, J.; Horsman, G.P.; Luo, D.; Ji, X.; Huang, S.X. Tropolone ring construction in the biosynthesis of rubrolone B, a cationic tropolone alkaloid from endophytic Streptomyces. Org. Lett. 2016, 18, 1254–1257. [Google Scholar] [CrossRef] [PubMed]
- Palleroni, N.J.; Reichelt, K.E.; Mueller, D.; Epps, R.; Tabenkin, B.; Bull, D.N.; Schuep, W.; Berger, J. Production of a novel red pigment, rubrolone, by Streptomyces echinoruber sp. nov. I. taxonomy, fermentation and partial purification. J. Antibiot. 1978, 31, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Schuep, W.; Blount, J.F.; Williams, T.H.; Stempel, A. Production of a novel red pigment, rubrolone, by Streptomyces echinoruber Sp. nov. II. chemistry and structure elucidation. J. Antibiot. 1978, 31, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C.; Draeger, S.; Rommert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Krohn, K.; Biele, C.; Drogies, K.H.; Steingrover, K.; Aust, H.J.; Draeger, S.; Schulz, B. Fusidilactones, a new group of polycyclic lactones from an endophyte, Fusidium sp. Eur. J. Org. Chem. 2002. [CrossRef]
- Qin, S.; Krohn, K.; Flörke, U.; Schulz, B.; Draeger, S.; Pescitelli, G.; Salvadori, P.; Antus, S.; Kurtán, T. Two new fusidilactones from the fungal endophyte Fusidium sp. Eur. J. Org. Chem. 2009, 2009, 3279–3284. [Google Scholar] [CrossRef]
- Wang, J.S.; Hsung, R.P.; Ghosh, S.K. Stereoselective ketal-tethered intramolecular Diels-Alder cycloadditions. An approach to the 2-oxadecalin spiroketal core of antifungal agent fusidilactione C. Org. Lett. 2004, 6, 1939–1942. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Huang, W.Y.; Chen, J.R.; Yan, W.Z.; Xie, D.Q.; Tan, R.X. Cephalosol: An antimicrobial metabolite with an unprecedented skeleton from endophytic Cephalosporium acremonium IFB-E007. Chem. Eur. J. 2008, 14, 10670–10674. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, T.; Takenaka, Y.; Nagakura, N.; Hamada, N. 6H-dibenzo[b,d]pyran-6-one derivatives from the cultured lichen mycobionts of Graphis spp. and their biosynthetic origin. Phytochemistry 2003, 62, 71–75. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, N.; Cheng, B.; Zhai, H. Total synthesis of (+/−)-cephalosol. Org. Lett. 2012, 14, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Luo, D.; Yamashita, K.; Oshima, Y. Structures and biomimetic synthesis of novel alpha-pyrone polyketides of an endophytic Penicillium sp. in Catharanthus roseus. Org. Lett. 2013, 15, 1020–1023. [Google Scholar] [CrossRef] [PubMed]
- Eade, S.J.; Walter, M.W.; Byrne, C.; Odell, B.; Rodriguez, R.; Baldwin, J.E.; Adlington, R.M.; Moses, J.E. Biomimetic synthesis of pyrone-derived natural products: Exploring chemical pathways from a unique polyketide precursor. J. Org. Chem. 2008, 73, 4830–4839. [Google Scholar] [CrossRef] [PubMed]
- Barnes, E.C.; Jumpathong, J.; Lumyong, S.; Voigt, K.; Hertweck, C. Daldionin, an unprecedented binaphthyl derivative, and diverse polyketide congeners from a fungal orchid endophyte. Chem. Eur. J. 2016, 22, 4551–4555. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Hussain, H.; Ahmed, I.; Draeger, S.; Schulz, B.; Meier, K.; Steinert, M.; Pescitelli, G.; Kurtán, T.; Flörke, U.; et al. Viburspiran, an antifungal member of the octadride class of maleic anhydride natural products. Eur. J. Org. Chem. 2011, 2011, 808–812. [Google Scholar] [CrossRef]
- Ebrahim, W.; Aly, A.H.; Wray, V.; Proksch, P.; Debbab, A. Unusual octalactones from Corynespora cassiicola, an endophyte of Laguncularia racemosa. Tetrahedron Lett. 2013, 54, 6611–6614. [Google Scholar] [CrossRef]
- Zhu, T.J.; Du, L.; Hao, P.F.; Lin, Z.J.; Gu, Q.Q. Citrinal A, a novel tricyclic derivative of citrinin, from an algicolous fungus Penicillium sp. i-1-1. Chin. Chem. Lett. 2009, 20, 917–920. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, H.; Ma, H.; Jiang, J.; Sun, W.; Cheng, L.; Zhang, G.; Zhang, Y. Citrinal B, a new secondary metabolite from endophytic fungus Colletotrichum capsici and structure revision of citrinal A. Tetrahedron Lett. 2016, 57, 4250–4253. [Google Scholar] [CrossRef]
- Brady, S.F.; Singh, M.P.; Janso, J.E.; Clardy, J. Cytoskyrins A and B, new BIA active bisanthraquinones isolated from an endophytic fungus. Org. Lett. 2000, 2, 4047–4049. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Papageorgiou, C.D.; Piper, J.L.; Chadha, R.K. The cytoskyrin cascade: A facile entry into cytoskyrin A, deoxyrubroskyrin, rugulin, skyrin, and flavoskyrin model systems. Angew. Chem. Int. Ed. 2005, 44, 5846–5851. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Lim, Y.H.; Papageorgiou, C.D.; Piper, J.L. Total synthesis of (+)-rugulosin and (+)-2,2′-epi-cytoskyrin A through cascade reactions. Angew. Chem. Int. Ed. 2005, 44, 7917–7921. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Lim, Y.H.; Piper, J.L.; Papageorgiou, C.D. Total syntheses of 2,2′-epi-cytoskyrin A, rugulosin, and the alleged structure of rugulin. J. Am. Chem. Soc. 2007, 129, 4001–4013. [Google Scholar] [CrossRef] [PubMed]
- Lhamo, S.; Wang, X.-B.; Li, T.-X.; Wang, Y.; Li, Z.-R.; Shi, Y.-M.; Yang, M.-H.; Kong, L.-Y. Three unusual indole diketopiperazine alkaloids from a terrestrial-derived endophytic fungus, Aspergillus sp. Tetrahedron Lett. 2015, 56, 2823–2826. [Google Scholar] [CrossRef]
- Meng, L.H.; Wang, C.Y.; Mandi, A.; Li, X.M.; Hu, X.Y.; Kassack, M.U.; Kurtan, T.; Wang, B.G. Three diketopiperazine alkaloids with spirocyclic skeletons and one bisthiodiketopiperazine derivative from the mangrove-derived endophytic fungus Penicillium brocae MA-231. Org. Lett. 2016, 18, 5304–5307. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Zhou, G.; Zhu, M.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Neosartoryadins A and B, fumiquinazoline alkaloids from a mangrove-derived fungus Neosartorya udagawae HDN13-313. Org. Lett. 2016, 18, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.H.; Xu, M.Y.; Li, H.J.; Li, J.Q.; Chen, Y.X.; Ma, W.Z.; Li, Y.P.; Xu, J.; Yang, D.P.; Lan, W.J. Amino acid-directed strategy for inducing the marine-derived fungus Scedosporium apiospermum F41-1 to maximize alkaloid diversity. Org. Lett. 2017, 19, 4888–4891. [Google Scholar] [CrossRef] [PubMed]
- Jiao, R.H.; Xu, S.; Liu, J.Y.; Ge, H.M.; Ding, H.; Xu, C.; Zhu, H.L.; Tan, R.X. Chaetominine, a cytotoxic alkaloid produced by endophytic Chaetomium sp. IFB-E015. Org. Lett. 2006, 8, 5709–5712. [Google Scholar] [CrossRef] [PubMed]
- Nakao, Y.; Kuo, J.; Yoshida, W.Y.; Kelly, M.; Scheuer, P.J. More kapakahines from the marine sponge Cribrochalina olemda. Org. Lett. 2003, 5, 1387–1390. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Liang, K.; Tong, X.; Ding, M.; Li, D.; Xia, C. Biomimetic synthesis of (−)-chaetominine epimers via copper-catalyzed radical cyclization. Tetrahedron 2015, 71, 3699–3704. [Google Scholar] [CrossRef]
- DarkinRattray, S.J.; Gurnett, A.M.; Myers, R.W.; Dulski, P.M.; Crumley, T.M.; Allocco, J.J.; Cannova, C.; Meinke, P.T.; Colletti, S.L.; Bednarek, M.A.; et al. Apicidin: A novel antiprotozoal agent that inhibits parasite histone deacetylase. Proc. Natl. Acad. Sci. USA 1996, 93, 13143–13147. [Google Scholar] [CrossRef]
- Singh, S.B.; Zink, D.L.; Liesch, J.M.; Dombrowski, A.W.; Darkin-Rattray, S.J.; Schmatz, D.M.; Goetz, M.A. Structure, histone deacetylase, and antiprotozoal activities of apicidins B and C, congeners of apicidin with proline and valine substitutions. Org. Lett. 2001, 3, 2815–2818. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.Y.; Ahn, J.W.; Kim, H.S.; Lee, J.; Yoon, J.H. Apicidin inhibits cell growth by downregulating IGF-1R in salivary mucoepidermoid carcinoma cells. Oncol. Rep. 2015, 33, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Bauden, M.; Tassidis, H.; Ansari, D. In vitro cytotoxicity evaluation of HDAC inhibitor apicidin in pancreatic carcinoma cells subsequent time and dose dependent treatment. Toxicol. Lett. 2015, 236, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lai, Z.; Huang, W.; Ling, H.; Lin, M.; Tang, S.; Liu, Y.; Tao, Y. Apicidin inhibited proliferation and invasion and induced apoptosis via mitochondrial pathway in non-small cell lung cancer GLC-82 Cells. Anticancer Agents Med. Chem. 2017, 17, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Ratnaweera, P.B.; Williams, D.E.; Patrick, B.O.; de Silva, E.D.; Andersen, R.J. Solanioic acid, an antibacterial degraded steroid produced in culture by the fungus Rhizoctonia solani isolated from tubers of the medicinal plant Cyperus rotundus. Org. Lett. 2015, 17, 2074–2077. [Google Scholar] [CrossRef] [PubMed]
- Wipf, P.; Halter, R.J. Chemistry and biology of wortmannin. Org. Biomol. Chem. 2005, 3, 2053–2061. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Ross, L.; Tamayo, G.; Clardy, J. Asterogynins: Secondary metabolites from a Costa Rican endophytic fungus. Org. Lett. 2010, 12, 4661–4663. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.-E.; Yang, Z.-D.; Sheng, L.; Zhou, S.-Y.; Li, S.; Yao, X.-J.; Zhi, K.-K.; Wang, Y.-G.; Zhang, F. Secovironolide, a novel furanosteroid scaffold with a five-membered B ring from the endophytic fungus Talaromyces wortmannii LGT-4. Tetrahedron Lett. 2015, 56, 6754–6757. [Google Scholar] [CrossRef]
- Fu, G.-C.; Yang, Z.-D.; Zhou, S.-Y.; Li, X.-M.; Yu, H.-T.; Yao, X.-J.; Fang, J.-G.; Shu, Z.-M.; Xue, H.-Y.; Wang, Y.-G. Wortmannines A–C, three novel wortmannin derivatives with an unusual five-membered B ring from the endophytic fungus Talaromyces wortmannii LGT-4. Tetrahedron Lett. 2016, 57, 4608–4611. [Google Scholar] [CrossRef]
- Suzuki, T.; Kobayashi, S. Concise approach to pupukeanane skeleton: Synthetic study of chloropupukeananin. Org. Lett. 2010, 12, 2920–2923. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Liu, S.; Jiang, L.; Chen, X.; Guo, L.; Che, Y. Chloropupukeananin, the first chlorinated pupukeanane derivative, and its precursors from Pestalotiopsis fici. Org. Lett. 2008, 10, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miyajima, Y.; Suzuki, K.; Iwakiri, K.; Koshimizu, M.; Hirai, G.; Sodeoka, M.; Kobayashi, S. Unexpected Diels-Alder/carbonyl-ene cascade toward the biomimetic synthesis of chloropupukeananin. Org. Lett. 2013, 15, 1748–1751. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Liu, S.C.; Zheng, Z.H.; Chen, X.L.; Zhang, H.; Guo, L.D.; Che, Y.S. Chloropestolide A, an antitumor metabolite with an unprecedented spiroketal skeleton from Pestalotiopsis fici. Org. Lett. 2009, 11, 2836–2839. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Niu, S.; Lu, X.; Chen, X.; Zhang, H.; Guo, L.; Che, Y. Unique metabolites of Pestalotiopsis fici suggest a biosynthetic hypothesis involving a Diels-Alder reaction and then mechanistic diversification. Chem. Commun. 2010, 46, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Bruhn, T.; Guo, L.; Gotz, D.C.; Brun, R.; Stich, A.; Che, Y.; Bringmann, G. Chloropupukeanolides C-E: Cytotoxic pupukeanane chlorides with a spiroketal skeleton from Pestalotiopsis fici. Chem. Eur. J. 2011, 17, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ge, H.; Zou, J.H.; Tao, X.; Chen, R.; Dai, J. Periconianone A, a new 6/6/6 carbocyclic sesquiterpenoid from endophytic fungus Periconia sp. with neural anti-inflammatory activity. Org. Lett. 2014, 16, 1410–1413. [Google Scholar] [CrossRef] [PubMed]
- Liffert, R.; Linden, A.; Gademann, K. Total synthesis of the sesquiterpenoid periconianone a based on a postulated biogenesis. J. Am. Chem. Soc. 2017, 139, 16096–16099. [Google Scholar] [CrossRef] [PubMed]
- Pulici, M.; Sugawara, F.; Koshino, H.; Uzawa, J.; Yoshida, S.; Lobkovsky, E.; Clardy, J. Pestalotiopsins A and B: New caryophyllenes from an endophytic fungus of Taxus brevifolia. J. Org. Chem. 1996, 61, 2122–2124. [Google Scholar] [CrossRef]
- Ding, L.; Goerls, H.; Dornblut, K.; Lin, W.; Maier, A.; Fiebig, H.H.; Hertweck, C. Bacaryolanes A–C, rare bacterial caryolanes from a mangrove endophyte. J. Nat. Prod. 2015, 78, 2963–2967. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhao, Y.; Huang, G.D.; Wu, W.S. Four new compounds from Sinacalia tangutica. Helv. Chim. Acta 2008, 91, 1894–1901. [Google Scholar] [CrossRef]
- Brady, S.F.; Singh, M.P.; Janso, J.E.; Clardy, J. Guanacastepene, a fungal-derived diterpene antibiotic with a new carbon skeleton. J. Am. Chem. Soc. 2000, 122, 2116–2117. [Google Scholar] [CrossRef]
- Brady, S.F.; Bondi, S.M.; Clardy, J. The guanacastepenes: A highly diverse family of secondary metabolites produced by an endophytic fungus. J. Am. Chem. Soc. 2001, 123, 9900–9901. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Janso, J.E.; Luckman, S.W.; Brady, S.F.; Clardy, J.; Greenstein, M.; Maiese, W.M. Biological activity of guanacastepene, a novel diterpenoid antibiotic produced by an unidentified fungus CR115. J. Antibiot. 2000, 53, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.K.; Hughes, C.C.; Kennedy-Smith, J.J.; Gradl, S.N.; Trauner, D. Total synthesis of (−)-heptemerone B and (−)-guanacastepene E. J. Am. Chem. Soc. 2006, 128, 17057–17062. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.Z.; Sha, C.K. Stereoselective total syntheses of guanacastepenes N and O. Org. Lett. 2015, 17, 3486–3489. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.P.; Liang, X.R.; Yin, X.L.; Wang, G.; Ji, N.Y. Absolute configurations of unique harziane diterpenes from Trichoderma species. Org. Lett. 2012, 14, 3815–3817. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Huang, H.; Li, H.; Sun, X.; Huang, H.; Lu, Y.; Lin, Y.; Long, Y.; She, Z. Asperterpenoid A, a new sesterterpenoid as an inhibitor of Mycobacterium tuberculosis protein tyrosine phosphatase B from the culture of Aspergillus sp. 16-5c. Org. Lett. 2013, 15, 721–723. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, Y.; Chen, S.; Liu, Y.; Lu, Y.; Chen, D.; Lin, Y.; Huang, X.; She, Z. Aspterpenacids A and B, two sesterterpenoids from a mangrove endophytic fungus Aspergillus terreus H010. Org. Lett. 2016, 18, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.E.; Huang, H.R.; Shao, C.L.; Xia, X.K.; Ma, L.; Huang, X.S.; Lu, Y.J.; Lin, Y.C.; Long, Y.H.; She, Z.G. Asperterpenols A and B, new sesterterpenoids isolated from a mangrove endophytic fungus Aspergillus sp. 085242. Org. Lett. 2013, 15, 2522–2525. [Google Scholar] [CrossRef] [PubMed]
- Skellam, E. The biosynthesis of cytochalasans. Nat. Prod. Rep. 2017, 34, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tao, X.; Liu, J.; Chen, R.; Zhang, M.; Li, L.; Fang, X.; Yu, L.-Y.; Dai, J. Periconiasin G, a new cytochalasan with unprecedented 7/6/5 tricyclic ring system from the endophytic fungus Periconia sp. Tetrahedron Lett. 2016, 57, 796–799. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, D.; Zhang, M.; Liu, X.; Chen, R.; Zhao, J.; Li, L.; Wang, N.; Dai, J. Periconiasins I and J, two new cytochalasans from an endophytic fungus Periconia sp. Tetrahedron Lett. 2016, 57, 5794–5797. [Google Scholar] [CrossRef]
- Zhang, D.; Tao, X.; Chen, R.; Liu, J.; Li, L.; Fang, X.; Yu, L.; Dai, J. Pericoannosin A, a polyketide synthase-nonribosomal peptide synthetase hybrid metabolite with new carbon skeleton from the endophytic fungus Periconia sp. Org. Lett. 2015, 17, 4304–4307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.W.; Ge, H.L.; Xie, D.; Chen, R.D.; Zou, J.H.; Tao, X.Y.; Dai, J.G. Periconiasins A–C, new cytotoxic cytochalasans with an unprecedented 9/6/5 tricyclic ring system from endophytic fungus Periconia sp. Org. Lett. 2013, 15, 1674–1677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-W.; Tao, X.-Y.; Liu, J.-M.; Chen, R.-D.; Zhang, M.; Fang, X.-M.; Yu, L.-Y.; Dai, J.-G. A new polyketide synthase−nonribosomal peptide synthetase hybrid metabolite from plant endophytic fungus Periconia sp. Chin. Chem. Lett. 2016, 27, 640–642. [Google Scholar] [CrossRef]
- Tian, C.; Lei, X.; Wang, Y.; Dong, Z.; Liu, G.; Tang, Y. Total syntheses of periconiasins A–E. Angew. Chem. Int. Ed. 2016, 55, 6992–6996. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Wang, H.; Li, L.; Song, B.; Chen, H.; Zhang, H.; Liu, X.; Zou, Z. Trichodermone, a spiro-cytochalasan with a tetracyclic nucleus (7/5/6/5) skeleton from the plant endophytic fungus Trichoderma gamsii. J. Nat. Prod. 2014, 77, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Wang, H.; Li, L.; Chen, A.J.; Chen, L.; Chen, H.; Zhang, H.; Liu, X.; Zou, Z. Trichoderones A and B: Two pentacyclic cytochalasans from the plant endophytic fungus Trichoderma gamsii. Eur. J. Org. Chem. 2012, 2012, 2516–2519. [Google Scholar] [CrossRef]
- Yan, B.C.; Wang, W.G.; Hu, D.B.; Sun, X.; Kong, L.M.; Li, X.N.; Du, X.; Luo, S.H.; Liu, Y.; Li, Y.; et al. Phomopchalasins A and B, two cytochalasans with polycyclic-fused skeletons from the endophytic fungus Phomopsis sp. shj2. Org. Lett. 2016, 18, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhang, X.; Feng, H.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Campyridones A–D, pyridone alkaloids from a mangrove endophytic fungus Campylocarpon sp. HDN13-307. Tetrahedron 2016, 72, 5679–5683. [Google Scholar] [CrossRef]
- Maloney, K.N.; Hao, W.; Xu, J.; Gibbons, J.; Hucul, J.; Roll, D.; Brady, S.F.; Schroeder, F.C.; Clardy, J. Phaeosphaeride A, an inhibitor of STAT3-dependent signaling isolated from an endophytic fungus. Org. Lett. 2006, 8, 4067–4070. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Okamoto, I.; Morita, N.; Kiyotani, T.; Tamura, O. Synthesis of the proposed structure of phaeosphaeride A. Org. Biomol. Chem. 2011, 9, 5825–5832. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Kobayashi, Y.; Nakamura, M.; Tamura, O.; Kogen, H. Establishment of relative and absolute configurations of phaeosphaeride A: Total synthesis of ent-phaeosphaeride A. J. Org. Chem. 2015, 80, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Abzianidze, V.V.; Poluektova, E.V.; Bolshakova, K.P.; Panikorovskii, T.L.; Bogachenkov, A.S.; Berestetskiy, A.O. Crystal structure of natural phaeosphaeride A. Acta Crystallogr. Sect. E 2015, 71 Pt 8, o625–o626. [Google Scholar] [CrossRef] [PubMed]
- Chatzimpaloglou, A.; Yavropoulou, M.P.; Rooij, K.E.; Biedermann, R.; Mueller, U.; Kaskel, S.; Sarli, V. Total synthesis and biological activity of the proposed structure of phaeosphaeride A. J. Org. Chem. 2012, 77, 9659–9667. [Google Scholar] [CrossRef] [PubMed]
- Abzianidze, V.V.; Prokofieva, D.S.; Chisty, L.A.; Bolshakova, K.P.; Berestetskiy, A.O.; Panikorovskii, T.L.; Bogachenkov, A.S.; Holder, A.A. Synthesis of natural phaeosphaeride A derivatives and an in vitro evaluation of their anti-cancer potential. Bioorg. Med. Chem. Lett. 2015, 25, 5566–5569. [Google Scholar] [CrossRef] [PubMed]
- Li, C.S.; Ding, Y.; Yang, B.J.; Miklossy, G.; Yin, H.Q.; Walker, L.A.; Turkson, J.; Cao, S. A new metabolite with a unique 4-pyranone-gamma-lactam-1,4-thiazine moiety from a Hawaiian-plant associated fungus. Org. Lett. 2015, 17, 3556–3559. [Google Scholar] [CrossRef] [PubMed]
- Trenti, F.; Cox, R.J. Structural revision and biosynthesis of the fungal phytotoxins phyllostictines A and B. J. Nat. Prod. 2017, 80, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.F.; Kurtan, T.; Yang, X.H.; Mandi, A.; Geng, M.Y.; Ye, B.P.; Taglialatela-Scafati, O.; Guo, Y.W. Penibruguieramine A, a novel pyrrolizidine alkaloid from the endophytic fungus Penicillium sp. GD6 associated with Chinese mangrove Bruguiera gymnorrhiza. Org. Lett. 2014, 16, 1390–1393. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, S.; Kim, S. Biomimetic total synthesis of (−)-penibruguieramine A using memory of chirality and dynamic kinetic resolution. Angew. Chem. Int. Ed. 2015, 54, 10875–10878. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Strobel, G.; Harper, J.; Lobkovsky, E.; Clardy, J. Cryptocin, a potent tetramic acid antimycotic from the endophytic fungus Cryptosporiopsis cf. quercina. Org. Lett. 2000, 2, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Spiteller, M. Natural products containing ‘decalin’ motif in microorganisms. Nat. Prod. Rep. 2014, 31, 1175–1201. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Rao, M.; Ou, J.; Yin, J.; Lu, W.; Liu, M.; Pang, X.; Gao, S. Total synthesis and biological studies of cryptocin and derivatives of equisetin and fusarisetin A. Org. Biomol. Chem. 2014, 12, 7591–7597. [Google Scholar] [CrossRef] [PubMed]
- Kontnik, R.; Clardy, J. Codinaeopsin, an antimalarial fungal polyketide. Org. Lett. 2008, 10, 4149–4151. [Google Scholar] [CrossRef] [PubMed]
- Ondeyka, J.G.; Helms, G.L.; Hensens, O.D.; Goetz, M.A.; Zink, D.L.; Tsipouras, A.; Shoop, W.L.; Slayton, L.; Dombrowski, A.W.; Polishook, J.D.; et al. Nodulisporic acid A, a novel and potent insecticide from a Nodulisporium sp. isolation, structure determination, and chemical transformations. J. Am. Chem. Soc. 1997, 119, 8809–8816. [Google Scholar] [CrossRef]
- Gao, S.S.; Li, X.M.; Williams, K.; Proksch, P.; Ji, N.Y.; Wang, B.G. Rhizovarins A–F, indole-diterpenes from the mangrove-derived endophytic fungus Mucor irregularis QEN-189. J. Nat. Prod. 2016, 79, 2066–2074. [Google Scholar] [CrossRef] [PubMed]
- Ji, N.Y.; Liu, X.H.; Miao, F.P.; Qiao, M.F. Aspeverin, a new alkaloid from an algicolous strain of Aspergillus versicolor. Org. Lett. 2013, 15, 2327–2329. [Google Scholar] [CrossRef] [PubMed]
- Mugishima, T.; Tsuda, M.; Kasai, Y.; Ishiyama, H.; Fukushi, E.; Kawabata, J.; Watanabe, M.; Akao, K.; Kobayashi, J. Absolute stereochemistry of citrinadins A and B from marine-derived fungus. J. Org. Chem. 2005, 70, 9430–9435. [Google Scholar] [CrossRef] [PubMed]
- Finefield, J.M.; Frisvad, J.C.; Sherman, D.H.; Williams, R.M. Fungal origins of the bicyclo[2.2.2]diazaoctane ring system of prenylated indole alkaloids. J. Nat. Prod. 2012, 75, 812–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levinson, A.M. Total synthesis of aspeverin via an iodine(III)-mediated oxidative cyclization. Org. Lett. 2014, 16, 4904–4907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Mandi, A.; Li, X.M.; Du, F.Y.; Wang, J.N.; Li, X.; Kurtan, T.; Wang, B.G. Varioxepine A, a 3H-oxepine-containing alkaloid with a new oxa-cage from the marine algal-derived endophytic fungus Paecilomyces variotii. Org. Lett. 2014, 16, 4834–4837. [Google Scholar] [CrossRef] [PubMed]
- Doveston, R.G.; Taylor, R.J.K. An expedient synthesis of the proposed biosynthetic precursor of the oxepine natural product, janoxepin. Tetrahedron Lett. 2012, 53, 2533–2536. [Google Scholar] [CrossRef]
- Hemberger, Y.; Xu, J.; Wray, V.; Proksch, P.; Wu, J.; Bringmann, G. Pestalotiopens A and B: Stereochemically challenging flexible sesquiterpene-cyclopaldic acid hybrids from Pestalotiopsis sp. Chem. Eur. J. 2013, 19, 15556–15564. [Google Scholar] [CrossRef] [PubMed]
- Li, C.S.; Ren, G.; Yang, B.J.; Miklossy, G.; Turkson, J.; Fei, P.; Ding, Y.; Walker, L.A.; Cao, S. Meroterpenoids with antiproliferative activity from a Hawaiian-plant associated fungus Peyronellaea coffeae-arabicae FT238. Org. Lett. 2016, 18, 2335–2338. [Google Scholar] [CrossRef] [PubMed]
- Yaegashi, J.; Praseuth, M.B.; Tyan, S.W.; Sanchez, J.F.; Entwistle, R.; Chiang, Y.M.; Oakley, B.R.; Wang, C.C. Molecular genetic characterization of the biosynthesis cluster of a prenylated isoindolinone alkaloid aspernidine A in Aspergillus nidulans. Org. Lett. 2013, 15, 2862–2865. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Sun, X.; Li, N.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Isoindolone-containing meroperpenoids from the endophytic fungus Emericella nidulans HDN12-249. Org. Lett. 2016, 18, 4670–4673. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, C.W. The isolation and structure of cyclopiazonic acid, a toxic metabolite of Penicillium cyclopium Westling. Tetrahedron 1968, 24, 2101–2119. [Google Scholar] [CrossRef]
- Zhou, M.; Miao, M.M.; Du, G.; Li, X.N.; Shang, S.Z.; Zhao, W.; Liu, Z.H.; Yang, G.Y.; Che, C.T.; Hu, Q.F.; et al. Aspergillines A–E, highly oxygenated hexacyclic indole-tetrahydrofuran-tetramic acid derivatives from Aspergillus versicolor. Org. Lett. 2014, 16, 5016–5019. [Google Scholar] [CrossRef] [PubMed]
- Seshime, Y.; Juvvadi, P.R.; Tokuoka, M.; Koyama, Y.; Kitamoto, K.; Ebizuka, Y.; Fujii, I. Functional expression of the Aspergillus flavus PKS-NRPS hybrid CpaA involved in the biosynthesis of cyclopiazonic acid. Bioorg. Med. Chem. Lett. 2009, 19, 3288–3292. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Walsh, C.T. Cyclopiazonic acid biosynthesis in Aspergillus sp.: Characterization of a reductase-like R* domain in cyclopiazonate synthetase that forms and releases cyclo-acetoacetyl-l-tryptophan. Biochemistry 2009, 48, 8746–8757. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Walsh, C.T. Characterization of cyclo-acetoacetyl-L-tryptophan dimethylallyltransferase in cyclopiazonic acid biosynthesis: Substrate promiscuity and site directed mutagenesis studies. Biochemistry 2009, 48, 11032–11044. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |
Figure 1.
Structures of divergolides A–D (1–4).

Scheme 1.
The proposed biosynthetic pathway for divergolides A–D (1–4), starting from the ansamycin starter unit AHBA. The polyketide backbone is proposed to be disrupted through a putative Baeyer-Villiger oxidation.
Scheme 1.
The proposed biosynthetic pathway for divergolides A–D (1–4), starting from the ansamycin starter unit AHBA. The polyketide backbone is proposed to be disrupted through a putative Baeyer-Villiger oxidation.

Figure 2.
Structures of actinoallolides A–E (5–9) and microcarpalide (10).

Figure 3.
Structures of blennolides A–G (11–17) and noduliprevenone (18).

Figure 4.
Structures of microsphaeropsones A–C (19–21), isofusidienols A–D (22–25) and lycopodiellactone (26).
Figure 4.
Structures of microsphaeropsones A–C (19–21), isofusidienols A–D (22–25) and lycopodiellactone (26).

Figure 5.
Structures of compounds 27–35.

Scheme 2.
Bioinspired tandem dimerization-spiroketalization for the total synthesis of pestalospirane B (31).
Scheme 2.
Bioinspired tandem dimerization-spiroketalization for the total synthesis of pestalospirane B (31).

Figure 6.
Structures of (±)-torreyanic acid (36) and its analogue 37.

Figure 7.
Structures of compounds 38–43.

Scheme 3.
Biosynthesis of rubrolone B (43) on the basis of the data from feeding experiments.

Figure 8.
Structures of compounds 44–49.

Scheme 4.
Plausible biosynthetic pathway of citreoviripyrone A (46).

Figure 9.
Structures of citrinals A–B (50–51) and cytoskyrins A–B (52–53).

Figure 10.
Structures of aspertryptanthrins A–C (54–56) and spirobrocazines A–B (57–58).

Figure 11.
Structures of compounds 59–64.

Figure 12.
Structures of compounds 65–71.

Figure 13.
Structures of compounds 72–83.

Scheme 5.
(A) Proposed biosynthetic pathway for periconianone A (79); (B) Synthetic strategy to 79 inspired by the biogenetic hypothesis of 79.
Scheme 5.
(A) Proposed biosynthetic pathway for periconianone A (79); (B) Synthetic strategy to 79 inspired by the biogenetic hypothesis of 79.

Figure 14.
Structures of diterpenoids 84–86.

Figure 15.
Structures of sesterterpenoids 87–91.

Figure 16.
Structures of compounds 92–99.

Figure 17.
Structures of phomopchalasins A–B (100–101) and campyridones A–D (102–105).

Figure 18.
Structures of compounds 106–111.

Scheme 6.
Proposed biosynthetic pathway for paraphaeosphaeride A (108).

Figure 19.
Structures of compounds 112–116.

Figure 20.
Structures of pestalotiopens A–B (117–118) and peyronellins A–C (119–121).

Figure 21.
Structures of emericellolides A–C (122–124).

Figure 22.
Structures of aspergillines A–E (125–129).

Figure 23.
Selected endophyte-derived secondary metabolites with significant biological activities.


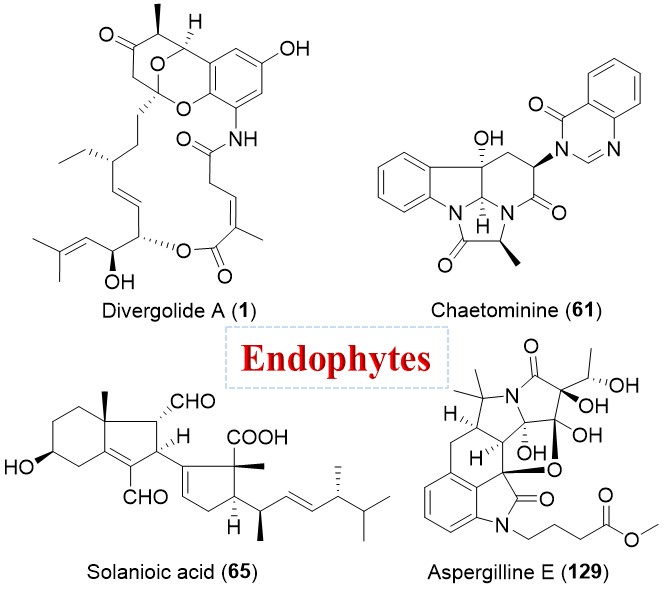{kind=link}
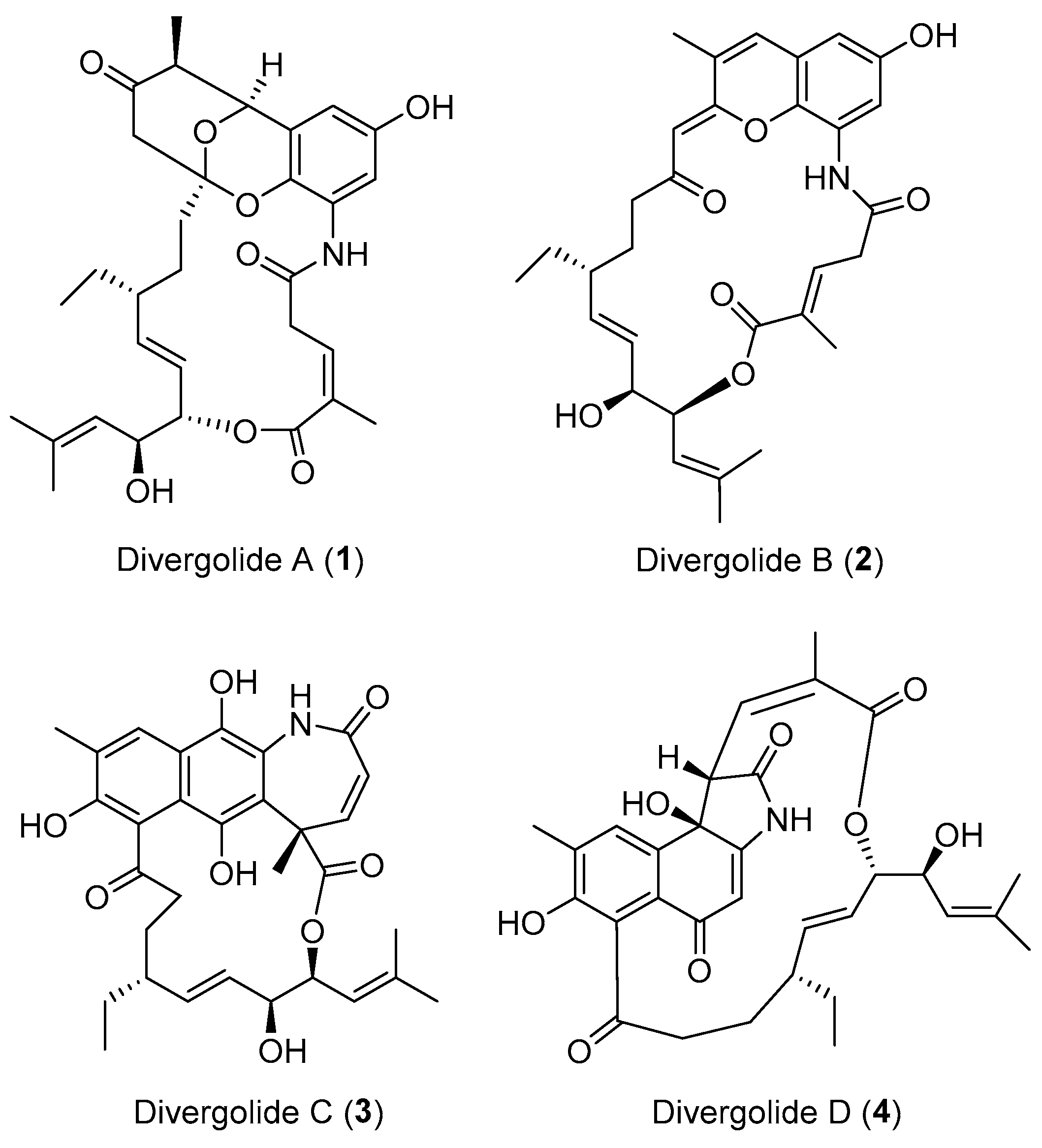{kind=link}
{kind=link}
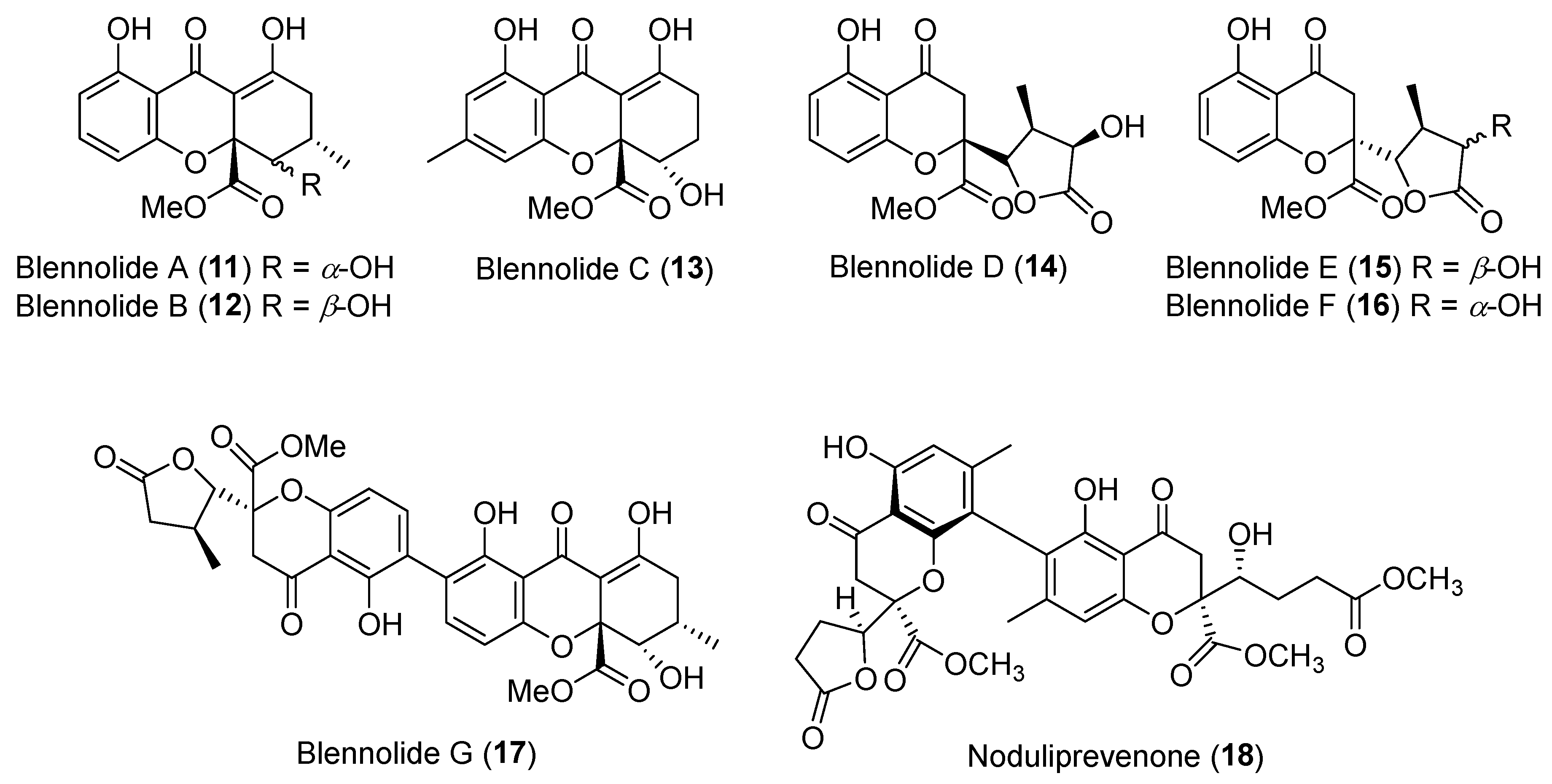{kind=link}
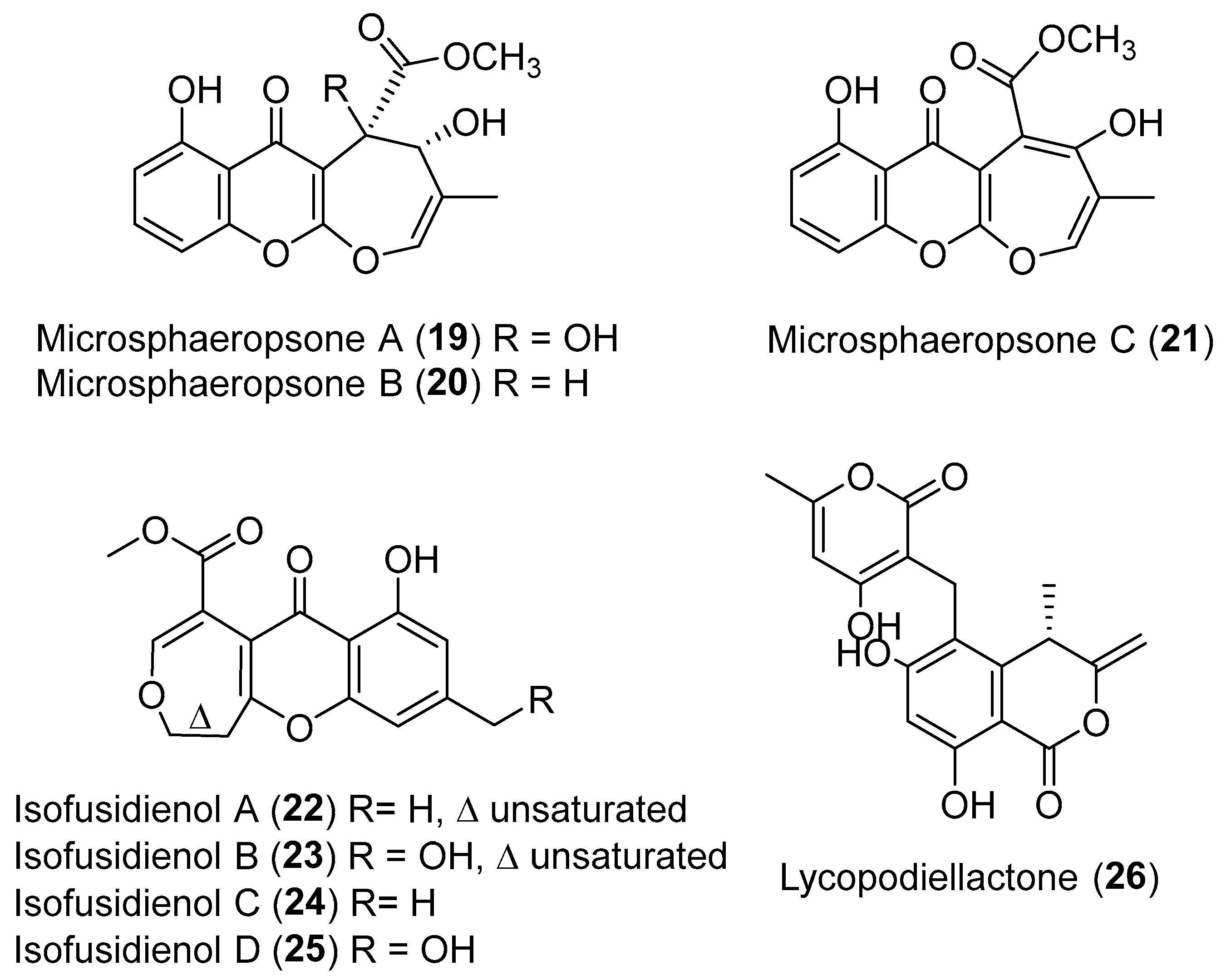{kind=link}
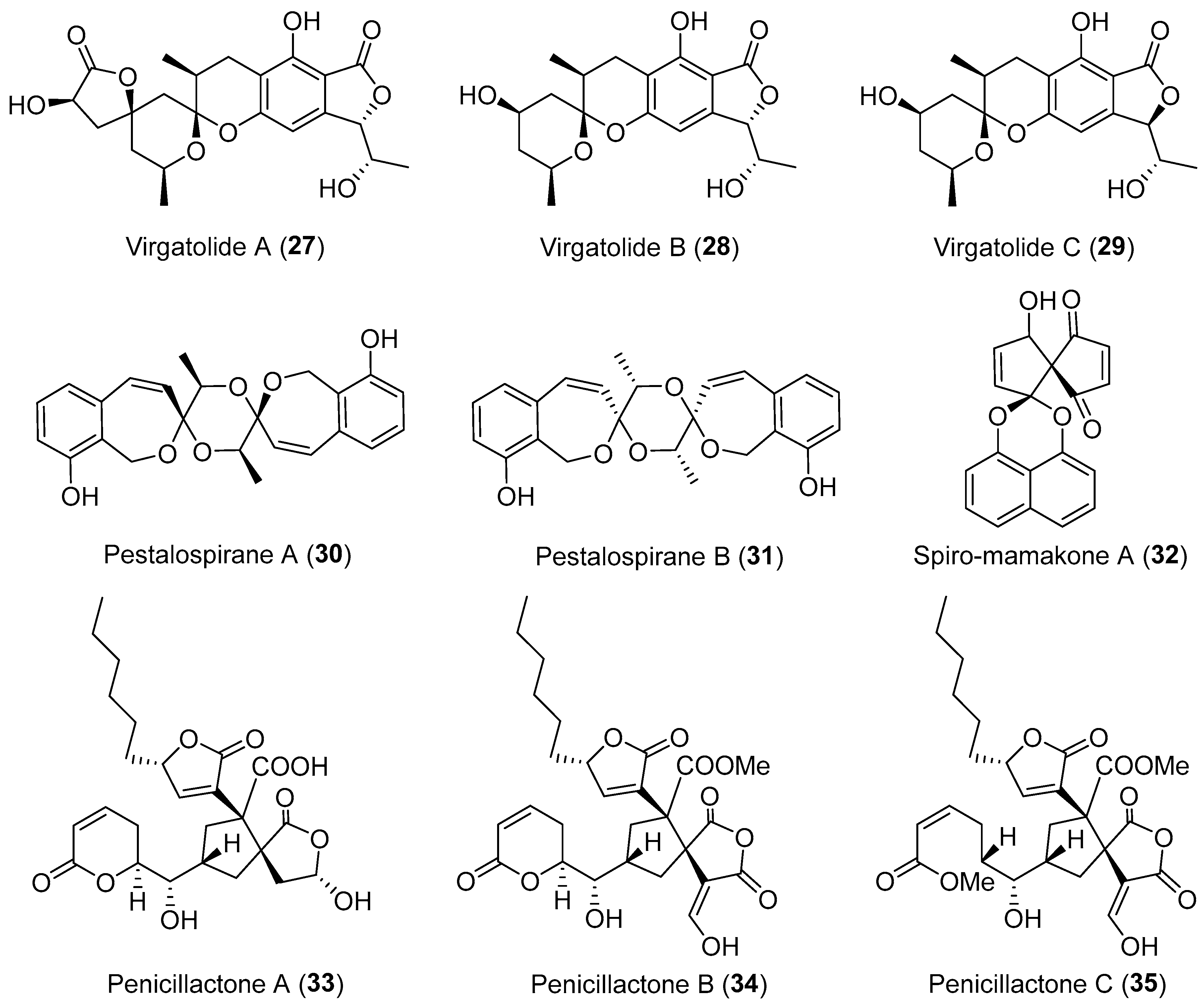{kind=link}
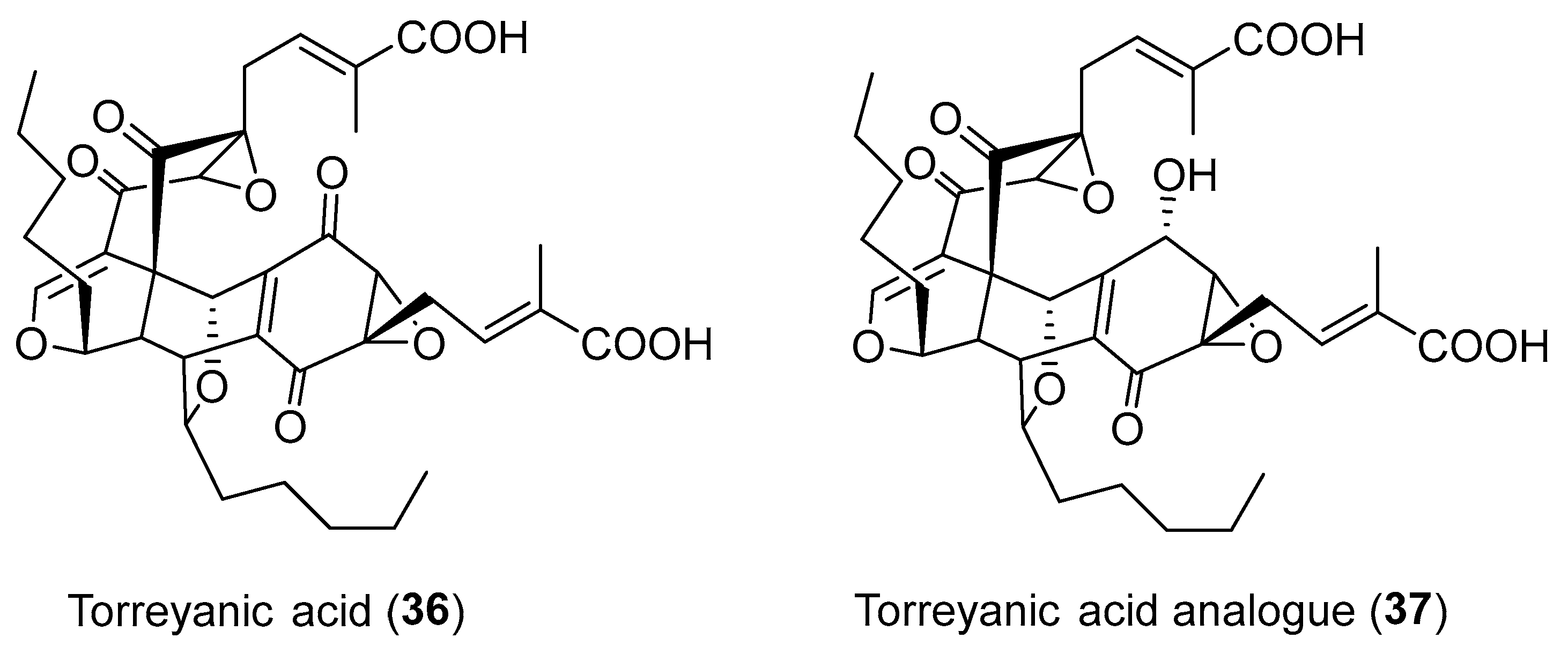{kind=link}
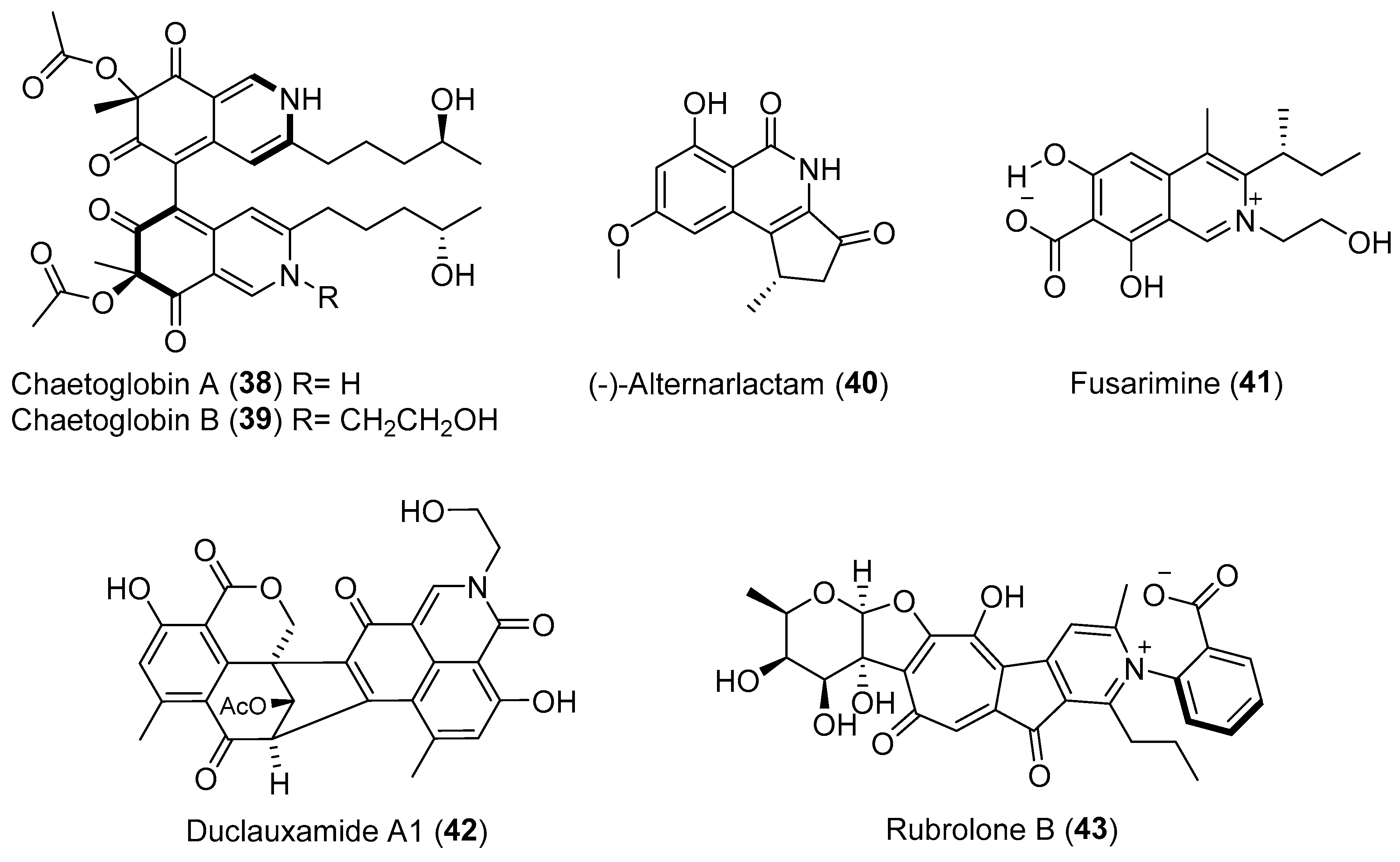{kind=link}
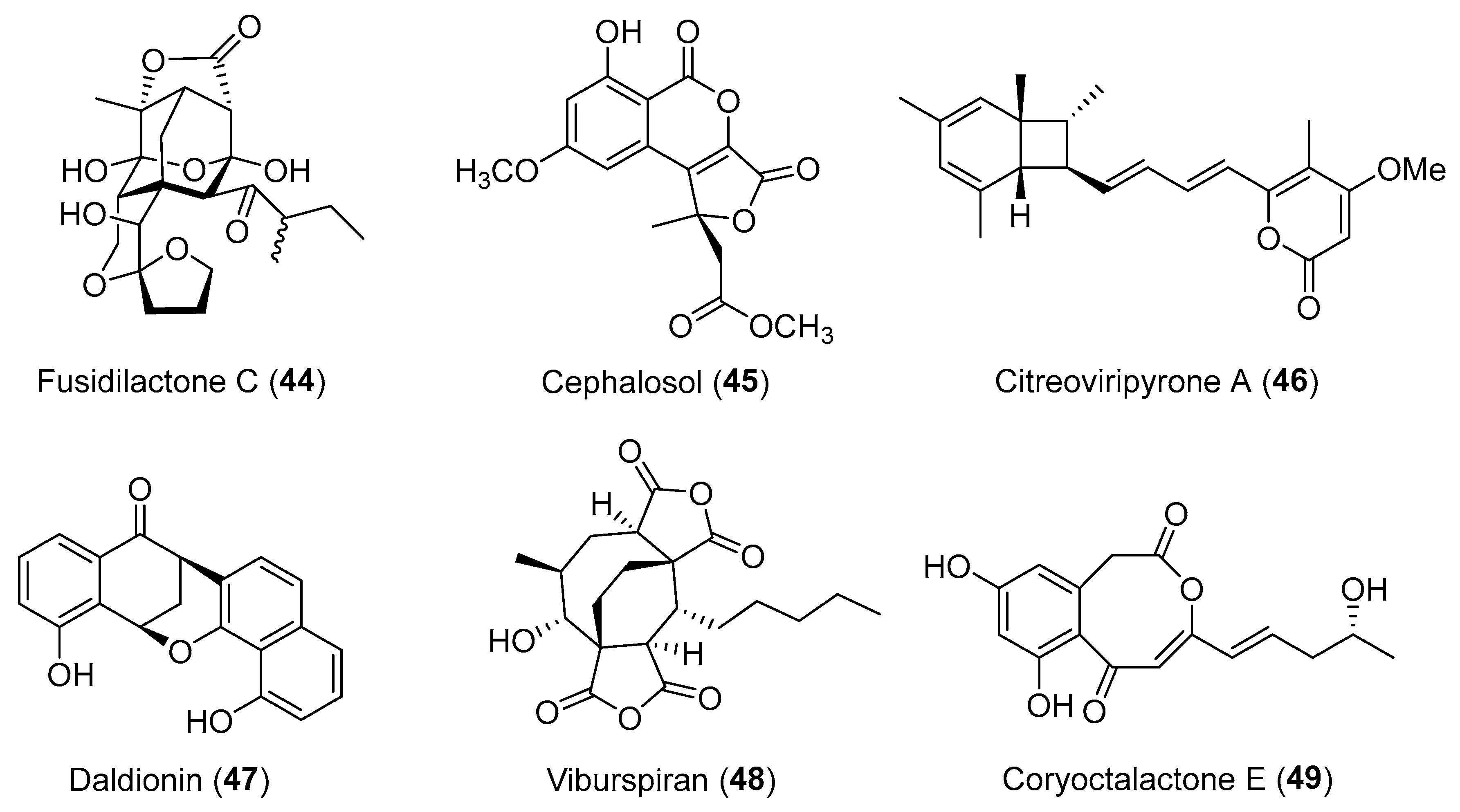{kind=link}
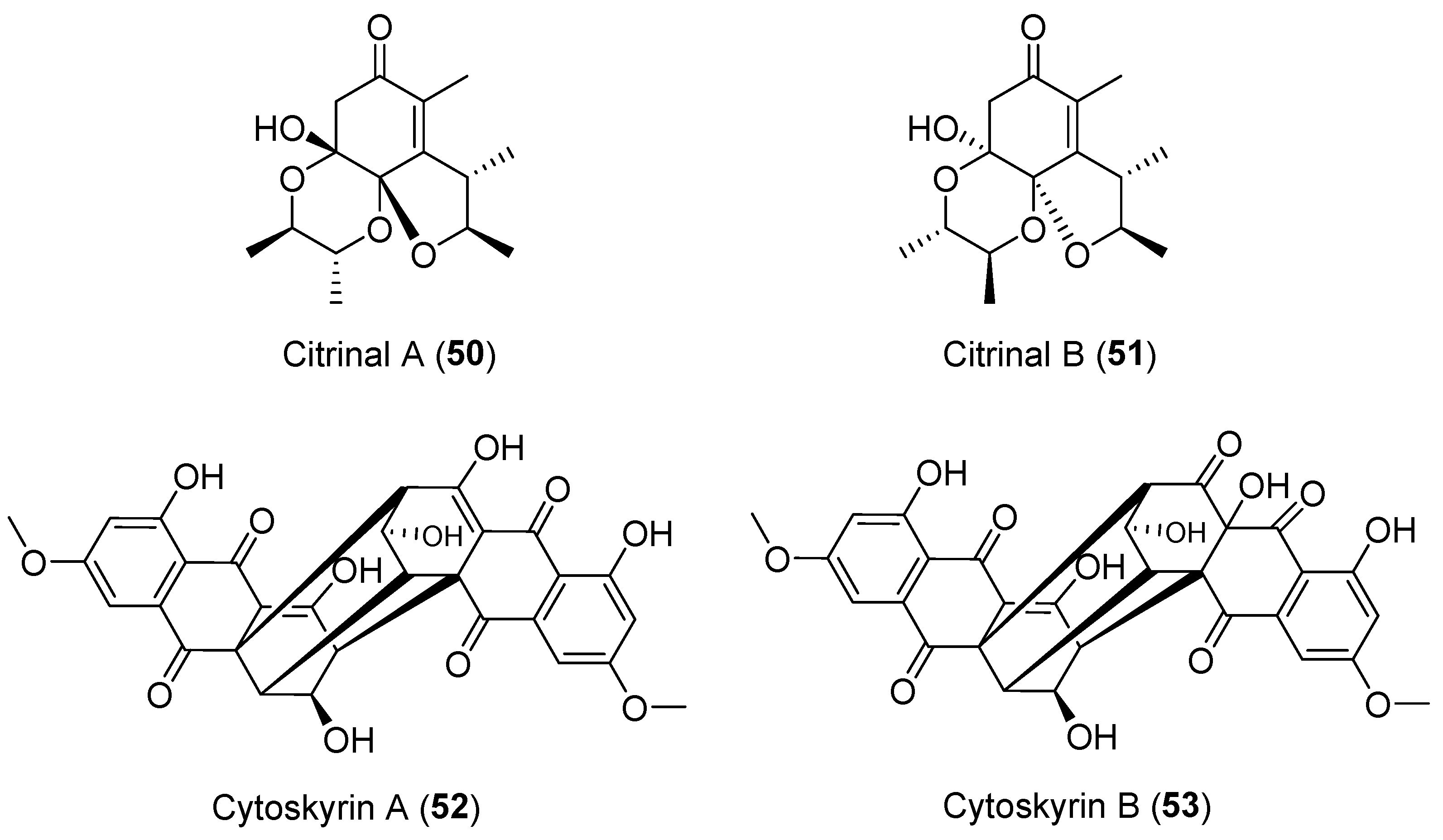{kind=link}
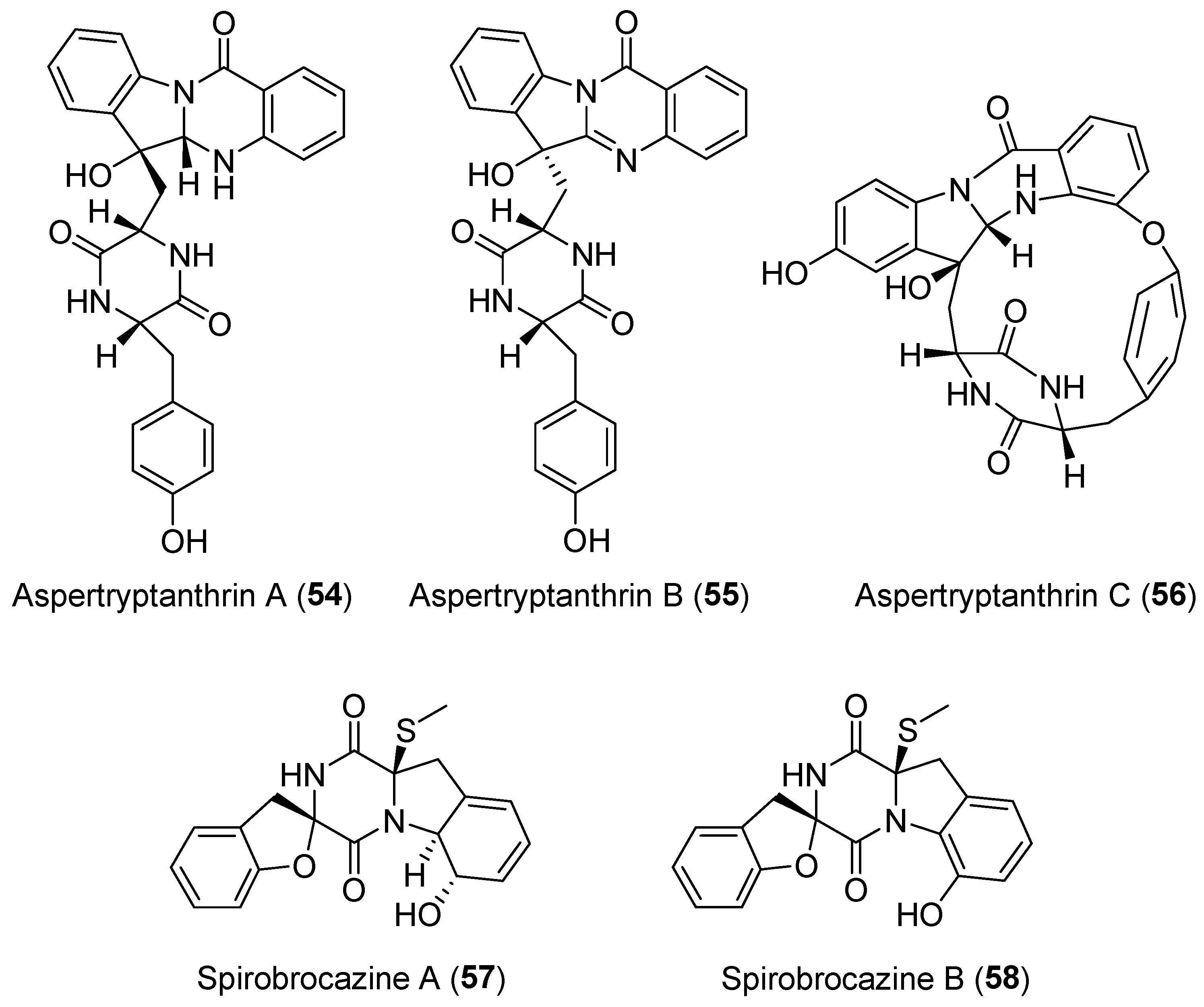{kind=link}
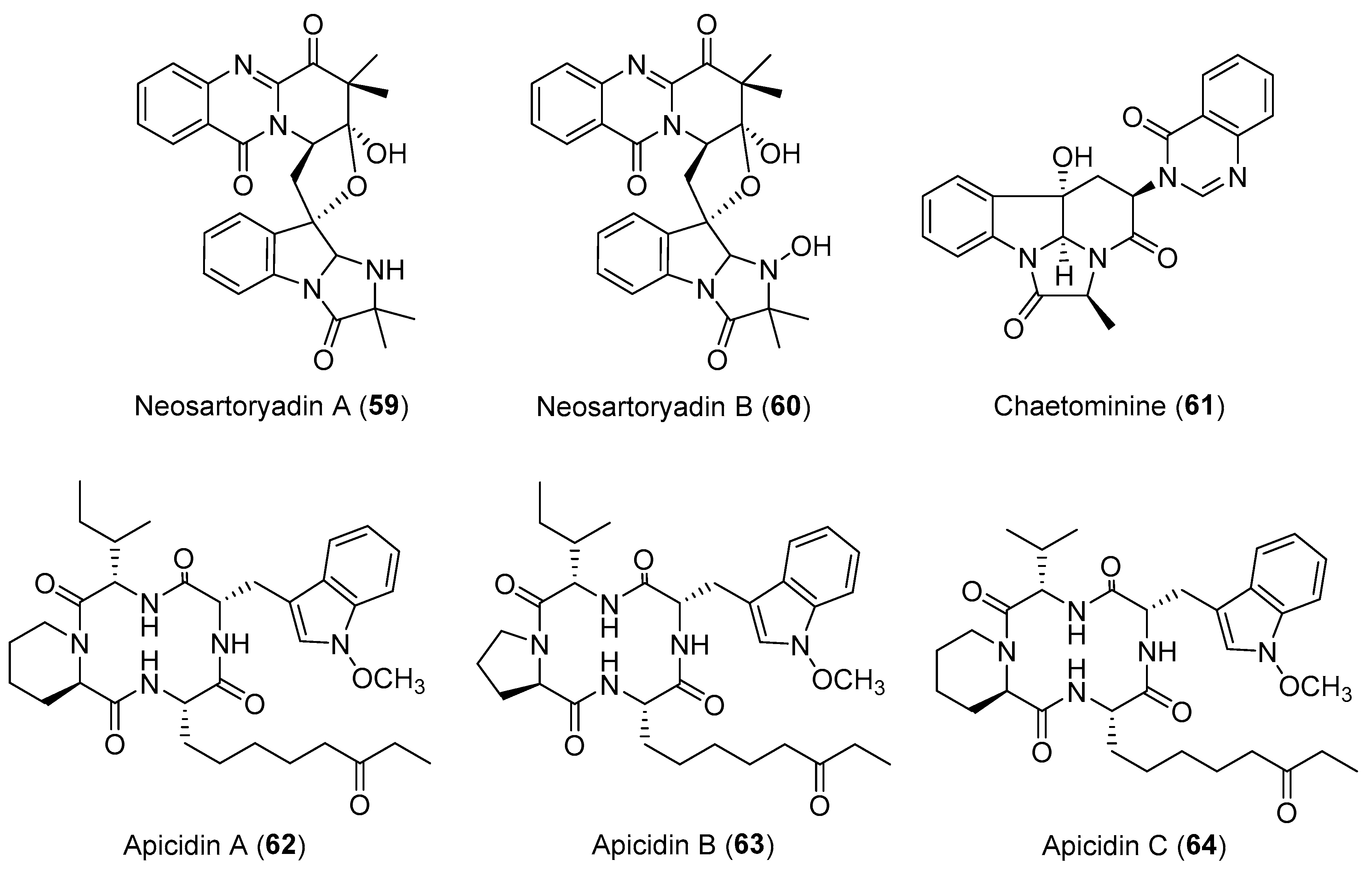{kind=link}
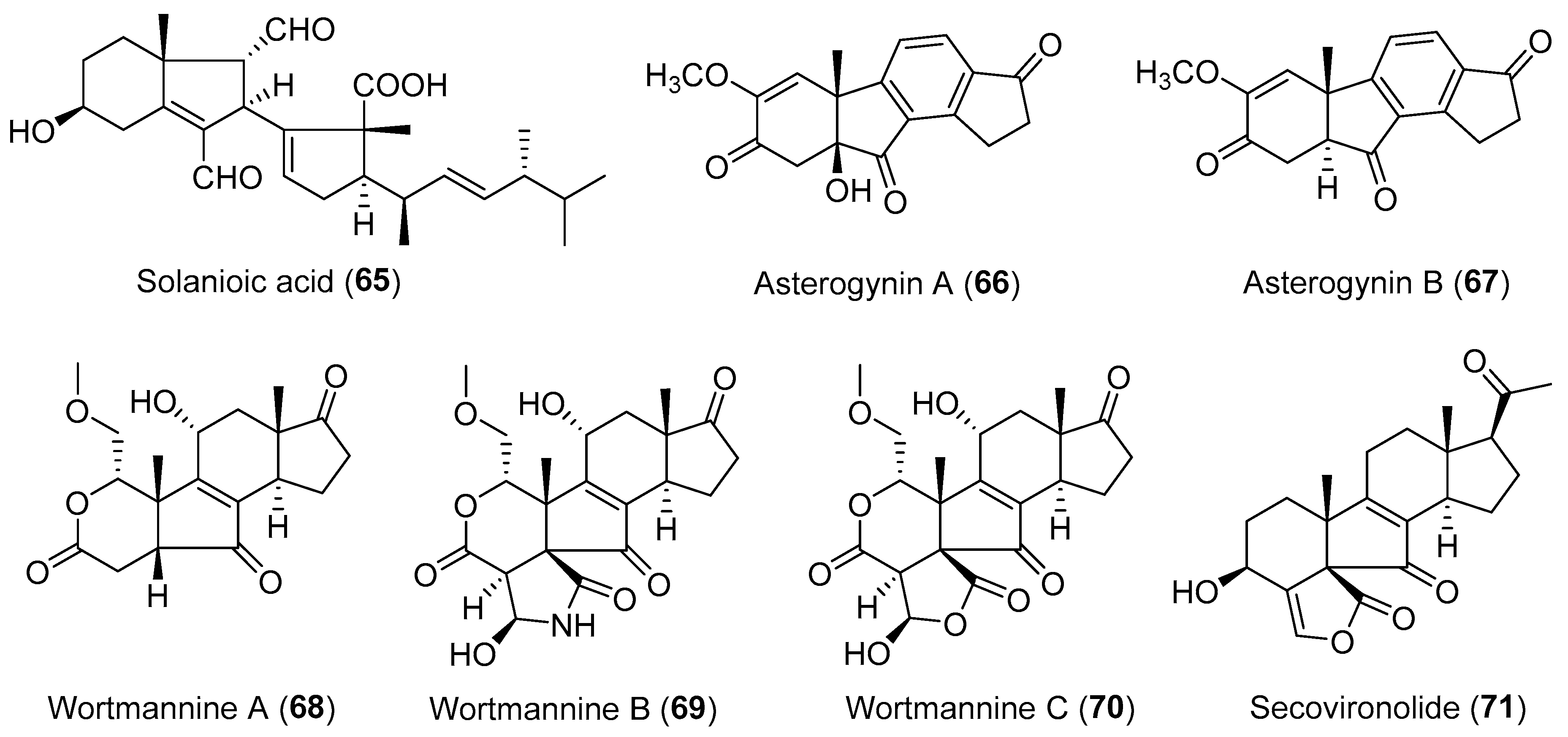{kind=link}
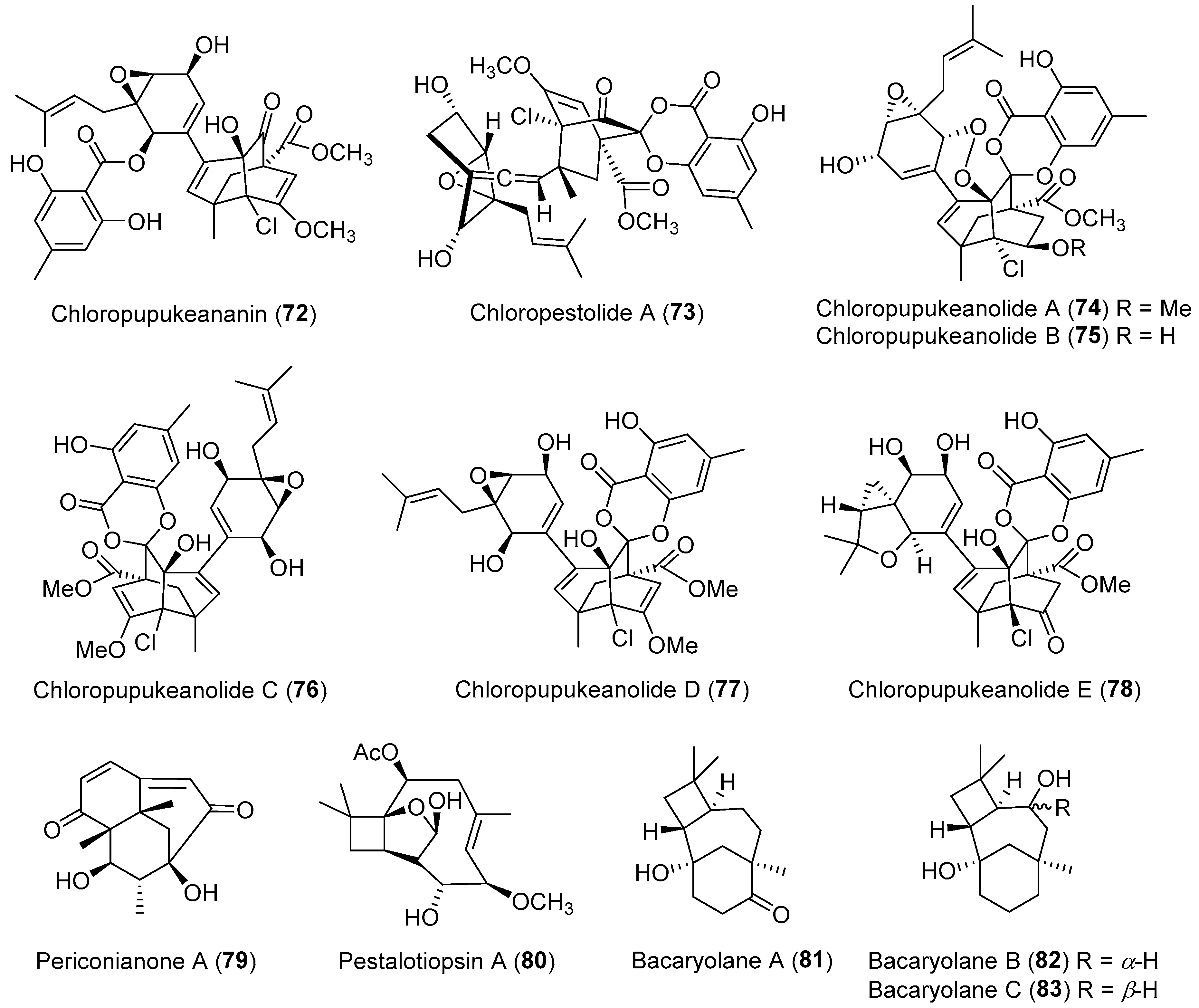{kind=link}
{kind=link}
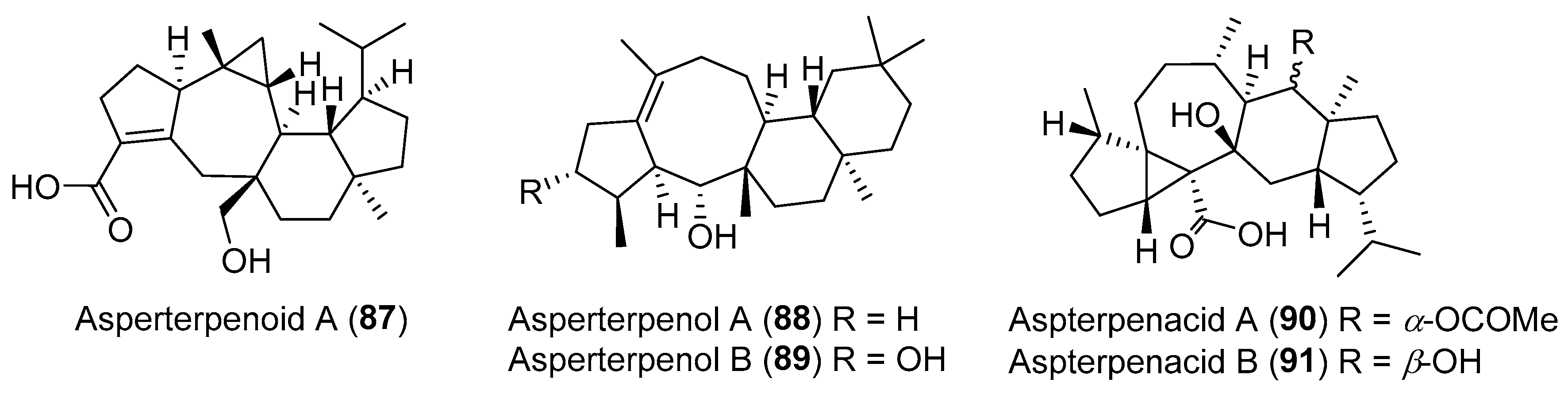{kind=link}
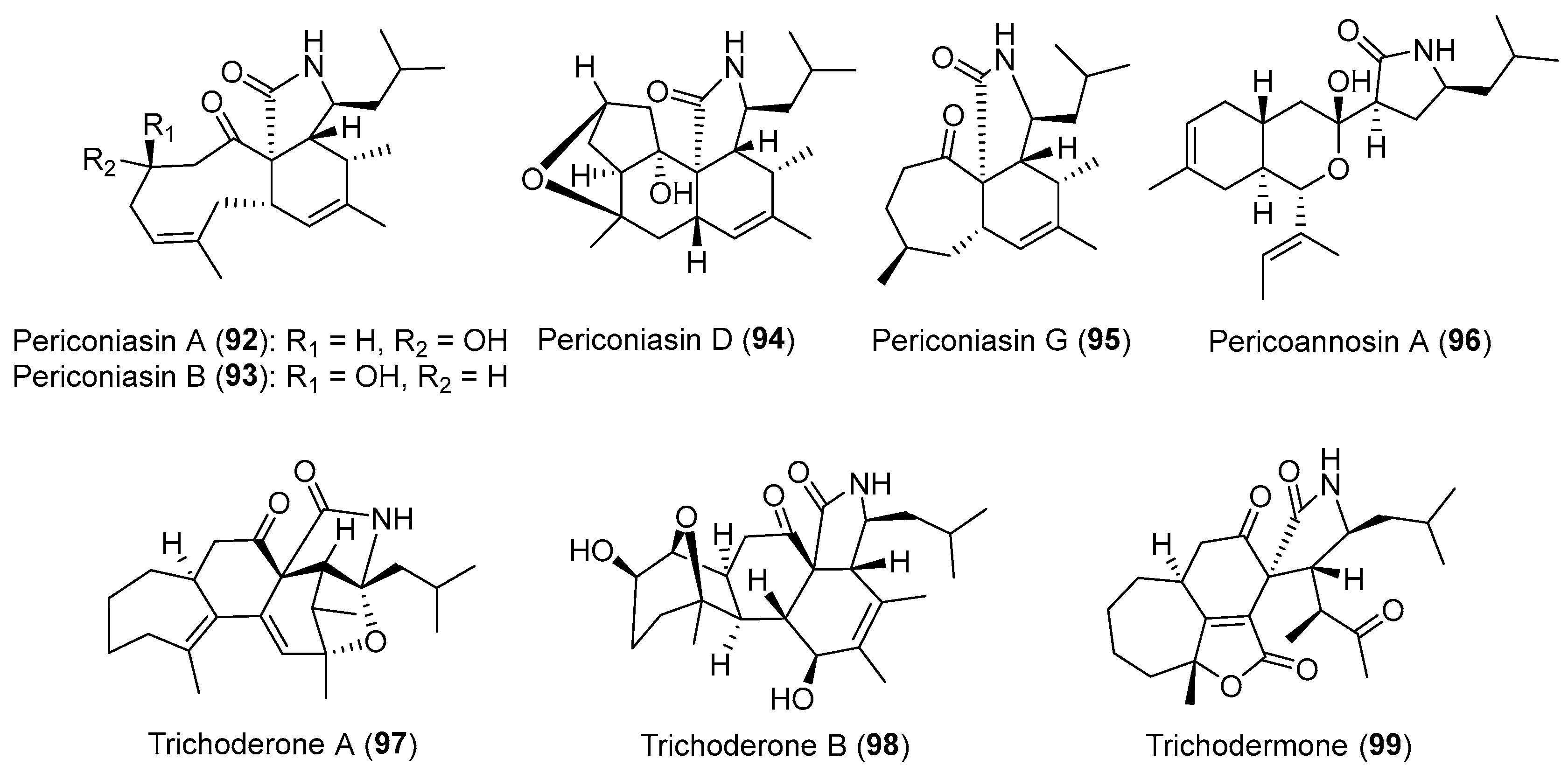{kind=link}
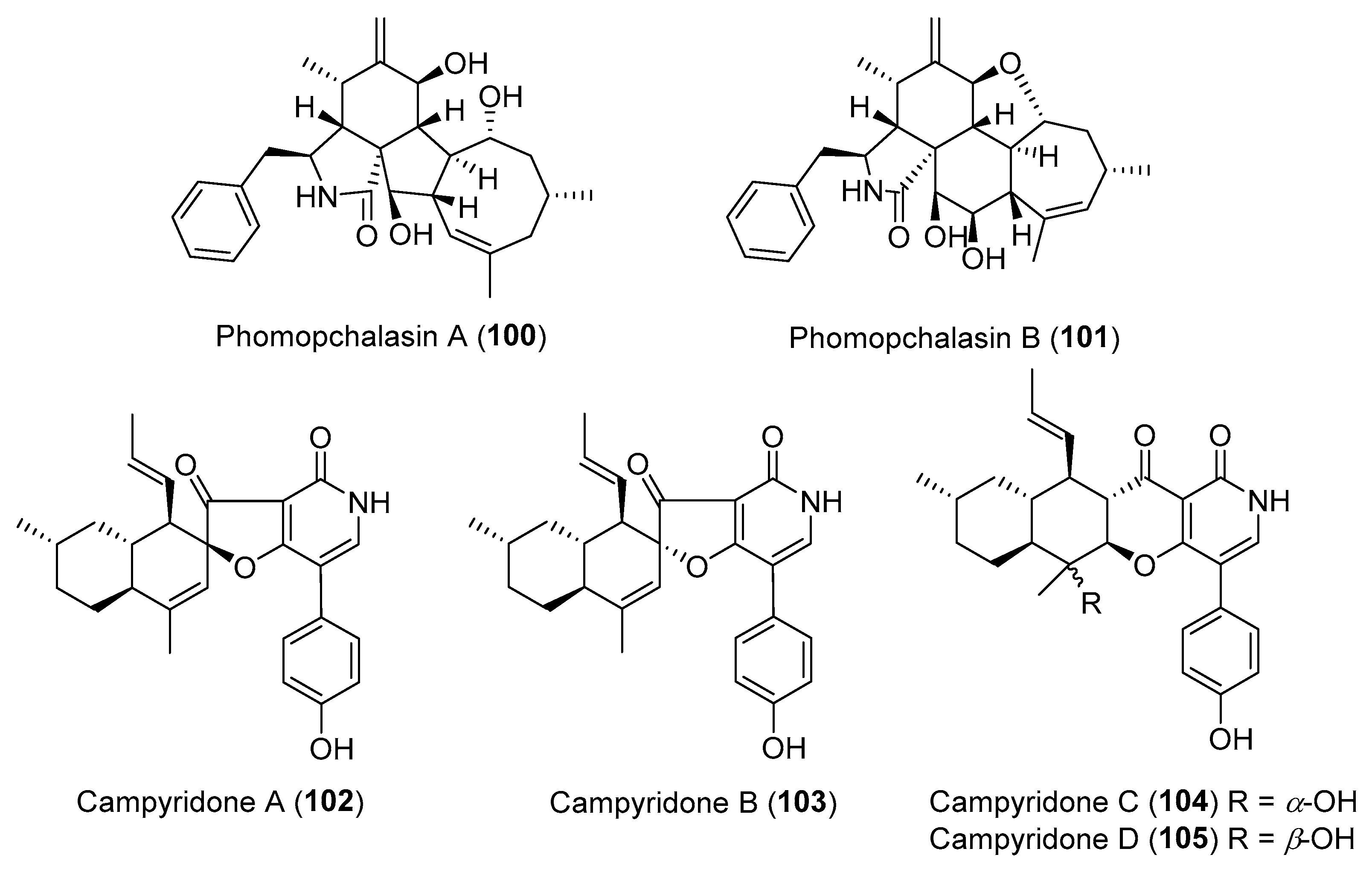{kind=link}
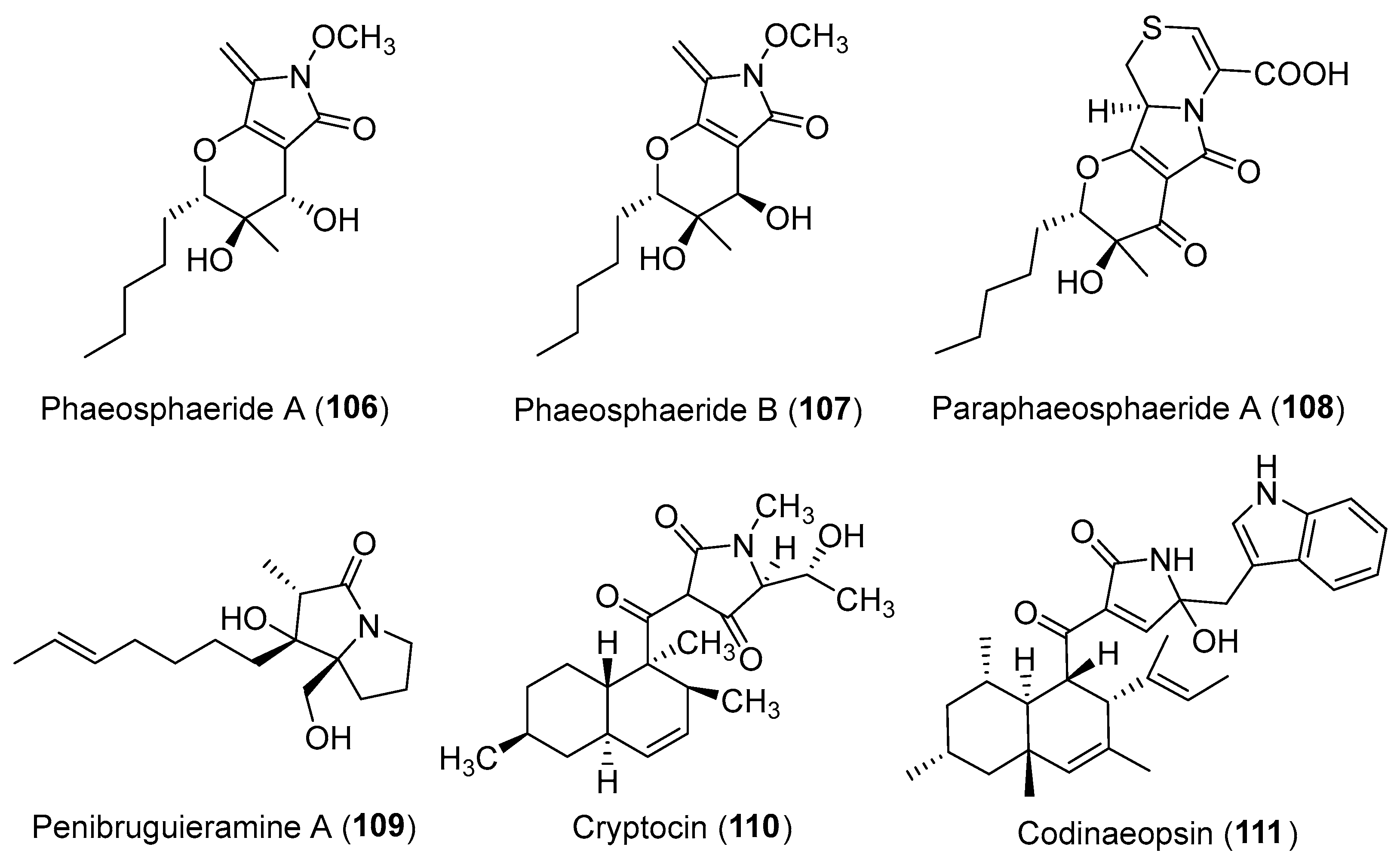{kind=link}
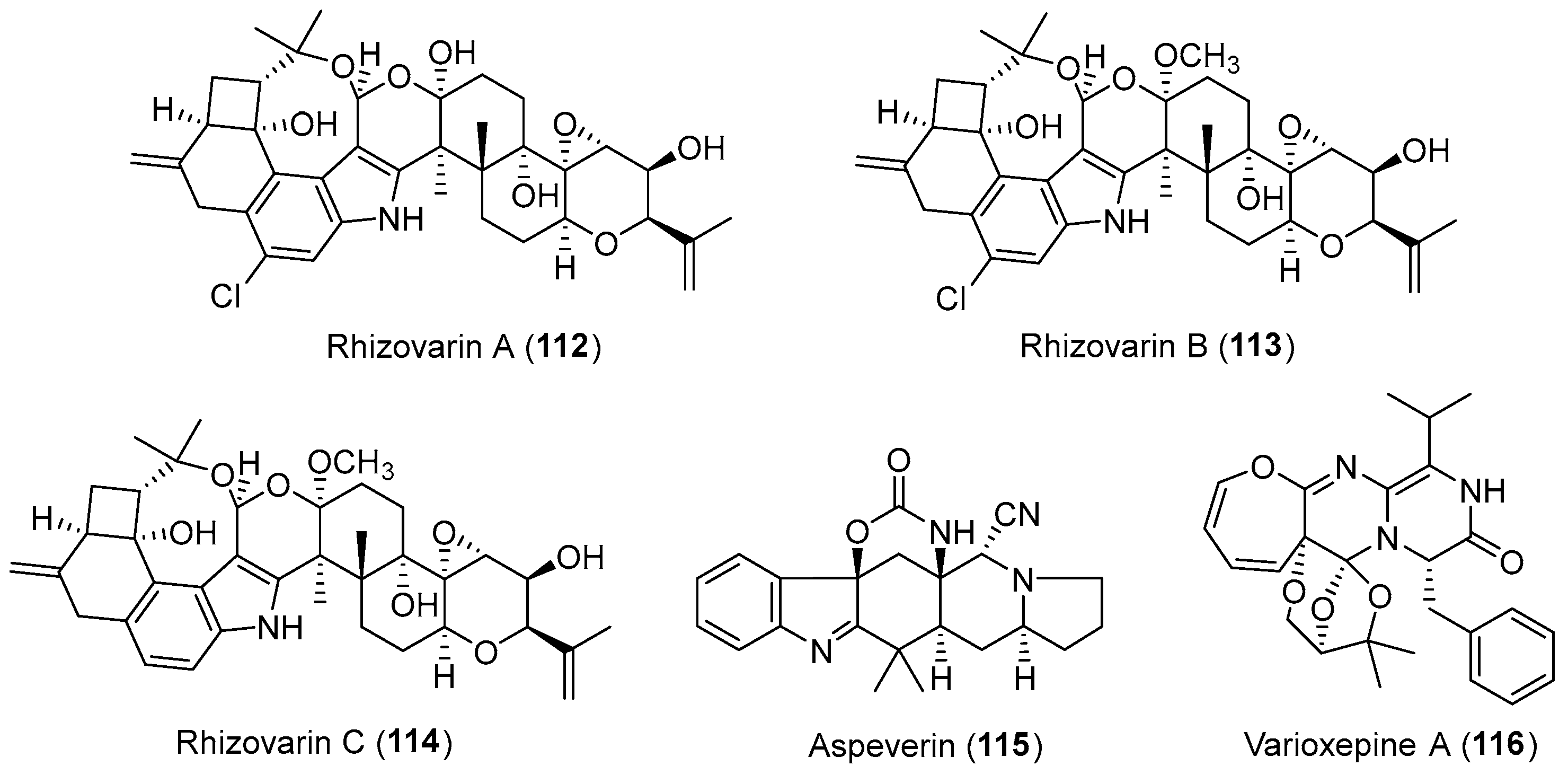{kind=link}
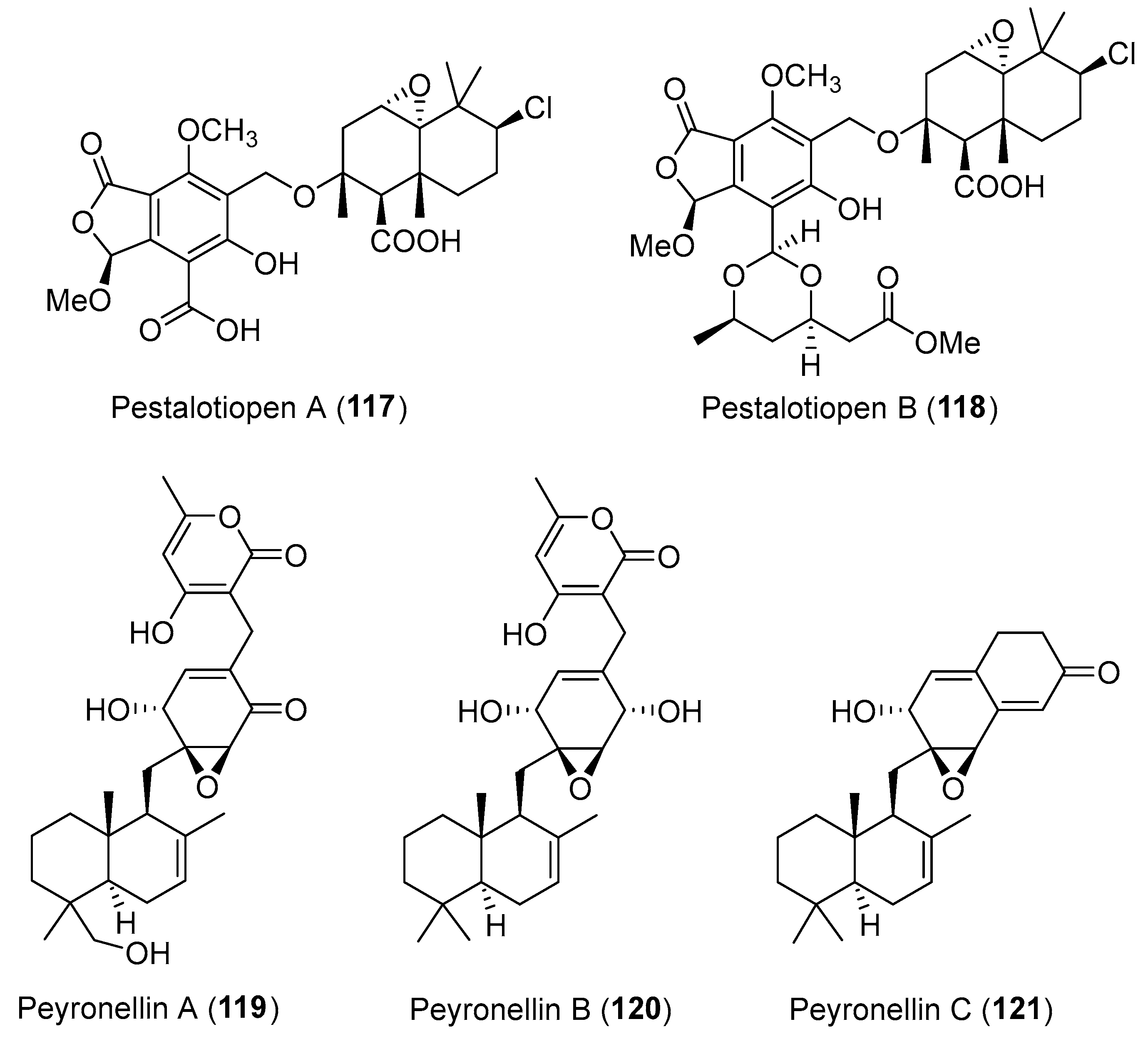{kind=link}
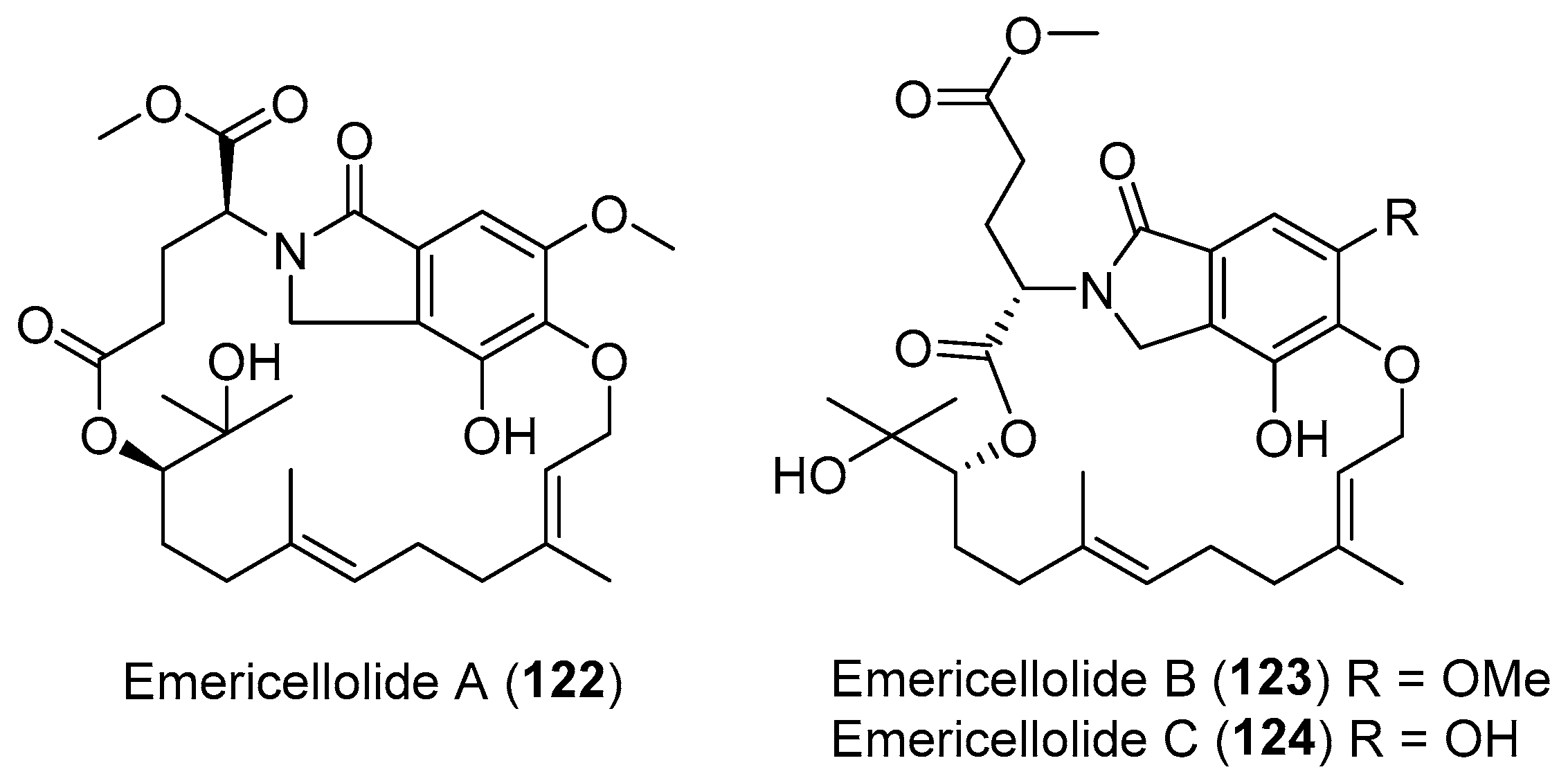{kind=link}
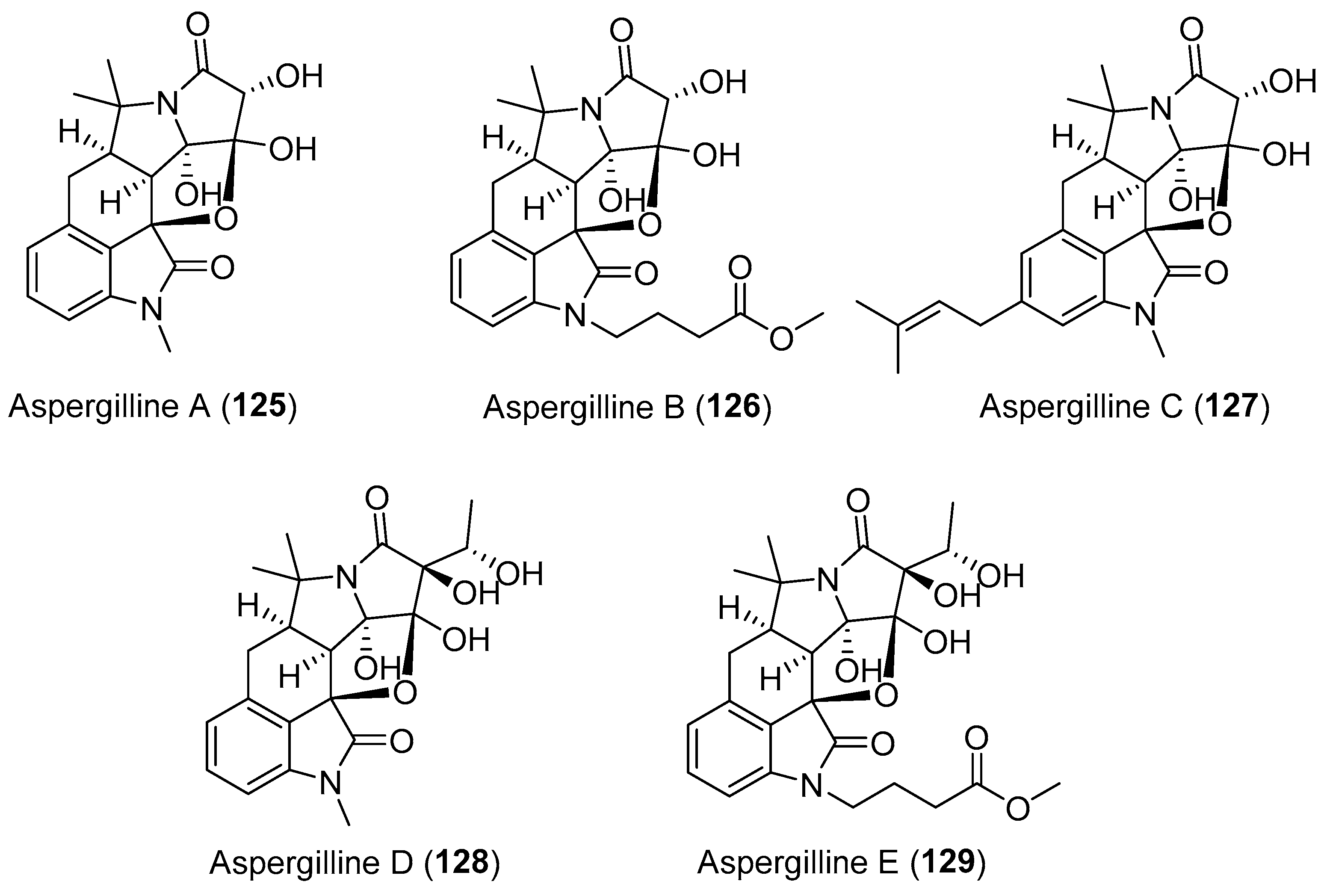{kind=link}
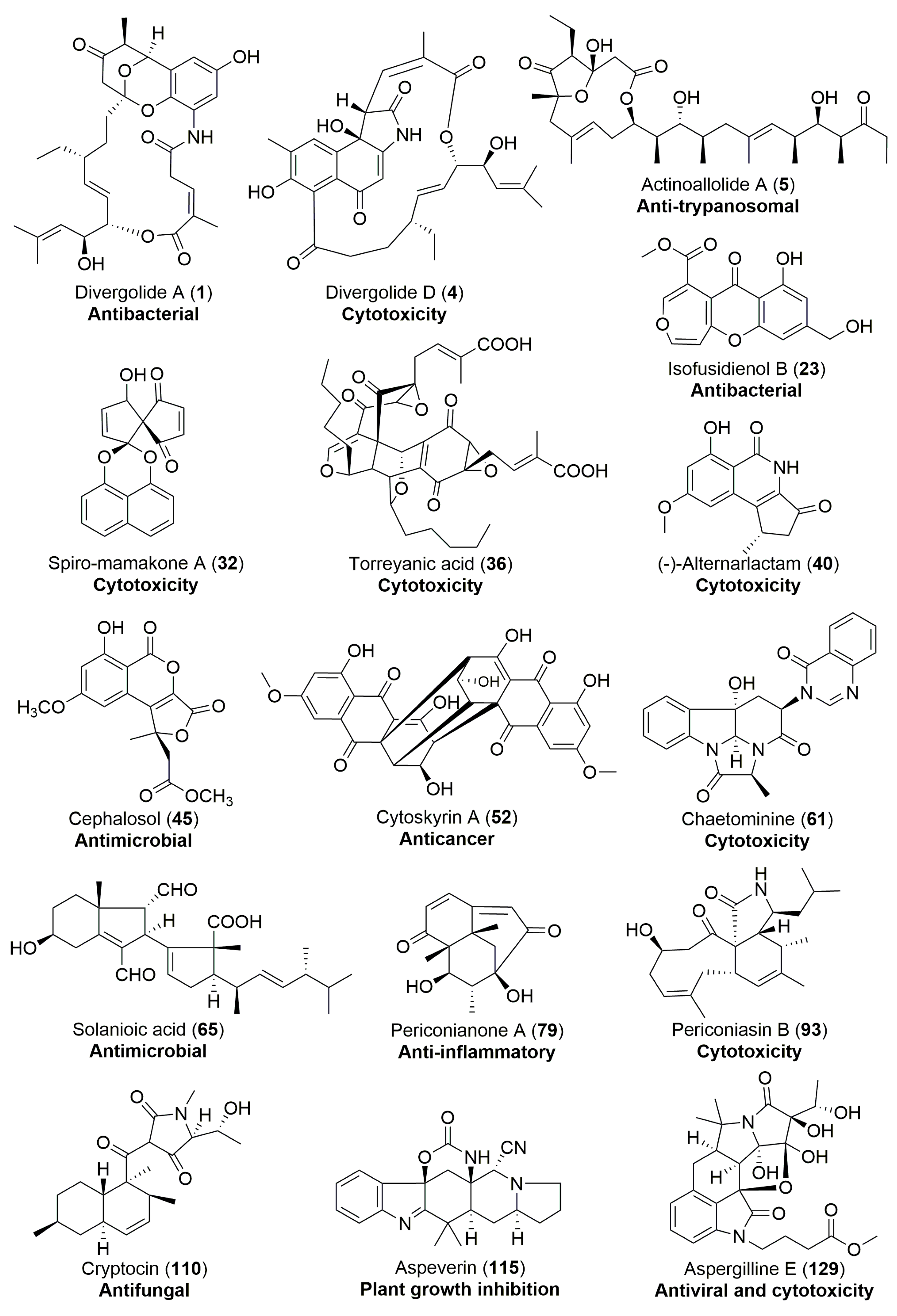{kind=link}
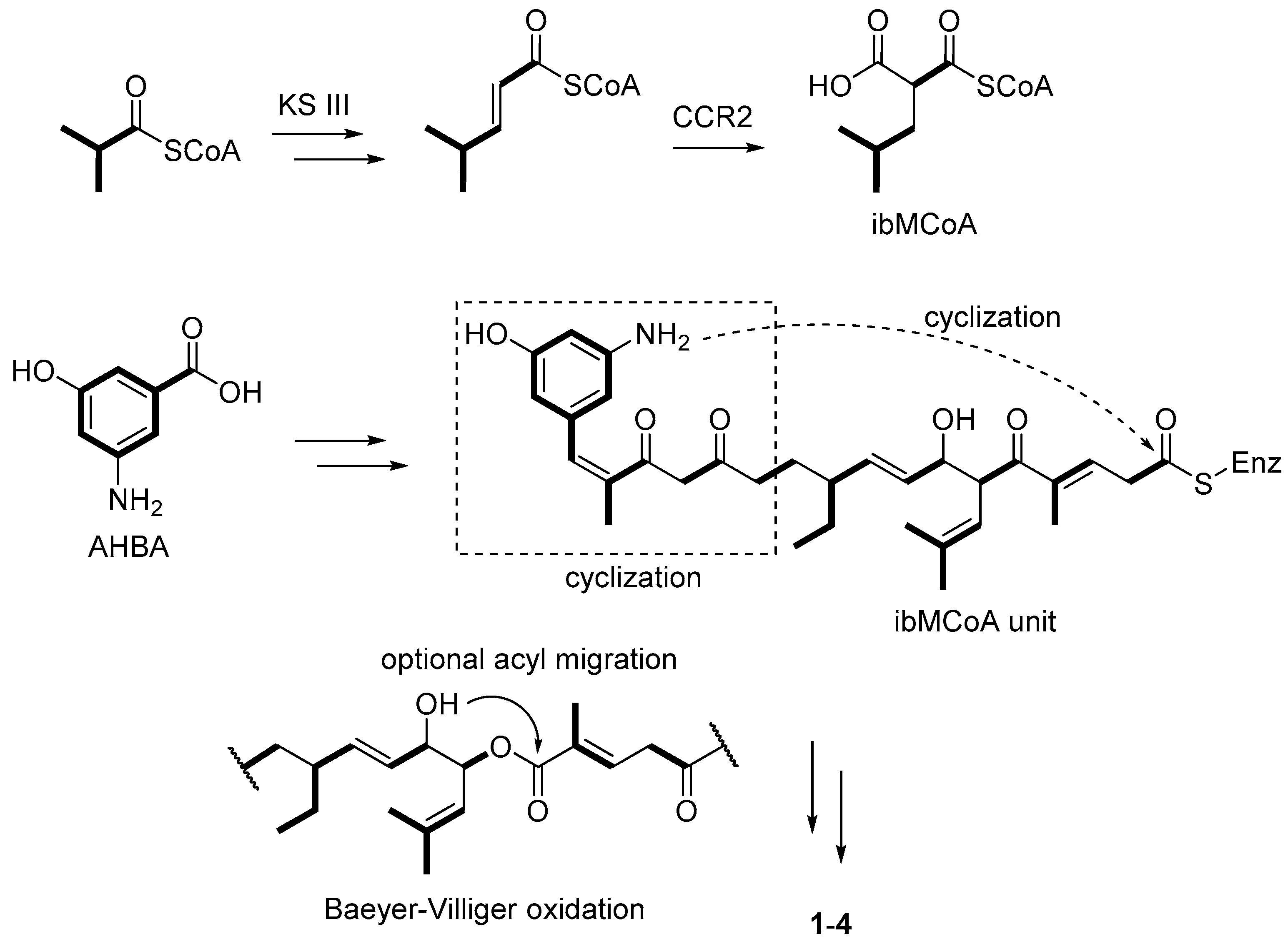{kind=link}
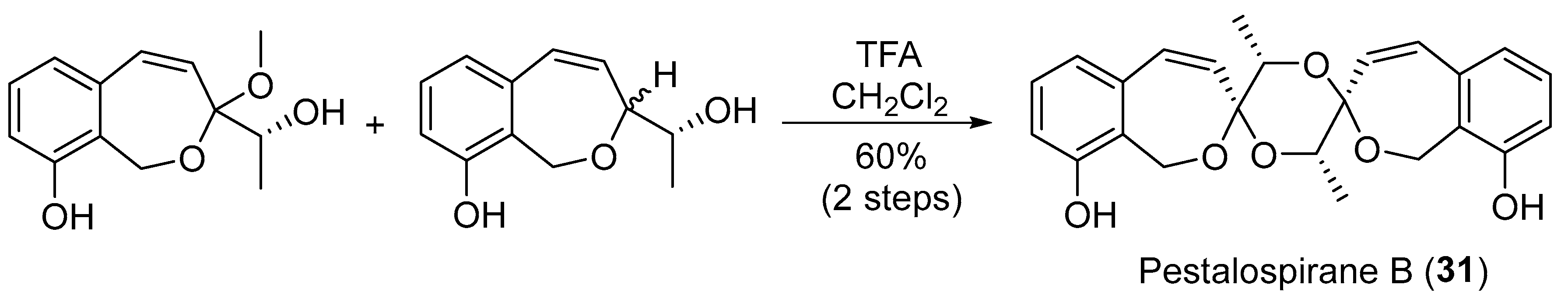{kind=link}
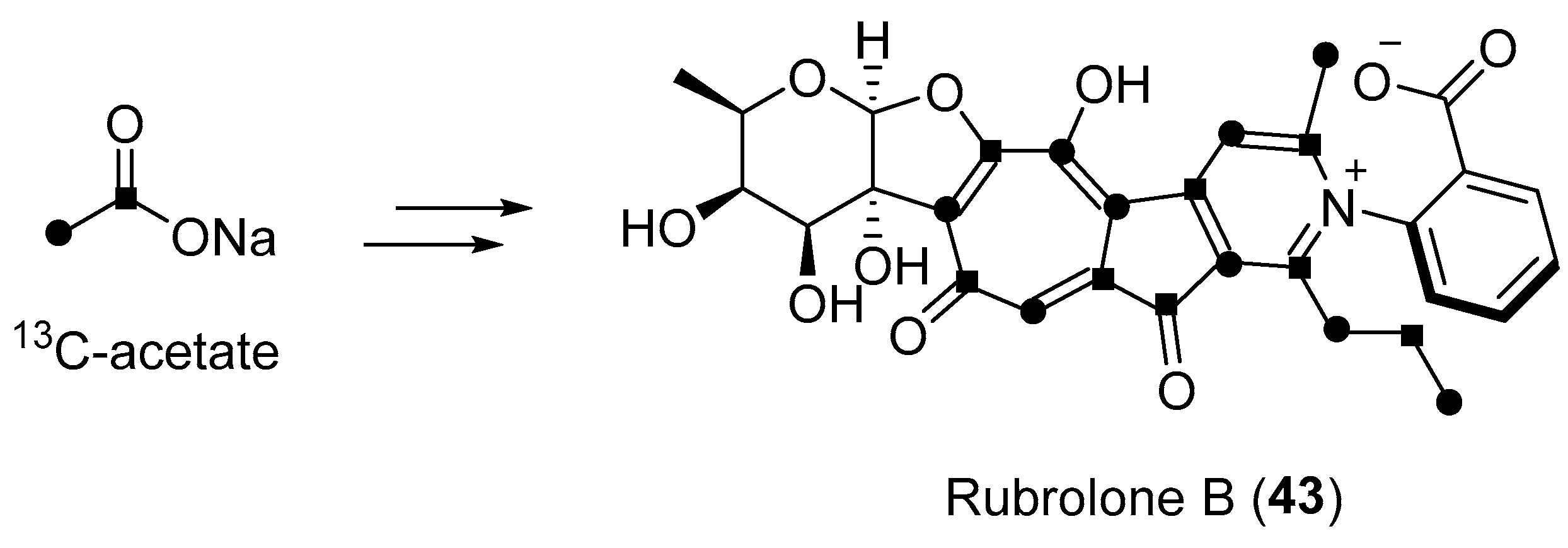{kind=link}
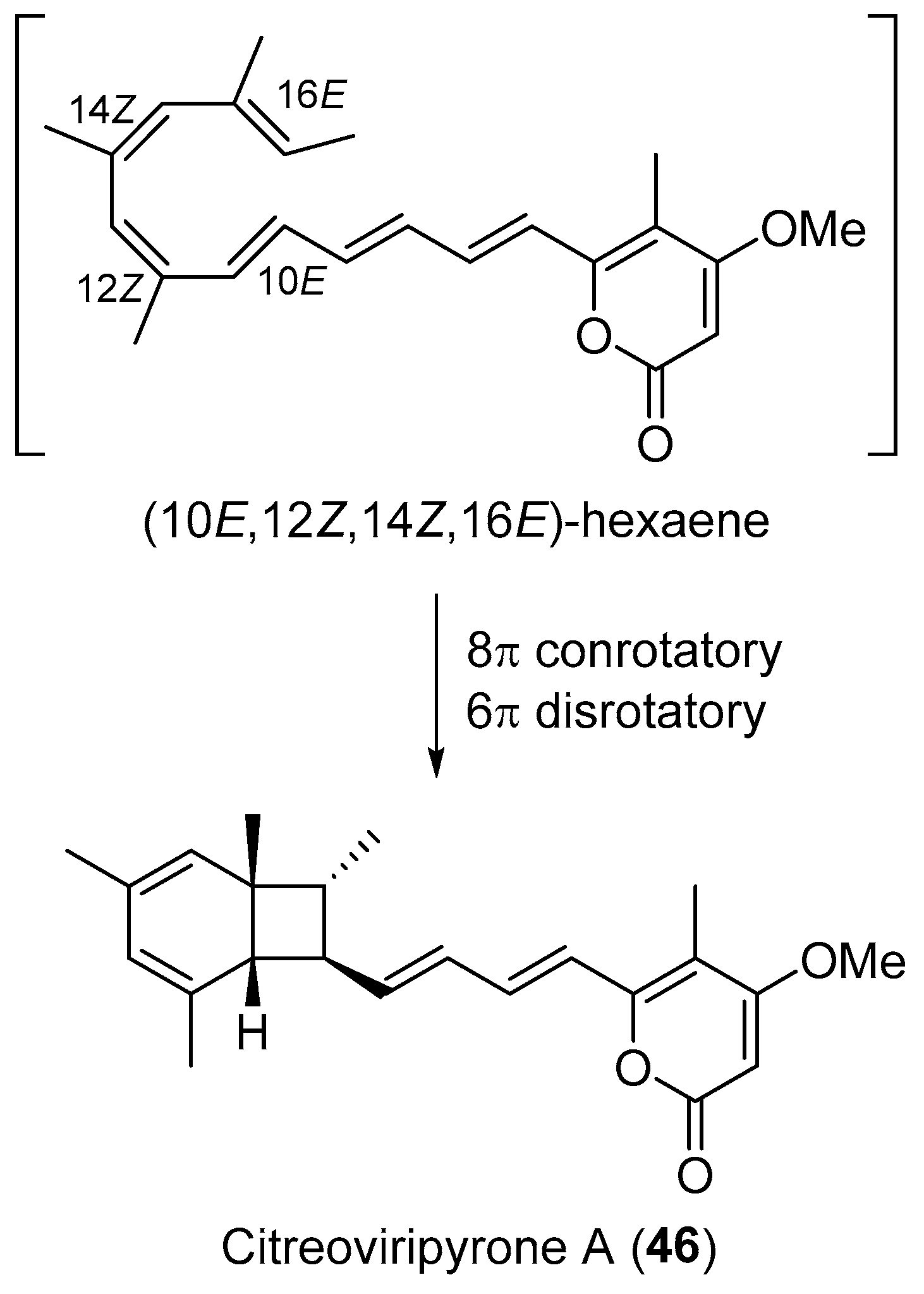{kind=link}
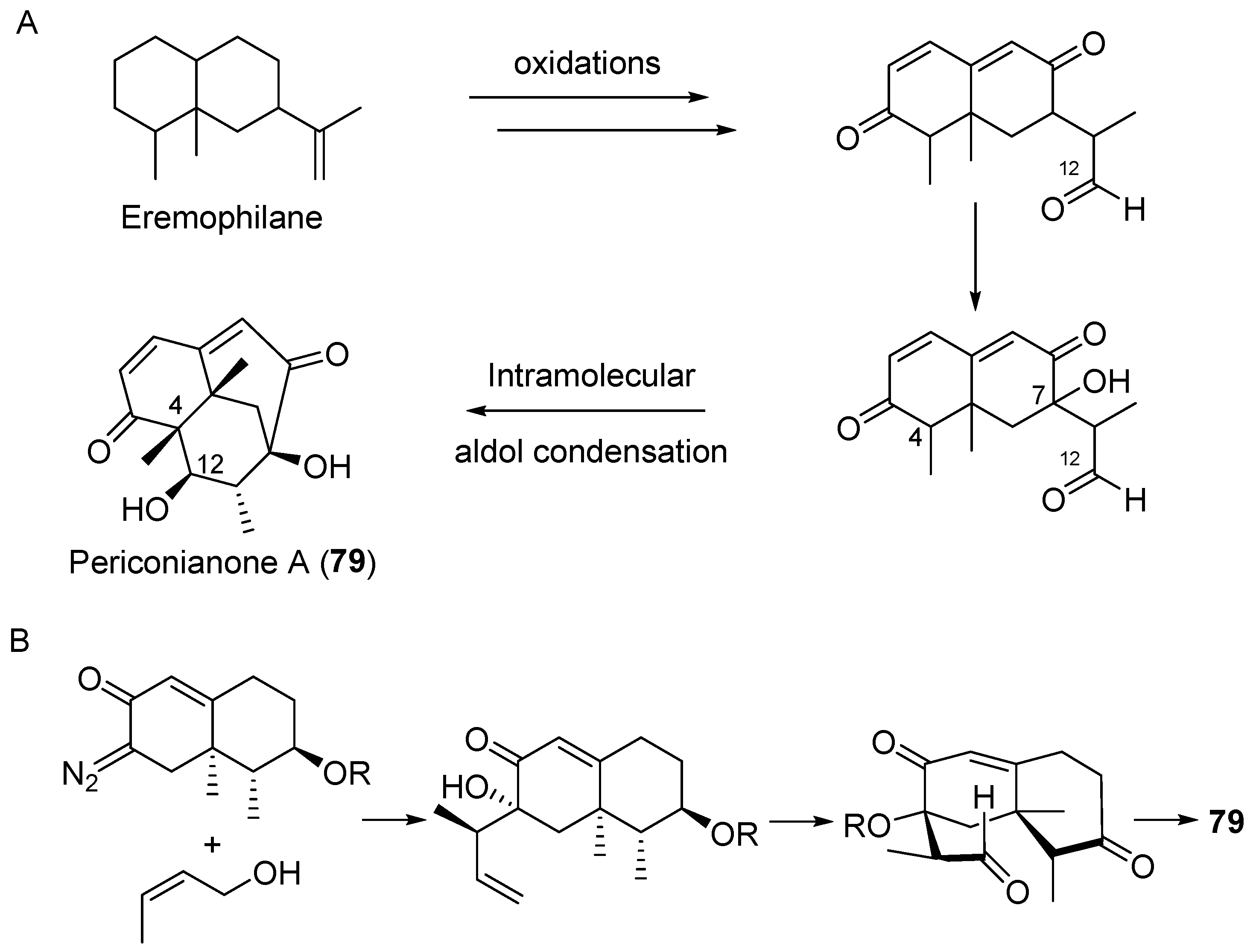{kind=link}
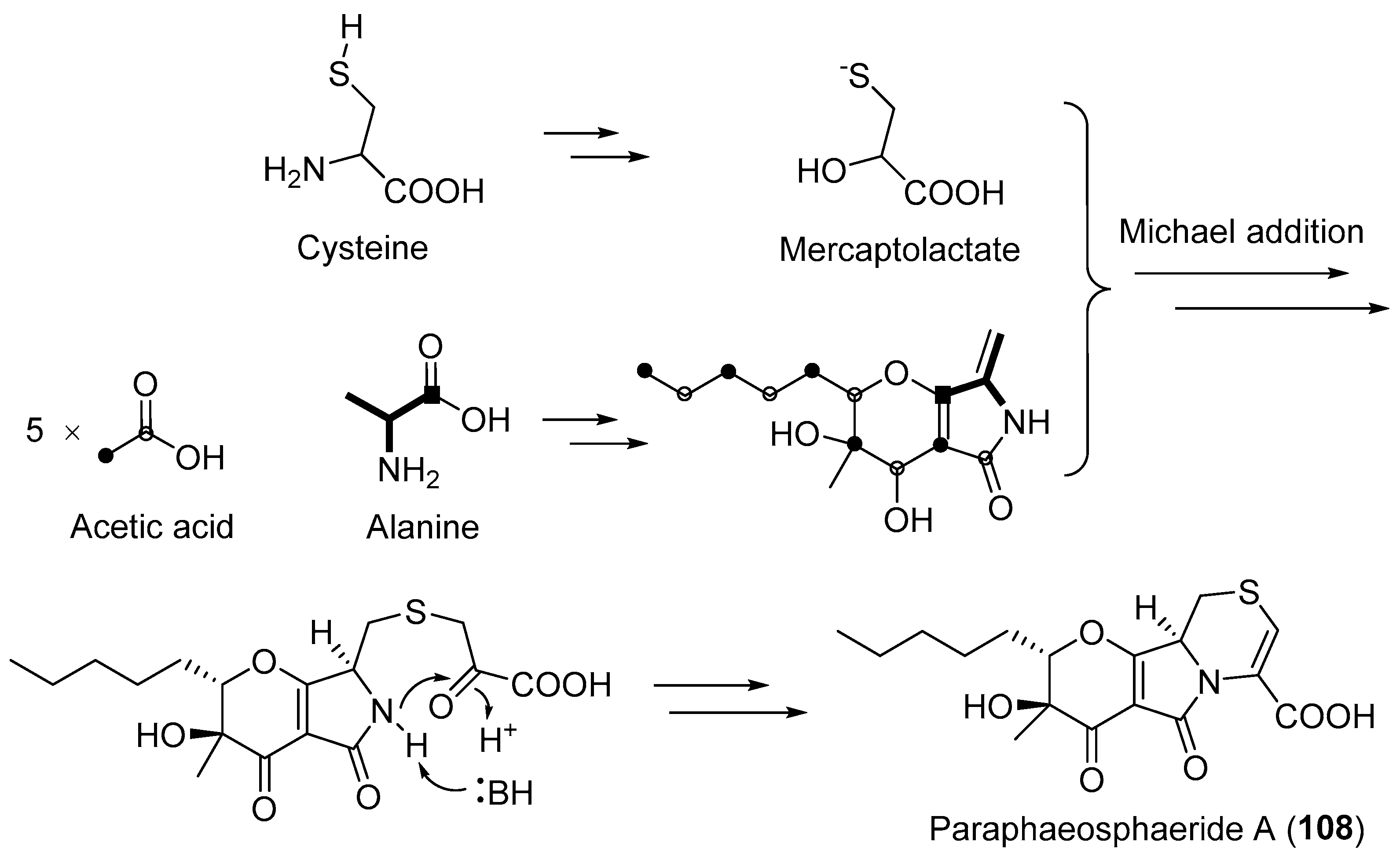{kind=link}
Table 1.
Structurally novel natural products from endophytic bacteria or fungi.
| Microorganisms | Origin | Secondary Metabolites (Figures) | Reported Activities a | Ref. |
|---|---|---|---|---|
| Streptomyces sp. | Marine, Bruguiera gymnorrhiza | Divergolides A–D (1–4) (Figure 1) | Antibacterial, cytotoxicity | [28] |
| Streptomyces sp. | Terrestrial, Camellia sinensis | Rubrolone B (43) (Figure 7) | NR b | [61] |
| Streptomyces sp. | Marine, Bruguiera gymnorrhiza | Bacaryolanes A–C (81–83) (Figure 13) | NR | [108] |
| Chalara sp. | Terrestrial, Artemisia vulgaris | Isofusidienols A–D (22–25) (Figure 4) | Antibacterial | [39] |
| A nonsporulating fungus | Terrestrial, Knightia excelsa | Spiro-mamakone A (32) (Figure 5) | Antibacterial, cytotoxicity | [45] |
| Penicillium brocae | Marine | Spirobrocazines A–B (57–58) (Figure 10) | Antibacterial, cytotoxicity | [83] |
| Pestalotiopsis fici | Terrestrial | Chloropupukeananin (72) (Figure 13) | Antibacterial, antiviral | [100] |
| Unidentified fungus | Terrestrial, Daphnopsis americana | Guanacastepene A (84) (Figure 14) | Antibacterial | [110,111] |
| Blennoria sp. | Terrestrial, Carpobrotus edulis | Blennolides A–G (11–17) (Figure 3) | Antifungal, antialgal | [36] |
| Cryptosporiopsis sp. | Terrestrial, Viburnum tinus | Viburspiran (48) (Figure 8) | Antifungal | [74] |
| Cryptosporiopsis cf. quercina | Terrestrial, Triptergyium wilfordii | Cryptocin (110) (Figure 18) | Antifungal | [140] |
| Cephalosporium acremonium | Terrestrial, Trachelospermum jasminoides | Cephalosol (45) (Figure 8) | Antimicrobial | [68] |
| Daldinia eschscholtzii | Terrestrial, Paphiopedilum exul | Daldionin (47) (Figure 8) | Antimicrobial | [73] |
| Rhizoctonia solani | Terrestrial, Cyperus rotundus | Solanioic acid (65) (Figure 12) | Antimicrobial | [94] |
| Trichoderma spp. | Marine, alga Codium fragile | Harziandione (85) and harzianone (86) (Figure 14) | Antimicrobial | [115] |
| Paecilomyces variotii | Marine | Varioxepine A (116) (Figure 19) | Antimicrobial | [150] |
| Pestalotiopsis sp. | Marine, Rhizophora mucronata | Pestalotiopens A–B (117–118) (Figure 20) | Antimicrobial | [152] |
| Pestalotiopsis fici | Terrestrial | Chloropestolide A (73) (Figure 13) | Anti-HIV, cytotoxicity | [102] |
| Periconia sp. | Terrestrial, Annona muricata | Pericoannosin A (96) (Figure 16) | Anti-HIV | [122] |
| Neosartorya udagawae | Marine, Aricennia marina | Neosartoryadins A–B (59–60) (Figure 11) | Antiviral | [84] |
| Periconia sp. | Terrestrial, Annona muricata | periconiasins (92–95) (Figure 16) | Antiviral, cytotoxicity | [120,121,123] |
| Aspergillus versicolor | Terrestrial, polyphylla var. yunnanensis | Aspergillines A–E (125–129) (Figure 22) | Antiviral, cytotoxicity | [157] |
| Periconia sp. | Terrestrial, Annonsa muricata | Periconianone A (79) (Figure 13) | Anti-inflammatory | [105] |
| Unidentified fungus | Terrestrial, Vochysia guatemalensis | Codinaeopsin (111) (Figure 18) | Antimalarial | [143] |
| Phomopsis sp. | Terrestrial, Isodon eriocalyx var. laxiflora | Phomopchalasins A–B (100–101) (Figure 17) | Antimigratory activity | [128] |
| Fusarium pallidoroseum | Terrestrial | Apicidins A–C (62–64) (Figure 11) | Antiprotozoal, anticancer | [90] |
| Actinoallomurus fulvus | Terrestrial, Capsicum frutescens | Actinoallolides A–E (5–9) (Figure 2) | Anti-trypanosomal | [34] |
| Aspergillus sp. | Marine | Asperterpenoid A (87) (Figure 15) | Antituberculosis | [116] |
| Aspergillus sp. | Marine | Asperterpenols A–B (88–89) (Figure 15) | Acetylcholinesterase inhibition | [118] |
| Cytospora sp. | Terrestrial, Conocarpus erecta | Cytoskyrins A–B (52–53) (Figure 9) | BIA activity | [78] |
| Unidentified fungus | Terrestrial, Ficus microcarpa L. | Microcarpalide (10) (Figure 2) | Cytotoxicity | [35] |
| Nodulisporium sp. | Marine, Alga | Noduliprevenone (18) (Figure 3) | Cytotoxicity | [37] |
| Pestalotiopsis virgatula | Terrestrial, Dracontomelon duperreanum | Virgatolides A–C (27–29) (Figure 5) | Cytotoxicity | [41] |
| Pestalotiopsis microspora | Terrestrial, Torreya taxifolia | (±)-torreyanic acid (36) (Figure 6) | Cytotoxicity | [50] |
| Chaetomium globosum | Terrestrial, Imperata cylindrical | Chaetoglobins A–B (38–39) (Figure 7) | Cytotoxicity | [56] |
| Alternaria sp. | Terrestrial, Carex aridula | (-)-Alternarlactam (40) (Figure 7) | Cytotoxicity | [57] |
| Penicillium manginii | Terrestrial, Panax notoginseng | Duclauxamide A1 (42) (Figure 7) | Cytotoxicity | [59] |
| Penicillium sp. | Terrestrial, Catharanthus roseus | Citreoviripyrone A (46) (Figure 8) | Cytotoxicity | [71] |
| Chaetomium sp. | Terrestrial, Adenophora axilliflora | Chaetominine (61) (Figure 11) | Cytotoxicity | [86] |
| Pestalotiopsis fici | Terrestrial | Chloropupukeanolides A–E (74–78) (Figure 13) | Cytotoxicity | [103,104] |
| Trichoderma gamsii | Terrestrial, Panax notoginseng | Trichoderones A–B (97–98) (Figure 16) | Cytotoxicity | [127] |
| Trichoderma gamsii | Terrestrial, Panax notoginseng | Trichodermone (99) (Figure 16) | Cytotoxicity | [126] |
| Campylocarpon sp. | Marine, Sonneratia caseolaris | Campyridones A–D (102–105) (Figure 17) | Cytotoxicity | [129] |
| Mucor irregularis | Marine, Rhizophora stylosa | Rhizovarins A–C (112–114) (Figure 19) | Cytotoxicity | [145] |
| Peyronellaea coffeae-arabicae | Terrestrial, Pritchardia lowreyana | Peyronellins A–C (119–121) (Figure 20) | Cytotoxicity | [153] |
| Pestalotiopsis sp. | Terrestrial, Taxus brevifolia | Pestalotiopsin A (80) (Figure 13) | Immunosuppressive | [107] |
| Penicillium dangeardii | Terrestrial, Lysidice rhodostegia | Penicillactones A–C (33–35) (Figure 5) | Inhibitors of the release of β-glucuronidase | [48] |
| Phaeosphaeria avenaria | Terrestrial | Phaeosphaeride A–B (106–107) (Figure 18) | Inhibiting STAT3 activity | [130] |
| Aspergillus versicolor | Marine, green alga Codium fragile | Aspeverin (115) (Figure 19) | Marine plant growth inhibition | [146] |
| Microsphaeropsis sp. | Terrestrial, Lycium intricatum | Microsphaeropsones A–C (19–21) (Figure 4) | NR | [38] |
| Paraphaeosphaeria neglecta | Terrestrial, Lycopodiella cernua | Lycopodiellactone (26) (Figure 4) | NR | [40] |
| Pestalotiopsis virgatula | Terrestrial, Terminalia chebula | Pestalospiranes A–B (30–31) (Figure 5) | NR | [43] |
| Pestalotiopsis sp. | Terrestrial, Clavaroids sp. | Torreyanic acid analogue (37) (Figure 6) | NR | [53] |
| Fusarium sp. | Terrestrial, Melia azedarach | Fusarimine (41) (Figure 7) | NR | [58] |
| Fusidium sp. | Terrestrial, Mentha arvensis | Fusidilactone C (44) (Figure 8) | NR | [65,66] |
| Corynespora cassiicola | Marine, Laguncularia racemosa | Coryoctalactone E (49) (Figure 8) | NR | [75] |
| Colletotrichum capsici | Terrestrial, Siegesbeckia pubescens | Citrinals A–B (50–51) (Figure 9) | NR | [76,77] |
| Aspergillus sp. | Terrestrial, Melia azedarach L. | Aspertryptanthrins A–C (54–56) (Figure 10) | NR | [82] |
| Chalara alabamensis | Terrestrial, Asterogyne martiana | Asterogynins A–B (66–67) (Figure 12) | NR | [96] |
| Talaromyces wortmannii | Terrestrial, Tripterygium wilfordii | Wortmannines A–C (68–70) (Figure 12) | NR | [98] |
| Talaromyces wortmanni | Terrestrial, Tripterygium wilfordii | Secovironolide (71) (Figure 12) | NR | [97] |
| Aspergillus sp. | Marine, Kandelia obovata | Aspterpenacids A–B (90–91) (Figure 15) | NR | [117] |
| Paraphaeosphaeria neglecta | Terrestrial, Lycopodiella cernua | Paraphaeosphaeride A (108) (Figure 18) | NR | [136] |
| Penicillium sp. | Marine, Bruguiera gymnorrhiza | Penibruguieramine A (109) (Figure 18) | NR | [138] |
| Emericella nidulans | Terrestrial, Tamarix chinensis Lour | Emericellolides A–C (122–124) (Figure 21) | NR | [155] |
a Organize the bioactivity using alphabetical order. b NR: not reported in references or have reported in literature to have no biological activities.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gao, H.; Li, G.; Lou, H.-X. Structural Diversity and Biological Activities of Novel Secondary Metabolites from Endophytes. Molecules 2018, 23, 646. https://doi.org/10.3390/molecules23030646
AMA Style
Gao H, Li G, Lou H-X. Structural Diversity and Biological Activities of Novel Secondary Metabolites from Endophytes. Molecules. 2018; 23(3):646. https://doi.org/10.3390/molecules23030646
Chicago/Turabian StyleGao, Han, Gang Li, and Hong-Xiang Lou. 2018. "Structural Diversity and Biological Activities of Novel Secondary Metabolites from Endophytes" Molecules 23, no. 3: 646. https://doi.org/10.3390/molecules23030646