Preeclampsia with Intrauterine Growth Restriction Generates Morphological Changes in Endothelial Cells Associated with Mitochondrial Swelling—An In Vitro Study

 ,
,  ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Ethics Statement
2.2. Study Design
2.3. Biochemical Analyses
2.4. Cell Culture
2.5. Experimental Design
2.6. Endothelial Cell Viability
2.7. Cell Proliferation
2.8. Detection of Reactive Oxygen Species (ROS)
2.9. Light Microscopy
2.10. Electron Microscopy
2.11. Statistical Analysis
3. Results
3.1. Patient Description
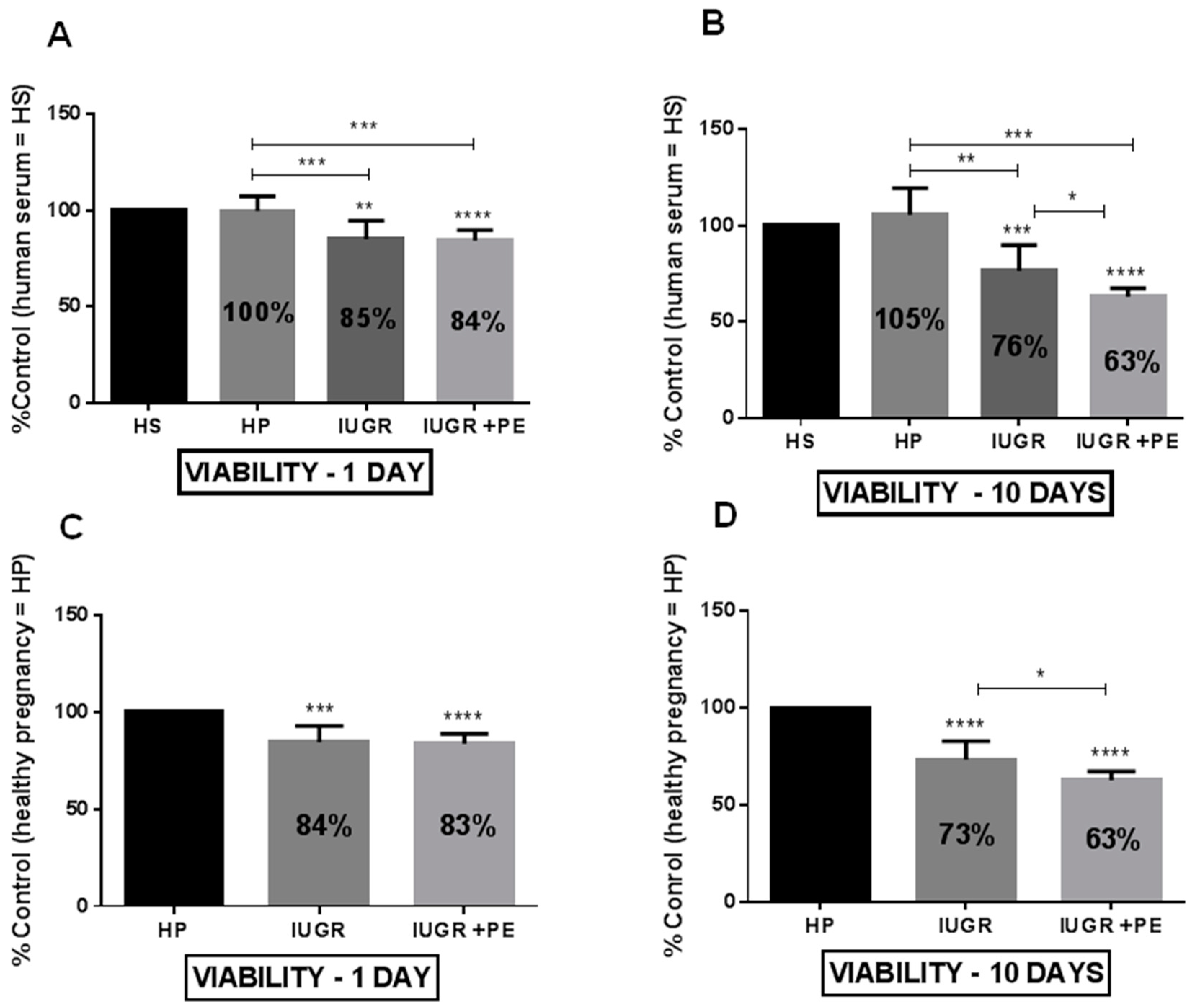
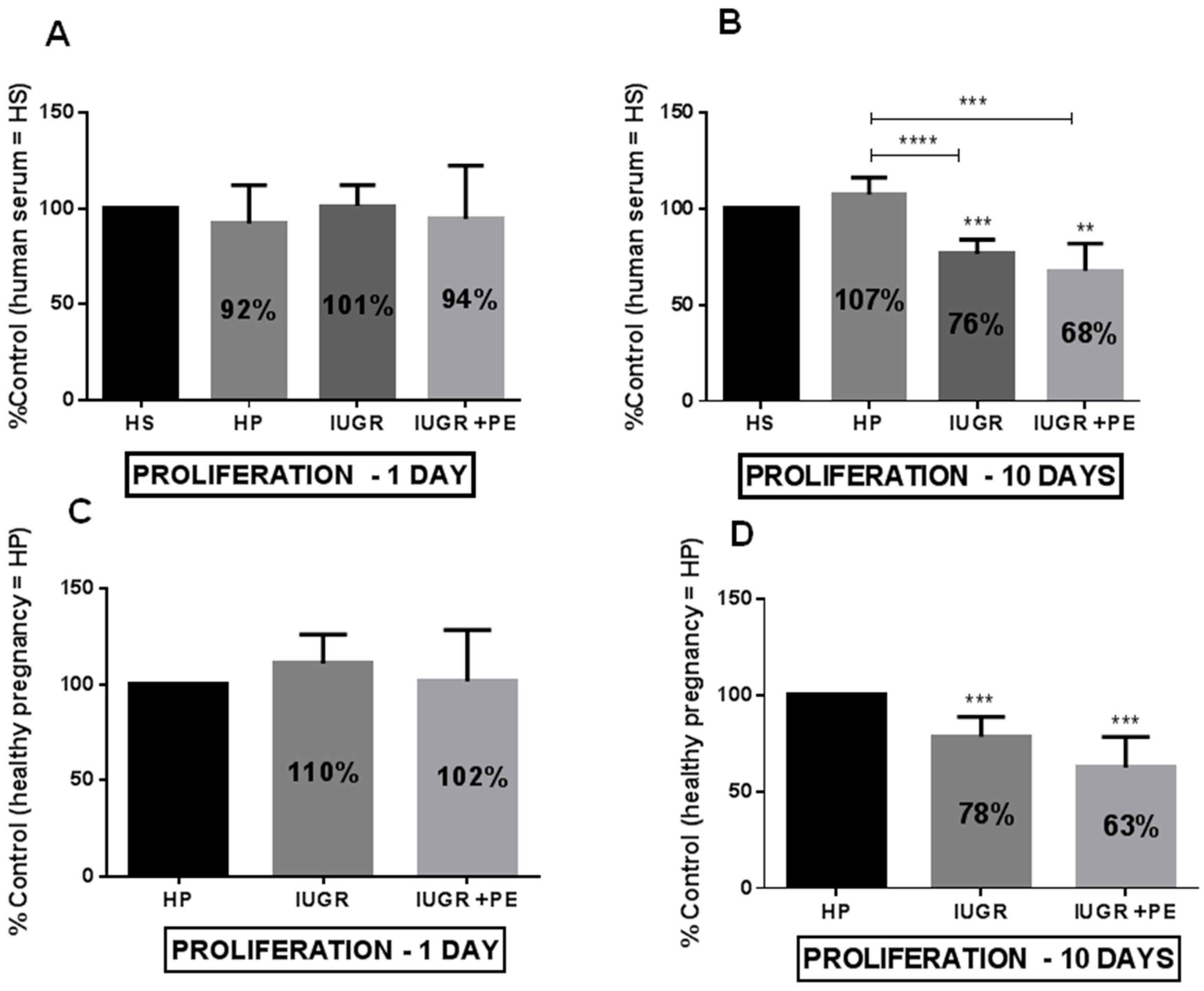
3.2. Endothelial Cell Viability and Proliferation
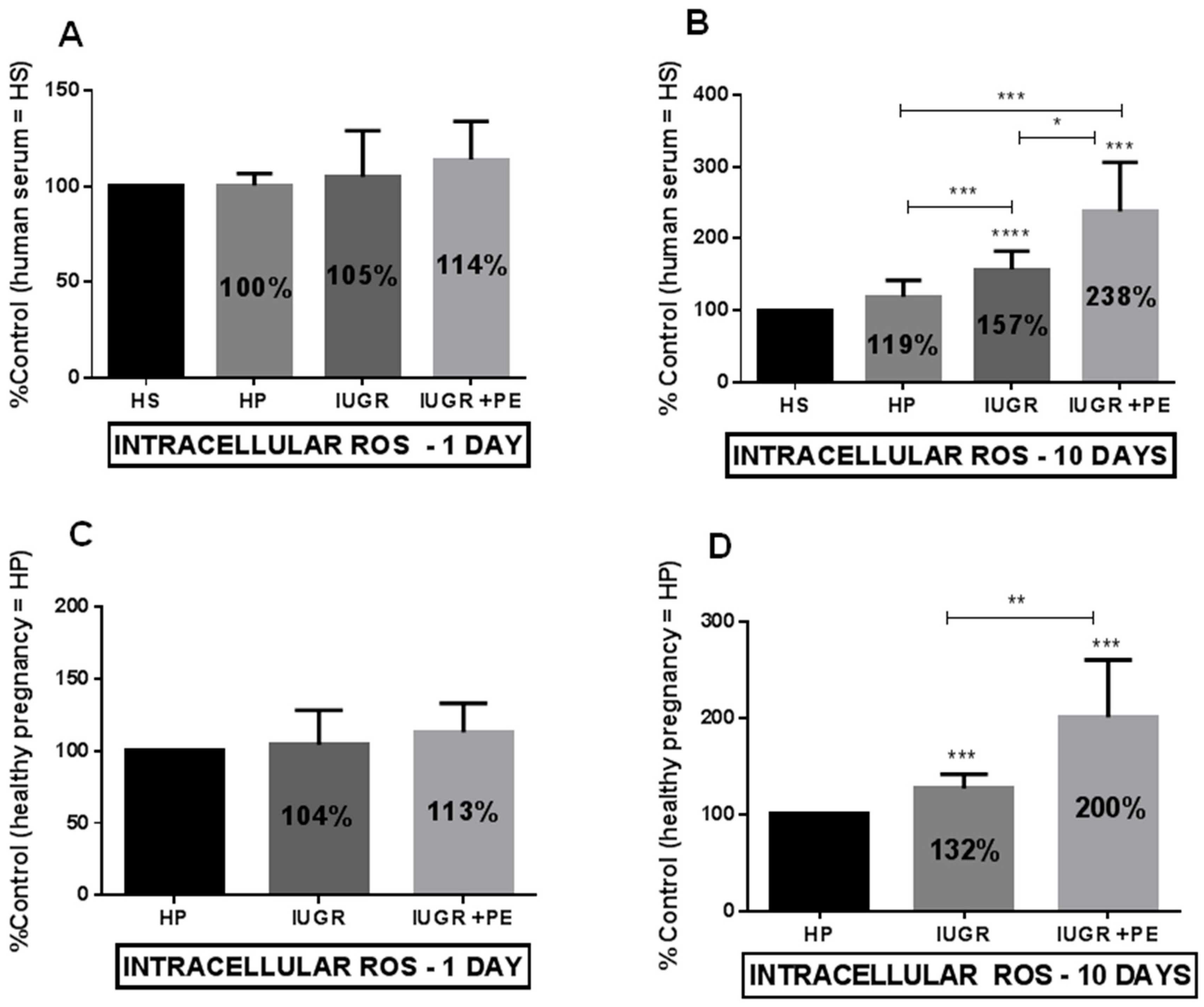
3.3. Reactive Oxygen Species (ROS) Generation
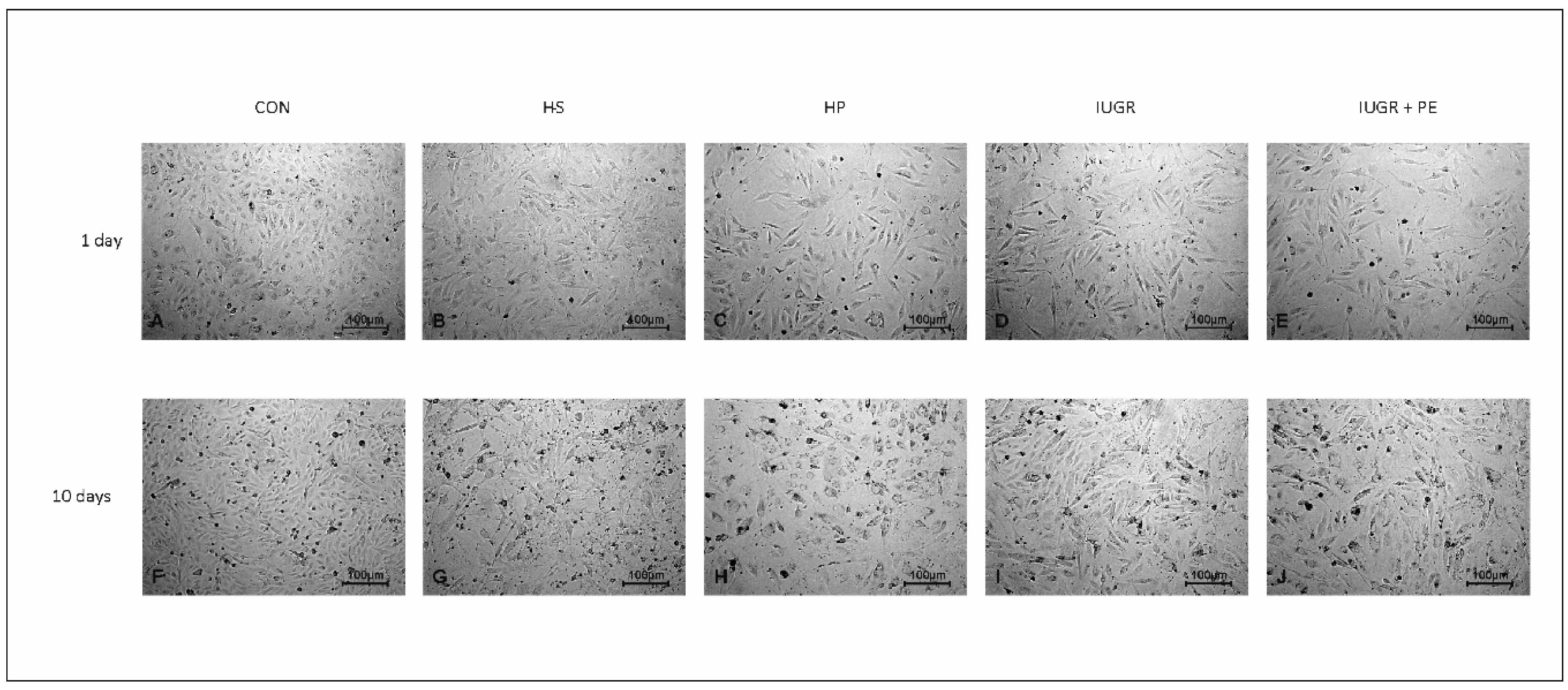
3.4. Light Microscopy
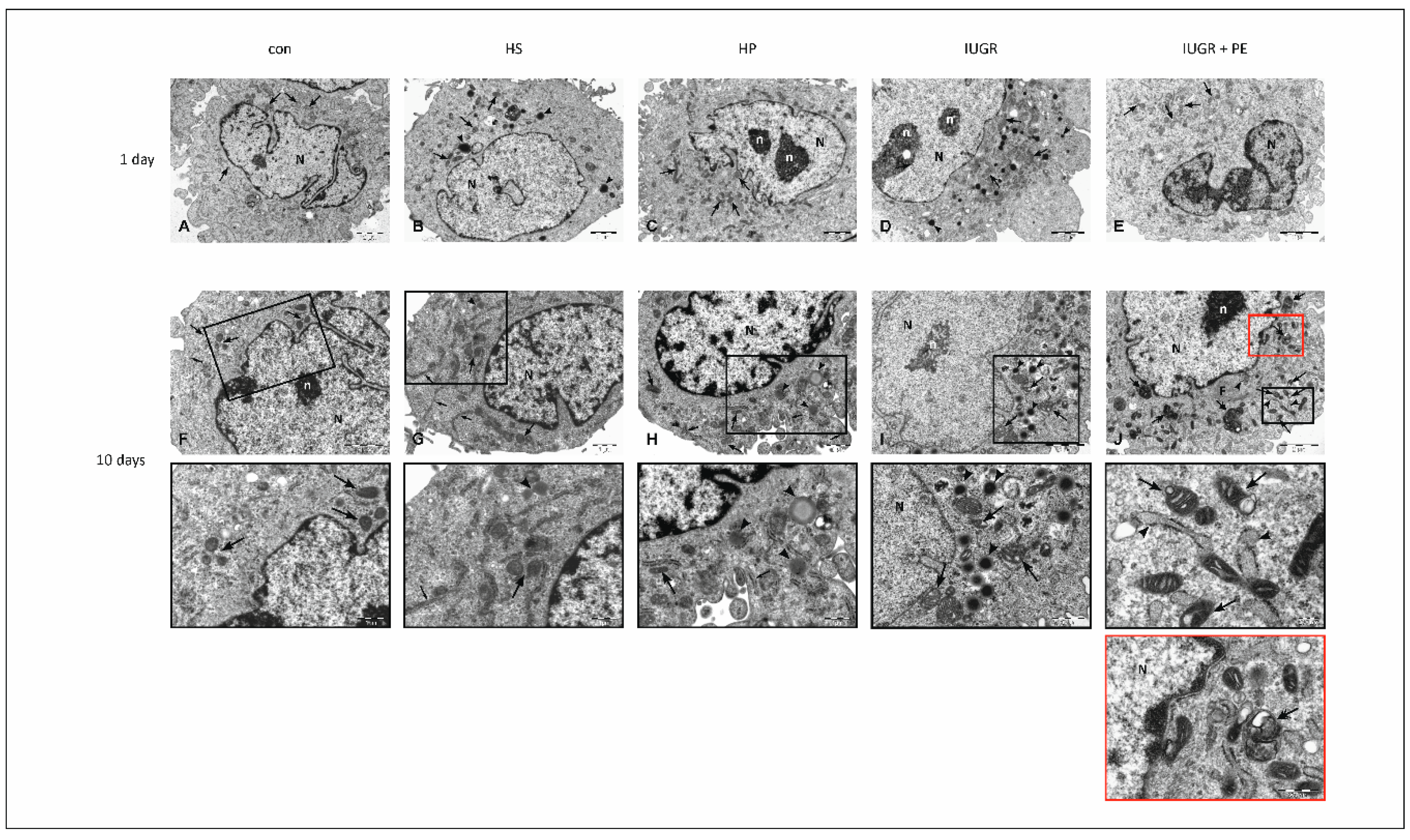
3.5. Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef]
- Roland, C.S.; Hu, J.; Ren, C.E.; Chen, H.; Li, J.; Varvoutis, M.S.; Leaphart, L.W.; Byck, D.B.; Zhu, X.; Jiang, S.W. Morphological changes of placental syncytium and their implications for the pathogenesis of preeclampsia. Cell. Mol. Life Sci. 2016, 73, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Bonello, S.; Zahringer, C.; Belaiba, R.S.; Djordjevic, T.; Hess, J.; Michiels, C.; Kietzmann, T.; Gorlach, A. Reactive oxygen species activate the HIF-1alpha promoter via a functional NFkappaB site. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Pathophysiology of placental-derived fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S745–S761. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.E.; Min, J.Y.; Merchan, J.; Lim, K.H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef]
- Jonsdottir, L.S.; Arngrimsson, R.; Geirsson, R.T.; Sigvaldason, H.; Sigfusson, N. Death rates from ischemic heart disease in women with a history of hypertension in pregnancy. Acta Obstet. Gynecol. Scand. 1995, 74, 772–776. [Google Scholar] [CrossRef] [PubMed]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.H.; Yuan, H.T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Yinon, Y.; Kingdom, J.C.; Odutayo, A.; Moineddin, R.; Drewlo, S.; Lai, V.; Cherney, D.Z.; Hladunewich, M.A. Vascular dysfunction in women with a history of preeclampsia and intrauterine growth restriction: Insights into future vascular risk. Circulation 2010, 122, 1846–1853. [Google Scholar] [CrossRef]
- Crispi, F.; Dominguez, C.; Llurba, E.; Martin-Gallan, P.; Cabero, L.; Gratacos, E. Placental angiogenic growth factors and uterine artery Doppler findings for characterization of different subsets in preeclampsia and in isolated intrauterine growth restriction. Am. J. Obstet. Gynecol. 2006, 195, 201–207. [Google Scholar] [CrossRef]
- Roberts, J.M.; Edep, M.E.; Goldfien, A.; Taylor, R.N. Sera from preeclamptic women specifically activate human umbilical vein endothelial cells in vitro: Morphological and biochemical evidence. Am. J. Reprod. Immunol. 1992, 27, 101–108. [Google Scholar] [CrossRef]
- Matsubara, K.; Matsubara, Y.; Hyodo, S.; Katayama, T.; Ito, M. Role of nitric oxide and reactive oxygen species in the pathogenesis of preeclampsia. J. Obstet. Gynaecol. Res. 2010, 36, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gu, Y.; Zhang, Y.; Lewis, D.F. Evidence of endothelial dysfunction in preeclampsia: Decreased endothelial nitric oxide synthase expression is associated with increased cell permeability in endothelial cells from preeclampsia. Am. J. Obstet. Gynecol. 2004, 190, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Edgell, C.J.; McDonald, C.C.; Graham, J.B. Permanent cell line expressing human factor VIII-related antigen established by hybridization. Proc. Natl. Acad. Sci. USA 1983, 80, 3734–3737. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Witowski, J.; Korybalska, K.; Wisniewska, J.; Breborowicz, A.; Gahl, G.M.; Frei, U.; Passlick-Deetjen, J.; Jorres, A. Effect of glucose degradation products on human peritoneal mesothelial cell function. J. Am. Soc. Nephrol. 2000, 11, 729–739. [Google Scholar]
- Krysko, D.V.; Vanden, B.T.; D’Herde, K.; Vandenabeele, P. Apoptosis and necrosis: Detection, discrimination and phagocytosis. Methods 2008, 44, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Vanlangenakker, N.; Vanden, B.T.; Krysko, D.V.; Festjens, N.; Vandenabeele, P. Molecular mechanisms and pathophysiology of necrotic cell death. Curr. Mol. Med. 2008, 8, 207–220. [Google Scholar] [CrossRef]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef]
- Pedersen, P.L. ATP synthase. The machine that makes ATP. Curr. Biol. 1994, 4, 1138–1141. [Google Scholar] [CrossRef]
- Antico Arciuch, V.G.; Elguero, M.E.; Poderoso, J.J.; Carreras, M.C. Mitochondrial regulation of cell cycle and proliferation. Antioxid. Redox Signal. 2012, 16, 1150–1180. [Google Scholar] [CrossRef] [PubMed]
- Sweet, S.; Singh, G. Changes in mitochondrial mass, membrane potential, and cellular adenosine triphosphate content during the cell cycle of human leukemic (HL-60) cells. J. Cell. Physiol. 1999, 180, 91–96. [Google Scholar] [CrossRef]
- Genc, H.; Uzun, H.; Benian, A.; Simsek, G.; Gelisgen, R.; Madazli, R.; Guralp, O. Evaluation of oxidative stress markers in first trimester for assessment of preeclampsia risk. Arch. Gynecol. Obstet. 2011, 284, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Manton, K.G. The role of oxidative damage in mitochondria during aging: A review. Front. Biosci. 2004, 9, 1100–1117. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R.; Nyska, A. Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L.; Webster, R.P. Vascular biology of preeclampsia. J. Thromb. Haemost. 2009, 7, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Guzy, R.D.; Schumacker, P.T. Oxygen sensing by mitochondria at complex III: The paradox of increased reactive oxygen species during hypoxia. Exp. Physiol. 2006, 91, 807–819. [Google Scholar] [CrossRef]
- Sanchez-Aranguren, L.C.; Prada, C.E.; Riano-Medina, C.E.; Lopez, M. Endothelial dysfunction and preeclampsia: Role of oxidative stress. Front. Physiol. 2014, 5, 372. [Google Scholar] [CrossRef]
- Holland, O.J.; Cuffe, J.S.M.; Dekker, N.M.; Callaway, L.; Kwan Cheung, K.A.; Radenkovic, F.; Perkins, A.V. Placental mitochondrial adaptations in preeclampsia associated with progression to term delivery. Cell Death Dis. 2018, 9, 1150. [Google Scholar] [CrossRef]
- Nakamura, M.; Sekizawa, A.; Purwosunu, Y.; Okazaki, S.; Farina, A.; Wibowo, N.; Shimizu, H.; Okai, T. Cellular mRNA expressions of anti-oxidant factors in the blood of preeclamptic women. Prenat. Diagn. 2009, 29, 691–696. [Google Scholar] [CrossRef]
- Hayashi, T.; Rizzuto, R.; Hajnoczky, G.; Su, T.P. MAM: More than just a housekeeper. Trends Cell Biol. 2009, 19, 81–88. [Google Scholar] [CrossRef]
- Huang, C.Y.; Chiang, S.F.; Lin, T.Y.; Chiou, S.H.; Chow, K.C. HIV-1 Vpr triggers mitochondrial destruction by impairing Mfn2-mediated ER-mitochondria interaction. PLoS ONE 2012, 7, e33657. [Google Scholar] [CrossRef] [PubMed]
- Klecker, T.; Westermann, B. Mitochondria are clamped to vacuoles for lipid transport. Dev. Cell 2014, 30, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Dulak, J.; Loboda, A.; Jozkowicz, A. Effect of heme oxygenase-1 on vascular function and disease. Curr. Opin. Lipidol. 2008, 19, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Turpin, C.A.; Sakyi, S.A.; Owiredu, W.K.; Ephraim, R.K.; Anto, E.O. Association between adverse pregnancy outcome and imbalance in angiogenic regulators and oxidative stress biomarkers in gestational hypertension and preeclampsia. BMC Pregnancy Childbirth 2015, 15, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerencser, A.A.; Doczi, J.; Torocsik, B.; Bossy-Wetzel, E.; dam-Vizi, V. Mitochondrial swelling measurement in situ by optimized spatial filtering: Astrocyte-neuron differences. Biophys. J. 2008, 95, 2583–2598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Y.D.; Wang, H.; Kho, S.H.; Rinkiko, S.; Sheng, X.; Shen, H.M.; Zhu, Y.Z. Hydrogen sulfide protects HUVECs against hydrogen peroxide induced mitochondrial dysfunction and oxidative stress. PLoS ONE 2013, 8, e53147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, A.D.; Siniossoglou, S. Function of lipid droplet-organelle interactions in lipid homeostasis. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1459–1468. [Google Scholar] [CrossRef]
- Hubel, C.A.; Roberts, J.M.; Taylor, R.N.; Musci, T.J.; Rogers, G.M.; McLaughlin, M.K. Lipid peroxidation in pregnancy: New perspectives on preeclampsia. Am. J. Obstet. Gynecol. 1989, 161, 1025–1034. [Google Scholar] [CrossRef]
- Lorentzen, B.; Endresen, M.J.; Hovig, T.; Haug, E.; Henriksen, T. Sera from preeclamptic women increase the content of triglycerides and reduce the release of prostacyclin in cultured endothelial cells. Thromb. Res. 1991, 63, 363–372. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, J.; Yu, X.; Shi, L.; Du, W.; Hu, L.; Liu, C.; Cao, Y. SIRT1 regulates accumulation of oxidized LDL in HUVEC via the autophagy-lysosomal pathway. Prostaglandins Other Lipid Mediat. 2016, 122, 37–44. [Google Scholar] [CrossRef]
- Dong, G.; Yang, S.; Cao, X.; Yu, N.; Yu, J.; Qu, X. Low shear stressinduced autophagy alleviates cell apoptosis in HUVECs. Mol. Med. Rep. 2017, 15, 3076–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Sun, X.; Jiang, L.; Liu, X.; Chen, M.; Yao, X.; Sun, Q.; Yang, G. 6-Gingerol induces autophagy to protect HUVECs survival from apoptosis. Chem. Biol. Interact. 2016, 256, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Normal Pregnancy Control (n = 25) | IUGR (n = 25) | IUGR + PE (n = 24) |
|---|---|---|---|
| Age, years | 32 ± 6 | 29 ± 6 | 30 ± 6 |
| Number of pregnancy, median (min-max) | 2 (1–8) | 1 (1–8) | 1 (1–5) |
| Primiparous, n (%) | 7 (28%) | 14 (56%) | 13 (34%) |
| Gestational age, week, median (min-max) | 39 (38–41) | 36 (28–40) | 34 (28–40) |
| Newborn weight, g | 3531 ± 476 | 2032 ± 720 **** | 1734 ± 684 **** |
| SYS, mmHg | 109.6 ± 6.6 | 112.9 ± 21.7 †††† | 132.9 ± 18.6 **** |
| DIA, mmHg | 69.2 ± 5.53 | 68.6 ± 9.7 †††† | 82.7 ± 12.86 ** |
| Urine protein, g/L / 24 h | below 0.03 | 0,15 ± 0 †††† | 3.07 ± 4.31 **** |
| hsCRP, mg/L | 5.3 ± 3.4 | 6.2 ± 9.0 | 5.4 ± 4.7 |
| VEGF, pg/mL | 139.0 ± 37.5 | 36.3 ± 20.2 **** | 40.9 ± 2.2 **** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Formanowicz, D.; Malińska, A.; Nowicki, M.; Kowalska, K.; Gruca-Stryjak, K.; Bręborowicz, G.; Korybalska, K. Preeclampsia with Intrauterine Growth Restriction Generates Morphological Changes in Endothelial Cells Associated with Mitochondrial Swelling—An In Vitro Study. J. Clin. Med. 2019, 8, 1994. https://doi.org/10.3390/jcm8111994
Formanowicz D, Malińska A, Nowicki M, Kowalska K, Gruca-Stryjak K, Bręborowicz G, Korybalska K. Preeclampsia with Intrauterine Growth Restriction Generates Morphological Changes in Endothelial Cells Associated with Mitochondrial Swelling—An In Vitro Study. Journal of Clinical Medicine. 2019; 8(11):1994. https://doi.org/10.3390/jcm8111994
Chicago/Turabian StyleFormanowicz, Dorota, Agnieszka Malińska, Marcin Nowicki, Katarzyna Kowalska, Karolina Gruca-Stryjak, Grzegorz Bręborowicz, and Katarzyna Korybalska. 2019. "Preeclampsia with Intrauterine Growth Restriction Generates Morphological Changes in Endothelial Cells Associated with Mitochondrial Swelling—An In Vitro Study" Journal of Clinical Medicine 8, no. 11: 1994. https://doi.org/10.3390/jcm8111994