

Applications of Molecular Markers for Developing Abiotic-Stress-Resilient Oilseed Crops
 , , , , ,
, , , , ,  , ,
, ,
Abstract
:1. Introduction
2. Applications of Molecular Markers in Development of Abiotic-Stress-Tolerant Oilseed Crops
2.1. Drought
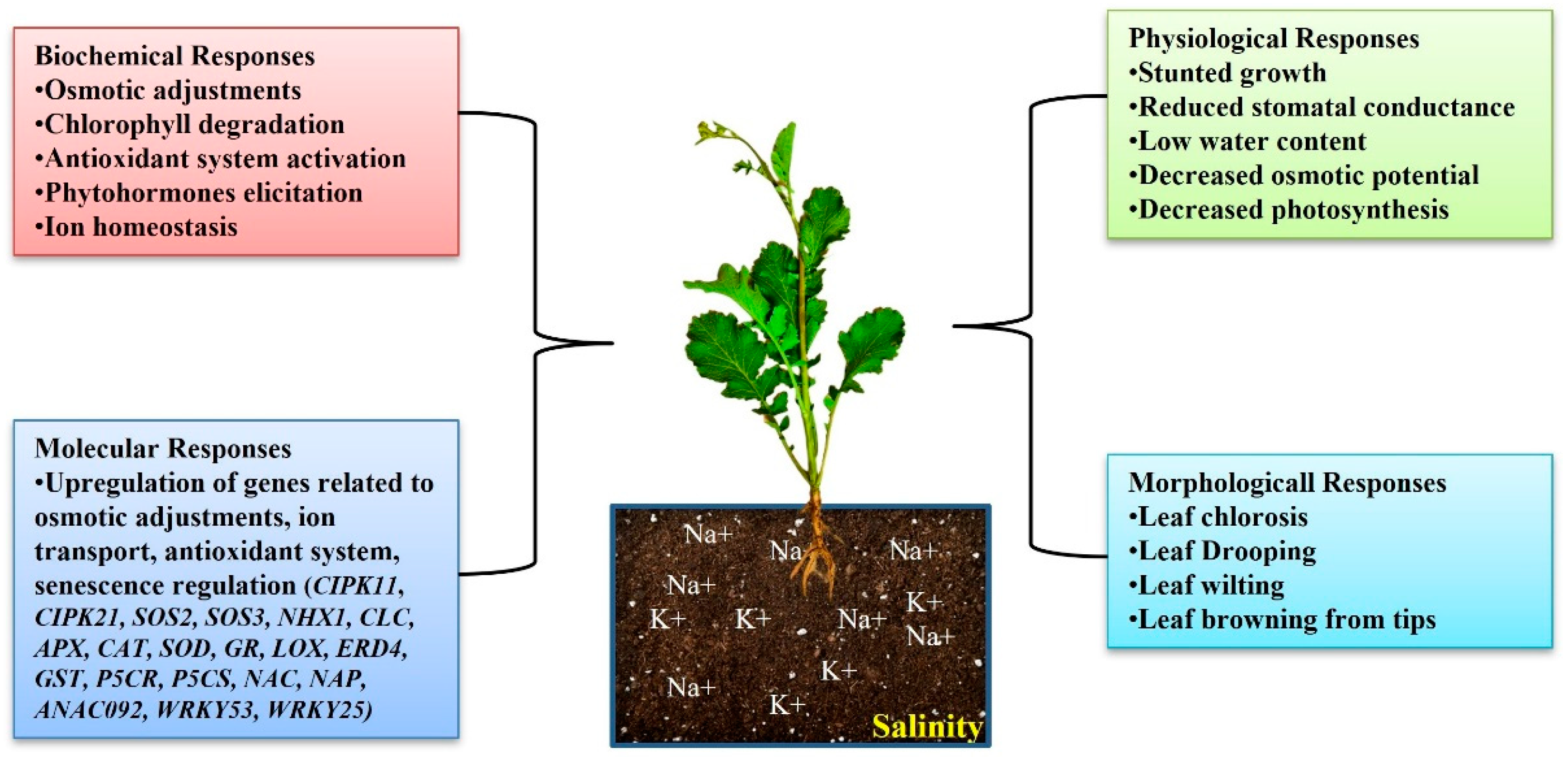
2.2. Salinity
2.3. Heavy Metal Stress
2.4. Flooding
2.5. Cold Stress
2.6. Heat Stress
3. Advent of a New Era for Development of Molecular Markers
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- NMOOP. Present Status of Oilseed Crops and Vegetable Oils in India. 2018; pp. 1–72. Available online: https://www.nfsm.gov.in/StatusPaper/NMOOP2018.pdf (accessed on 20 November 2022).
- Mittal, S. Structural Shift in Demand for Food: Projections for 2020; Working Paper No. 184; Indian Council for Research on International Economic Relations (ICRIER): New Delhi, India, 2006. [Google Scholar]
- DaMatta, F.M.; Grandis, A.; Arenque, B.C.; Buckeridge, M.S. Impacts of climate changes on crop physiology and food quality. Food Res. Int. 2010, 43, 1814–1823. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; Volume 2. [Google Scholar]
- Chauhan, J.S.; Singh, K.H.; Singh, V.V.; Kumar, S. Hundred years of rapeseed-mustard breeding in India: Accomplishments and future strategies. Indian J. Agric. Sci. 2011, 81, 1093–1109. [Google Scholar]
- Singh, J.; Sharma, P.C.; Singh, V. Breeding Mustard (Brassica juncea) for Salt Tolerance: Problems and Prospects. In Brassica Breeding and Biotechnology; Islam, A., Hossain, M., Islam, A., Eds.; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Wu, W.; Shah, F.; Duncan, R.W.; Ma, B.L. Grain yield, root growth habit and lodging of eight oilseed rape genotypes in response to a short period of heat stress during flowering. Agric. For. Meteorol. 2020, 287, 107954. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. Short-term heat stress during flowering results in a decline in Canola seed productivity. J. Agron. Crop Sci. 2022, 208, 486–496. [Google Scholar] [CrossRef]
- Wei, Y.; Jin, J.; Jiang, S.; Ning, S.; Liu, L. Quantitative Response of Soybean Development and Yield to Drought Stress during Different Growth Stages in the Huaibei Plain, China. Agronomy 2018, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Desclaux, D.; Huynh, T.T.; Roumet, P. Identification of soybean plant characteristics that indicate the timing of drought stress. Crop Sci. 2000, 40, 716–722. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Ullah, I.; Ali, S.; Kang, S.M.; Lee, I.J. Alleviation of salt stress response in soybean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Ann. Microbiol. 2019, 69, 797–808. [Google Scholar] [CrossRef]
- VanToai, T.T.; Hoa, T.T.C.; Hue, N.T.N.; Nguyen, H.T.; Shannon, J.G.; Rahman, M.A. Flooding tolerance of soybean [Glycine max (L.) Merr.] germplasm from Southeast Asia under field and screen-house environments. Open Agric. J. 2010, 4, 38–46. [Google Scholar] [CrossRef]
- Rhine, M.D.; Stevens, G.; Shannon, G.; Wrather, A.; Sleper, D. Yield and Nutritional Responses to Waterlogging of Soybean Cultivars. Irrig. Sci. 2010, 28, 135–142. [Google Scholar] [CrossRef]
- Staniak, M.; Czopek, K.; Stępień-Warda, A.; Kocira, A.; Przybyś, M. Cold Stress during Flowering Alters Plant Structure, Yield and Seed Quality of Different Soybean Genotypes. Agronomy 2021, 11, 2059. [Google Scholar] [CrossRef]
- Kalarani, M.K.; Senthil, A.; Thangaraj, M. Effect of water stress on morpho-physiological traits of sunflower (Helianthus annus L.) genotypes. Madras Agric. J. 2004, 91, 224–239. [Google Scholar]
- Hussain, M.; Malik, M.A.; Farooq, M.; Ashraf, M.Y.; Cheema, M.A. Improving drought tolerance by exogenous application of glycinebetaine and salicylic acid in sunflower. J. Agron. Crop Sci. 2008, 194, 193–199. [Google Scholar] [CrossRef]
- Snowden, R.; Wheeler, B.D. Iron toxicity to fenplant species. J. Ecol. 1993, 81, 35–46. [Google Scholar]
- Wang, L.; Zhang, Y.; Qi, X.; Li, D.; Wei, W.; Zhang, X. Global gene expression responses to waterlogging in roots of sesame (Sesamum indicum L.). Acta Physiol. Plant. 2012, 34, 2241–2249. [Google Scholar] [CrossRef]
- Golestani, M.; Pakniyat, H. Evaluation of drought tolerance indices in sesame lines. J. Sci. Technol. Agric. Nat. Resour. 2007, 11, 141–150. [Google Scholar]
- Heidari, M.; Galavi, M.; Hassani, M. Effect of sulfur and iron fertilizers on yield, yield components and nutrient uptake in sesame (S. indicum L.) under water stress. Afr. J. Biotechnol. 2011, 10, 8816–8822. [Google Scholar] [CrossRef] [Green Version]
- Joshan, Y.; Sani, B.; Jabbari, H.; Mozafari, H.; Moaveni, P. Effect of drought stress on oil content and fatty acids composition of some safflower genotypes. Plant Soil Environ. 2019, 65, 563–567. [Google Scholar] [CrossRef] [Green Version]
- Hamidou, F.; Halilou, O.; Vadez, V. Assessment of groundnut under combined heat and drought stress. J. Agron. Crop Sci. 2013, 199, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Manavalan, L.P.; Guttikonda, S.K.; Tran, L.-S.; Nguyen, H.T. Physiological and Molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [Green Version]
- Mian, M.A.R.; Ashley, D.A.; Boerma, H.R. An additional QTL for water use efficiency in soybean. Crop Sci. 1998, 38, 390–393. [Google Scholar] [CrossRef]
- Gopalakrishna, R.; Kumar, G.; KrishnaPrasad, B.T.; Mathew, M.K.; Udaya Kumar, M. A Stress-responsive gene from groundnut, Gdi-15, is homologous to flavonol 3-o-glucosyltransferase involved in anthocyanin biosynthesis. Biochem. Biophys. Res. Commun. 2001, 284, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Rauf, S. Breeding sunflower (Helianthus annuus L.) for drought tolerance. Commun. Biometry Crop Sci. 2008, 3, 29–44. [Google Scholar]
- Hervé, D.; Fabre, F.; Berrios, E.F.; Leroux, N.; Al Chaarani, G.; Planchon, C.; Sarrafi, A.; Gentzbittel, L. QTL analysis of photosynthesis and water status traits in sunflower (Helianthus annuus L.) under greenhouse conditions. J. Exp. Bot. 2001, 52, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Meng, R.; Newman, M.; Gao, G.; Pittman, R.N.; Prakash, C.S. Microsatellites as DNA markers in cultivated peanut (Arachis hypogaea L.). BMC Plant Biol. 2003, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Ulfat, M.; Athar, H.R.; Ashraf, M.; Akram, N.A.; Jamil, A. Appraisal of physiological and biochemical selection criteria for evaluation of salt tolerance in canola (Brassica napus L.). Pak. J. Bot. 2007, 39, 1593–1608. [Google Scholar]
- Rosegrant, M.W.; Fernandez, M.; Sinha, A.; Alder, J.; Ahammad, H.; De Fraiture, C.; Eickhour, B.; Fonseca, J.; Huang, J.; Koyama, O.; et al. Looking into the future for agriculture and AKST. In International Assessment of Agricultural Knowledge, Science and Technology for Development (IAASTD): Agriculture at a Crossroads, Global Report; McIntyre, B.D., Herren, H.R., Wakhungu, J., Watson, R.T., Eds.; Island Press: Washington, DC, USA, 2009; pp. 307–376. [Google Scholar]
- Turner, N. Optimizing water use. In Proceedings of the Crop Science, Progress and Prospects, Third International Crop Science Congress, Hambourg, Germany, 17–22 August 2000; CABI: Wallingford, UK, 2001; pp. 119–135. [Google Scholar]
- Pennisi, E. The Blue Revolution, Drop by Drop, Gene by Gene. Science 2008, 320, 171–173. [Google Scholar] [CrossRef]
- Boyer, J.S. Plant Productivity and Environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef]
- Gleick, P.H. The World’s Water 1998-1999: The Biennial Report on Freshwater Resources; Island Press: Washington, DC, USA, 1998. [Google Scholar]
- Araus, J.L. Plant Breeding and Drought in C3 Cereals: What Should We Breed For? Ann. Bot. 2002, 89, 925–940. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Ostrowska, A. Drought-Stress Induced Physiological and Molecular Changes in Plants. Int. J. Mol. Sci. 2022, 23, 4698. [Google Scholar] [CrossRef]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Biol. 1996, 47, 377–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Pozo, J.C.; Ramirez-Parra, E. Deciphering the molecular bases for drought tolerance in Arabidopsis autotetraploids. Plant Cell Environ. 2014, 37, 2722–2737. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Govinal Badiger, B.; Ravi, K.; Nagaraj, K.M. Drought tolerance mechanisms and their molecular basis. In Plant Abiotic Stress, 2nd ed.; Jenks, M.A., Hasegawa, P.M., Eds.; Wiley: Hoboken, NJ, USA, 2014; pp. 15–46. [Google Scholar]
- Alqudah, A.M.; Samarah, N.H.; Mullen, R.E. Drought Stress Effect on Crop Pollination, Seed Set, Yield and Quality. In Alternative Farming Systems, Biotechnology, Drought Stress and Ecological Fertilisation; Lichtfouse, E., Ed.; Springer: Dordrecht, The Netherlands, 2011; Volume 6, pp. 193–213. [Google Scholar]
- Islam, M.S.; Akhter, M.M.; Sabagh, A.E.; Liu, L.Y.; Nguyen, N.T.; Ueda, A.; Masaoka, Y.; Saneoka, H. Comparative Studies on Growth and Physiological Responses to Saline and Alkaline Stresses of Foxtail Millet (“SetariaItalica” L.) and Proso Millet (“Panicum miliaceum” L.). Aust. J. Crop Sci. 2011, 5, 1269–1277. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Good, A.G.; Maclagan, J.L. Effects of drought stress on the water relations in Brassica species. Can. J. Plant Sci. 1993, 73, 525–529. [Google Scholar] [CrossRef] [Green Version]
- Arya, H.; Singh, M.B.; Bhalla, P.L. Towards Developing Drought-smart Soybeans. Front. Plant Sci. 2021, 12, 750664. [Google Scholar] [CrossRef]
- Chen, H.; Kumawat, G.; Yan, Y.; Fan, B.; Xu, D. Mapping and validation of a major QTL for primary root length of soybean seedlings grown in hydroponic conditions. BMC Genom. 2021, 22, 132. [Google Scholar] [CrossRef]
- Mian, M.A.R.; Bailey, M.A.; Ashley, D.A.; Wells, R.; Carter, T.E.; Parrott, W.A.; Boerma, H.R. Molecular markers associated with water use efficiency and leaf ash in soybean. Crop Sci. 1996, 36, 1252–1257. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Baird, W.V. The ribosomal small-subunit protein S28 gene from Helianthus annuus (Asteraceae) is down-regulated in response to drought, high salinity, and abscisic acid. Am. J. Bot. 2003, 90, 526–531. [Google Scholar] [CrossRef]
- Dezar, C.A.; Gago, G.M.; Gonzalez, D.H.; Chan, R.L. Hahb-4, a sunflower homeobox-leucine zipper gene, is a developmental regulator and confers drought tolerance to Arabidopsis thaliana plants. Transgenic Res. 2005, 14, 429–440. [Google Scholar] [CrossRef]
- Herrera-Rodríguez, M.B.; Pérez-Vicente, R.; Maldonado, J.-M. Expression of asparagine synthetase genes in sunflower (Helianthus annuus) under various environmental stresses. Plant Physiol. Biochem. 2007, 45, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Poormohammad Kiani, S.; Grieu, P.; Maury, P.; Hewezi, T.; Gentzbittel, L.; Sarrafi, A. Genetic variability for physiological traits under drought conditions and differential expression of water stress-associated genes in sunflower (Helianthus annuus L.). Theor. Appl. Genet. 2007, 114, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, Y.A.S.; García-Moreno, M.J.; Redondo-Nevado, J.; Velasco, L.; Pérez-Vich, B. Development and characterization of genomic microsatellite markers in safflower (Carthamus tinctorius L.): Development of SSR markers in safflower. Plant Breed. 2011, 130, 237–241. [Google Scholar] [CrossRef]
- Tang, J.; Lou, Z.; Wang, Y.; Guo, M. Expression of a Small Heat Shock Protein (CTL-Hsyapr) screened by CDNA-AFLP approach is correlated with hydroxysafflor Yellow A in safflower (Carthamus tinctorius L.). Biochem. Syst. Ecol. 2010, 38, 722–730. [Google Scholar] [CrossRef]
- Chapman, M.A.; Hvala, J.; Strever, J.; Matvienko, M.; Kozik, A.; Michelmore, R.W.; Tang, S.; Knapp, S.J.; Burke, J.M. Development, polymorphism, and cross-taxon utility of EST-SSR markers from safflower (Carthamus tinctorius L.). Theor. Appl. Genet. 2009, 120, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Mayerhofer, R.; Archibald, C.; Bowles, V.; Good, A.G. Development of molecular markers and linkage maps for the Carthamus Species C. tinctorius and C. oxyacanthus. Genome 2010, 53, 266–276. [Google Scholar] [CrossRef]
- García-Moreno, M.J.; Velasco, L.; Pérez-Vich, B. Transferability of non-genic microsatellite and gene-based sunflower markers to safflower. Euphytica 2010, 175, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Mirzahashemi, M.; Mohammadi-nejad, G.; Golkar, P. A QTL linkage map of safflower for yield under drought stress at reproductive stage. Iran. J. Genet. Plant Breed. 2015, 4, 20–27. [Google Scholar]
- Stalker, H.T.; Mozingo, L.G. Molecular markers of Arachis and marker-assisted selection. Peanut Sci. 2001, 28, 117–123. [Google Scholar] [CrossRef]
- Hopkins, M.S.; Casa, A.M.; Wang, T.; Mitchell, S.E.; Dean, R.E.; Kochert, G.D.; Kresovich, S. Discovery and characterization of polymorphic simple sequence repeats (SSRs) in peanut. Crop Sci. 1999, 39, 1243–1247. [Google Scholar] [CrossRef]
- Ferguson, M.E.; Burow, M.D.; Schulze, S.R.; Bramel, P.J.; Paterson, A.H.; Kresovich, S.; Mitchell, S. Microsatellite identification and characterization in peanut (A. hypogaea L.). Theor. Appl. Genet. 2003, 108, 1064–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, K.; Vadez, V.; Isobe, S.; Mir, R.R.; Guo, Y.; Nigam, S.N.; Gowda, M.V.C.; Radhakrishnan, T.; Bertioli, D.J.; Knapp, S.J.; et al. Identification of several small main-effect QTLs and a large number of epistatic QTLs for drought tolerance related traits in groundnut (Arachis hypogaea L.). Theor. Appl. Genet. 2011, 122, 1119–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautami, B.; Pandey, M.K.; Vadez, V.; Nigam, S.N.; Ratnakumar, P.; Krishnamurthy, L.; Radhakrishnan, T.; Gowda, M.V.C.; Narasu, M.L.; Hoisington, D.A.; et al. Quantitative trait locus analysis and construction of consensus genetic map for drought tolerance traits based on three recombinant inbred line populations in cultivated groundnut (Arachis hypogaea L.). Mol. Breed. 2012, 30, 757–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, F.; Gill, R.A.; Ali, B.; Farooq, M.A.; Xu, L.; Najeeb, U.; Zhou, W. Sesame. In Breeding Oilseed Crops for Sustainable Production; Gupta, S.K., Ed.; Elsevier: San Diego, CA, USA, 2016; pp. 135–147. [Google Scholar]
- Sun, J.; Rao, Y.; Le, M.; Yan, T.; Yan, X.; Zhou, H. Effects of drought stress on sesame growth and yield characteristics and comprehensive evaluation of drought tolerance. Chin. J. Oil Crop Sci. 2010, 32, 525–533. [Google Scholar]
- Boureima, S.; Eyletters, M.; Diouf, M.; Diop, T.A.; Van Damme, P. Sensitivity of seed germination and seedling radicle growth to drought stress in sesame (Sesamum indicum L.). Res. J. Environ. Sci. 2011, 5, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Dossa, K.; Li, D.; Zhou, R.; Yu, J.; Wang, L.; Zhang, Y.; You, J.; Liu, A.; Mmadi, M.A.; Fonceka, D.; et al. The genetic basis of drought tolerance in the high oil crop Sesamum indicum. Plant Biotechnol. J. 2019, 17, 1788–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dossa, K.; Niang, M.; Assogbadjo, A.E.; Cissé, N.; Diouf, D. Whole genome homology-based identification of candidate genes for drought tolerance in sesame (Sesamum indicum L.). Afr. J. Biotechnol. 2016, 15, 1464–1475. [Google Scholar]
- Li, D.; Dossa, K.; Zhang, Y.; Wei, X.; Wang, L.; Zhang, Y.; Liu, A.; Zhou, R.; Zhang, X. GWAS uncovers differential genetic bases for drought and salt tolerances in sesame at the germination stage. Genes 2018, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Dossa, K.; Diouf, D.; Cissé, N. Genome-wide investigation of Hsf genes in sesame reveals their segmental duplication expansion and their active role in drought stress response. Front. Plant Sci. 2016, 7, 1522. [Google Scholar] [CrossRef]
- Liu, W.; Li, D.; Ren, G.; Zhang, Y.; Wen, F.; Han, J.; Zhang, X. Drought resistance of sesame germplasm resources and association analysis at adult stage. Sci. Agric. Sin. 2017, 50, 625–639. [Google Scholar]
- Liang, J.; Sun, J.; Ye, Y.; Yan, X.; Yan, T.; Rao, Y.; Zhou, H.; Le, M. QTL mapping of PEG-induced drought tolerance at the early seedling stage in sesame using whole genome re-sequencing. PLoS ONE 2021, 16, e0247681. [Google Scholar] [CrossRef] [PubMed]
- Dossa, K.; Wei, X.; Li, D.; Fonceka, D.; Zhang, Y.; Wang, L.; Yu, J.; Boshou, L.; Diouf, D.; Cissé, N. Insight into the AP2/ERF transcription factor superfamily in sesame and expression profiling of DREB subfamily under drought stress. BMC Plant Biol. 2016, 16, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, E.A. Response to abiotic stress. In Biochemistry & Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 1158–1203. [Google Scholar]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- FAO. The World Map of Salt Affected Soil [WWW Document]. Food and Agriculture Organization, United Nations. 2021. Available online: https://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/global-map-of-salt-affected-soils/en/ (accessed on 12 May 2021).
- Machado, R.; Serralheiro, R. Soil salinity: Effect on vegetable crop growth. management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.Y.; Sarwar, G.; Ashraf, M.; Afaf, R.; Sattar, A. Salinity induced changes in α-amylase activity during germination and early cotton seedling growth. Biol. Plant. 2002, 45, 589–591. [Google Scholar] [CrossRef]
- Seckin, B.; Sekmen, A.H.; Türkan, İ. An enhancing effect of exogenous mannitol on the antioxidant enzyme activities in roots of wheat under salt stress. J. Plant Growth Regul. 2009, 28, 12–20. [Google Scholar] [CrossRef]
- Anuradha, S.; Seeta Ram Rao, S. Effect of brassinosteroids on salinity stress induced inhibition of seed germination and seedling growth of rice (Oryza sativa L.). Plant Growth Regul. 2001, 33, 151–153. [Google Scholar] [CrossRef]
- Bybordi, A. The influence of salt stress on seed germination, growth and yield of canola cultivars. Not. Bot. Horti Agrobot. Cluj Napoca 2010, 38, 128–133. [Google Scholar]
- Flagella, Z.; Giuliani, M.M.; Rotunno, T.; Di Caterina, R.; De Caro, A. Effect of saline water on oil yield and quality of a high oleic sunflower (Helianthus annuus L.) Hybrid. Eur. J. Agron. 2004, 21, 267–272. [Google Scholar] [CrossRef]
- Sinaki, J.M.; Majidi, E.; Shirani-Rad, A.L.; Moormohammadi, G.; Zarei, G. The effect of water deficit during growth stage of canola (Brassica napus L). Am.-Eurasian J. Agric. Environ. Sci. 2007, 2, 417–422. [Google Scholar]
- Raza, A.; Tabassum, J.; Fakhar, A.Z.; Sharif, R.; Chen, H.; Zhang, C.; Ju, L.; Fotopoulos, V.; Siddique, K.H.M.; Singh, R.K.; et al. Smart reprograming of plants against salinity stress using modern biotechnological tools. Crit. Rev. Biotechnol. 2022, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Nayidu, N.; Bollina, V.; Kagale, S. Oilseed crop productivity under salt stress. In Ecophysiology and Responses of Plants under Salt Stress; Springer: New York, NY, USA, 2013; pp. 249–265. [Google Scholar]
- Kumar, D. Salt tolerance in oilseed Brassicas—Present status and future prospects. Plant Breed. Abstr. 1995, 65, 1439–1447. [Google Scholar]
- Lee, G.J.; Carter, T.E., Jr.; Villagarcia, M.R.; Li, Z.; Zhou, X.; Gibbs, M.O.; Boerma, H.R. A Major QTL Conditioning salt tolerance in S-100 soybean and descendent cultivars. Theor. Appl. Genet. 2004, 109, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Hamwieh, A.; Tuyen, D.D.; Cong, H.; Benitez, E.R.; Takahashi, R.; Xu, D.H. Identification and Validation of a major QTL for salt tolerance in soybean. Euphytica 2011, 179, 451–459. [Google Scholar] [CrossRef]
- Chen, H.; Cui, S.; Fu, S.; Gai, J.; Yu, D. Identification of quantitative trait loci associated with salt tolerance during seedling growth in soybean (Glycine max L.). Aust. J. Agric. Res. 2008, 59, 1086–1091. [Google Scholar] [CrossRef]
- Hamwieh, A.; Xu, D. Conserved salt tolerance quantitative trait locus (QTL) in wild and cultivated soybeans. Breed. Sci. 2008, 58, 355–359. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.I.; Lyra, D.-A.; Farooq, M.; Nikoloudakis, N.; Khalid, N. Salt and drought stresses in safflower: A Review. Agron. Sustain. Dev. 2016, 36, 4. [Google Scholar] [CrossRef] [Green Version]
- Amini, H.; Arzani, A.; Bahrami, F. Seed yield and some physiological traits of safflower as affected by water deficit stress. Int. J. Plant Prod. 2013, 7, 597–614. [Google Scholar]
- Johnson, R.C.; Kisha, T.J.; Evans, M.A. Characterizing safflower germplasm with AFLP molecular markers. Crop Sci. 2007, 47, 1728–1736. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; von Witzke-Ehbrecht, S.; Maass, B.L.; Becker, H.C. Relationships among different geographical groups, agro-morphology, fatty acid composition and RAPD marker diversity in safflower (Carthamus tinctorius). Genet. Resour. Crop Evol. 2009, 56, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Lexer, C.; Welch, M.E.; Durphy, J.L.; Rieseberg, L.H. Natural selection for salt tolerance quantitative trait loci (QTLs) in wild sunflower hybrids: Implications for the origin of Helianthus paradoxus, a diploid hybrid species. Mol. Ecol. 2003, 12, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Lexer, C.; Lai, Z.; Rieseberg, L.H. Candidate gene polymorphisms associated with salt tolerance in wild sunflower hybrids: Implications for the origin of Helianthus paradoxus, a diploid hybrid species. New Phytol. 2004, 161, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelist, C.; Lexer, C.; Dillmann, C.; Sicard, D.; Rieseberg, L.H. Microsatellite signature of ecological selection for salt tolerance in a wild sunflower hybrid species, Helianthus paradoxus. Mol. Ecol. 2006, 15, 4623–4634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteros, M.J. Identification and confirmation of QTL conditioning drought tolerance in nepalese soybean. In Proceedings of the 11th Biennial Conference on the Molecular and Cellular Biology of the Soybean, Lincoln, NE, USA, 5–8 August 2006. [Google Scholar]
- Bhatnagar, S.; King, C.A.; Purcell, L.; Ray, J.D. Identification and mapping of quantitative trait loci associated with crop response to water-deficit stress in soybean [Glycine max (L.) Merr.]. In Proceedings of the ASA-CSSA-SSSA International Annual Meeting Poster Abstract, Salt Lake City, UT, USA, 6–10 November 2005. [Google Scholar]
- Specht, J.E.; Chase, K.; Macrander, M.; Graef, G.L.; Chung, J.; Markwell, J.P.; Germann, M.; Orf, J.H.; Lark, K.G. Soybean response to water: A QTL analysis of drought tolerance. Crop Sci. 2001, 41, 493–509. [Google Scholar] [CrossRef]
- Jegadeesan, S.; Yu, K.; Poysa, V.; Gawalko, E.; Morrison, M.J.; Shi, C.; Cober, E. Mapping and validation of simple sequence repeat markers linked to a major gene controlling seed cadmium accumulation in soybean [Glycine max (L.) Merr]. Theor. Appl. Genet. 2010, 121, 283–294. [Google Scholar] [CrossRef]
- Kassem, M.A.; Meksem, K.; Kang, C.H.; Njiti, V.N.; Kilo, V.; Wood, A.J.; Lightfoot, D.A. Loci underlying resistance to manganese toxicity mapped in a soybean recombinant inbred line population of ‘Essex’ × ‘Forrest’. Plant Soil 2004, 260, 197–204. [Google Scholar] [CrossRef]
- VanToai, T.T.; St. Martin, S.K.; Chase, K.; Boru, G.; Schnipke, V.; Schmitthenner, A.F.; Lark, K.G. Identification of a QTL associated with tolerance of soybean to soil waterlogging. Crop Sci. 2001, 41, 1247–1252. [Google Scholar] [CrossRef] [Green Version]
- Githiri, S.M.; Watanabe, S.; Harada, K.; Takahashi, R. QTL Analysis of flooding tolerance in soybean at an early vegetative growth stage. Plant Breed. 2006, 125, 613–618. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Vuong, T.D.; VanToai, T.; Lee, J.D.; Wu, X.; Mian, M.A.R.; Dorrance, A.E.; Shannon, J.G.; Nguyen, H.T. Mapping of quantitative trait loci associated with resistance to Phytophthora sojae and flooding tolerance in soybean. Crop Sci. 2012, 52, 2481–2493. [Google Scholar] [CrossRef]
- Cornelious, B.; Chen, P.; Hou, A.; Shi, A.; Shannon, J.G. Yield potential and waterlogging tolerance of selected near-isogenic lines and recombinant inbred lines from two southern soybean populations. J. Crop. Improv. 2006, 16, 97–111. [Google Scholar] [CrossRef]
- Cornelious, B.; Chen, P.; Chen, Y.; de Leon, N.; Shannon, J.G.; Wang, D. Identification of QTLs underlying water-logging tolerance in soybean. Mol. Breed. 2005, 16, 103–112. [Google Scholar] [CrossRef]
- Hewezi, T.; Léger, M.; El Kayal, W.; Gentzbittel, L. transcriptional profiling of sunflower plants growing under low temperatures reveals an extensive down-regulation of gene expression associated with chilling sensitivity. J. Exp. Bot. 2006, 57, 3109–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, P.; Di Rienzo, J.; Fernandez, L.; Hopp, H.E.; Paniego, N.; Heinz, R.A. Transcriptomic Identification of candidate genes involved in sunflower responses to chilling and salt stresses based on CDNA microarray analysis. BMC Plant Biol. 2008, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funatsuki, H.; Kawaguchi, K.; Matsuba, S.; Sato, Y.; Ishimoto, M. Mapping of QTL associated with chilling tolerance during reproductive growth in soybean. Theor. Appl. Genet. 2005, 111, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.B.; Qiu, P.C.; Jiang, H.W.; Liu, C.Y.; Xin, D.W.; Li, C.D.; Hu, G.H.; Chen, Q.S. Dissection of genetic overlap of drought and low-temperature tolerance QTLs at the germination stage using backcross introgression lines in soybean. Mol. Biol. Rep. 2012, 39, 6087–6094. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Ohnishi, S.; Senda, M.; Miyoshi, T.; Ishimoto, M.; Kitamura, K.; Funatsuki, H. A novel major quantitative trait locus controlling seed development at low temperature in soybean (Glycine max). Theor. Appl. Genet. 2009, 118, 1477–1488. [Google Scholar] [CrossRef]
- Zeng, X.; Li, W.; Wu, Y.; Liu, F.; Luo, J.; Cao, Y.; Zhu, L.; Li, Y.; Li, J.; You, Q.; et al. Fine mapping of a dominant thermo-sensitive genic male sterility gene (BntsMs) in Rapeseed (Brassica napus) with AFLP- and Brassica rapa-derived PCR markers. Theor. Appl. Genet. 2014, 127, 1733–1740. [Google Scholar] [CrossRef]
- Kole, C.; Thormann, C.; Karlsson, B.; Palta, J.; Gaffney, P.; Yandell, B.; Osborn, T. Comparative mapping of loci controlling winter survival and related traits in oilseed Brassica rapa and B. napus. Mol. Breed. 2002, 9, 201–210. [Google Scholar] [CrossRef]
- Poodineh, M.; Nezhad, N.M.; Mohammadi-Nejad, G.; Fakheri, B.A.; Ebrahimi, F. Identification of safflower (Carthamus tinctorius L.) QTL under drought stress and normal conditions. Ind. Crops Prod. 2021, 171, 113889. [Google Scholar] [CrossRef]
- Dhungana, S.K.; Kim, H.-S.; Kang, B.-K.; Seo, J.-H.; Kim, H.-T.; Shin, S.-O.; Oh, J.-H.; Baek, I.-Y. Identification of QTL for tolerance to flooding stress at seedling stage of soybean (Glycine max L. Merr.). Agronomy 2021, 11, 908. [Google Scholar] [CrossRef]
- Raskin, I.; Ensley, B.D. Phytoremediation of Toxic Metals: Using Plants to Clean Up the Environment; John Wiley and Sons, Inc.: New York, NY, USA, 2000; p. 304. [Google Scholar]
- McLaughlin, M.J.; Parker, D.R.; Clarke, J.M. Metals and micronutrients—Food safety issues. Field Crops Res. 1999, 60, 143–163. [Google Scholar] [CrossRef]
- Soudek, P.; Katrusáková, A.; Sedlácek, L.; Petrová, S.; Kocí, V.; Marsík, P.; Griga, M.; Vanek, T. Effect of heavy metals on inhibition of root elongation in 23 cultivars of flax (Linum usitatissimum L.). Arch. Environ. Contam. Toxicol. 2010, 59, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Mo, Z.; Wang, H.; Wang, Z.J.; Cao, Z.H. The transportation, time-dependent distribution of heavy metals in paddy crops. Chemosphere 2003, 50, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Adverse effects of cadmium on plants and possible mitigation of Cd-induced damages. In Cadmium: Characteristics, Sources of Exposure, Health and Environmental Effects; Nova Science: New York, NY, USA, 2013; pp. 1–48. [Google Scholar]
- ul Hassan, Z.; Ali, S.; Ahmad, R.; Rizwan, M.; Abbas, F.; Yasmeen, T.; Iqbal, M. Biochemical and molecular responses of oilseed crops to heavy metal stress. In Oilseed Crops: Yield and Adaptations under Environmental Stress; Wiley & Sons: Chichester, UK, 2017; pp. 236–248. [Google Scholar] [CrossRef]
- Ashraf, M.Y.; Azhar, N.; Ashraf, M.; Hussain, M.; Arshad, M. Influence of lead on growth and nutrient accumulation in canola (Brassica napus L.) cultivars. J. Environ. Biol. 2011, 32, 659. [Google Scholar] [PubMed]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Kilo, V.; Lightfoot, D.A. Genetic analysis of resistance to manganese toxicity in soybean using molecular markers Soybean. Genet. Newsl. 1996, 23, 155–157. [Google Scholar]
- Chugh, V.; Kaur, N.; Gupta, A.K.; Rai, A. The seed biochemical signature as a potent marker for water logging tolerance in maize. Plant Stress 2022, 4, 100085. [Google Scholar] [CrossRef]
- Ahmed, F.; Rafii, M.Y.; Ismail, M.R.; Juraimi, A.S.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Waterlogging tolerance of crops: Breeding, mechanism of tolerance, molecular approaches, and future prospects. Biomed Res. Int. 2013, 2013, 963525. [Google Scholar] [CrossRef] [Green Version]
- Ploschuk, R.A.; Miralles, D.J.; Striker, G.G. A quantitative review of soybean responses to waterlogging: Agronomical, morpho-physiological and anatomical traits of tolerance. Plant Soil 2022, 475, 237–252. [Google Scholar] [CrossRef]
- Wiraguna, E. Adaptation of legume seeds to waterlogging at germination. Crops 2022, 2, 111–119. [Google Scholar] [CrossRef]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of waterlogging tolerance in plants: Research progress and prospects. Front. Plant Sci. 2020, 11, 627331. [Google Scholar] [CrossRef] [PubMed]
- Valliyodan, B.; Ye, H.; Song, L.; Murphy, M.; Shannon, J.G.; Nguyen, H.T. Genetic diversity and genomic strategies for improving drought and waterlogging tolerance in soybeans. J. Exp. Bot. 2017, 68, 1835–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ofori, A.; Becker, H.C.; Kopisch-Obuch, F.J. Effect of crop improvement on genetic diversity in oilseed Brassica rapa (Turnip-Rape) cultivars, detected by SSR markers. J. Appl. Genet. 2008, 49, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Z.; Creelman, R.A.; Zhu, J.-K. From laboratory to field. Using information from Arabidopsis to engineer salt, cold, and drought tolerance in crops. Plant Physiol. 2004, 135, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K.; Seki, M. Regulatory network of gene expression in the drought and cold stress responses. Curr. Opin. Plant Biol. 2003, 6, 410–417. [Google Scholar] [CrossRef]
- Lata, C.; Prasad, M. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 2011, 62, 4731–4748. [Google Scholar] [CrossRef] [Green Version]
- Chinnusamy, V.; Zhu, J.-K.; Sunkar, R. Gene regulation during cold stress acclimation in plants. Methods Mol. Biol. 2010, 639, 39–55. [Google Scholar]
- Chen, L.; Zhong, H.; Ren, F.; Guo, Q.-Q.; Hu, X.-P.; Li, X.-B. A novel cold-regulated gene, COR25, of Brassica napusis involved in plant response and tolerance to cold stress. Plant Cell Rep. 2011, 30, 463–471. [Google Scholar] [CrossRef]
- Kreps, J.A.; Wu, Y.; Chang, H.-S.; Zhu, T.; Wang, X.; Harper, J.F. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol. 2002, 130, 2129–2141. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Chugh, V.; Sahi, G.K.; Chhuneja, P. Wheat: Mechanisms and genetic means for improving heat tolerance. In Improving Crop Resistance to Abiotic Stress; Wiley-VCH Verlag GmbH & Co., KGaA: Weinheim, Germany, 2012; pp. 657–694. [Google Scholar]
- Singh, S.K.; Kakani, V.G.; Brand, D.; Baldwin, B.; Reddy, K.R. Assessment of cold and heat tolerance of winter-grown canola (Brassica napus L.) cultivars by pollen-based parameters. J. Agron. Crop Sci. 2008, 194, 225–236. [Google Scholar] [CrossRef]
- Sionit, N.; Strain, B.R.; Flint, E.P. Interaction of temperature and CO2 enrichment on soybean: Photosynthesis and seed yield. Can. J. Plant Sci. 1987, 67, 629–636. [Google Scholar] [CrossRef]
- Egli, D.B.; Wardlaw, I.F. Temperature response of seed growth characteristics of soybeans. Agron. J. 1980, 72, 560–564. [Google Scholar] [CrossRef]
- Maestri, E.; Klueva, N.; Perrotta, C.; Gulli, M.; Nguyen, H.T.; Marmiroli, N. Molecular genetics of heat tolerance and heat shock proteins in cereals. Plant Mol. Biol. 2002, 48, 667–681. [Google Scholar] [CrossRef]
- Gomez Selvaraj, M.; Narayana, M.; Schubert, A.M.; Ayers, J.L.; Baring, M.R.; Burow, M.D. Identification of QTLs for Pod and kernel traits in cultivated peanut by bulked segregant analysis. Electron. J. Biotechnol. 2009, 12, 3–4. [Google Scholar] [CrossRef]
- Wang, L.; Xia, Q.; Zhang, Y.; Zhu, X.; Zhu, X.; Li, D.; Ni, X.; Gao, Y.; Xiang, H.; Wei, X.; et al. Updated sesame genome assembly and fine mapping of plant height and seed coat color QTLs using a new high-density genetic map. BMC Genom. 2016, 17, 31. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Wang, Y.; Song, C.; Zhou, J.; Qiu, L.; Huang, H.; Wang, Y. A single origin and moderate bottleneck during domestication of soybean (Glycine max): Implications from microsatellites and nucleotide sequences. Ann. Bot. 2010, 106, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Wang, Z.; Tu, J.; Zhang, J.; Yu, F.; McVetty, P.B.E.; Li, G. An ultradense genetic recombination map for Brassica napus, consisting of 13551 SRAP markers. Theor. Appl. Genet. 2007, 114, 1305–1317. [Google Scholar] [CrossRef]
- Xia, Z.; Tsubokura, Y.; Hoshi, M.; Hanawa, M.; Yano, C.; Okamura, K.; Ahmed, T.A.; Anai, T.; Watanabe, S.; Hayashi, M.; et al. An integrated high-density linkage map of soybean with RFLP, SSR, STS, and AFLP markers using a single F2population. DNA Res. 2007, 14, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Tonnis, B.; Morris, B.; Wang, R.B.; Zhang, A.L.; Pinnow, D.; Wang, M.L. Variation in seed fatty acid composition and sequence divergence in the FAD2 gene coding region between wild and cultivated sesame. J. Agric. Food Chem. 2014, 62, 11706–11710. [Google Scholar] [CrossRef]
- Talukder, Z.I.; Ma, G.; Hulke, B.S.; Jan, C.-C.; Qi, L. Linkage mapping and genome-wide association studies of the Rfgene cluster in sunflower (Helianthus annuus L.) and their distribution in world sunflower collections. Front. Genet. 2019, 10, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.-S.; Yu, T.-F.; He, G.-H.; Chen, M.; Zhou, Y.-B.; Chai, S.-C.; Xu, Z.-S.; Ma, Y.-Z. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses. BMC Genom. 2014, 15, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, D.; Xiao, Y.; Yang, W.; Ye, W.; Wang, B.; Younas, M.; Wu, J.; Liu, K. Association mapping of six yield-related traits in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2014, 127, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Huang, C.; Zhang, L.; Liu, H.; Yu, J.; Hu, Z.; Hua, W. Systematic Analysis of Hsf family genes in the Brassica napus Genome reveals novel responses to heat, drought and high CO2 stresses. Front. Plant Sci. 2017, 8, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Meng, M.; Yan, Z.; Lin, Z.; Nie, X.; Yang, X. Genome-wide association mapping of stress-tolerance traits in cotton. Crop J. 2019, 7, 77–88. [Google Scholar] [CrossRef]
- Dowell, J.A.; Reynolds, E.C.; Pliakas, T.P.; Mandel, J.R.; Burke, J.M.; Donovan, L.A.; Mason, C.M. Genome-wide association mapping of floral traits in cultivated sunflower (Helianthus annuus). J. Hered. 2019, 110, 275–286. [Google Scholar] [CrossRef]
- Jiang, H.; Huang, L.; Ren, X.; Chen, Y.; Zhou, X.; Xia, Y.; Huang, J.; Lei, Y.; Yan, L.; Wan, L.; et al. Diversity characterization and association analysis of agronomic traits in a Chinese peanut (Arachis hypogaea L.) mini-core collection: Diversity characterization and association analysis of peanut. J. Integr. Plant Biol. 2014, 56, 159–169. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, Y.; Lü, H.; Li, D.; Wang, L.; Zhang, X. Association analysis for quality traits in a diverse panel of chinese sesame (Sesamum indicum L.) germplasm: Association analysis for quality. J. Integr. Plant Biol. 2013, 55, 745–758. [Google Scholar] [CrossRef]
- Yu, J.; Golicz, A.A.; Lu, K.; Dossa, K.; Zhang, Y.; Chen, J.; Wang, L.; You, J.; Fan, D.; Edwards, D.; et al. Insight into the evolution and functional characteristics of the pan-genome assembly from sesame landraces and modern cultivars. Plant Biotechnol. J. 2019, 17, 881–892. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Huang, J.; Liu, S.; Liu, X.; Li, R.; Luo, J.; Fu, Z. Improved assembly and annotation of the sesame genome. DNA Res. 2022, 29, dsac041. [Google Scholar] [CrossRef]
- Bowers, J.E.; Pearl, S.A.; Burke, J.M. Genetic mapping of millions of SNPs in safflower (Carthamus tinctorius L.) via whole-genome resequencing. G3 Genes Genomes Genet. 2016, 6, 2203–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Liu, H.; Zhan, W.; Yu, Z.; Qin, E.; Liu, S.; Yang, T.; Xiang, N.; Kudrna, D.; Chen, Y.; et al. The chromosome-scale reference genome of safflower (Carthamus tinctorius) provides insights into linoleic acid and flavonoid biosynthesis. Plant Biotechnol. J. 2021, 19, 1725–1742. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Liang, Z.; Yan, T.; Xu, Y.; Xuan, L.; Tang, J.; Zhou, G.; Lohwasser, U.; Hua, S.; Wang, H. Whole-genome resequencing of a worldwide collection of rapeseed accessions reveals the genetic basis of ecotype divergence. Mol. Plant 2019, 12, 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paritosh, K.; Pradhan, A.K.; Pental, D. A highly contiguous genome assembly of Brassica nigra (BB) and revised nomenclature for the pseudochromosomes. BMC Genom. 2020, 21, 887. [Google Scholar] [CrossRef]
- Pegadaraju, V.; Nipper, R.; Hulke, B.; Qi, L.; Schultz, Q. De novo sequencing of sunflower genome for SNP discovery using RAD (Restriction site Associated DNA) approach. BMC Genom. 2013, 14, 556. [Google Scholar] [CrossRef]
- Dimitrijevic, A.; Horn, R. Sunflower hybrid breeding: From markers to genomic selection. Front. Plant Sci. 2018, 8, 2238. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Pan, C.; Fan, W.; Liu, W.; Zhao, H.; Li, D.; Wang, S.; Hu, L.; He, B.; Qian, K. A Chromosome-level Genome Assembly of Wild Castor Provides New Insights into its Adaptive Evolution in Tropical Desert. Genom. Proteom. Bioinform. 2022, 20, 42–59. [Google Scholar] [CrossRef]
- Dmitriev, A.A.; Pushkova, E.N.; Novakovskiy, R.O.; Beniaminov, A.D.; Rozhmina, T.A.; Zhuchenko, A.A.; Bolsheva, N.L.; Muravenko, O.V.; Povkhova, L.V.; Dvorianinova, E.M. Genome sequencing of fiber flax cultivar atlant using oxford nanopore and illumina platforms. Front. Genet. 2021, 11, 590282. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.; Ren, L.; Farmer, A.D.; Pandey, M.K. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, N.; Choudhary, S.; Naaz, N.; Sharma, N.; Laskar, R.A. Recent advancements in molecular marker-assisted selection and applications in plant breeding programmes. Genet. Eng. Biotechnol. 2021, 19, 128. [Google Scholar] [CrossRef] [PubMed]
- Morgante, M.; Salamini, F. From plant genomics to breeding practice. Curr. Opin. BioTechnol. 2003, 14, 214–219. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Crop | Abiotic Stress | Yield Reduction | References |
|---|---|---|---|---|
| 1. | Mustard | Moisture stress | 17–94% | [5] |
| 2. | Mustard | Salinity | 50–90% | [6] |
| 3. | Mustard | Heat stress | 34% | [7] |
| 4. | Mustard | Heat stress | >54% | [8] |
| 5. | Soybean | Drought | 73–82% | [9] |
| 6. | Soybean | Drought | 50% | [10] |
| 7. | Soybean | Salinity | Up to 40% | [11] |
| 8. | Soybean | Flooding | Up to 25% | [12] |
| 9. | Soybean | Flooding | 20–39% | [13] |
| 10. | Soybean | Cold stress | 24% | [14] |
| 11. | Sunflower | Drought | Up to 40% | [15,16] |
| 12. | Sesame | Waterlogging | 50–90% | [17,18] |
| 13. | Sesame | Drought | 28% | [19,20] |
| 14. | Safflower | Drought | 17.2% | [21] |
| 15. | Groundnut | Drought | 55–72% | [22] |
| Mapping Population/Genotypes | Trait | Marker Used/Markers Linked to QTL | Crop | Reference |
|---|---|---|---|---|
| Hutcheson × PI471938, 140 F4 population | Drought tolerance | SSR (Satt226) | Soybean | [97] |
| Jackson × KS4895, 81 RILs | Drought tolerance | SSR (Sat_044) | Soybean | [98] |
| Minsoy × Noir 1, 236 RILs | Drought tolerance | SSR (Satt205-Satt489) | Soybean | [99] |
| S-100 × Tokyo, 116 F2 population | Drought tolerance | RFLP (A489H) | Soybean | [25] |
| Young×PI416937, 120 F4 population | Drought tolerance | RFLP (B031-1, A089-1, cr497-1, K375-1, A063-1) | Soybean | [47] |
| TAG 24 × ICGV 86031, RILs | Drought tolerance | SSRs | Groundnut | [61] |
| ICGS 76 × CSMG 84-1 and ICGS 44 × ICGS 76, RILs | Drought tolerance | SSRs | Groundnut | [62] |
| Mex.22-191 × IL.111 F3 population | Drought tolerance | SSRs and ISSRs | Safflower | [57] |
| 400 accessions including landraces and modern cultivars | Drought tolerance | SNPs | Sesame | [66] |
| S-100 × Tokyo RILs | Salt stress | SSRs (Sat_091) | Soybean | [86] |
| FT-Abyara × C01 and Jindou No. 690197, RILs | Salt stress | SSRs (Sat_091) | Soybean | [87] |
| Kefeng No. 1 × Nannong1138-2, RILs | Salt stress | SSRs (Sat_164 and Sat_358) | Soybean | [88] |
| Jackson (PI548657) × JWS156-1, F2 | Salt stress | SSRs | Soybean | [89] |
| 490 accessions | Drought and salt stress | SNPs | Sesame | [68] |
| AC Hime × Westag-97, RILs | Cadmium toxicity | SSRs (SatK147, SacK149, and SattK152) | Soybean | [100] |
| Essex × Forest, RILs | Manganese toxicity | SSRs (Satt291, Satt239, OEO2) | Soybean | [101] |
| Archer × ‘Minsoy and Archer × Noir I’, RILs | Flooding | SSRs (Sat_064) | Soybean | [102] |
| Misuzudaizu × Moshidou Gong 503, RILs | Flooding | SSRs | Soybean | [103] |
| S99-2281 × PI 408105A, RILs | Flooding | SSRs and SNPs (Sct_033, BARC-024569-04982, BARC-016279-02316) | Soybean | [104] |
| A5403 × Archer (Population 1) × P9641 × Archer (Population 2), F6:11 RILs | Flooding | SSRs (Satt385, Satt269, Sat_064) | Soybean | [105,106] |
| Santiago II and Melody | Cold stress | ESTs | Sunflower | [107] |
| HA89 | Cold stress | ESTs | Sunflower | [108] |
| Hayahikari × Toyomusume, RILs | Cold stress | SSRs | Soybean | [109] |
| Hongfeng11 × Harosoy, BC2F3 | Cold stress | SSRs | Soybean | [110] |
| RILs | Cold stress | SSRs | Soybean | [111] |
| TE5A, BC2 | Heat stress | AFLPs, SCARs | Rapeseed | [112] |
| Jinhuangma (JHM) and Zhushanbai (ZSB), landraces | Drought stress | SNPs | Sesame | [71] |
| RILs (Per × R500) and DH lines (Major × Stellar) | Cold stress | RFLPs, AFLPs | Mustard and canola | [113] |
| Mex.22−191 × Goldasht, F9 RILs | Drought | AFLPs | safflower | [114] |
| K099 × Fendou 16, F7 RILs | Drought | SSR (Sat_165 and Satt621) | Soybean | [46] |
| NTS116 × Danbaekkong, RILs | Flooding | SNPs | Soybean | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chugh, V.; Kaur, D.; Purwar, S.; Kaushik, P.; Sharma, V.; Kumar, H.; Rai, A.; Singh, C.M.; Kamaluddin; Dubey, R.B. Applications of Molecular Markers for Developing Abiotic-Stress-Resilient Oilseed Crops. Life 2023, 13, 88. https://doi.org/10.3390/life13010088
Chugh V, Kaur D, Purwar S, Kaushik P, Sharma V, Kumar H, Rai A, Singh CM, Kamaluddin, Dubey RB. Applications of Molecular Markers for Developing Abiotic-Stress-Resilient Oilseed Crops. Life. 2023; 13(1):88. https://doi.org/10.3390/life13010088
Chicago/Turabian StyleChugh, Vishal, Dasmeet Kaur, Shalini Purwar, Prashant Kaushik, Vijay Sharma, Hitesh Kumar, Ashutosh Rai, Chandra Mohan Singh, Kamaluddin, and R. B. Dubey. 2023. "Applications of Molecular Markers for Developing Abiotic-Stress-Resilient Oilseed Crops" Life 13, no. 1: 88. https://doi.org/10.3390/life13010088