
Cryopreservation of Cyanobacteria and Eukaryotic Microalgae Using Exopolysaccharide Extracted from a Glacier Bacterium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. EPS Yielded by Pseudomonas sp. BGI-2
2.2. Extraction of EPS
2.3. Cyanobacterial and Microalgal Strains Used for Cryopreservation Assay
2.4. Cryopreservation of Cyanobacteria and Microalgae
2.5. Growth Recovery
2.6. Effect of Heat Sterilization on the Cryoprotective Activity of EPS
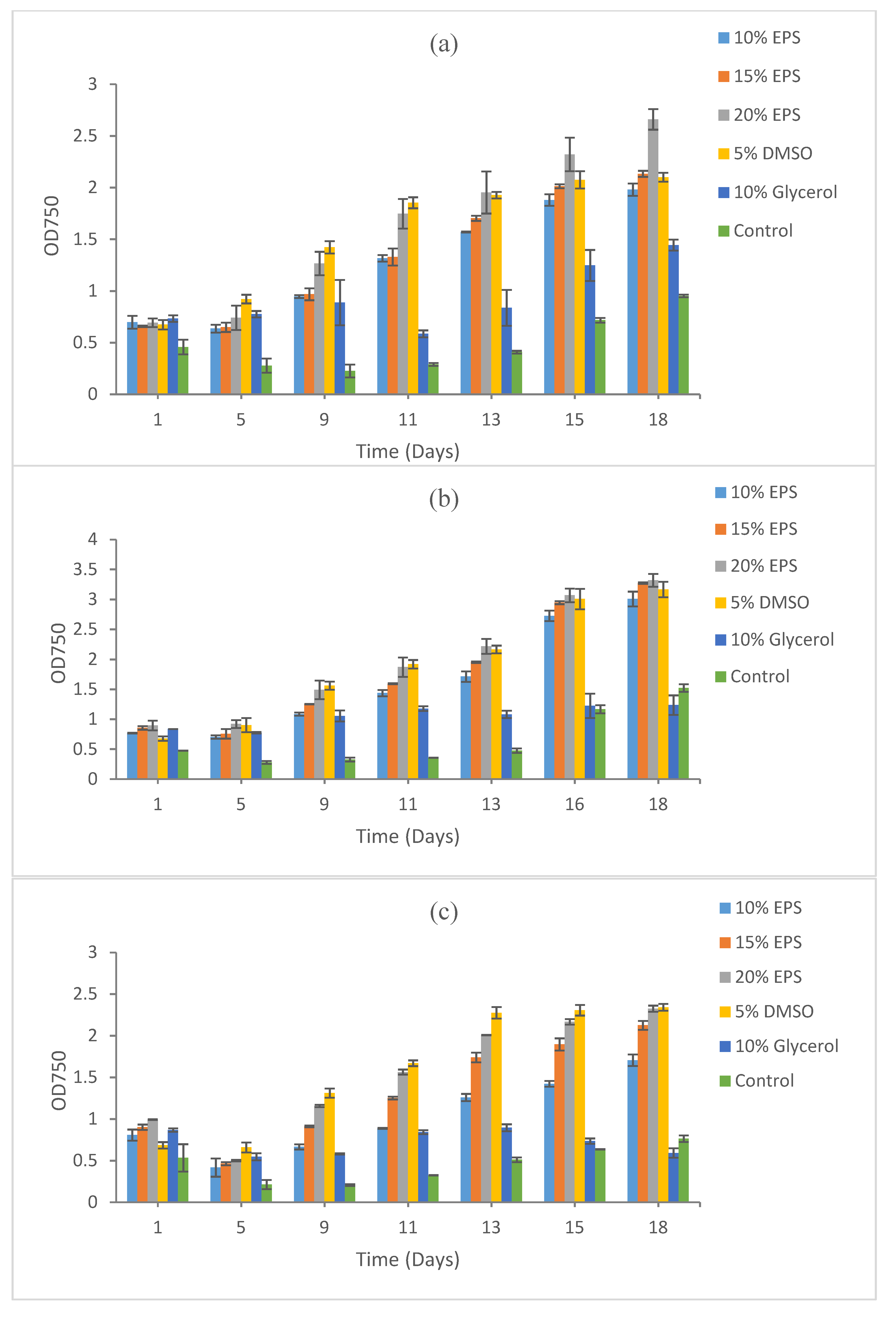
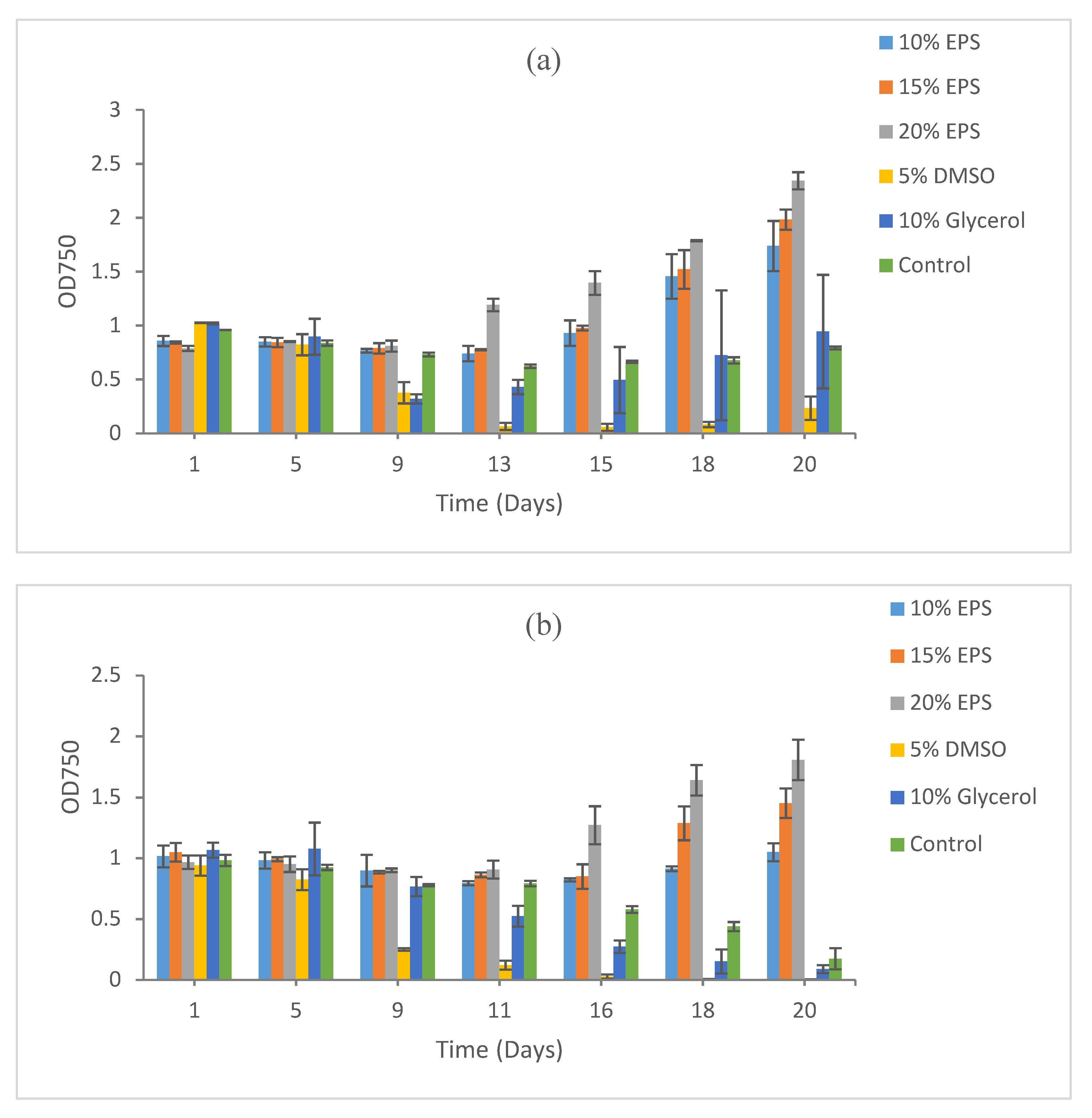
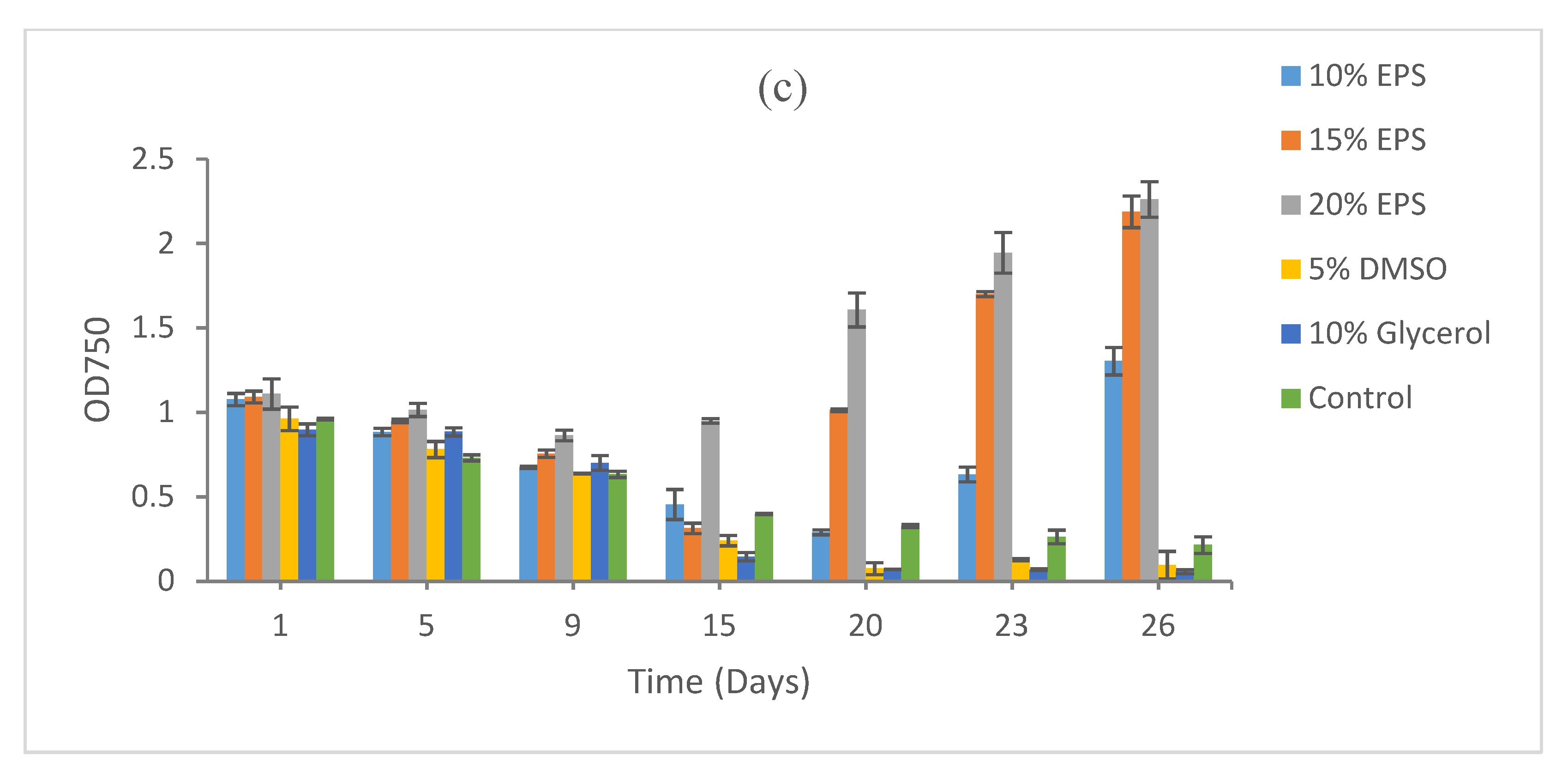
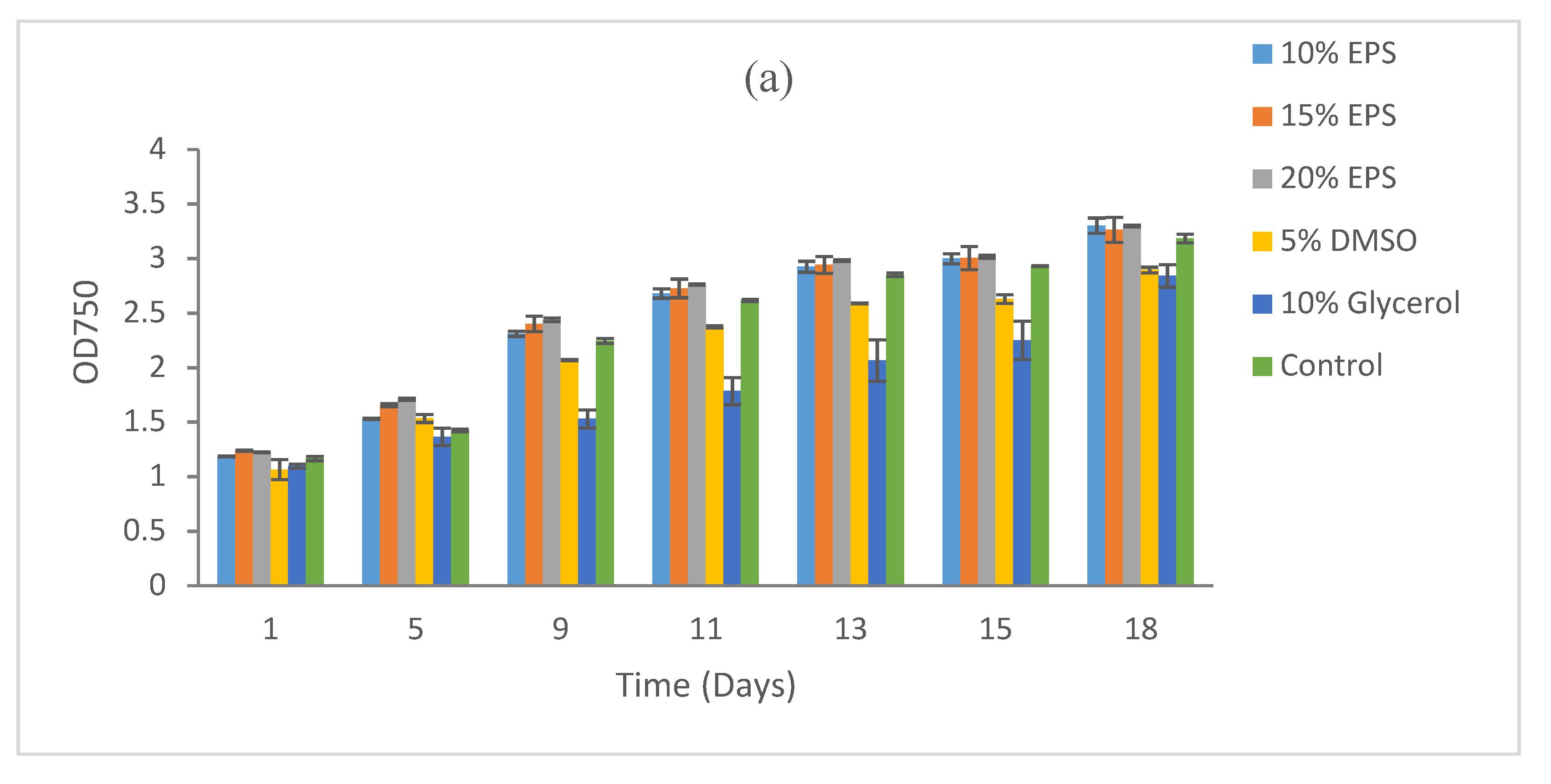
3. Results
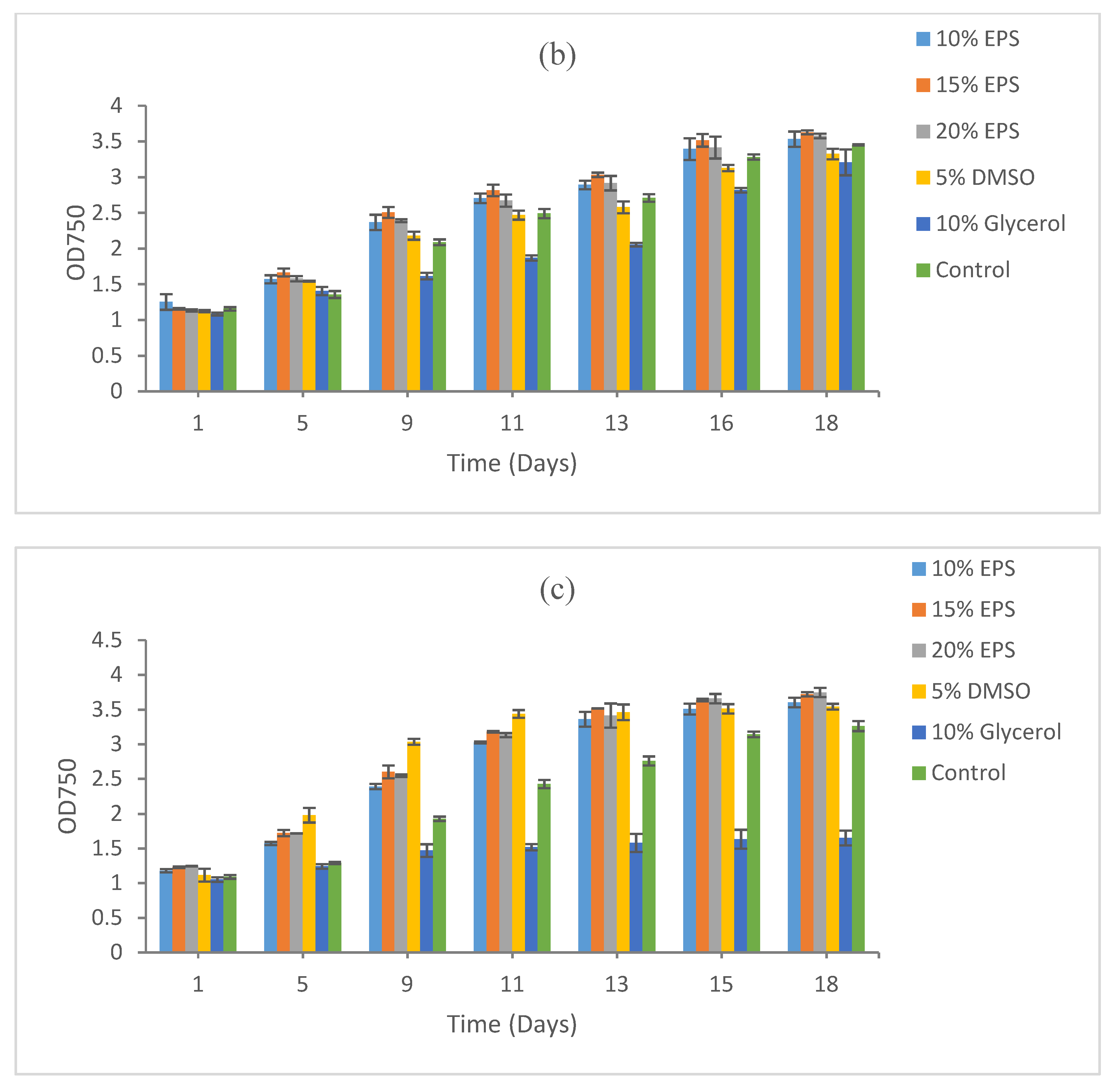
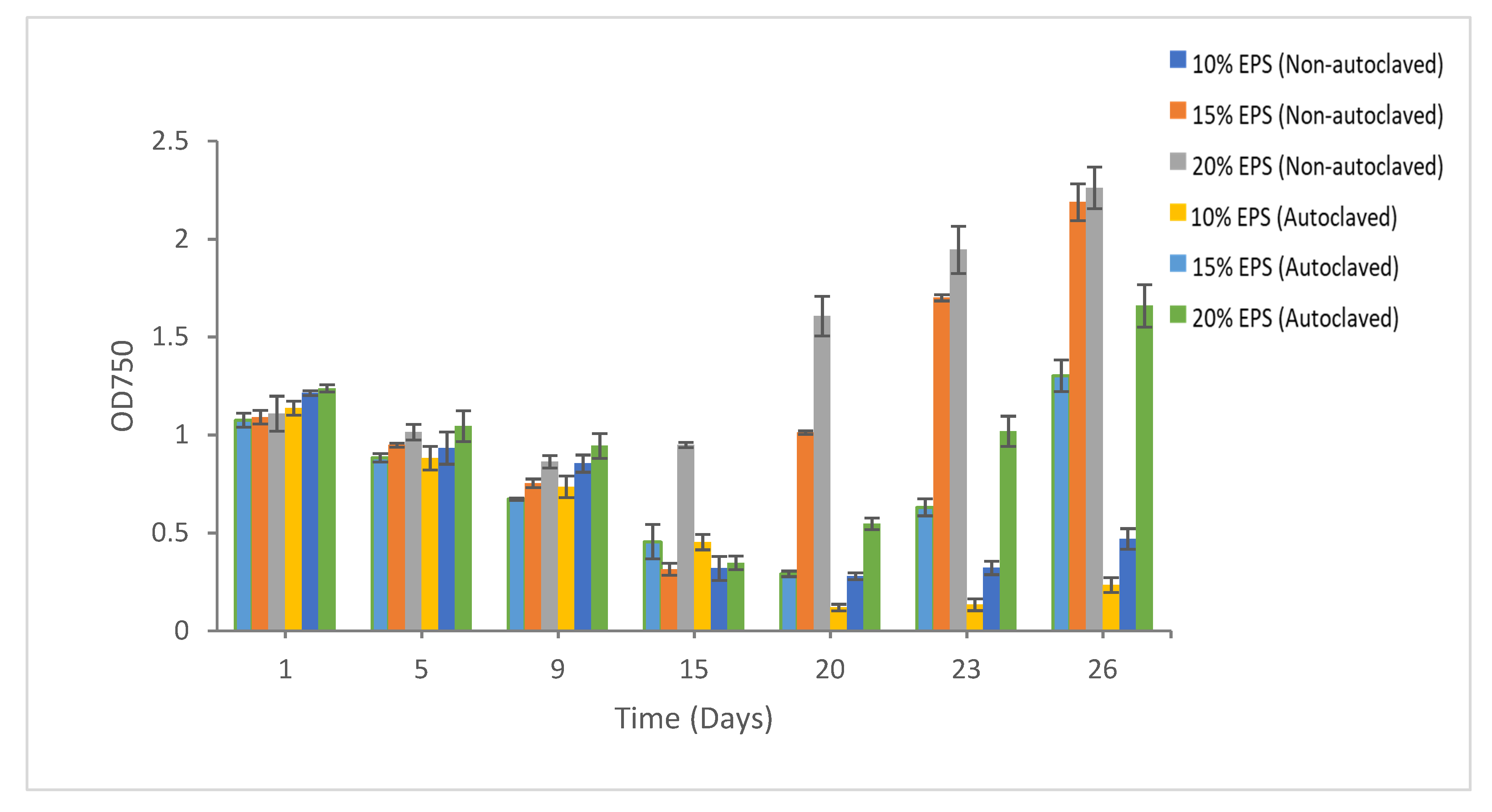
3.1. Growth Recovery of Cyanobacteria
3.1.1. Synechococcus sp. CBW1003
3.1.2. Synechococcus sp. CB0101
3.1.3. Microcystis aeruginosa PCC7806
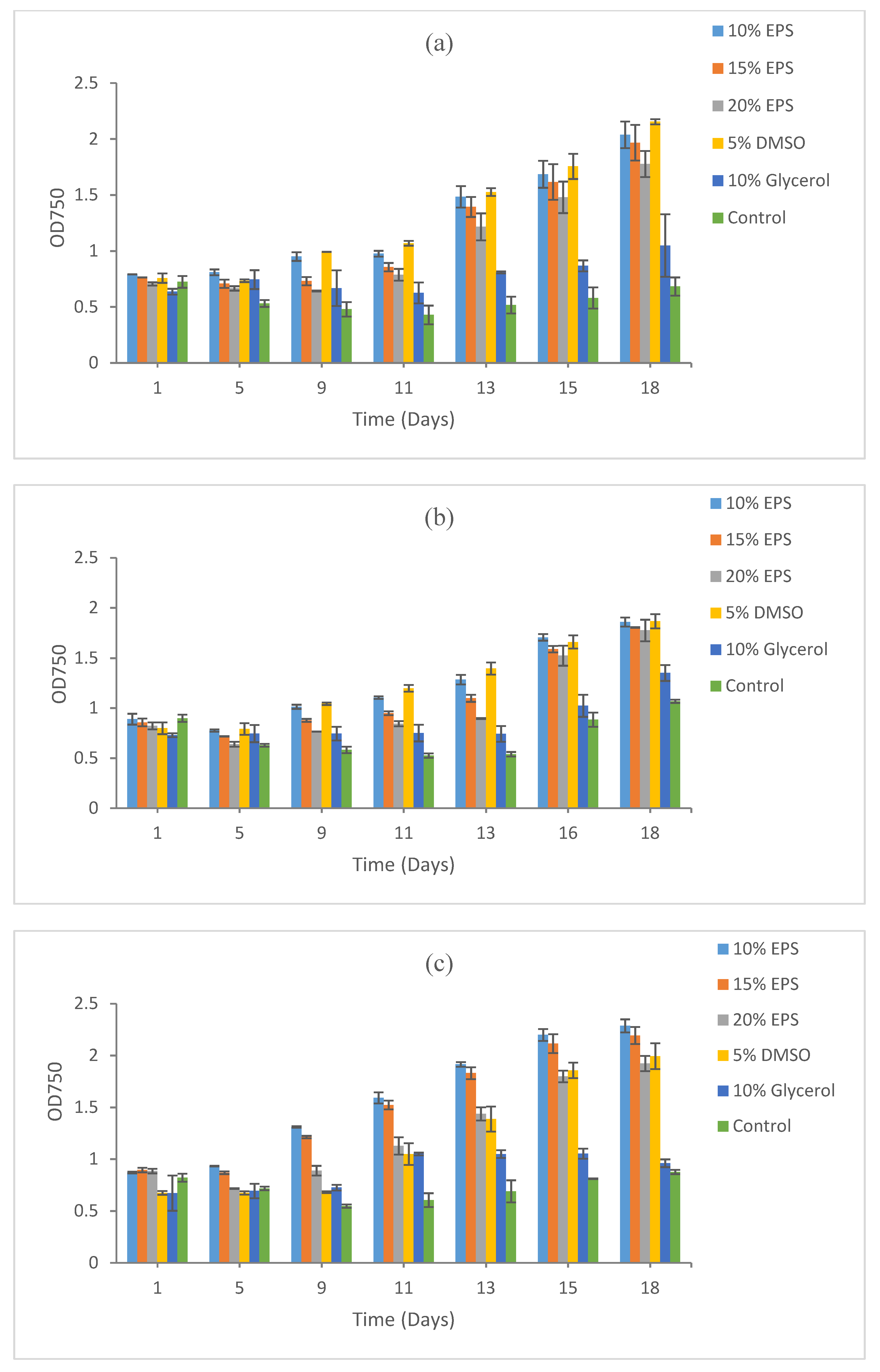
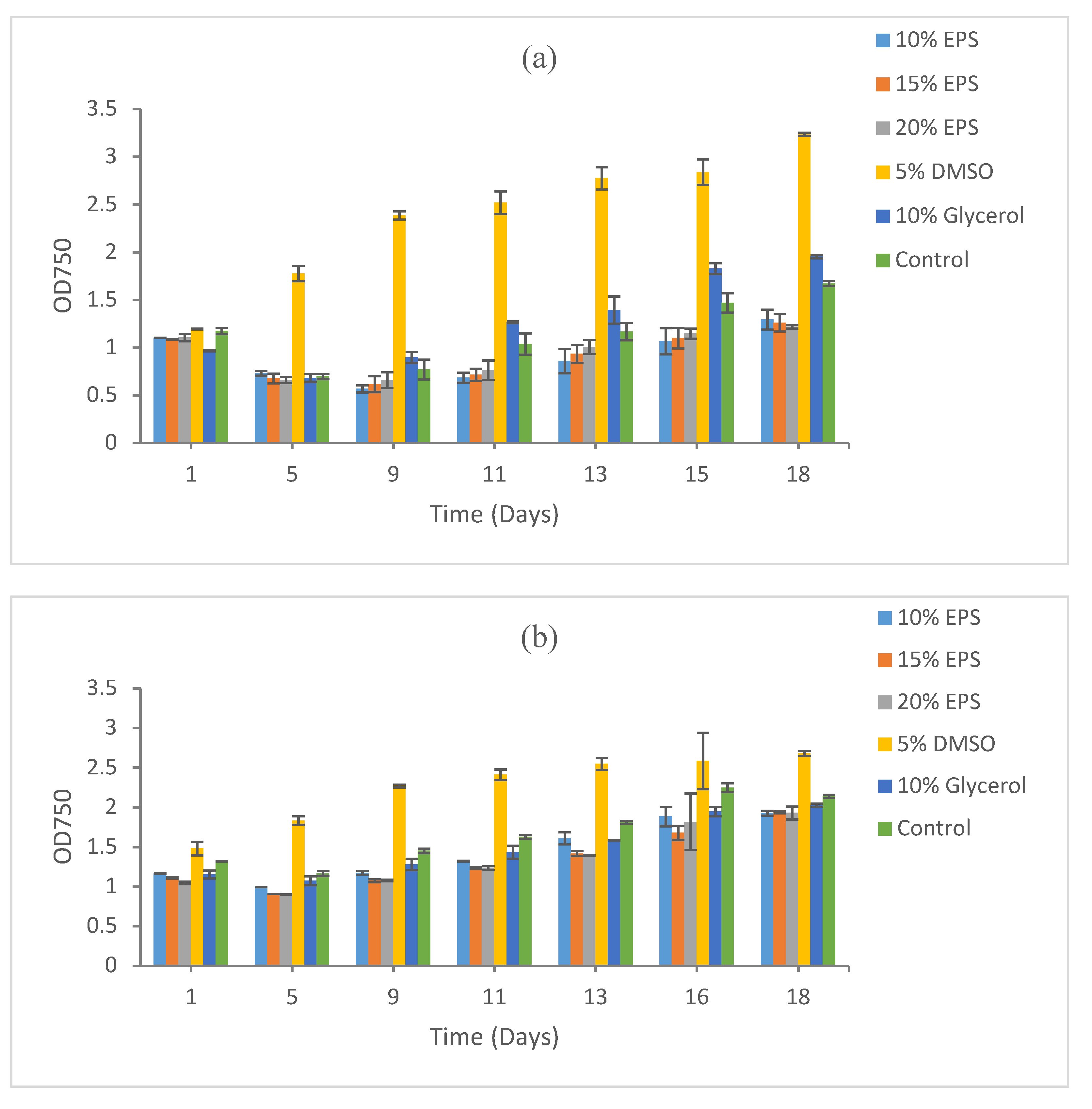
3.2. Growth Recovery of Eukaryotic Microalgae
3.2.1. Scenedesmus sp. HTB1
3.2.2. Chlorella vulgaris
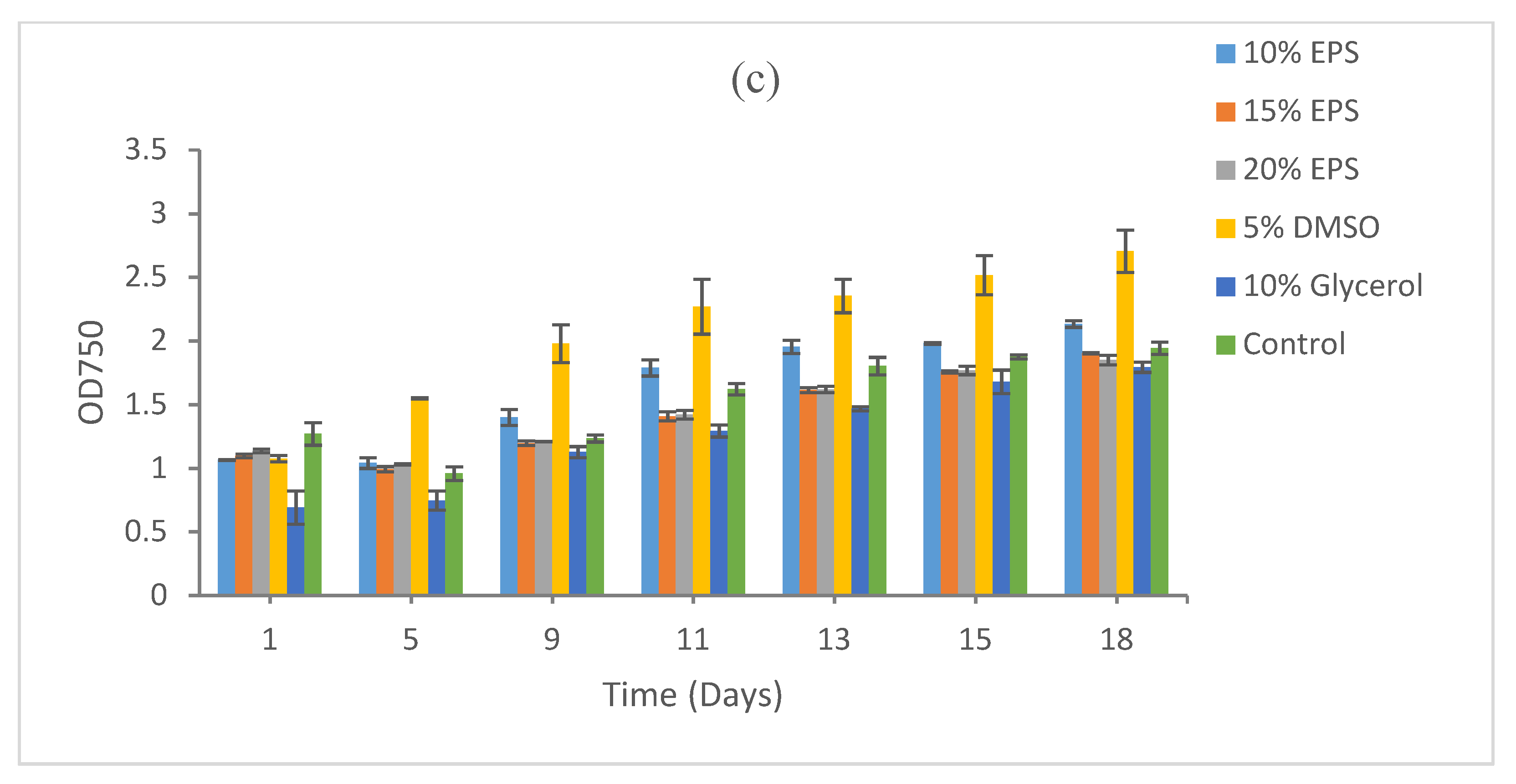
3.3. Effect of Heat Sterilization on the Cryoprotective Activity of the EPS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karlsson, J.O.; Toner, M. Long-term storage of tissues by cryopreservation: Critical issues. Biomaterials 1996, 17, 243–256. [Google Scholar] [CrossRef]
- Dando, T.R.; Bousfeld, I.J. Maintenance of Microorganisms and Cultured Cells: A Manual of Laboratory Methods; Kirsop, B.E., Doyle, A., Eds.; Academic Press: Cambridge, MA, USA, 1991; pp. 57–64. [Google Scholar]
- Day, J.G. Cyanobacteria: An Economic Perspective; Sharma, N.K., Rai, A.K., Stal, L.J., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 319–327. [Google Scholar]
- Kapoore, R.V.; Huete-Ortega, M.; Day, J.G.; Stanley, M.S.; Vaidyanathan, S. Effects of cryopreservation on viability and functional stability of an industrially relevant alga. Sci. Rep. 2019, 9, 2093. [Google Scholar] [CrossRef]
- Lorenz, M.; Friedl, T.; Day, J.G. Maintenance of actively metabolizing microalgal cultures. Algal Cult. Tech. 2005, 145, 50011-1. [Google Scholar]
- Day, J.G.; DeVille, M.M. Cryopreservation of algae. In Cryopreservation and Freeze-Drying Protocols; Humana Press: Totowa, NJ, USA, 1995; pp. 81–89. [Google Scholar]
- Bui, T.V.; Ross, I.L.; Jakob, G.; Hankamer, B. Impact of procedural steps and cryopreservation agents in the cryopreservation of chlorophyte microalgae. PLoS ONE 2013, 8, e78668. [Google Scholar] [CrossRef]
- Chian, R.C.; Quinn, P. Cryobiology: An overview. In Fertility Cryopreservation; Cambridge University Press: Oxford, UK, 2010; pp. 1–9. [Google Scholar]
- Fahy, G.M. The relevance of cryoprotectant “toxicity” to cryobiology. Cryobiology 1986, 23, 1–13. [Google Scholar] [CrossRef]
- Best, B.P. Cryoprotectant toxicity: Facts, issues, and questions. Rejuvenation Res. 2015, 18, 422–436. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.Z.; Su, B.X.; Lyu, S.S.; Hide, G.; Lun, Z.R.; Lai, D.H. Trehalose, an easy, safe and efficient cryoprotectant for the parasitic protozoan Trypanosoma brucei. Acta Trop. 2016, 164, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.A.; Kim, Y.H.; Kim, B.J.; Kim, B.G.; Kim, K.J.; Auh, J.H.; Schmidt, J.A.; Ryu, B.Y. Cryopreservation in trehalose preserves functional capacity of murine spermatogonial stem cells. PloS ONE 2013, 8, e54889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wowk, B. How cryoprotectants work. Cryonic 2007, 28, 3–7. [Google Scholar]
- Rampelotto, P.H. Extremophiles and extreme environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Olijve, L.L.; Meister, K.; DeVries, A.L.; Duman, J.G.; Guo, S.; Bakker, H.J.; Voets, I.K. Blocking rapid ice crystal growth through non-basal plane adsorption of antifreeze proteins. Proc. Natl. Acad. Sci. USA 2016, 113, 3740–3745. [Google Scholar] [CrossRef] [Green Version]
- Deming, J.W.; Young, J.N. The role of exopolysaccharides in microbial adaptation to cold habitats. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 259–284. [Google Scholar]
- Krembs, C.; Deming, J.W. The role of exopolymers in microbial adaptation to sea ice. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin/Heidelberg, Germany, 2008; pp. 247–264. [Google Scholar]
- Carrion, O.; Delgado, L.; Mercade, E. New emulsifying and cryoprotective exopolysaccharide from Antarctic Pseudomonas sp. ID1. Carbohydr. Polym. 2015, 117, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I.W. Bacterial exopolysaccharides. In Advances in Microbial Physiology; Academic Press: Cambridge, MA, USA, 1972; Volume 8, pp. 143–213. [Google Scholar]
- Krembs, C.; Eicken, H.; Junge, K.; Deming, J.W. High concentrations of exopolymeric substances in Arctic winter sea ice: Implications for the polar ocean carbon cycle and cryoprotection of diatoms. Deep Sea Res. Part I Oceanogr. Res. Pap. 2002, 49, 2163–2181. [Google Scholar] [CrossRef]
- Nichols, C.M.; Guezennec, J.; Bowman, J.P. Bacterial exopolysaccharides from extreme marine environments with special consideration of the Southern Ocean, sea ice, and deep-sea hydrothermal vents: A review. Mar. Biotechnol. 2005, 7, 253–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, P.; Chen, F.; Hassan, F.; Sosa, A.; Khan, S.; Badshah, M.; Shah, A.A. Bacterial community characterization of Batura Glacier in the karakoram range of pakistan. Int. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ali, P.; Shah, A.A.; Hassan, F.; Hertkorn, N.; Consior, M.; Chen, F. Production, optimization and partial characterization of exopolysaccharide extracted from psychrotrophic Pseudomonas sp. BGI-2 isolated from the ice of Batura glacier, Pakistan. Front. Microbiol. 2020. [Google Scholar] [CrossRef]
- Tamaru, Y.; Takani, Y.; Yoshida, T.; Sakamoto, T. Crucial role of extracellular polysaccharides in desiccation and freezing tolerance in the terrestrial cyanobacterium Nostoc commune. Appl. Environ. Microbiol. 2005, 71, 7327–7333. [Google Scholar] [CrossRef] [Green Version]
- Casillo, A.; Parrilli, E.; Sannino, F.; Mitchell, D.E.; Gibson, M.I.; Marino, G.; Lanzetta, R.; Parrilli, M.; Cosconati, S.; Novellino, E.; et al. Structure-activity relationship of the exopolysaccharide from a psychrophilic bacterium: A strategy for cryoprotection. Carbohydr. Polym. 2017, 156, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.B.; Chen, X.L.; He, H.L.; Zhang, X.Y.; Xie, B.B.; Yu, Y.; Chen, B.; Zhou, B.C.; Zhang, Y.Z. Structure and ecological roles of a novel exopolysaccharide from the Arctic sea ice bacterium Pseudoalteromonas sp. strain SM20310. Appl. Environ. Microbiol. 2013, 79, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, S.N.; Cresswell-Maynard, T.; Thomas, D.N.; Underwood, G.J. Production and characterization of the intra-and extracellular carbohydrates and polymeric substances (EPS) of three sea-ice diatom species, and evidence for a cryoprotective role for EPS. J. Phycol. 2012, 48, 1494–1509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Zhang, S.C.; Liu, X.Z.; Xu, Y.J.; Hu, J.H.; Xu, Y.Y.; Li, J.; Chen, S.L. Toxicity and protective efficiency of cryoprotectants to flounder (Paralichthys olivaceus) embryos. Theriogenology 2005, 63, 763–773. [Google Scholar] [CrossRef]
- Ali, P.; Shah, A.A.; Hasan, F.; Cai, H.; Sosa, A.; Chen, F. Draft genome sequence of a cold-adapted Pseudomonas sp. strain, BGI-2, isolated from the ice of Batura Glacier, Pakistan. Microbiol. Resour. Announc. 2019, 8, e00320-e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, M.M. Simple conditions for growth of unicellular blue-green algae on plates 1, 2. J. Phycol. 1968, 4, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Waterbury, J.B.; Willey, J.M. Isolation and growth of marine planktonic cyanobacteria. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1988; Volume 167, pp. 100–105. [Google Scholar]
- Xu, Y.; Jiao, N.; Chen, F. Novel psychrotolerant picocyanobacteria isolated from Chesapeake Bay in the winter. J. Phycol. 2015, 51, 782–790. [Google Scholar] [CrossRef]
- Chen, F.; Wang, K.; Kan, J.; Bachoon, D.S.; Lu, J.; Lau, S.; Campbell, L. Phylogenetic diversity of Synechococcus in the Chesapeake Bay revealed by Ribulose-1, 5-bisphosphate carboxylase-oxygenase (RuBisCO) large subunit gene (rbcL) sequences. Aquat. Microb. Ecol. 2004, 36, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, F.; Chen, F. High throughput screening of CO2-tolerating microalgae using GasPak bags. Aquat. Biosyst. 2013, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Gaget, V.; Chiu, Y.T.; Lau, M.; Humpage, A.R. From an environmental sample to a long-lasting culture: The steps to better isolate and preserve cyanobacterial strains. J. Appl. Phycol. 2017, 29, 309–321. [Google Scholar] [CrossRef]
- Esteves-Ferreira, A.A.; Corrêa, D.M.; Carneiro, A.P.; Rosa, R.M.; Loterio, R.; Araújo, W.L. Comparative evaluation of different preservation methods for cyanobacterial strains. J. Appl. Phycol. 2013, 25, 919–929. [Google Scholar] [CrossRef]
- Nakanishi, K.; Deuchi, K.; Kuwano, K. Cryopreservation of four valuable strains of microalgae, including viability and characteristics during 15 years of cryostorage. J. Appl. Phycol. 2012, 24, 1381–1385. [Google Scholar] [CrossRef]
- Aray-Andrade, M.M.; Uyaguari-Diaz, M.I.; Bermudez, J.R. Short-term deleterious effects of standard isolation and cultivation methods on new tropical freshwater microalgae strains. PeerJ 2018, 6, e5143. [Google Scholar] [CrossRef]
- Hanslick, J.L.; Lau, K.; Noguchi, K.K.; Olney, J.W.; Zorumski, C.F.; Mennerick, S.; Farber, N.B. Dimethyl sulfoxide (DMSO) produces widespread apoptosis in the developing central nervous system. Neurobiol. Dis. 2009, 34, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Hua, T.C.; Sun, D.W.; Liu, B.; Yang, G.; Cao, Y. Cryopreservation of tissue-engineered dermal replacement in Me2SO: Toxicity study and effects of concentration and cooling rates on cell viability. Cryobiology 2007, 55, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Saadaoui, I.; Al Emadi, M.; Bounnit, T.; Schipper, K.; Al Jabri, H. Cryopreservation of microalgae from desert environments of Qatar. J. Appl. Phycol. 2016, 28, 2233–2240. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, P.; Fucich, D.; Shah, A.A.; Hasan, F.; Chen, F. Cryopreservation of Cyanobacteria and Eukaryotic Microalgae Using Exopolysaccharide Extracted from a Glacier Bacterium. Microorganisms 2021, 9, 395. https://doi.org/10.3390/microorganisms9020395
Ali P, Fucich D, Shah AA, Hasan F, Chen F. Cryopreservation of Cyanobacteria and Eukaryotic Microalgae Using Exopolysaccharide Extracted from a Glacier Bacterium. Microorganisms. 2021; 9(2):395. https://doi.org/10.3390/microorganisms9020395
Chicago/Turabian StyleAli, Pervaiz, Daniel Fucich, Aamer Ali Shah, Fariha Hasan, and Feng Chen. 2021. "Cryopreservation of Cyanobacteria and Eukaryotic Microalgae Using Exopolysaccharide Extracted from a Glacier Bacterium" Microorganisms 9, no. 2: 395. https://doi.org/10.3390/microorganisms9020395