
Neopestalotiopsis Species Associated with Flower Diseases of Macadamia integrifolia in Australia


1
Queensland Alliance for Agriculture & Food Innovation, The University of Queensland, Ecosciences Precinct, Dutton Park, QLD 4102, Australia
2
Centre for Crop Health, University of Southern Queensland, Toowoomba, QLD 4350, Australia
3
School of Agriculture & Food Sciences, The University of Queensland, Gatton, QLD 4343, Australia
*
Authors to whom correspondence should be addressed.
J. Fungi 2021, 7(9), 771; https://doi.org/10.3390/jof7090771
Submission received: 26 August 2021
/
Revised: 9 September 2021
/
Accepted: 15 September 2021
/
Published: 17 September 2021
(This article belongs to the Section Fungal Evolution, Biodiversity and Systematics)
Abstract
:Macadamia (Macadamia integrifolia) is native to eastern Australia and produces an edible nut that is extensively cultivated in commercial orchards in several countries. Little is known about the diversity of fungi associated with diseases of macadamia inflorescences. A survey of fungi associated with the dry flower disease of macadamia detected several isolates of Neopestalotiopsis (Pestalotiopsidaceae, Sordariomycetes). Five new species of Neopestalotiopsis were identified based on molecular phylogenetic analyses of concatenated gene sequences of the internal transcribed spacer (ITS), β-tubulin (TUB), and the translation elongation factor 1-alpha (TEF1α). The new species are named Neopestalotiopsis drenthii, N. maddoxii, N. olumideae, N. vheenae, and N. zakeelii, and are described by molecular, morphological, and cultural characteristics. The ecology of the isolates and their pathogenic, saprophytic, or commensal ability were not determined.
1. Introduction
Macadamia (Macadamia integrifolia) is a tree nut crop that is cultivated for its high-value kernel in tropical and subtropical regions in Australia, Asia, Africa, South America, and the U.S.A. Four species, M. integrifolia, M. tetraphylla, M. ternifolia, and M. jansenii, are native to Australia [1]. Macadamia integrifolia and M. tetraphylla produce edible kernels, whereas M. ternifolia and M. jansenii produce small and inedible nuts that contain high levels of cyanogenic glycosides [2]. Several new diseases caused by fungal and oomyceteous pathogens have been reported on macadamia with the expansion of its production area [3,4,5].
Diseases of flowers and fruit result in significant yield losses and poor-quality kernels [6,7,8]. A mature macadamia tree may produce over 10,000 racemes (inflorescences) at peak anthesis, with each raceme typically having 100–300 flowers (Figure 1a) [9,10]. Flower diseases can result in poor pollination efficiency, with less than 1% of the flowers producing fruit, as well as a reduction in the potential of the flower to bear fruit. Macadamia racemes may be affected by fungal pathogens at different developmental stages. Macadamia inflorescences have four growth stages: small green buds on the rachis (stage 1); florets that turn from light green to white and are partially up to fully open, with stamens that pull away from the stigmas (stage 2); fully opened flowers with sepals that turn brown at peak anthesis (stage 3); sepals that drop and flowers with swollen fertilized embryos (stage 4) [11]. Most of the diseases that affect macadamia inflorescences are flower blights [12]. A diversity of fungi has been associated with flower blights of macadamia in Australia [13], including species of Botrytis [14], Cladosporium [15], Neopestalotiopsis, and Pestalotiopsis [8].
The incidence of macadamia dry flower disease caused by Pestalotiopsis macadamiae and Neopestalotiopsis macadamiae is on the increase in Australian macadamia plantations [8]. Dry flower disease is characterized by the necrotic blight of flowers (Figure 1b). Akinsanmi et al. [16] suggested that multiple species of Pestalotiopsis and Neopestalotiopsis were responsible for dry flower epidemics in Australia.
Pestalotiopsis (Pestalotiopsidaceae, Sordariomycetes) was reclassified in 2014 and two new genera, Neopestalotiopsis and Pseudopestalotiopsis, were introduced [17]. Molecular phylogenies based on the combined sequences of three gene regions, including the internal transcribed spacer (ITS) region of rDNA, β-tubulin (TUB), and the translation elongation factor 1-alpha (TEF1α), have been used to delimit species within the pestalotioid genera (Pestalotiopsis, Neopestalotiopsis, and Pseudopestalotiopsis) [17]. Morphological identification of pestalotioid species is unreliable as species often have overlapping conidial measurements [18,19].
Several species of Neopestalotiopsis are phytopathogens in tropical and subtropical regions, causing leaf spot, dry flower, fruit rot, fruit scab, and trunk diseases on a range of crops [5,8,20,21,22,23,24,25,26]. Flower and leaf diseases on macadamia caused by Neopestalotiopsis spp. have been reported in Australia [5,8], Brazil [26], and China [25]. Many new pestalotioid species have been introduced in recent years [27,28,29,30,31,32,33]. Many unidentified isolates of Neopestalotiopsis were obtained from macadamia racemes with dry flower symptoms. However, there is little information about their identity and the diversity of fungi that cause dry flower disease on macadamia in Australia. The aim of this study was to identify the species of Neopestalotiopsis associated with the dry flowers of macadamia in Australia.
2. Materials and Methods
2.1. Sample Collection and Isolation
The isolates included in this study were collected from macadamia racemes with symptoms of dry flower disease. Samples were obtained from commercial macadamia orchards in Queensland (QLD) and New South Wales (NSW), Australia in 2019 and 2020. The samples were surface sterilized and incubated, as described by Akinsanmi et al. [8]. Monoconidial cultures of 13 isolates were established, as described by Akinsanmi et al. [34], and cryopreserved in a sterile 15% glycerol solution at −80 °C. Living cultures of the isolates were deposited in the Queensland Plant Pathology Herbarium (BRIP), Brisbane, Australia.
2.2. Cultural and Morphological Studies
Colony characteristics of cultures on ½-potato dextrose agar (PDA; Difco Laboratories, Franklin Lakes, NJ, USA.) medium were recorded after 7 d incubation at 25 °C. Fungal morphology was recorded from colonies grown in the dark for 14 d at 25 °C on PDA as well as on autoclaved pine needles on water agar. Fungal structures were examined in lactic acid on slide mounts under a Leica DM5500B compound microscope (Wetzlar, Germany) with Nomarski differential interference contrast illumination, and images were taken with a Leica DFC 500 camera. Measurements of at least 30 conidia and other fungal structures were taken at 1000× magnification. Novel species were registered in MycoBank [35].
2.3. DNA Extraction, PCR Amplification, and Sequencing
Genomic DNA was extracted from approx. 40 mg mycelium from colonies grown on PDA for 14 d. The mycelium was homogenized using TissueLyser (Qiagen, Chadstone, Australia) for 2 min at 30 Hz, and DNA was extracted using the BioSprint 96 DNA Plant Kit on a robotic platform (Qiagen, Chadstone, Australia). DNA concentration was determined with a BioDrop Duo spectrophotometer (BioDrop, Cambridge, England) and adjusted to 10 ng µL−1. The DNA of each isolate served as the template for the PCR amplifications using the reactions and thermal cyclic conditions described by Prasannath et al. [5]. Briefly, each reaction was performed in a 25 μL reaction volume, with 1 μL each of 10 μM forward and reverse primers, PCR reaction mix, and 2 μL of DNA template. PCR amplification was performed in SuperCycler Thermal Cycler (Kyratec, Wembley, Australia) at 95 °C for 2 min, followed by 35 cycles at 95 °C for 30 s, 55 °C for 30 s, and at 72 °C for 1 min, with a final extension step at 72 °C for 5 min. Three loci, ITS, TUB, and TEF1α, were amplified and sequenced using the primer pairs ITS4/ITS5 [36], BT2A/BT2B [37], and EF1-526F/EF1-1567R [38], respectively. The quality of PCR amplicons was checked on 1% agarose gel electrophoresis stained with GelRed (Biotium, Melbourne, Australia) under UV light by Molecular Imager GelDoc (Bio-Rad Laboratories Inc., Gladesville, Australia). The targeted PCR products were purified and sequenced in both directions at Macrogen Inc. (Seoul, South Korea).
2.4. Phylogenetic Analyses
The DNA sequences were assembled in Geneious Prime v. 2021.0.3 (Biomatters Ltd., San Diego, CA, USA.), manually trimmed and aligned to produce consensus sequences for each locus. The consensus sequences generated in this study were deposited in GenBank (Table 1). The sequences were compared against the NCBI GenBank nucleotide database using BLASTn to check the closest phylogenetic matches. The sequences of the ex-type isolates of the Neopestalotiopsis species were retrieved from GenBank (Table 1) and aligned with the sequences generated from our isolates using MAFFT v. 7.3.8.8 [39] in Geneious. Ambiguously aligned positions in each multiple alignment were excluded using Gblocks v. 0.91b [40]. The concatenated three-locus sequence dataset (ITS + TEF1α + TUB) of Neopestalotiopsis consisted of 63 taxa, with the outgroup taxon Pestalotiopsis diversiseta MFLUCC 12-0287 (Table 1). The combined sequence data matrix was manually improved with BioEdit v. 7.2.5 [41] and gaps were treated as missing data. Phylogenetic trees were generated from Maximum Likelihood (ML), Bayesian Inference (BI), and Maximum Parsimony (MP) analyses.
ML analysis was implemented using RAxML v. 8.2.11 [50] in Geneious. The search option was set to rapid bootstrapping, and the analysis was run using the GTR-GAMMAI evolution model with 1000 bootstrap iterations. BI analysis was conducted with MrBayes v. 3.2.1 [51] in Geneious to calculate posterior probabilities by the Markov Chain Monte Carlo (MCMC) method. The GTR-GAMMAI nucleotide substitution model was applied in BI analysis. Four MCMC chains were run simultaneously, starting from random trees for 1,000,000 generations. The temperature of the heated chain was set to 0.15 and trees were sampled every 200 generations until the average standard deviation of split frequencies reached 0.01 (stop value). Burn-in was set at 25%, after which the likelihood values were stationary. MP analysis was performed with PAUP v. 4.0b10 [52]. Trees were inferred using a heuristic search strategy, with 100 random stepwise addition and tree-bisection-reconnection (TBR) branch swapping. Max-trees were set to 5000 and bootstrap support values were evaluated for tree branches with 1000 replications [53]. Phylograms were visualized in FigTree v. 1.4.4 [54] and annotated in Adobe Illustrator 2021.
The Genealogical Concordance Phylogenetic Species Recognition (GCPSR) concept and a pairwise homoplasy index (PHI) test were used to determine species boundaries [55]. The PHI test was performed using SplitsTree4 v. 4.17.1 [56] to determine the recombination level within phylogenetically closely related species. The concatenated three-locus dataset (ITS + TEF1α + TUB) was used for the analyses. PHI test results (Fw) >0.05 indicated no significant recombination within the dataset. The relationships between closely related taxa were visualized in split graphs with both the Log-Det transformation and splits decomposition options.
3. Results
3.1. Phylogenetic Analyses
The concatenated sequence data matrix comprised 1367 base pairs (bp) (476 for ITS, 464 for TEF1α, and 427 for TUB), of which 935 bp were constant, 238 bp were parsimony-uninformative, and 174 bp were parsimony-informative. ML analysis yielded a best scoring tree, with a final ML optimization value of −6493.745 and the following model parameters: alpha—0.597, Π(A)—0.231, Π(C)—0.276, Π(G)—0.218, and Π(T)—0.274. Similar tree topologies were obtained by ML, BI, and MP methods, and the best scoring ML tree is shown in Figure 2. ML bootstrap values, BI posterior probabilities, and MP bootstrap values (MLBS/BIPP/MPBS) are given at nodes of the phylogram (Figure 2). The phylogenetic tree inferred from the concatenated alignment resolved the 13 Neopestalotiopsis isolates from symptomatic macadamia inflorescences (dry flower disease) into five well-supported monophyletic clades that represent novel species of Neopestalotiopsis (Figure 2).
3.2. Taxonomy
Neopestalotiopsis drenthii Prasannath, Akinsanmi & R.G. Shivas, sp. nov. (Figure 3).
MycoBank: MB840916.
Etymology: Named after Andre Drenth, in recognition of his many contributions to the study of tropical and subtropical plant diseases.
Type: AUSTRALIA, Queensland, Mackay, from flower blight of M. integrifolia, 3 October 2019, O.A. Akinsanmi (Holotype BRIP 72264a, includes ex-type culture). GenBank: MZ303787 (ITS); MZ312680 (TUB); MZ344172 (TEF1α).
Description: Conidiomata pycnidial on PDA, globose, 200–400 µm diam., solitary or aggregated in clusters, exudes black conidial masses. Conidiophores reduced to conidiogenous cells. Conidiogenous cells ampulliform, hyaline, smooth, 5–20 × 2–5 μm. Conidia fusiform to ellipsoidal, straight or curved, 24–30 × 7–9 µm, 4-septate; basal cell conical, 4–6.5 µm, hyaline, smooth, thin-walled; with a single appendage filiform, unbranched, centric, 4–7 µm long; three median cells doliiform, 16–19 µm, smooth, versicolored, septa darker than the rest of the cell (second cell from base pale brown, 3.5–6.5 μm long; third cell medium to dark brown, 3.5–6.5 μm long; fourth cell medium to dark brown, 4–6 μm long); apical cell conical to subcylindrical, 3–5 µm long, hyaline, smooth, thin-walled; with 2–3 apical tubular appendages unbranched, filiform, 15–22 µm long. Sexual morph not seen.
Culture characteristics: Colonies on PDA after 7 d at 25 °C reach 80 mm diam., producing white aerial mycelia with copious pycnidia after two weeks; reverse cream.
Habitat and distribution: Racemes of M. integrifolia (Proteaceae); Australia.
Other material examined: AUSTRALIA, Queensland, Mackay, from flower blight of M. integrifolia, 3 October 2019, O.A. Akinsanmi (living culture, BRIP 72263a).
Notes: Neopestalotiopsis drenthii is closely related to N. surinamensis. A pairwise nucleotide comparison between N. drenthii ex-type strain (BRIP 72264a) and N. surinamensis ex-type strain (CBS 450.74) showed 2 bp differences (Identities 534/536, 2 gaps) in ITS, 0 bp differences (Identities 428/428, no gaps) in TUB, and 7 bp differences (Identities 474/481, no gaps) in TEF1α sequences in GenBank. Neopestalotiopsis drenthii and N. surinamensis have similar sized conidia, but tubular apical appendages of N. drenthii are shorter than the 18–27 µm of N. surinamensis [17].
Neopestalotiopsis maddoxii Prasannath, Akinsanmi & R.G. Shivas, sp. nov. (Figure 4).
MycoBank: MB840917.
Etymology: Named after Craig Maddox, in recognition of his research contributions to macadamia crop protection in Australia.
Type: AUSTRALIA, Queensland, Alloway, from flower blight of M. integrifolia, 22 Sep. 2019, Tim O’ Dale (Holotype BRIP 72266a, includes ex-type culture). GenBank: MZ303782 (ITS); MZ312675 (TUB); MZ344167 (TEF1α).
Description: Conidiomata pycnidial on PDA, globose, 200–500 µm diam., solitary or aggregated in clusters, exudes dark slimy conidial droplets. Conidiophores reduced to conidiogenous cells. Conidiogenous cells ampulliform, hyaline, smooth, 5–15 × 2–5 μm. Conidia fusiform to clavate, straight or curved, 25–30 × 7–11 µm, 4-septate; basal cell conical, 3–5.5 µm, hyaline, smooth, thin-walled; with a single appendage filiform, unbranched, centric, 4–7 µm long; three median cells doliiform, 18–23 µm, smooth, versicolored, septa darker than the rest of the cell (second cell from base pale brown, 4.5–7.5 μm long; third cell medium to dark brown, 4.5–7.5 μm long; fourth cell medium to dark brown, 5–7 μm long); apical cell subcylindrical, 3–5 µm long, hyaline, smooth, thin-walled; with 3 apical tubular appendages unbranched, filiform, 15–27 µm long. Sexual morph not seen.
Culture characteristics: Colonies on PDA after 7 d at 25 °C reach 70 mm diam., with cream aerial mycelium, forming abundant pycnidia near the center after two weeks; reverse cream to buff.
Habitat and distribution: Racemes of M. integrifolia (Proteaceae); Australia.
Other material examined: AUSTRALIA, Queensland, Nambour, from flower blight of M. integrifolia, 22 Aug. 2019, K. Prasannath (living culture, BRIP 72260a); Bundaberg, from flower blight of Macadamia sp., 25 August 2019, O.A. Akinsanmi (living culture, BRIP 72262a); Gympie, from flower blight of M. integrifolia, 12 September 2019, M. Boote (living culture, BRIP 72272a); Maleny, from flower blight of M. integrifolia, 20 September 2019, O.A. Akinsanmi (living cultures, BRIP 72275a and BRIP 72284a).
Notes: Neopestalotiopsis maddoxii was sister to a clade containing N. javaensis, N. mesopotamica, and N. rosae. Neopestalotiopsis maddoxii differed from N. javaensis in ITS (Identities 534/539, 3 gaps); TUB (Identities 429/431, no gaps); TEF1α (Identities 480/486, 4 gaps). Neopestalotiopsis maddoxii differed from N. mesopotamica in ITS (Identities 534/539, 3 gaps); TUB (Identities 410/414, no gaps); TEF1α (Identities 466/472, no gaps). Neopestalotiopsis maddoxii differed from N. rosae in ITS (Identities 534/539, 3 gaps); TUB (Identities 430/432, no gaps); TEF1α (Identities 478/481, no gaps). Neopestalotiopsis maddoxii is morphologically indistinguishable from N. javaensis, N. mesopotamica, and N. rosae [17].
Neopestalotiopsis olumideae Prasannath, Akinsanmi & R.G. Shivas, sp. nov. (Figure 5).
MycoBank: MB840918.
Etymology: Named after Olumide Jeff-Ego, in recognition of her research contributions to macadamia diseases in Australia.
Type: AUSTRALIA, Queensland, Maleny, from flower blight of M. integrifolia, 20 Sep. 2019, O.A. Akinsanmi (Holotype BRIP 72273a, includes ex-type culture). GenBank: MZ303790 (ITS); MZ312683 (TUB); MZ344175 (TEF1α).
Description: Conidiomata pycnidial on PDA, globose, 200–400 µm diam., mostly solitary. Conidiophores reduced to conidiogenous cells. Conidiogenous cells ampulliform to cylindrical, hyaline, smooth, 5–15 × 2–5 μm. Conidia fusiform to ellipsoidal, straight or curved, 27–31 × 7–9 µm, 4-septate; basal cell conical, 4.5–7 µm, hyaline, smooth, thin-walled; with a single appendage filiform, unbranched, centric, 3–5 µm long; three median cells doliiform, 18–20 µm, smooth, versicolored, septa darker than the rest of the cell (second cell from base pale brown, 4–7 μm long; third cell medium to dark brown, 4–7 μm long; fourth cell medium to dark brown, 4.5–6.6 μm long); apical cell conical to subcylindrical, 3.5–5.5 µm long, hyaline, smooth, thin-walled; with 2–3 apical tubular appendages unbranched, filiform, 8–17 µm long. Sexual morph not seen.
Culture characteristics: Colonies on PDA after 7 d at 25 °C reach 60 mm diam., with whitish cottony aerial mycelium; reverse cream to buff.
Habitat and distribution: Racemes of M. integrifolia (Proteaceae); Australia.
Other material examined: AUSTRALIA, Queensland, Maleny, from flower blight of M. integrifolia, 20 September 2019, O.A. Akinsanmi (living culture, BRIP 72283a).
Notes: Neopestalotiopsis olumideae was phylogenetically close to N. protearum, N. acrostichi and N. pernambucana. Neopestalotiopsis olumideae differed from N. protearum in ITS (Identities 517/521, no gaps); TUB (Identities 436/438, no gaps); TEF1α (Identities 469/475, 3 gaps). Neopestalotiopsis olumideae differed from N. acrostichi in ITS (Identities 506/506, no gaps); TUB (Identities 435/436, no gaps); TEF1α (Identities 464/471, 1 gap). Neopestalotiopsis olumideae is morphologically indistinguishable from N. protearum, N. acrostichi, and N. pernambucana [17,30].
Neopestalotiopsis vheenae Prasannath, Akinsanmi & R.G. Shivas, sp. nov. (Figure 6).
MycoBank: MB840919.
Etymology: Named after Vheena Mohankumar, in recognition of her research studies into fungal diseases of macadamia crops in Australia.
Type: AUSTRALIA, New South Wales, Rosebank, from flower blight of M. integrifolia, 16 October 2019, P. Fraser (Holotype BRIP 72293a, includes ex-type culture). GenBank: MZ303792 (ITS); MZ312685 (TUB); MZ344177 (TEF1α).
Description: Conidiomata pycnidial on PDA, globose, 200–500 µm diam., solitary or aggregated in clusters, exudes black slimy conidial droplets. Conidiophores reduced to conidiogenous cells. Conidiogenous cells ampulliform to cylindrical, hyaline, smooth, 5–10 × 3–5 μm. Conidia fusiform to clavate, straight or curved, 22–26 × 8–11 µm, 4-septate; basal cell conical, 3–5 µm, hyaline, smooth, thin-walled; with a single appendage filiform, unbranched, centric, 4–6.5 µm long; three median cells doliiform, 17–20 µm, smooth, versicolored, septa darker than the rest of the cell (second cell from base pale brown, 4–7 μm long; third cell medium to dark brown, 4–7 μm long; fourth cell medium to dark brown, 4.5–6.6 μm long); apical cell conical to subcylindrical, 3–4.5 µm long, hyaline, smooth, thin-walled; mostly with 3 apical tubular appendages unbranched, filiform, 15–25 µm long. Sexual morph not seen.
Culture characteristics: Colonies on PDA after 7 d at 25 °C reach 80 mm diam., with white dense aerial mycelium, pycnidia abundant; reverse buff.
Habitat and distribution: Racemes of M. integrifolia (Proteaceae); Australia.
Notes: Neopestalotiopsis vheenae ex-type strain (BRIP 72293a) had identical ITS, TUB, and TEF1α sequences to isolate BRIP 70210, which was previously identified as N. clavispora [5]. Neopestalotiopsis vheenae causes yellow halo spot of macadamia in Australia [5]. In this phylogenetic analysis, N. vheenae was sister to N. sichuanensis. BLASTn searches in GenBank showed that N. vheenae ex-type (BRIP 72293a) differed from N. sichuanensis ex-type (CFCC 5338) by 1 bp (Identities 510/511, no gaps) in ITS, 1 bp (Identities 422/423, no gaps) in TUB, and 10 bp (Identities 482/492, 6 gaps) in TEF1α sequences. Neopestalotiopsis vheenae is morphologically indistinguishable from N. sichuanensis [33].
Neopestalotiopsis zakeelii Prasannath, Akinsanmi & R.G. Shivas, sp. nov. (Figure 7).
MycoBank: MB840920.
Etymology: Named after Mohamed Cassim Mohamed Zakeel, in recognition of his research into the diagnosis of new and emerging diseases on macadamia in Australia.
Type: AUSTRALIA, Queensland, Landsborough, from flower blight of M. integrifolia, 20 September 2019, A. Woodford (Holotype BRIP 72282a, includes ex-type culture). GenBank: MZ303789 (ITS); MZ312682 (TUB); MZ344174 (TEF1α).
Description: Conidiomata pycnidial on PDA, globose, 100–250 µm diam., mostly solitary. Conidiophores reduced to conidiogenous cells. Conidiogenous cells ampulliform, hyaline, smooth, 5–20 × 2–5 μm. Conidia fusiform to ellipsoidal, straight or curved, 27–33 × 7–9 µm, 4-septate; basal cell conical, 4.5–7 µm, hyaline, smooth, thin-walled; with a single appendage filiform, unbranched, centric, 7–10 µm long; three median cells doliiform, 17–22 µm, smooth, versicolored, septa darker than the rest of the cell (second cell from base pale brown, 4.5–7.5 μm long; third cell medium to dark brown, 4.5–7.5 μm long; fourth cell medium to dark brown, 5–7 μm long); apical cell conical to subcylindrical, 3–5.5 µm long, hyaline, smooth, thin-walled; with 2–3 apical tubular appendages unbranched, filiform, 25–37 µm long. Sexual morph not seen.
Culture characteristics: Colonies on PDA after 7 d at 25 °C reach 55 mm diam., with white sparse aerial mycelium; reverse cream.
Habitat and distribution: Racemes of M. integrifolia (Proteaceae); Australia.
Other material examined: AUSTRALIA, Queensland, Nambour, from flower blight of M. integrifolia, 22 August 2019, K. Prasannath (living culture, BRIP 72271a).
Notes: Neopestalotiopsis zakeelii is most closely related to N. vitis and N. australis. Neopestalotiopsis zakeelii (ex-type: BRIP 72282a) differed from N. vitis (ex-type: MFLUCC 15-1265) by 4 bp (Identities 416/420, no gaps) in ITS; 0 bp (Identities 319/319, no gaps) in TUB; 4 bp (Identities 369/373, no gaps) in TEF1α. Neopestalotiopsis zakeelii differed from N. australis (ex-type: CBS 114159) by 4 bp (Identities 526/530, no gaps) in ITS; 3 bp (Identities 430/433, no gaps) in TUB; 6 bp (Identities 478/484, no gaps) in TEF1α. Neopestalotiopsis australis, N. vitis, and N. zakeelii are morphologically indistinguishable [17,22].
There was no evidence of significant genetic recombination (Fw > 0.05) between the novel species of Neopestalotiopsis and closely related species (Figure 8). The results confirmed that these taxa were significantly different from the existing species of Neopestalotiopsis.
4. Discussion
Five novel species, Neopestalotiopsis drenthii, N. maddoxii, N. olumideae, N. vheenae, and N. zakeelii, were discovered in isolates obtained from macadamia inflorescences with dry flower disease and subsequently described. There was no evidence of significant genetic recombination events between these species and their closest relatives.
Pestalotioid fungi (Pestalotiopsidaceae, Sordariomycetes) are species-rich asexual taxa, which are common pathogens on many crops [30,32,57]. Multi-locus phylogenetic analyses segregated Neopestalotiopsis and Pseudopestalotiopsis from Pestalotiopsis [17]. These three genera are morphologically similar in having 5-celled conidia with tubular apical appendages [17]. Norphanphoun et al. [30] found that concatenated gene sequences of ITS, TUB, and TEF1α resolved Pestalotiopsis and Pesudopestalotiopsis, while additional genes may be required to provide a better delimitation of Neopestalotiopsis spp.
There were 49 species names recognized in Neopestalotiopsis [58] prior to the five species described in this study. Neopestalotiopsis drenthii, N. maddoxii, N. olumideae, N. vheenae, and N. zakeelii formed well-supported monophyletic clades in the phylogenetic analysis. The topology of our phylogeny is similar to those generated in earlier studies [29,57].
Pestalotiopsis macadamiae and N. macadamiae were first reported as the causal fungi of dry flower disease of macadamia by Akinsanmi et al. [8]. These fungi are considered endemic to Australia and have likely co-evolved with macadamia. Pestalotiopsis macadamiae has been reported outside Australia on macadamia leaves in China [59]. The present study found a high diversity of Neopestalotiopsis spp. on macadamia racemes with dry flower symptoms. It is not known whether Neopestalotiopsis drenthii, N. maddoxii, N. olumideae, N. vheenae, and N. zakeelii are pathogens or saprobes. The role of these fungi in the pathogenicity of dry flower disease remains to be demonstrated by Koch’s postulates. Living cultures of N. drenthii, N. maddoxii, N. olumideae, N. vheenae, and N. zakeelii are accessible in the BRIP culture collection for future research and comparative studies that may lead to the elucidation of their lifecycle and their role in dry flower disease.
5. Conclusions
Five fungal species, Neopestalotiopsis drenthii, N. maddoxii, N. olumideae, N. vheenae, and N. zakeelii, were described and illustrated. These fungi were isolated from inflorescences of macadamia with dry flower disease in Australia. The role that N. drenthii, N. maddoxii, N. olumideae, N. vheenae, and N. zakeelii play in dry flower disease of macadamia has yet to be determined. Hence, the pathogenicity of these novel species on macadamia racemes should be examined. Living cultures of the fungi are available for further study.
Author Contributions
Methodology, data curation, molecular analysis, and writing—original draft: K.P.; taxonomy—review and editing: R.G.S.; supervision and writing—review and editing: V.J.G.; fund acquisition, lead researcher, research project administration, research concept, supervision, and writing—review and editing: O.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
K.P. is a recipient of the University of Queensland Research Training Scholarship. The research was funded by Hort Innovation using the macadamia research and development levy and funds from the Australian Government—Project No. MC16018.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequence data are available in NCBI GenBank following the accession numbers in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hardner, C.M.; Peace, C.; Lowe, A.J.; Neal, J.; Pisanu, P.; Powell, M.; Schmidt, A.; Spain, C.; Williams, K. Genetic resources and domestication of macadamia. In Horticultural Reviews; Janick, J., Ed.; John Wiley: New York, NY, USA, 2009; pp. 1–125. [Google Scholar]
- Garg, M.L.; Blake, R.J.; Wills, R.B.; Clayton, E.H. Macadamia nut consumption modulates favourably risk factors for coronary artery disease in hypercholesterolemic subjects. Lipids 2007, 42, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Jeff-Ego, O.S.; Akinsanmi, O.A. Botryosphaeriaceae causing branch dieback and tree death of macadamia in Australia. Australas. Plant Pathol. 2019, 48, 59–64. [Google Scholar] [CrossRef]
- Jeff-Ego, O.S.; Drenth, A.; Topp, B.; Henderson, J.; Akinsanmi, O.A. Prevalence of Phytophthora species in macadamia orchards in Australia and their ability to cause stem canker. Plant Pathol. 2020, 69, 1270–1280. [Google Scholar] [CrossRef]
- Prasannath, K.; Galea, V.J.; Akinsanmi, O.A. Characterisation of leaf spots caused by Neopestalotiopsis clavispora and Colletotrichum siamense in macadamia in Australia. Eur. J. Plant Pathol. 2020, 156, 1219–1225. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Drenth, A. Economic returns from fungicide application to control husk spot of macadamia in Australia is influenced by spray efficiency, rates and costs of application. Crop Prot. 2012, 41, 35–41. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Drenth, A. Characterisation of husk rot in macadamia. Ann. Appl. Biol. 2017, 170, 104–115. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Nisa, S.; Jeff-Ego, O.S.; Shivas, R.G.; Drenth, A. Dry flower disease of macadamia in Australia caused by Neopestalotiopsis macadamiae sp. nov. and Pestalotiopsis macadamiae sp. nov. Plant Dis. 2017, 101, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Trueman, S.J. The reproductive biology of macadamia. Sci. Hortic. 2013, 150, 354–359. [Google Scholar] [CrossRef]
- Trueman, S.J.; Turnbull, C.G.N. Effects of cross-pollination and flower removal on fruit set in macadamia. Ann. Bot. 1994, 73, 23–32. [Google Scholar]
- Fitzell, R.D. Diseases and Disorders of Macadamias; NSW Agriculture: Wollongbar, NSW, Australia, 1994; pp. 1–31.
- Akinsanmi, O. Flower blights of macadamia caused by Botrytis cinerea, Pestalotiopsis macadamiae and Neopestalotiopsis macadamiae in Australia. In Proceedings of the International Congress of Plant Pathology (ICPP) 2018, Boston, MA, USA, 29 July–3 August 2018. [Google Scholar]
- Akinsanmi, O. Macadamia flower blights caused by a complex of pathogens: A redefinition of disease names. In Proceedings of the 7th International Macadamia Symposium, Kruger National Park, South Africa, 11–13 August 2015. [Google Scholar]
- Hunter, J.E.; Kunimoto, R.K. Reduction of macadamia nut set by Botrytis cinerea. Phytopathology 1973, 63, 939–941. [Google Scholar] [CrossRef]
- van den Berg, N.; Serfontein, S.; Christie, B.; Munro, C. First report of raceme blight caused by Cladosporium cladosporioides on macadamia nuts in South Africa. Plant Dis. 2008, 92, 484. [Google Scholar] [CrossRef]
- Akinsanmi, O.; Nisa, S.; Jeff-Ego, O.; Drenth, A. Multiple Pestalotiopsis and Neopestalotiopsis species cause flower blight of macadamia in Australia. Phytopathology 2016, 106, 122. [Google Scholar]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Groenewald, J.Z.; Xu, J.; Crous, P.W. Pestalotiopsis revisited. Stud. Mycol. 2014, 79, 121–186. [Google Scholar] [CrossRef] [Green Version]
- Jeewon, R.; Liew, E.C.Y.; Simpson, J.A.; Hodgkiss, I.J.; Hyde, K.D. Phylogenetic significance of morphological characters in the taxonomy of Pestalotiopsis species. Mol. Phylogenet. Evol. 2003, 27, 372–383. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Cai, L.; Chukeatirote, E.; Wu, W.P.; Sun, X.; Hyde, K.D. A multi-locus backbone tree for Pestalotiopsis, with a polyphasic characterization of 14 new species. Fungal Divers. 2012, 56, 95–129. [Google Scholar] [CrossRef] [Green Version]
- Ayoubi, N.; Soleimani, M.J. Strawberry fruit rot caused by Neopestalotiopsis iranensis sp. nov., and N. mesopotamica. Curr. Microbiol. 2016, 72, 329–336. [Google Scholar] [CrossRef]
- Chamorro, M.; Aguado, A.; De los Santos, B. First report of root and crown rot caused by Pestalotiopsis clavispora (Neopestalotiopsis clavispora) on strawberry in Spain. Plant Dis. 2016, 100, 1495. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Liu, M.; Maharachchikumbura, S.S.N.; Zhang, W.; Xing, Q.K.; Hyde, K.D.; Nilthong, S.; Li, X.H.; Yan, J.Y. Neopestalotiopsis vitis sp. nov. causing grapevine leaf spot in China. Phytotaxa 2016, 258, 63–74. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Laringnonl, P.; Hyde, K.D.; Al-Sady, A.; Liu, Z. Characterization of Neopestalotiopsis, Pestalotiopsis and Truncatella species associated with grapevine trunk diseases in France. Phytopathol. Mediterr. 2016, 55, 380–390. [Google Scholar]
- Solarte, F.; Muñoz, C.G.; Maharachchikumbura, S.S.N.; Álvarez, E. Diversity of Neopestalotiopsis and Pestalotiopsis spp., causal agents of guava scab in Colombia. Plant Dis. 2018, 102, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Xu, G.; Zheng, F.Q.; Zhou, J.; Zheng, L.; Miao, W.G.; Xie, C.P. First report of Neopestalotiopsis clavispora causing leaf spot on macadamia (Macadamia integrifolia) in China. Plant Dis. 2019, 104, 288. [Google Scholar] [CrossRef]
- Santos, C.C.; Domingues, J.L.; Santos, R.F.; Spósito, M.B.; Santos, A.; Novaes, Q.S. First report of Neopestalotiopsis clavispora causing leaf spot on macadamia in Brazil. Plant Dis. 2019, 103, 1790. [Google Scholar] [CrossRef]
- Freitas, E.F.S.; Da Silva, M.; Barros, M.V.P.; Kasuya, M.C.M. Neopestalotiopsis hadrolaeliae sp. nov., a new endophytic species from the roots of the endangered orchid Hadrolaelia jongheana in Brazil. Phytotaxa 2019, 416, 211–220. [Google Scholar] [CrossRef]
- Kumar, V.; Cheewangkoon, R.; Gentekaki, E.; Maharachchikumbura, S.S.N.; Brahmange, R.S.; Hyde, K.D. Neopestalotiopsis alpapicalis sp. nov., a new endophyte from tropical mangrove trees in Krabi Province (Thailand). Phytotaxa 2019, 393, 251–262. [Google Scholar] [CrossRef]
- Liu, X.; Tibpromma, S.; Zhang, F.; Xu, J.; Chethana, K.W.T.; Karunarathna, S.C.; Mortimer, P.E. Neopestalotiopsis cavernicola sp. nov. from Gem Cave in Yunnan Province, China. Phytotaxa 2021, 512, 1–27. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Jayawardena, R.S.; Chen, Y.; Wen, T.C.; Meepol, W.; Hyde, K.D. Morphological and phylogenetic characterization of novel pestalotioid species associated with mangroves in Thailand. Mycosphere 2019, 10, 531–578. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal Planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef] [PubMed]
- Huanluek, N.; Jjayawardena, R.S.; Maharachchikumbura, S.S.N.; Harishchandra, D.L. Additions to pestalotioid fungi in Thailand: Neopestalotiopsis hydeana sp. nov. and Pestalotiopsis hydei sp. nov. Phytotaxa 2021, 479, 23–43. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, X.L.; Tian, C.M. Identification and characterization of leaf-inhabiting fungi from Castanea plantations in China. J. Fungi 2021, 7, 64. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Mitter, V.; Simpfendorfer, S.; Backhouse, D.; Chakraborty, S. Identity and pathogenicity of Fusarium spp. isolated from wheat fields in Queensland and northern New South Wales. Aust. J. Agric. Res. 2004, 55, 97–107. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S. Primers for Elongation Factor 1-α (EF1-α); Insect Biocontrol Laboratory USDA, ARS, PSI: Beltsville, MD, USA, 2001; p. 4.
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; Claverie, J.M.; Gascuel, O. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bezerra, J.D.P.; Machado, A.R.; Firmino, A.L.; Rosado, A.W.C.; Souza, C.A.F.; Souza-Motta, C.M.; Freire, K.T.L.S.; Paiva, L.M.; Magalhaes, O.M.C.; Pereira, O.L.; et al. Mycological diversity description I. Acta Bot. Bras. 2018, 32, 656–666. [Google Scholar] [CrossRef] [Green Version]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Ma, X.Y.; Maharachchikumbura, S.S.N.; Chen, B.W.; Hyde, K.D.; McKenzie, E.H.C.; Chomnunti, P.; Kang, J.C. Endophytic pestalotiod taxa in Dendrobium orchids. Phytotaxa 2019, 419, 268–286. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvarado, P.; Edwards, J.; Moreno, G.; Sharma, R.; et al. Fungal Planet description sheets: 371–399. Persoonia 2015, 35, 264–327. [Google Scholar] [CrossRef]
- Silvério, M.L.; Cavalcanti, M.A.Q.; Silva, G.A.; Oliveira, R.J.V.; Bezerra, J.L. A new epifoliar species of Neopestalotiopsis from Brazil. Agrotropica 2016, 28, 151–158. [Google Scholar] [CrossRef]
- Jiang, N.; Bonthond, G.; Fan, X.L.; Tian, C.M. Neopestalotiopsis rosicola sp. nov. causing stem canker of Rosa chinensis in China. Mycotaxon 2018, 133, 271–283. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Guo, L.D.; Chukeatirote, E.; McKenzie, E.H.C.; Hyde, K.D. A destructive new disease of Syzygium samarangense in Thailand caused by the new species Pestalotiopsiss samarangensis. Trop. Plant Pathol. 2013, 38, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree: Tree Figure Drawing Tool (Version 1.4.4); Institute of Evolutionary Biology: Edinburgh, UK, 2018. [Google Scholar]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Gerardo-Lugo, S.S.; Tovar-Pedraza, J.M.; Maharachchikumbura, S.S.N.; Apodaca-Sánchez, M.A.; Beltran, H.; Correia, K.C.; Sauceda-Acosta, C.P.; Camacho-Tapia, M.; Hyde, K.D.; Marraiki, N.; et al. Characterization of Neopestalotiopsis species associated with mango grey leaf spot disease in Sinaloa, Mexico. Pathogens 2020, 9, 788. [Google Scholar] [CrossRef]
- Index Fungorum 2021. Available online: http://www.speciesfungorum.org/ (accessed on 23 August 2021).
- Senanayake, I.C.; Lian, T.T.; Mai, X.M.; Jeewon, R.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Zeng, Y.J.; Tian, S.L.; Xie, N. New geographical records of Neopestalotiopsis and Pestalotiopsis species in Guangdong Province, China. Asian J. Mycol. 2020, 3, 512–533. [Google Scholar] [CrossRef]
Figure 1.
Macadamia inflorescences (racemes). (a) Pendant racemes in tree canopy, and (b) dry flower disease.
Figure 1.
Macadamia inflorescences (racemes). (a) Pendant racemes in tree canopy, and (b) dry flower disease.

Figure 2.
Maximum Likelihood tree topology of Neopestalotiopsis based on a combined multi-locus alignment (ITS + TEF1α + TUB). Pestalotiopsis diversiseta (MFLUCC 12-0287) was used as an outgroup taxon. Maximum Likelihood bootstrap support values (>50%), Bayesian Inference posterior probabilities (>90%), and Maximum Parsimony bootstrap proportions (>50%) are displayed at the nodes, respectively. Strains of the newly described species are depicted in red.
Figure 2.
Maximum Likelihood tree topology of Neopestalotiopsis based on a combined multi-locus alignment (ITS + TEF1α + TUB). Pestalotiopsis diversiseta (MFLUCC 12-0287) was used as an outgroup taxon. Maximum Likelihood bootstrap support values (>50%), Bayesian Inference posterior probabilities (>90%), and Maximum Parsimony bootstrap proportions (>50%) are displayed at the nodes, respectively. Strains of the newly described species are depicted in red.

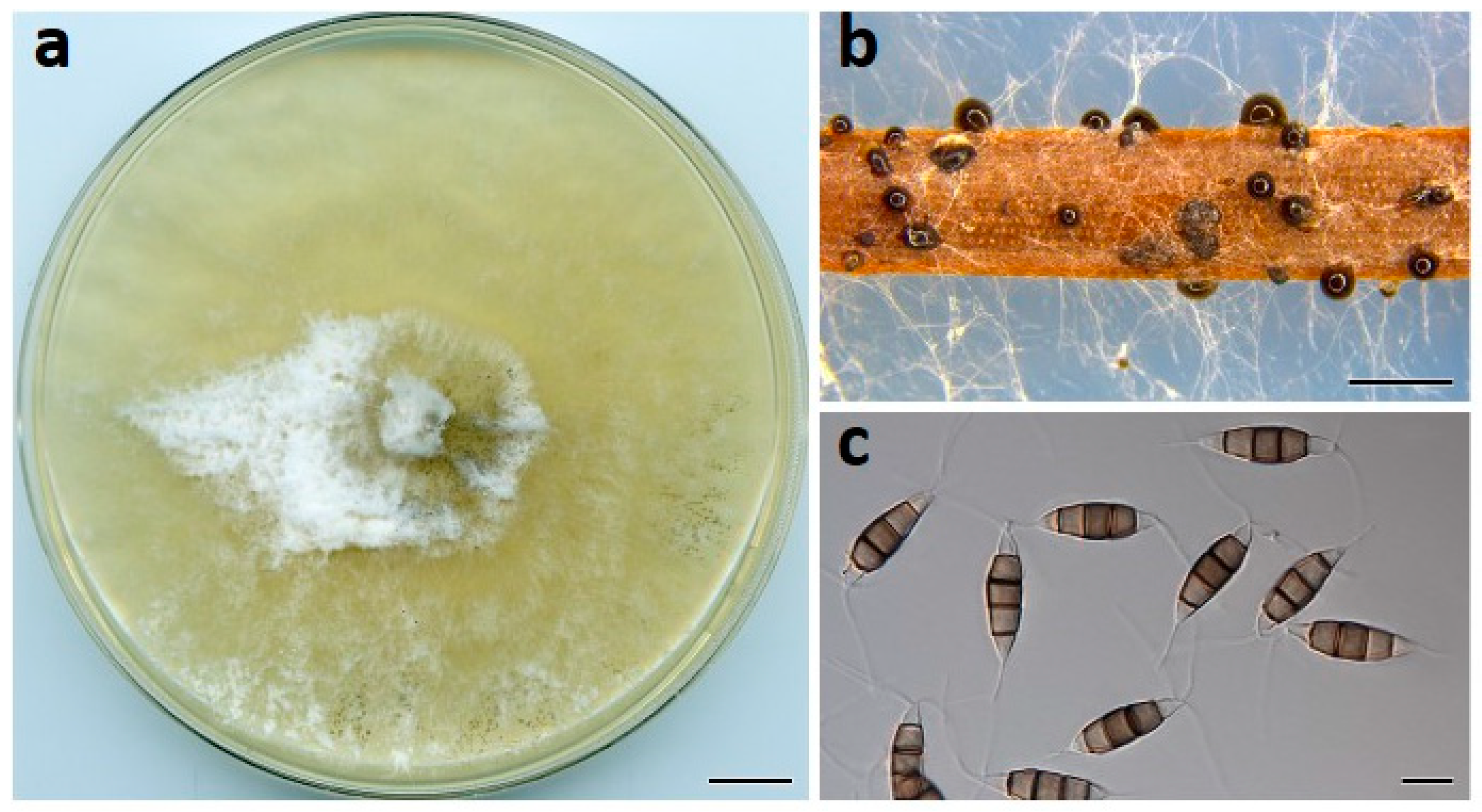
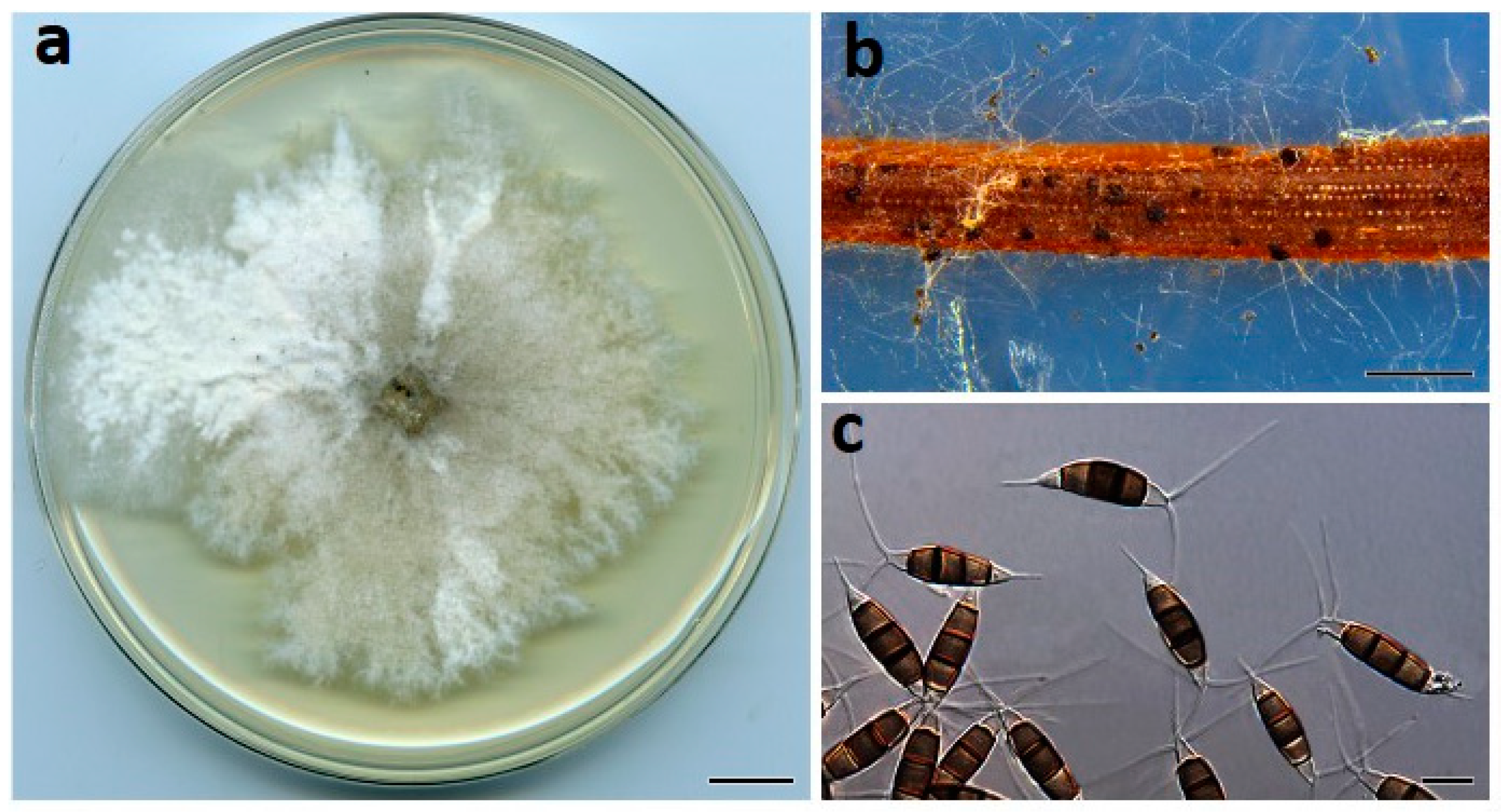
Figure 3.
Neopestalotiopsis drenthii (BRIP 72264a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.
Figure 3.
Neopestalotiopsis drenthii (BRIP 72264a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.

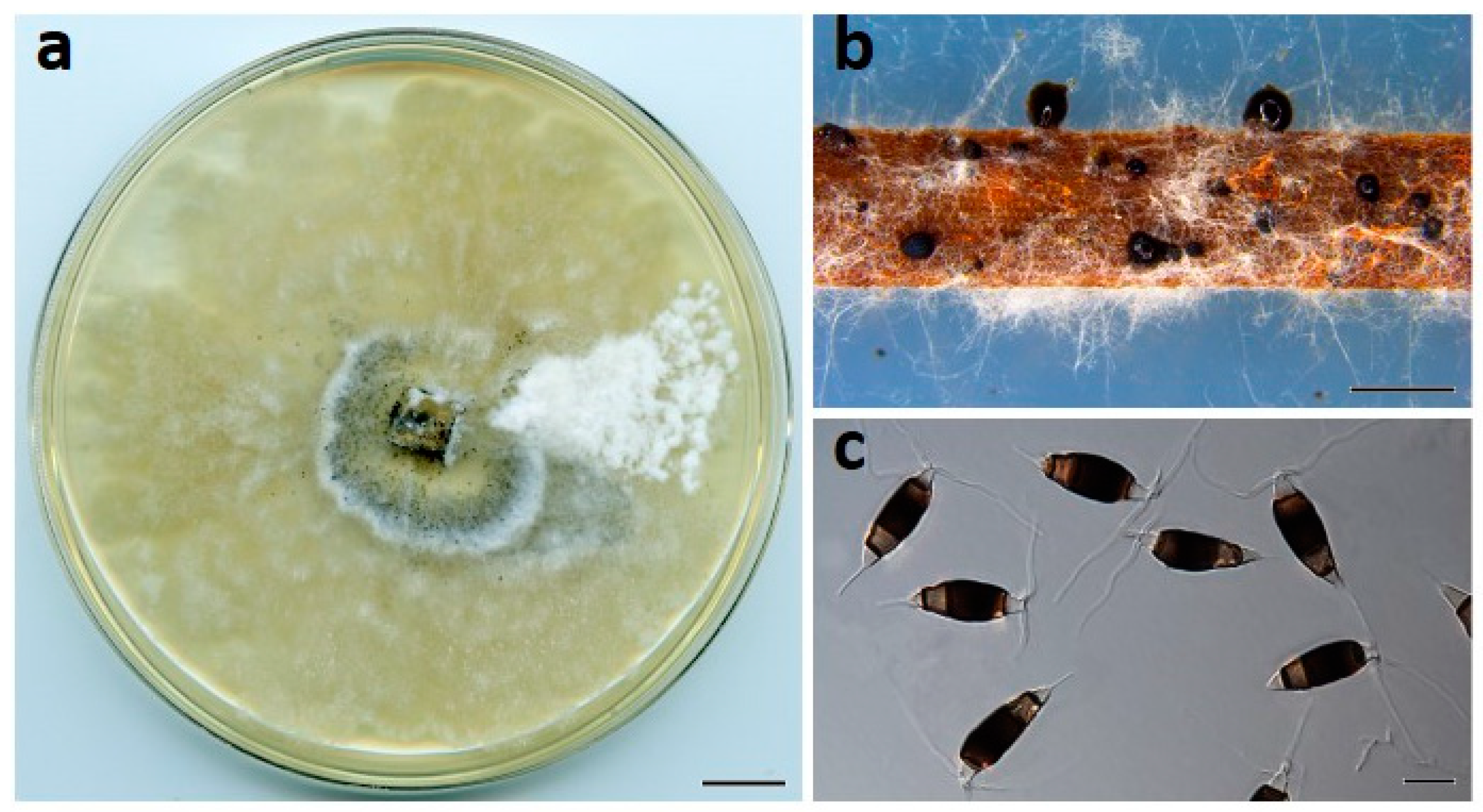
Figure 4.
Neopestalotiopsis maddoxii (BRIP 72266a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.
Figure 4.
Neopestalotiopsis maddoxii (BRIP 72266a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.

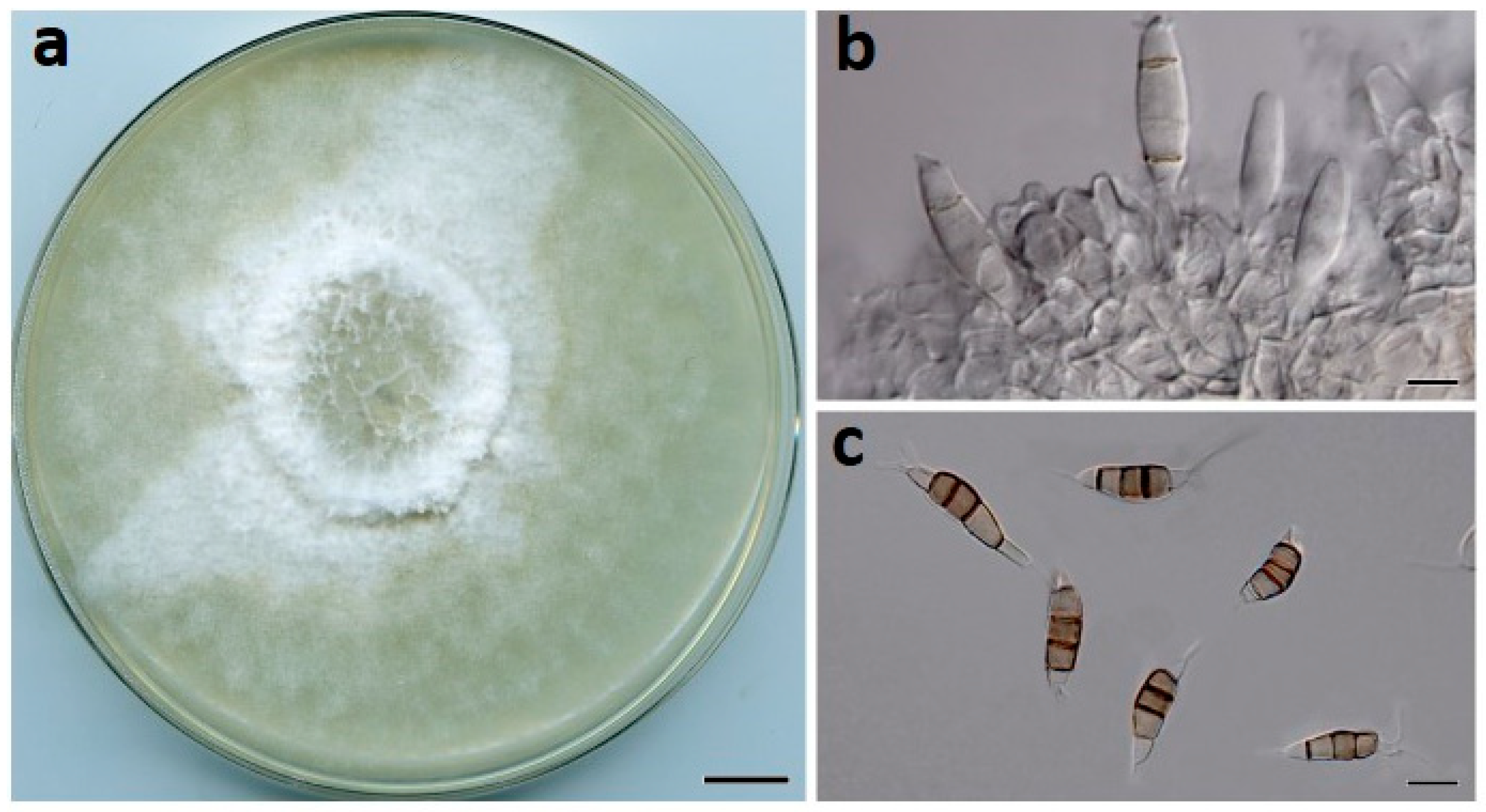
Figure 5.
Neopestalotiopsis olumideae (BRIP 72273a). (a) Two-week-old colony on PDA, (b) conidiogenous cells, and (c) conidia. Scale bars: a = 1 cm; b–c = 10 µm.
Figure 5.
Neopestalotiopsis olumideae (BRIP 72273a). (a) Two-week-old colony on PDA, (b) conidiogenous cells, and (c) conidia. Scale bars: a = 1 cm; b–c = 10 µm.

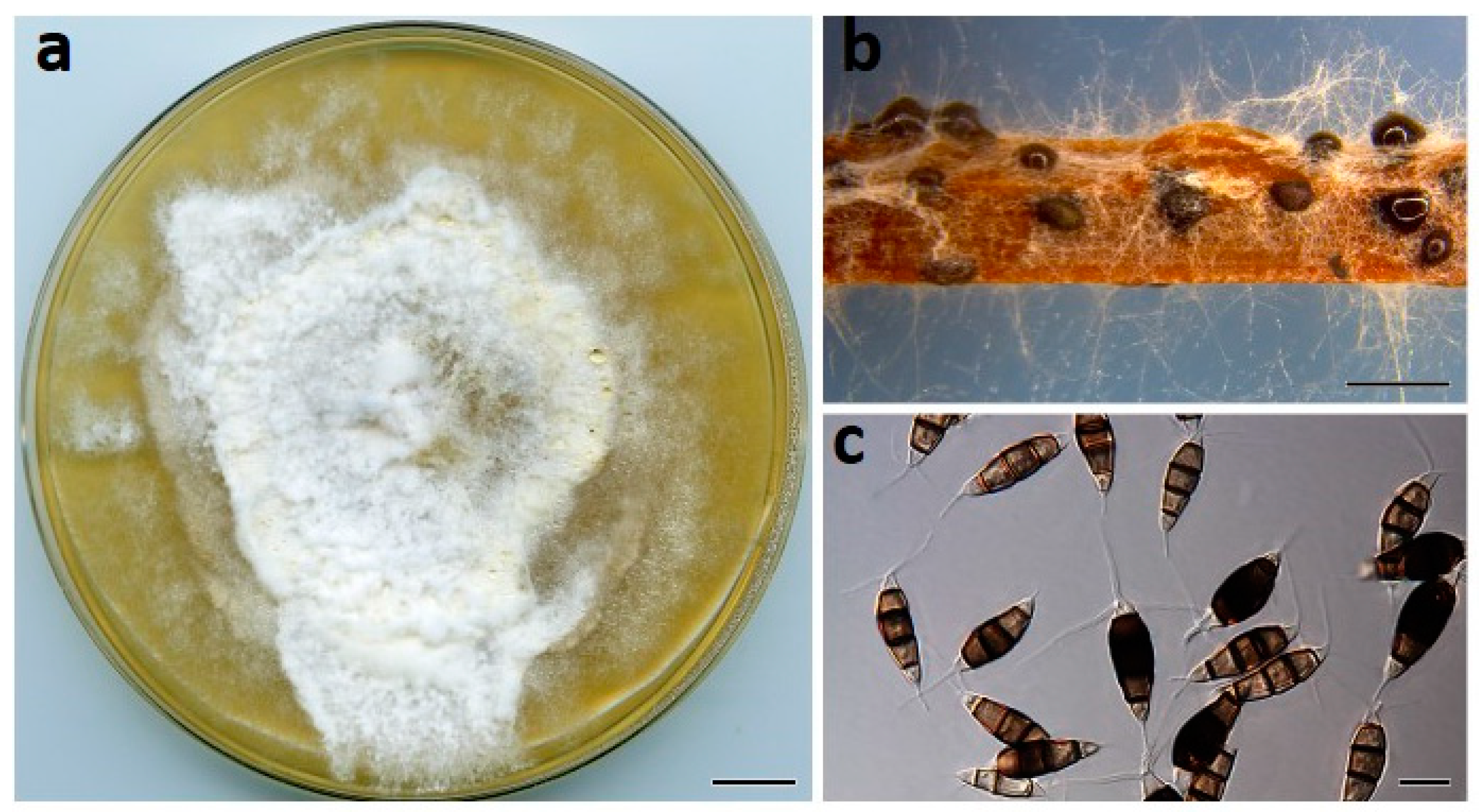
Figure 6.
Neopestalotiopsis vheenae (BRIP 72293a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.
Figure 6.
Neopestalotiopsis vheenae (BRIP 72293a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.

Figure 7.
Neopestalotiopsis zakeelii (BRIP 72282a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.
Figure 7.
Neopestalotiopsis zakeelii (BRIP 72282a). (a) Two-week-old colony on PDA, (b) conidiomata on pine needle agar, and (c) conidia. Scale bars: a = 1 cm; b = 1 mm; c = 10 µm.

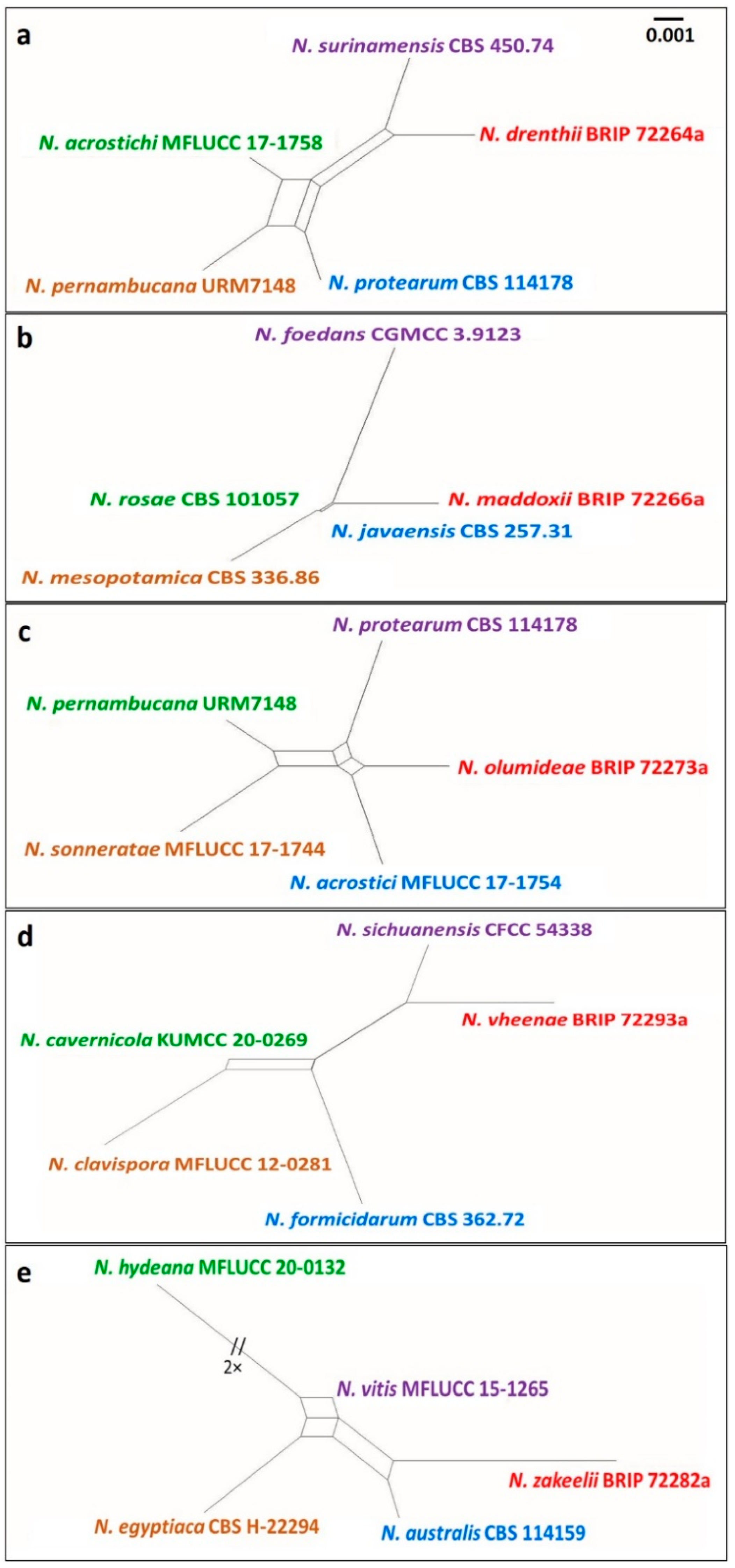
Figure 8.
Split graphs showing the result of PHI test of (a) Neopestalotiopsis drenthii (Fw = 0.735), (b) N. maddoxii (Fw = 0.118), (c) N. olumideae (Fw = 0.581), (d) N. vheenae (Fw = 0.876), and (e) N. zakeelii (Fw = 0.412) with their most closely related species using Log-Det transformation and splits decomposition options. The new taxon in each graph is shown in red font.
Figure 8.
Split graphs showing the result of PHI test of (a) Neopestalotiopsis drenthii (Fw = 0.735), (b) N. maddoxii (Fw = 0.118), (c) N. olumideae (Fw = 0.581), (d) N. vheenae (Fw = 0.876), and (e) N. zakeelii (Fw = 0.412) with their most closely related species using Log-Det transformation and splits decomposition options. The new taxon in each graph is shown in red font.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Neopestalotiopsis species and isolates used in the phylogenetic analyses, with GenBank accession numbers.
Table 1.
Neopestalotiopsis species and isolates used in the phylogenetic analyses, with GenBank accession numbers.
| Species | Strain 1 | Host/Substrate | Location | GenBank Accession Numbers 2 | Reference | ||
|---|---|---|---|---|---|---|---|
| ITS | TUB | TEF1α | |||||
| Neopestalotiopsis acrostichi | MFLUCC 17–1754 T | Acrostichum aureum | Thailand | MK764272 | MK764338 | MK764316 | [30] |
| N. alpapicalis | MFLUCC 17–2544 T | Rhizophora mucronata | Thailand | MK357772 | MK463545 | MK463547 | [28] |
| N. aotearoa | CBS 367.54 T | Canvas | New Zealand | KM199369 | KM199454 | KM199526 | [17] |
| N. asiatica | MFLUCC 12–0286 T | Prunus dulcis | China | JX398983 | JX399018 | JX399049 | [17] |
| N. australis | CBS 114159 T | Telopea sp. | Australia | KM199348 | KM199432 | KM199537 | [17] |
| N. brachiata | MFLUCC 17–555 T | Rhizophora apiculata | Thailand | MK764274 | MK764340 | MK764318 | [30] |
| N. brasiliensis | COAD 2166 T | Psidium guajava | Brazil | MG686469 | MG692400 | MG692402 | [42] |
| N. cavernicola | KUMCC 20–0269 T | Cave | China | MW545802 | MW557596 | MW550735 | [29] |
| N. chiangmaiensis | MFLUCC 18–0113 T | Pandanus sp. | Thailand | N/A | MH412725 | MH388404 | [43] |
| N. chrysea | MFLUCC 12–0261 T | Dead leaves | China | JX398985 | JX399020 | JX399051 | [19] |
| N. clavispora | MFLUCC 12–0281 T | Magnolia sp. | China | JX398979 | JX399014 | JX399045 | [19] |
| N. cocoes | MFLUCC 15–0152 T | Cocos nucifera | Thailand | KX789687 | N/A | KX789689 | [44] |
| N. cubana | CBS 600.96 T | Leaf Litter | Cuba | KM199347 | KM199438 | KM199521 | [17] |
| N. dendrobii | MFLUCC 14–0106 T | Dendrobium cariniferum | Thailand | MK993571 | MK975835 | MK975829 | [45] |
| N. drenthii | BRIP 72263a | Macadamia integrifolia | Australia | MZ303786 | MZ312679 | MZ344171 | This study |
| BRIP 72264a T | Macadamia integrifolia | Australia | MZ303787 | MZ312680 | MZ344172 | This study | |
| N. egyptiaca | CBS H–22294 T | Mangifera indica | Egypt | KP943747 | KP943746 | KP943748 | [46] |
| N. ellipsospora | MFLUCC 12–0283 T | Dead plant material | China | JX398980 | JX399016 | JX399047 | [19] |
| N. eucalypticola | CBS 264.37 T | Eucalyptus globulus | N/A | KM199376 | KM199431 | KM199551 | [17] |
| N. foedans | CGMCC 3.9123 T | Mangrove plant | China | JX398987 | JX399022 | JX399053 | [19] |
| N. formicidarum | CBS 362.72 T | Dead Formicidae (ant) | Cuba | KM199358 | KM199455 | KM199517 | [17] |
| N. hadrolaeliae | EHJ6a | Cattleya jongheana | Brazil | MK454709 | MK465120 | MK465122 | [27] |
| N. honoluluana | CBS 114495 T | Telopea sp. | USA | KM199364 | KM199457 | KM199548 | [17] |
| N. hydeana | MFLUCC 20–0132 T | Artocarpus heterophyllus | Thailand | MW266069 | MW251119 | MW251129 | [32] |
| N. iranensis | CBS 137768 T | Fragaria ananassa | Iran | KM074048 | KM074057 | KM074051 | [20] |
| N. javaensis | CBS 257.31 T | Cocos nucifera | Java | KM199357 | KM199437 | KM199543 | [17] |
| N. macadamiae | BRIP 63737c T | Macadamia sp. | Australia | KX186604 | KX186654 | KX186627 | [8] |
| N. maddoxii | BRIP 72260a | Macadamia integrifolia | Australia | MZ303780 | MZ312673 | MZ344165 | This study |
| BRIP 72262a | Macadamia integrifolia | Australia | MZ303781 | MZ312674 | MZ344166 | This study | |
| BRIP 72266a T | Macadamia integrifolia | Australia | MZ303782 | MZ312675 | MZ344167 | This study | |
| BRIP 72272a | Macadamia integrifolia | Australia | MZ303783 | MZ312676 | MZ344168 | This study | |
| BRIP 72275a | Macadamia integrifolia | Australia | MZ303784 | MZ312677 | MZ344169 | This study | |
| BRIP 72284a | Macadamia integrifolia | Australia | MZ303785 | MZ312678 | MZ344170 | This study | |
| N. magna | MFLUCC 12–652 T | Pteridium sp. | France | KF582795 | KF582793 | KF582791 | [17] |
| N. mesopotamica | CBS 336.86 T | Pinus brutia | Iraq | KM199362 | KM199441 | KM199555 | [17] |
| N. musae | MFLUCC 15–0776 T | Musa sp. | Thailand | KX789683 | KX789686 | KX789685 | [44] |
| N. natalensis | CBS 138.41 T | Acacia mollissima | South Africa | KM199377 | KM199466 | KM199552 | [17] |
| N. nebuloides | BRIP 66617 T | Sporobolus elongatus | Australia | MK966338 | MK977632 | MK977633 | [31] |
| N. olumideae | BRIP 72273a T | Macadamia integrifolia | Australia | MZ303790 | MZ312683 | MZ344175 | This study |
| BRIP 72283a | Macadamia integrifolia | Australia | MZ303791 | MZ312684 | MZ344176 | This study | |
| N. pandanicola | KUMCC 17–0175 | Pandanus sp. | China | N/A | MH412720 | MH388389 | [43] |
| N. pernambucana | URM7148 | Vismia guianensis | Brazil | KJ792466 | N/A | KU306739 | [47] |
| N. petila | MFLUCC 17–1737 T | Rhizophora mucronata | Thailand | MK764275 | MK764341 | MK764319 | [30] |
| N. phangngaensis | MFLUCC 18–0119 T | Pandanus sp. | Thailand | MH388354 | MH412721 | MH388390 | [43] |
| N. piceana | CBS 394.48 T | Picea sp. | UK | KM199368 | KM199453 | KM199527 | [17] |
| N. protearum | CBS 114178 T | Leucospermum cuneiforme | Zimbabwe | JN712498 | KM199463 | LT853201 | [17] |
| N. rhizophorae | MFLUCC 17–1551 T | Rhizophora mucronata | Thailand | MK764278 | MK764344 | MK764322 | [30] |
| N. rosae | CBS 101057 T | Rosa sp. | New Zealand | KM199359 | KM199429 | KM199523 | [17] |
| N. rosicola | CFCC 51992 T | Rosa chinensis | China | KY885239 | KY885245 | KY885243 | [48] |
| N. samarangensis | CBS 115451 | Unidentified tree | China | KM199365 | KM199447 | KM199556 | [49] |
| N. saprophytica | MFLUCC 12–0282 T | Magnolia sp. | China | JX398982 | JX399017 | JX399048 | [19] |
| N. sichuanensis | CFCC 54338 T | Castanea mollissima | China | MW166231 | MW218524 | MW199750 | [33] |
| N. sonneratae | MFLUCC 17–1744 T | Sonneronata alba | Thailand | MK764279 | MK764345 | MK764323 | [30] |
| N. steyaertii | IMI 192475 T | Eucalyptus viminalis | Australia | KF582796 | KF582794 | KF582792 | [17] |
| N. surinamensis | CBS 450.74 T | Soil under | Suriname | KM199351 | KM199465 | KM199518 | [17] |
| Elaeis guineensis | |||||||
| N. thailandica | MFLUCC 17–1730 T | Rhizophora mucronata | Thailand | MK764281 | MK764347 | MK764325 | [30] |
| N. umbrinospora | MFLUCC 12–0285 T | Unidentified plant | China | JX398984 | JX399019 | JX399050 | [19] |
| N. vheenae | BRIP 72293a T | Macadamia integrifolia | Australia | MZ303792 | MZ312685 | MZ344177 | This study |
| BRIP 70210 | Macadamia integrifolia | Australia | MN114212 | MN114214 | MN114213 | [5] 3 | |
| N. vitis | MFLUCC 15–1265 T | Vitis vinifera | China | KU140694 | KU140685 | KU140676 | [22] |
| N. zakeelii | BRIP 72271a | Macadamia integrifolia | Australia | MZ303788 | MZ312681 | MZ344173 | This study |
| BRIP 72282a T | Macadamia integrifolia | Australia | MZ303789 | MZ312682 | MZ344174 | This study | |
| N. zimbabwana | CBS 111495 T | Leucospermum | Zimbabwe | JX556231 | KM199456 | KM199545 | [17] |
| cunciforme | |||||||
| Pestalotiopsis diversiseta | MFLUCC 12–0287 T | Dead plant material | China | JX399009 | JX399040 | JX399073 | [19] |
1 BRIP: Queensland Plant Pathology Herbarium, Australia; CBS: Culture collection of the Centraalbureau voor Schimmelcultures, Fungal Biodiversity Centre, Utrecht, The Netherlands; CFCC: China Forestry Culture Collection Center, Research Institute of Forest Ecology, Environment and Protection, Beijing, China; CGMCC: China General Microbiological Culture Collection Center, Institute of Microbiology, Chinese Academy of Sciences, Beijing, China; COAD: Culture collection Coleção Octávio Almeida Drummond of the Universidade Federal de Viçosa, Viçosa, Brazil; HGUP: Plant Pathology Herbarium of Guizhou University, Guizhou, China; IMI: Culture collection of CABI Europe UK Centre, Egham, UK; KUMCC: Culture Collection of Kunming Institute of Botany, Chinese Academy of Sciences, Kunming, China; MFLUCC: Mae Fah Luang University Culture Collection, Chiang Rai, Thailand; URM: The Father Camille Torrend Herbarium, Pernambuco, Brazil. 2 ITS: internal transcribed spacer; TUB: β-tubulin; TEF1α: translation elongation factor 1-α. 3 as Neopestalotiopsis clavispora. Ex-type strains are labeled with T. N/A: Not available.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Prasannath, K.; Shivas, R.G.; Galea, V.J.; Akinsanmi, O.A. Neopestalotiopsis Species Associated with Flower Diseases of Macadamia integrifolia in Australia. J. Fungi 2021, 7, 771. https://doi.org/10.3390/jof7090771
AMA Style
Prasannath K, Shivas RG, Galea VJ, Akinsanmi OA. Neopestalotiopsis Species Associated with Flower Diseases of Macadamia integrifolia in Australia. Journal of Fungi. 2021; 7(9):771. https://doi.org/10.3390/jof7090771
Chicago/Turabian StylePrasannath, Kandeeparoopan, Roger G. Shivas, Victor J. Galea, and Olufemi A. Akinsanmi. 2021. "Neopestalotiopsis Species Associated with Flower Diseases of Macadamia integrifolia in Australia" Journal of Fungi 7, no. 9: 771. https://doi.org/10.3390/jof7090771
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.