
Rational Mutagenesis in the Lid Domain of Ribokinase from E. coli Results in an Order of Magnitude Increase in Activity towards D-arabinose
 , ,
, ,  , ,
, ,  , ,
, ,
Abstract
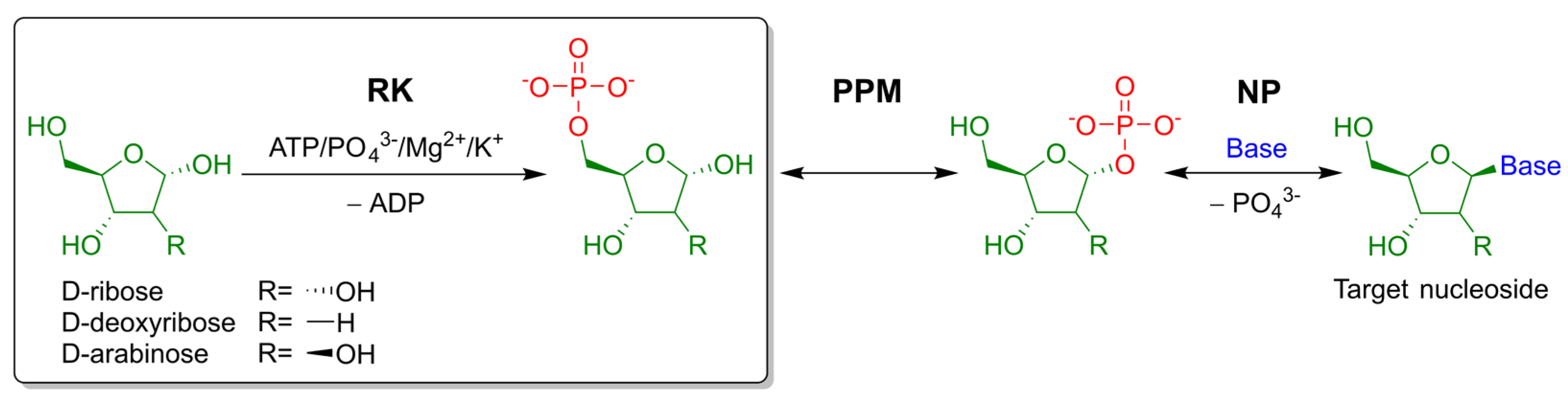
:1. Introduction
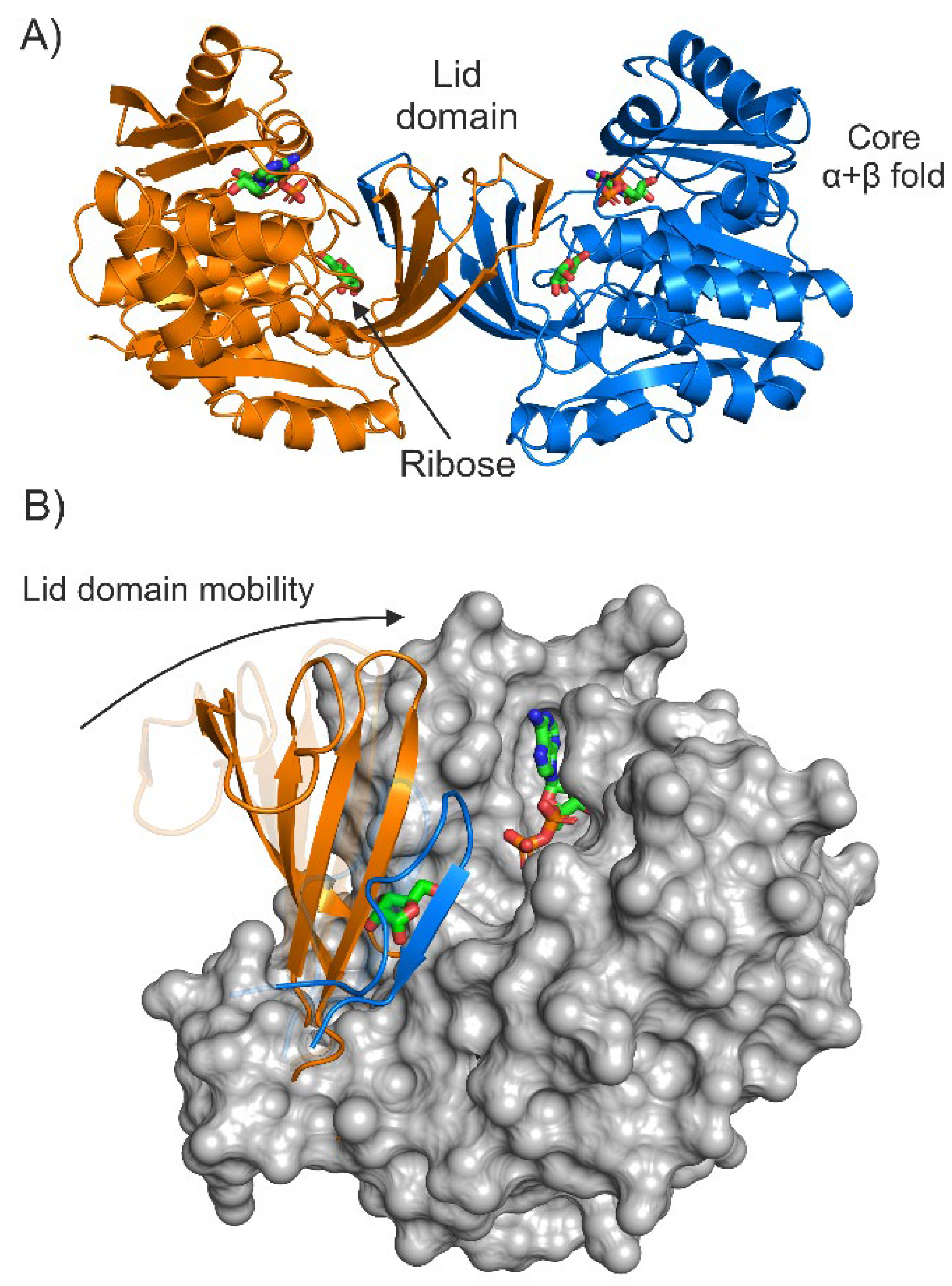
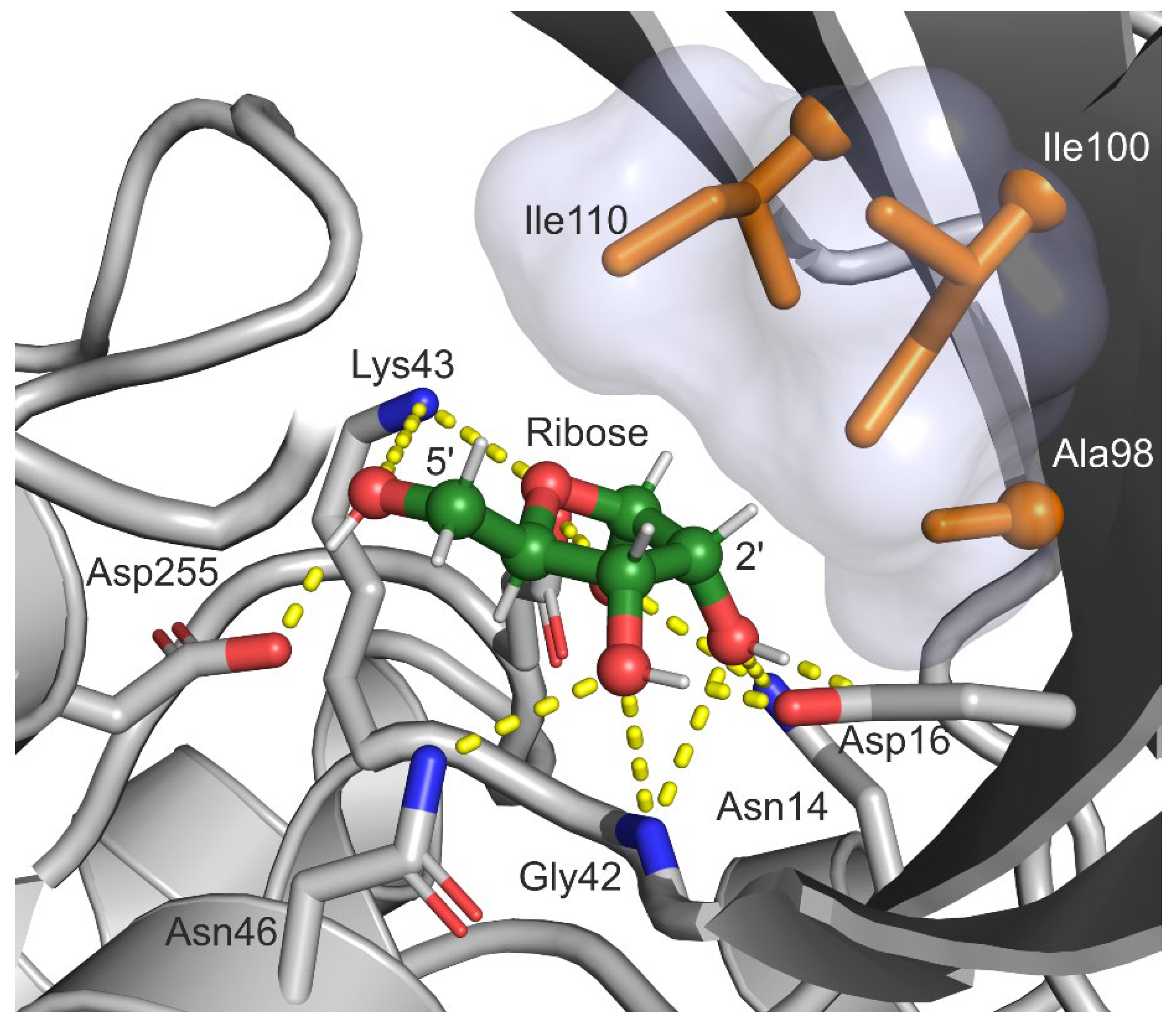
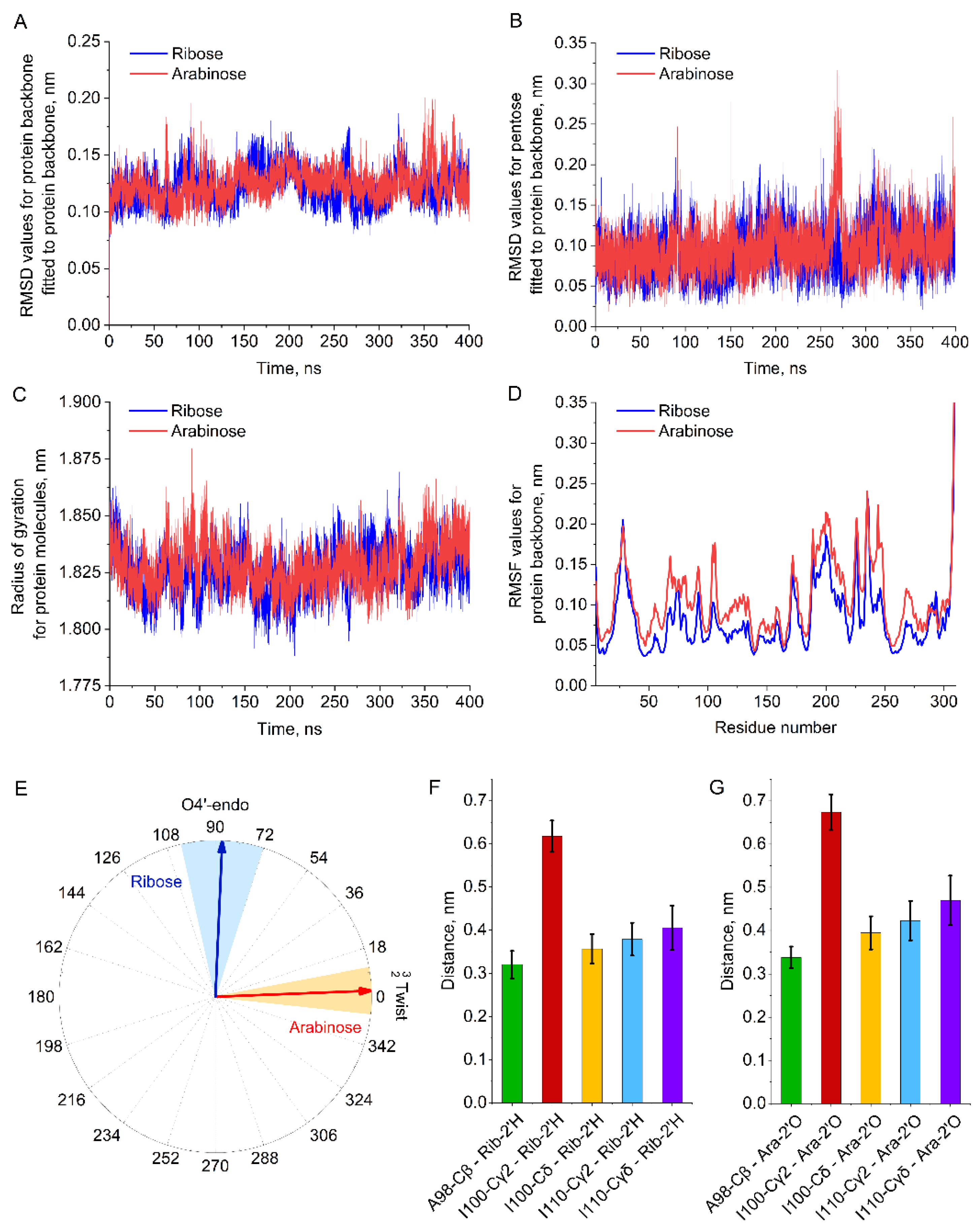
2. Results
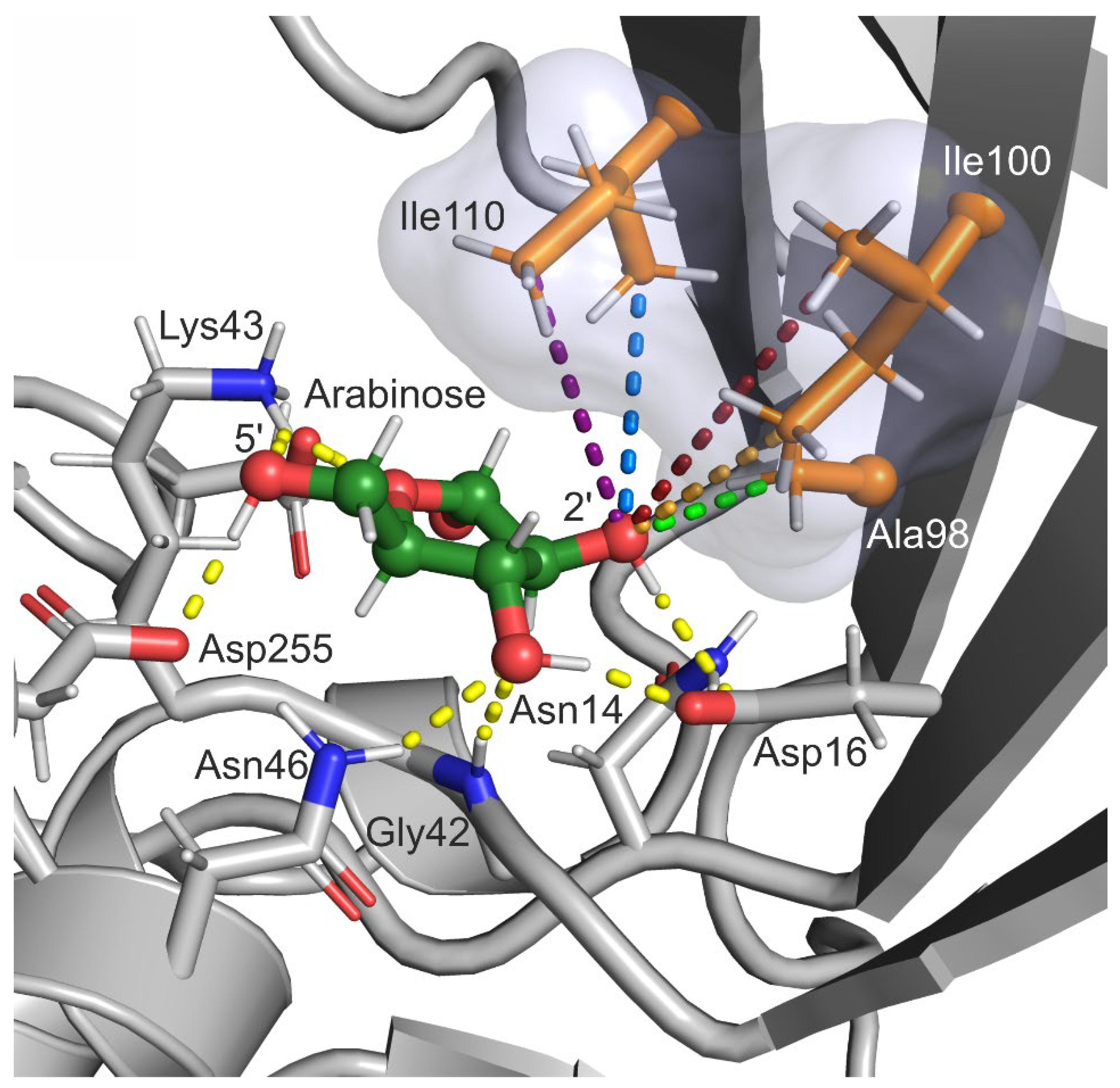
2.1. Selection of Design Hot Spots and Feasible Mutations
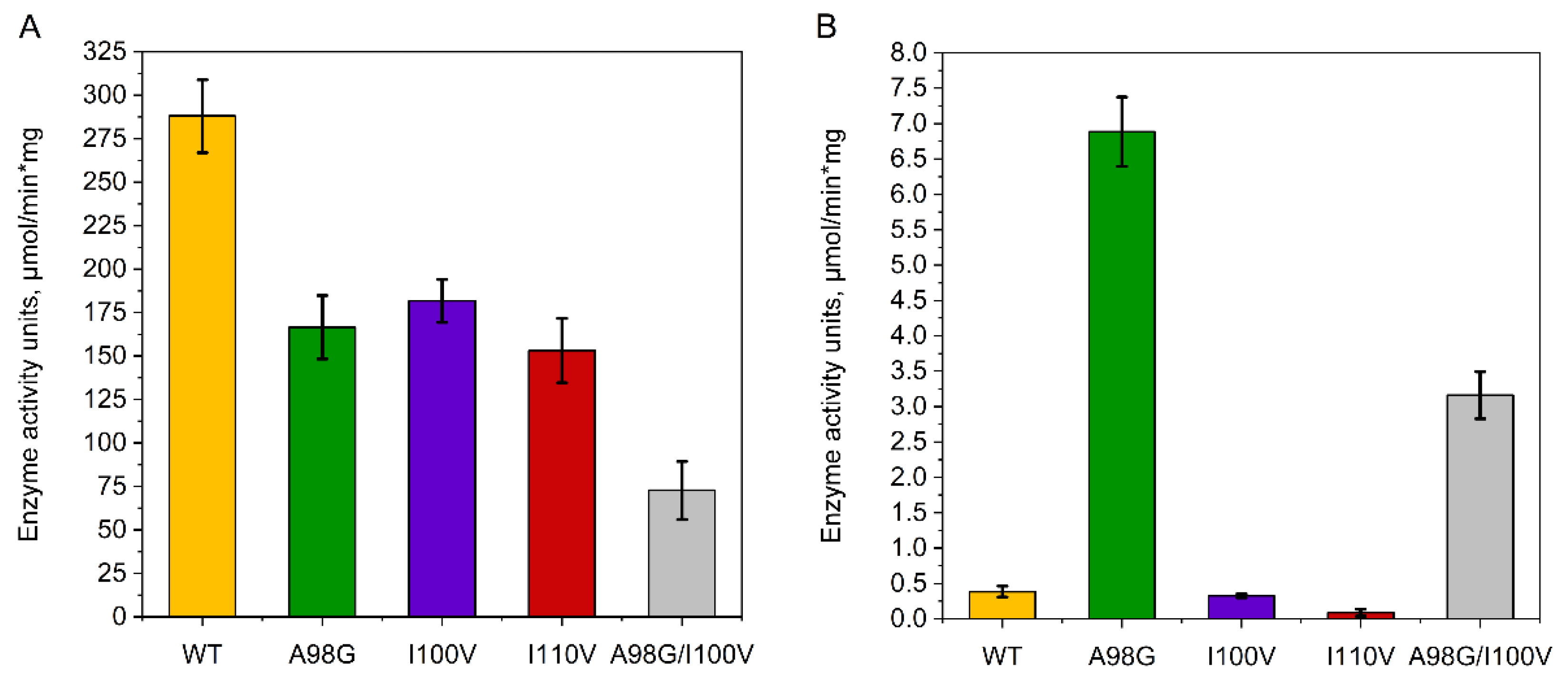
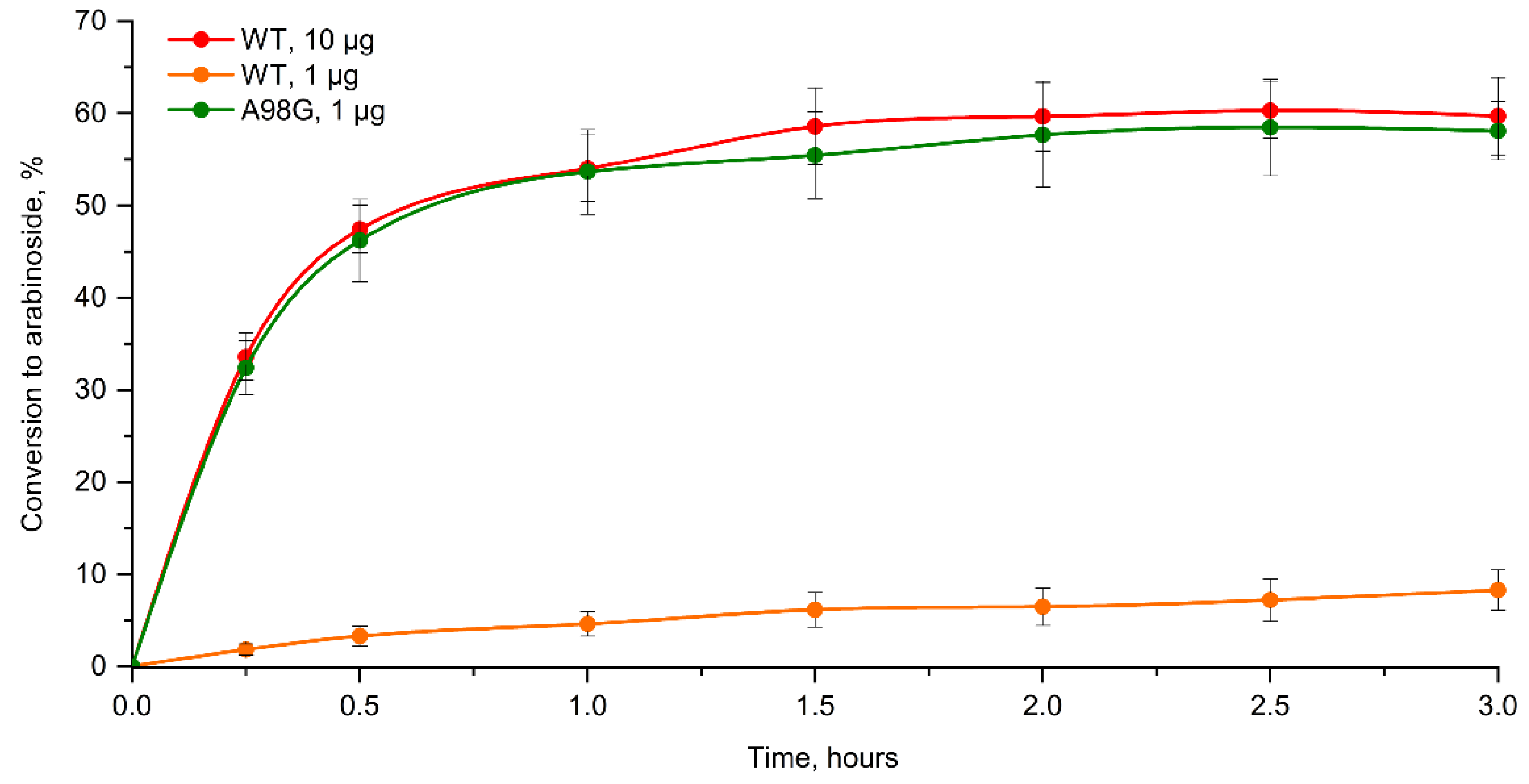
2.2. Enzyme Assay, Kinetic Studies and Cascade Synthesis
3. Discussion
4. Materials and Methods
4.1. Computational Approaches
4.2. Genetic Engineering and Producer Strain Cultivation
4.3. Purification of EcoRK
4.4. Enzyme Assay and Determination of Kinetic Parameters
4.5. Cascade Synthesis of 2-Chloroadenine Arabinoside
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mikhailopulo, I.A.; Miroshnikov, A.I. New trends in nucleoside biotechnology. Acta Nat. 2010, 2, 36–59. [Google Scholar] [CrossRef] [Green Version]
- Jordheim, L.P.; Durantel, D.; Zoulim, F.; Dumontet, C. Advances in the development of nucleoside and nucleotide analogues for cancer and viral diseases. Nat. Rev. Drug. Discov. 2013, 12, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Depaix, A.; Périgaud, C.; Peyrottes, S. Recent Trends in Nucleotide Synthesis. Chem. Rev. 2016, 116, 7854–7897. [Google Scholar] [CrossRef] [PubMed]
- Fateev, I.V.; Kostromina, M.A.; Abramchik, Y.A.; Eletskaya, B.Z.; Mikheeva, O.O.; Lukoshin, D.D.; Zayats, E.A.; Berzina, M.Y.; Dorofeeva, E.V.; Paramonov, A.S.; et al. Multi-Enzymatic Cascades in the Synthesis of Modified Nucleosides: Comparison of the Thermophilic and Mesophilic Pathways. Biomolecules 2021, 11, 586. [Google Scholar] [CrossRef]
- Scism, R.A.; Bachmann, B.O. Five-component cascade synthesis of nucleotide analogues in an engineered self-immobilized enzyme aggregate. ChemBioChem 2010, 11, 67–70. [Google Scholar] [CrossRef]
- Esipov, R.S.; Abramchik, Y.A.; Fateev, I.V.; Konstantinova, I.D.; Kostromina, M.A.; Muravyova, T.I.; Artemova, K.G.; Miroshnikov, A.I. A Cascade of Thermophilic Enzymes As an Approach to the Synthesis of Modified Nucleotides. Acta Nat. 2016, 8, 82–90. [Google Scholar] [CrossRef]
- Yehia, H.; Kamel, S.; Paulick, K.; Wagner, A.; Neubauer, P. Substrate spectra of nucleoside phosphorylases and their potential in the production of pharmaceutically active compounds. Curr. Pharm. Des. 2017, 23, 6913–6935. [Google Scholar] [CrossRef]
- Almendros, M.; Berenguer, J.; Sinisterra, J.V. Thermus thermophilus nucleoside phosphorylases active in the synthesis of nucleoside analogues. Appl. Environ. Microbiol. 2012, 78, 3128–3135. [Google Scholar] [CrossRef] [Green Version]
- Fateev, I.V.; Kharitonova, M.I.; Antonov, K.V.; Konstantinova, I.D.; Stepanenko, V.N.; Esipov, R.S.; Seela, F.; Temburnikar, K.W.; Seley-Radtke, K.L.; Stepchenko, V.A.; et al. Recognition of Artificial Nucleobases by E. coli Purine Nucleoside Phosphorylase versus its Ser90Ala Mutant in the Synthesis of Base-Modified Nucleosides. Chemistry 2015, 21, 13401–13419. [Google Scholar] [CrossRef]
- Miroshnikov, A.I.; Esipov, R.S.; Muravyova, T.I.; Konstantinova, I.D.; Fateev, I.V.; Mikhailopulo, I.A. A New Strategy for the Synthesis of Nucleosides: One-Pot Enzymatic Transformation of D-Pentoses into Nucleosides. Open. Conf. Proc. J. 2010, 1, 98–102. [Google Scholar] [CrossRef]
- Sigrell, J.A.; Cameron, A.D.; Jones, T.A.; Mowbray, S.L. Structure of Escherichia coli ribokinase in complex with ribose and dinucleotide determined to 1.8 A resolution: Insights into a new family of kinase structures. Structure 1998, 6, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Sigrell, J.A.; Cameron, A.D.; Mowbray, S.L. Induced fit on sugar binding activates ribokinase. J. Mol. Biol. 1999, 290, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Timofeev, V.I.; Abramchik, Y.A.; Muravyova, T.I.; Zhukhlistova, N.E.; Esipov, R.S.; Kuranova, I.P. Three-Dimensional Structure of Recombinant Thermophilic Ribokinase from Thermus species 2.9 in Complex with Adenosine Diphosphate. Crystallogr. Rep. 2021, 66, 769–776. [Google Scholar] [CrossRef]
- Paul, R.; Patra, M.D.; Sen, U. Crystal structure of apo and ligand bound vibrio cholerae ribokinase (Vc-RK): Role of monovalent cation induced activation and structural flexibility in sugar phosphorylation. Adv. Exp. Med. Biol. 2015, 842, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Chuvikovsky, D.V.; Esipov, R.S.; Skoblov, Y.S.; Chupova, L.A.; Muravyova, T.I.; Miroshnikov, A.I.; Lapinjoki, S.; Mikhailopulo, I.A. Ribokinase from E. coli: Expression, purification, and substrate specificity. Bioorg. Med. Chem. 2006, 14, 6327–6332. [Google Scholar] [CrossRef]
- Esipov, R.S.; Abramchik, Y.A.; Fateev, I.V.; Muravyova, T.I.; Skoblov, Y.S.; Kostromina, M.A.; Miroshnikov, A.I. Preparation and study of the substrate specificity of thermophilic ribokinase from Thermus sp. 2.9. Russ. J. Biopharm. 2016, 8, 3–12. [Google Scholar]
- Lukenbill, J.; Kalaycio, M. Fludarabine: A review of the clear benefits and potential harms. Leuk. Res. 2013, 37, 986–994. [Google Scholar] [CrossRef]
- Kisor, D.F. Nelarabine: A nucleoside analog with efficacy in T-cell and other leukemias. Ann. Pharmacother. 2005, 39, 1056–1063. [Google Scholar] [CrossRef]
- Murphy, T.; Yee, K. Cytarabine and daunorubicin for the treatment of acute myeloid leukemia. Expert. Opin. Pharmacother. 2017, 18, 1765–1780. [Google Scholar] [CrossRef]
- Whitley, R.; Alford, C.; Hess, F.; Buchanan, R. Vidarabine: A preliminary review of its pharmacological properties and therapeutic use. Drugs 1980, 20, 267–282. [Google Scholar] [CrossRef]
- Park, J.; van Koeverden, P.; Singh, B.; Gupta, R.S. Identification and characterization of human ribokinase and comparison of its properties with E. coli ribokinase and human adenosine kinase. FEBS Lett. 2007, 581, 3211–3216. [Google Scholar] [CrossRef] [PubMed]
- Fateev, I.V.; Antonov, K.V.; Konstantinova, I.D.; Muravyova, T.I.; Seela, F.; Esipov, R.S.; Miroshnikov, A.I.; Mikhailopulo, I.A. The chemoenzymatic synthesis of clofarabine and related 2′-deoxyfluoroarabinosyl nucleosides: The electronic and stereochemical factors determining substrate recognition by E. coli nucleoside phosphorylases. Beilstein. J. Org. Chem. 2014, 10, 1657–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, P.A.; Oh, J.; Lee, H.; Witte, C.P.; Rhee, S. Crystal structure and mutational analyses of ribokinase from Arabidopsis thaliana. J. Struct. Biol. 2019, 206, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Liliemark, J. The clinical pharmacokinetics of cladribine. Clin. Pharmacokinet. 1997, 32, 120–131. [Google Scholar] [CrossRef]
- Pui, C.H.; Jeha, S. Clofarabine. Nat. Rev. Drug. Discov. 2005, 4, 369–370. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- PLUMED consortium. Promoting transparency and reproducibility in enhanced molecular simulations. Nat. Methods 2019, 16, 670–673. [Google Scholar] [CrossRef] [Green Version]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the Amber ff99SB protein force field. Proteins 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, M.; Rahman, A. Strain fluctuations and elastic constants. J. Chem. Phys. 1982, 76, 2662–2666. [Google Scholar] [CrossRef]
- Van Gunsteren, W.F.; Berendsen, H.J.C. A Leap-Frog Algorithm for Stochastic Dynamics. Mol. Simul. 1988, 1, 173–185. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8592. [Google Scholar] [CrossRef] [Green Version]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Esipov, R.S.; Gurevich, A.I.; Chuvikovsky, D.V.; Chupova, L.A.; Muravyova, T.I.; Miroshnikov, A.I. Overexpression of Escherichia coli genes encoding nucleoside phosphorylases in the pET/Bl21(DE3) system yields active recombinant enzymes. Protein Expr. Purif. 2002, 24, 56–60. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vmax, μmol/min·mg | kcat, s−1 | KM, mM | |
|---|---|---|---|
| D-ribose: | |||
| Wild type | 340 ± 50 | 180 ± 30 | 0.15 ± 0.02 |
| A98G | 180 ± 20 | 97 ± 11 | 0.28 ± 0.03 |
| A98G/I100V | 130 ± 30 | 70 ± 16 | 0.24 ± 0.05 |
| D-arabinose: | |||
| Wild type | 3.6 ± 0.5 | 1.9 ± 0.3 | 470 ± 60 |
| A98G | 54 ± 5 | 29 ± 3 | 310 ± 30 |
| A98G/I100V | 33 ± 5 | 17 ± 3 | 730 ± 120 |
| ATP: | |||
| Wild type | 350 ± 40 | 185 ± 21 | 0.43 ± 0.05 |
| A98G | 160 ± 10 | 85 ± 5 | 0.58 ± 0.04 |
| A98G/I100V | 120 ± 10 | 63 ± 5 | 0.75 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zayats, E.A.; Fateev, I.V.; Kostromina, M.A.; Abramchik, Y.A.; Lykoshin, D.D.; Yurovskaya, D.O.; Timofeev, V.I.; Berzina, M.Y.; Eletskaya, B.Z.; Konstantinova, I.D.; et al. Rational Mutagenesis in the Lid Domain of Ribokinase from E. coli Results in an Order of Magnitude Increase in Activity towards D-arabinose. Int. J. Mol. Sci. 2022, 23, 12540. https://doi.org/10.3390/ijms232012540
Zayats EA, Fateev IV, Kostromina MA, Abramchik YA, Lykoshin DD, Yurovskaya DO, Timofeev VI, Berzina MY, Eletskaya BZ, Konstantinova ID, et al. Rational Mutagenesis in the Lid Domain of Ribokinase from E. coli Results in an Order of Magnitude Increase in Activity towards D-arabinose. International Journal of Molecular Sciences. 2022; 23(20):12540. https://doi.org/10.3390/ijms232012540
Chicago/Turabian StyleZayats, Evgeniy A., Ilya V. Fateev, Maria A. Kostromina, Yulia A. Abramchik, Dmitry D. Lykoshin, Daria O. Yurovskaya, Vladimir I. Timofeev, Maria Ya. Berzina, Barbara Z. Eletskaya, Irina D. Konstantinova, and et al. 2022. "Rational Mutagenesis in the Lid Domain of Ribokinase from E. coli Results in an Order of Magnitude Increase in Activity towards D-arabinose" International Journal of Molecular Sciences 23, no. 20: 12540. https://doi.org/10.3390/ijms232012540